
Abstract
Neuroprosthetics is a multidisciplinary field at the interface between neurosciences and biomedical engineering, which aims at replacing or modulating parts of the nervous system that get disrupted in neurological disorders or after injury. Although neuroprostheses have steadily evolved over the past 60 years in the field of sensory and motor disorders, their application to higher-order cognitive functions is still at a relatively preliminary stage. Nevertheless, a recent series of proof-of-concept studies suggest that electrical neuromodulation strategies might also be useful in alleviating some cognitive and memory deficits, in particular in the context of dementia. Here, we review the evolution of neuroprosthetics from sensorimotor to cognitive disorders, highlighting important common principles such as the need for neuroprosthetic systems that enable multisite bidirectional interactions with the nervous system.
Similar content being viewed by others

Introduction
Neural oscillations are ubiquitous throughout the nervous system, subserving sensory, motor, and cognitive functions in the brain and the spinal cord1. These oscillations are often disrupted in neurodegenerative and neuropsychiatric disorders2, after stroke3, traumatic brain injury4, or spinal cord injury (SCI)5. Despite different underlying mechanisms, abnormal or even abolished neural oscillations seem to be a hallmark of neurological disorders and to play a causative role in their behavioral symptoms6. In recent years, bioelectronic devices that interface with the nervous system and affect or reestablish these oscillations have become an alternative to pharmacological treatments, opening a new era of medicine consisting of electroceuticals, Brain-Computer or Brain-Machine Interfaces (BCI/BMI), and neuroprostheses7,8,9,10.
These bioelectronic devices can now treat some motor symptoms of Parkinson’s disease (PD)11 or restore motor function after SCI12,13,14. However, cognitive disorders still elude this new therapeutic modality, due to the intrinsic complexity of the neural mechanisms underlying cognition and to technological challenges for modulating the associated networks. Whether they result from brain injury or neurodegenerative diseases such as Alzheimer’s disease (AD), cognitive disorders do not yet have efficient pharmacological treatments, represent a major public health issue, and could potentially benefit from neuromodulation strategies15.
Here, we review the current state of the art on neuromodulation and neuroprosthetic approaches for sensorimotor and cognitive disorders, with a focus on invasive neurotechnologies tested in non-human primates (NHP) and humans. We first introduce the concept of motor neuroprostheses for motor impairments resulting from stroke or SCI, and somatosensory neuroprostheses for providing sensory feedback after limb amputation or paralysis. Next, we present deep brain stimulation (DBS) for motor and cognitive symptoms of PD. We then discuss the neural oscillations that characterize cognitive functions, especially learning and memory, and their alterations in memory disorders. This is followed by a state of the art on neuromodulation approaches for AD and other memory impairments. Finally, we present the enticing hypothesis that cognitive deficits would particularly benefit from the development of a new generation of neuroprosthetic systems that target large-scale brain network oscillations and facilitate the associated neurological functions.
Motor neuroprostheses
Motor neuroprostheses refer to a particular type of neuroprostheses that aim at restoring motor function by electrical stimulation of the structures involved in the generation of movement (muscles, peripheral nerves, spinal cord, or brain), after neuromotor disorders such as stroke or SCI16. The very first motor neuroprosthesis was a peroneal nerve stimulator invented in 1961 by Liberson and colleagues to treat foot drop after hemiplegia17. The term neuroprosthesis itself was first coined in the scientific literature in 1971 to refer to an intraspinal implant that allowed bladder voiding after paraplegia18. Since then, the definition of motor neuroprostheses has also been extended to technologies that extract motor commands from brain signals to control external devices, also called BCI or BMI (e.g. refs. 19,20). In this section, we briefly review the different types of motor neuroprostheses, ranging from neurostimulation of muscles, peripheral nerves, and spinal cord, to implantable BMIs.
Functional electrical stimulation (FES)
FES is a clinically approved neurostimulation technology that activates the efferent axons innervating specific muscles to produce a desired movement21. The stimulation can be delivered in the vicinity of the targeted muscle or to a motor nerve innervating it, in which case it is called peripheral nerve stimulation (PNS), using either non-invasive, percutaneous, or fully implanted electrodes. These electrodes are in turn connected to an electrical stimulator, which can typically control up to 16 independent channels. Such stimulation systems may simply be used to build up muscle strength, often referred to as NeuroMuscular Electrical Stimulation, or they can assist in functional tasks. Moreover, FES can serve as an assistive technology, reducing impairment in the execution of a given movement, or as part of a rehabilitation therapy that may lead to neuroplasticity and functional improvements, depending on the underlying disorder and its severity21.
FES has been applied over the past 60 years to both upper- and lower-limb motor tasks, such as standing, walking, reaching, and grasping21,22. Several FDA-approved or CE-marked systems are now commercially available for these applications. Clinically, FES has been mostly used for hemiplegia resulting from stroke, for example, as a foot-drop stimulator17, and for the rehabilitation of upper-limb motor function23. In the context of SCI, FES has also been extensively tested in research studies24,25 but has not yet become standard clinical practice. Overall, FES systems represent the main class of neuroprostheses that have been clinically accepted, but they remain mostly restricted to post-stroke motor rehabilitation. However, FES suffers from a conceptual limitation as it recruits motor axons in a non-physiological order. The large-diameter motor axons that produce large forces but are not resistant to fatigue are indeed recruited first26,27,28, which makes it challenging for FES to sustain large forces during extended periods of time. To circumvent this limitation, methods that target sensory afferents and activate reflex circuits within the spinal cord are desirable.
Spinal cord stimulation (SCS)
Well established as a treatment for chronic pain, SCS has recently received significant attention for its applications in motor control29. Epidural SCS can be delivered via percutaneous or fully implanted leads containing up to 16 electrodes placed in the posterior epidural space, which are then connected to an external or an implantable pulse generator (IPG). The mechanisms underlying its motor effects involve the activation of large-diameter afferent fibers located in the posterior roots, which in turn recruit motoneuron pools at the innervated spinal segment30,31.
The first observation that SCS may be used as a motor neuroprosthesis after SCI was made by Barolat and colleagues in 198632. They showed that one subject with incomplete SCI (i.e. with residual sensory or motor fibers crossing the injury site) regained voluntary motor control with SCS after several months of stimulation. A decade later, Dimitrijevic and colleagues demonstrated that SCS delivered over the lumbar spinal cord could produce rhythmic electromyographic responses and flexion-extension leg movements similar to stepping33. These results were obtained in six subjects with complete SCI (i.e. with no residual fiber across the injury) and provided indirect evidence for the existence of a Central Pattern Generator (CPG) in humans34. Later studies showed that varying stimulation frequency could also produce a bilateral extension of the lower limbs35,36.
The first combination of SCS with locomotor training was performed in 2002 in a subject with incomplete SCI placed in a partial weight-bearing system37,38,39. SCS led to an immediate facilitation of walking, which further improved during training but was not maintained without SCS. The combination of SCS and neurorehabilitation was then revived in 2011 by Harkema and colleagues, who showed that SCS allowed independent, full weight-bearing standing after 80 sessions of intensive training in one subject with complete SCI40. Importantly, this study re-discovered that SCS can enable voluntary movements of paralyzed muscles, as first observed by Barolat in 1986. Similar results were then confirmed in three more participants41,42 and later replicated at the Mayo Clinic43. One subject continued to train with SCS for 3.7 years, which led to a partial recovery of voluntary leg movements without SCS, indicating that SCS has the potential to trigger neuroplasticity mechanisms44.
A milestone for the application of SCS in people with SCI was reached in 2018. For the first time, three independent groups demonstrated in a total of six subjects that SCS combined with intensive rehabilitation, could enable independent overground walking13,45,46. At the Kentucky SCI Center, four participants with motor-complete SCI performed training sessions for standing, body-weight-supported treadmill stepping and overground walking, all with continuous SCS45. When using SCS, all participants achieved assisted standing and improved trunk stability while sitting. Moreover, the two participants with motor-complete, sensory-incomplete SCI gained the ability to walk overground with continuous SCS and assistive devices after a period of 15 and 85 weeks. Similarly, at the Mayo Clinic, an individual with chronic motor- and sensory-complete SCI was trained to perform step-like movements with SCS in a side-lying position43 and then on a treadmill and overground46. After 43 weeks of training and with continuous SCS, this participant was able to stand, step on a treadmill and walk overground with a walker and manual assistance for hip stability.
In parallel, Courtine and Bloch’s team in Lausanne pioneered a new paradigm termed spatiotemporal SCS13. Based on prior preclinical work47,48, they leveraged an IPG with real-time control capabilities to develop a stimulation protocol that alternated between the swing, weight acceptance, and propulsion phases of the gait cycle. These functionalities were targeted by spatially-specific electrode configurations, which were triggered either at a pre-defined pace or in real-time by residual kinematic events. Spatiotemporal SCS led to an immediate facilitation of walking and a long-term recovery of motor function within 6 months of training in three subjects with chronic incomplete SCI13. This approach was recently expanded to a total of nine participants49, including three patients with motor-complete SCI implanted with a new electrode array tailored for both leg and trunk motor functions14. Overall, these results have created a surge of interest to now integrate SCS into rehabilitation protocols after SCI.
Brain-controlled motor neuroprostheses
Both FES and SCS are neuroprostheses that can be controlled manually or by external kinematic events through external sensors that detect specific state changes. These state changes can also be extracted directly from brain signals through the use of BCI/BMI technologies. Such technologies record brain activity either non-invasively by electroencephalography (not discussed here), or invasively by electrocorticographic (ECoG) grids placed above or below the dura mater, or directly inside the brain by intracranial electrodes50.
Intracortical BMIs take their roots in early studies by Georgopoulos and colleagues in 1982, who showed that neurons in the motor cortex of NHPs performing a reaching task are tuned to the direction of movement51. With the advent of high-density microwire or microelectrode arrays (MEA), such as the Utah array, neurophysiologists then became able to record tens or hundreds of cells simultaneously from motor cortices. In the early 2000s, three groups led by Nicolelis, Donoghue and Schwarz, demonstrated in NHPs that these intracortical MEAs, combined with signal processing and machine learning, allowed to decode in real-time the intended hand trajectory and to control artificial devices such as a computer cursor or robotic arms20,52,53,54.
A few years later, Hochberg and colleagues translated these findings into a tetraplegic individual implanted with a 96-channel MEA in the hand area of the primary motor cortex55. Neural signals still showed movement-related modulation of spiking activity even after several years of paralysis. Moreover, algorithms to decode 2D movement intentions and discrete neural states enabled the participant to open and close a prosthetic hand, control a computer cursor and operate various software. The same group later demonstrated that two additional subjects could use this technology to control a robotic arm with three degrees of freedom (DOF) for the end-point trajectory and discrete states for hand grasping56. In parallel, Collinger and colleagues trained a participant implanted with two 96-channel MEAs in the motor cortex to control a robotic arm with up to seven DOF57. To control more complex movements that require bilateral coordination, intracortical BMIs can also be combined with additional control strategies, as demonstrated by Tenore’s work on the control of two prosthetic limbs (up to 12 DOF) for bimanual self-feeding58. In a recent application, Shenoy’s group expanded intracortical BMIs to the decoding of handwriting (31 characters), which allowed one subject to type sentences at speeds comparable with typical smartphone typing speeds59.
Less invasive than intracortical MEAs, ECoG implants consist of grids or strips of electrodes placed over the cortical surface or the dura mater, and are routinely used in neuromonitoring for epilepsy. In 2016, Ramsey’s group implanted subdural ECoG strips over the motor cortex of a person with amyotrophic lateral sclerosis, and connected them to an IPG with real-time recording capabilities60. This fully implanted BMI allowed the participant to control a typing program after 28 weeks of training. In 2019, Benabid and colleagues tested a new fully implantable ECoG recording system with 64 epidural electrodes placed bilaterally over the motor cortices in a subject with tetraplegia61. The participant trained to control a virtual avatar and a four-limb exoskeleton, achieving up to eight DOF without any recalibration over seven weeks. Recently, Chang’s group also applied subdural ECoGs to speech neuroprostheses, decoding both full words based on a 50-word vocabulary62, and individual letters to produce sentences from a >1000-word vocabulary63. Future applications will likely require the development of novel micro-ECoG implants characterized by smaller electrodes, increased electrode densities, and flexible substrates able to conform to the cortical surface64.
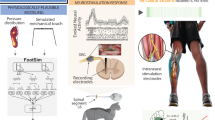
Lastly, BMIs should not be seen as a separate type of neuroprosthesis from neurostimulation methods, but rather as a potential source of control signals for operating them. Indeed, two groups showed in 2016–2017 that tetraplegic subjects with intracortical MEAs could control FES systems to achieve respectively finger, hand, and wrist movements65, or reaching and grasping movements involving the elbow, wrist, and hand66. Similarly, spatiotemporal SCS can be controlled directly by brain signals, as demonstrated in 2018 using intracortical MEAs in NHPs with a unilateral spinal cord injury12, and envisioned in humans using ECoG implants67. These various studies pave the way towards neuroprostheses that bidirectionally interface with the nervous system and harness both recording and stimulation capabilities (Fig. 1).

a Cortical implants typically used for sensorimotor neuroprostheses can be divided into two categories: intracortical MEAs such as the Utah array (10 × 10 Utah array, picture extracted with permission from ref. 55), and epidural or subdural ECoG strips or grids with different specifications (illustrated for a 8 × 8 ECoG grid, 4 mm contact diameter, 10 mm pitch, with permission from CorTec GmbH). These implants can both record neurophysiological signals and deliver electrical stimulation. b Peripheral and spinal implants for sensorimotor neuroprostheses target either the motor nerves or muscles in the case of FES (adapted with permission from ref. 66), the spinal cord in epidural SCS (adapted with permission from ref. 13), or the sensory nerves in PNS for somatosensory feedback (adapted with permission from ref. 80). All applications shown here used these implants for delivering electrical stimulation. Lightning bolts indicate neurostimulation.
Somatosensory neuroprostheses
Electrical neurostimulation can be used to induce movement as in motor neuroprostheses, but also to elicit somatic sensations such as touch or proprioception in individuals with limb amputation or paralysis68. In this section, we present somatosensory neuroprostheses that target either the peripheral sensory nerves of the somatosensory cortex, and the additional cognitive benefits associated with these bidirectional systems.
Peripheral somatosensory neuroprostheses
Peripheral somatosensory neuroprostheses, which are based on sensory PNS, have been developed to provide sensory feedback to amputees wearing a mechanical prosthesis of the missing limb. Early proof-of-concept studies by Clippinger and colleagues in the 70’s delivered stimulation through a single-channel non-penetrating cuff electrode wrapped around the median or sciatic nerve for, respectively upper- and lower-limb amputees69,70. Similar cuff electrodes have been used more recently in conjunction with an osseointegrated anchor to provide robust and chronic bidirectional communication with a prosthesis in upper-limb amputees over three to 7 years71,72. To generate more natural touch sensations, the traditional cuff electrode design has also been improved and scaled up to 16 stimulation contacts, which provided long-term natural tactile sensations in upper- and lower-limb amputees implanted for more than a year73,74.
The specificity of PNS depends on the ability to target specific bundles of nerve fibers, called fascicles, within a nerve. Higher specificity can therefore be achieved by electrodes that penetrate inside the nerve and reach individual fascicles. This led to the development of longitudinal intrafascicular electrodes, which were used to provide discrete and graded tactile sensations in upper-limb amputees75,76,77. Furthermore, fascicles innervating different body parts are somatotopically organized along the transverse section of a nerve, which motivated the design of transverse intrafascicular electrodes that offer higher spatial selectivity of multiple fascicles and have been applied to both the upper78,79 and lower extremities80,81. Finally, a slanted version of the Utah 96-MEA has also been used for transverse PNS and could evoke up to about a hundred different sensations in upper-limb amputees82,83.
Cortical somatosensory neuroprostheses
Somatosensory feedback can be delivered to the peripheral nerves, but also directly to the brain structures underlying somatosensation. The scientific foundations for cortical somatosensory neuroprostheses were first laid by Penfield and Boldrey back in 1937 during their mapping of sensory and motor cortices in epileptic patients84. These concepts were then revived in 1998 by Romo and colleagues, who demonstrated in NHPs that the sensation of mechanical flutter can be artificially induced by electrically stimulating quickly-adapting neurons in the somatosensory cortex at the frequency of the mimicked stimulus85,86. This observation led several other groups to show that intracortical microstimulation (ICMS) can be used as a cue to induce natural or learned behavioral responses87,88,89,90. Ultimately, ICMS was combined with intracortical BMIs to generate bidirectional neuroprostheses that extract motor commands from motor or association cortices and deliver feedback to the somatosensory cortex, as first demonstrated in NHPs in 201191,92.
These preclinical ICMS studies paved the way for the clinical translation of cortical somatosensory neuroprostheses. In 2016 and 2018, the groups of Gaunt and Andersen implanted paralyzed individuals with chronic intracortical MEAs in the somatosensory cortex93,94,95,96,97. ICMS was shown to evoke somatotopically organized tactile sensations93, which included both cutaneous and proprioceptive modalities94. These sensations depended on stimulation frequency95, were stable over months and even years96, and improved the quality of BMI control in motor tasks97. In 2022, Tenore’s group refined the intraoperative mapping procedure to target specific fingertips with this technology, which allowed a participant to distinguish up to seven finger-specific locations98. In parallel with these chronic implantations, acute experiments in epileptic patients have demonstrated that ECoG can also be used to deliver sensory feedback by stimulating the cortical surface, but provide lower dermatome specificity than intracortical MEAs99,100,101,102.
Cognitive benefits of bidirectional sensorimotor neuroprostheses
Finally, we would like to highlight that somatosensory feedback provides not only sensorimotor but also cognitive benefits to their users as shown by the groups of Raspopovic and others81,103,104,105,106,107. These benefits include an improved embodiment of the prosthesis and reduced abnormal phantom limb perceptions in amputees81,103,105,107, better multisensory integration104, more physiological kinematics and sensorimotor strategies106, decreased weight perception of the prosthesis107 and better cognitive integration in dual tasks81,107. These exciting outcomes provide a strong rationale for the clinical adoption of bidirectional and biomimetic neuroprostheses that improve sensorimotor and cognitive functions after injury (Fig. 1).
Neuromodulation for PD: from motor to cognitive symptoms
In parallel with the development of sensorimotor neuroprostheses, the electrical stimulation of deep brain structures, called DBS, was first introduced for Parkinson’s tremor by Benabid and colleagues in 1987108 and has since then become the main neuromodulation therapy for several motor disorders. PD is characterized by the progressive loss of dopaminergic neurons in the substantia nigra pars compacta (SNc), which results in a range of motor and non-motor symptoms. Motor symptoms include mainly tremor, limb stiffness (rigidity), slowness of movement (bradykinesia), impaired posture and balance. Non-motor symptoms include autonomic dysfunction, mood disorders, and cognitive impairment, which can range from Mild Cognitive Impairment (MCI) to Parkinson’s disease dementia (PDD) as the disease progresses109,110.
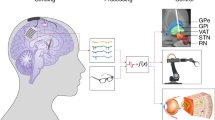
PD affects the whole basal ganglia (BG) circuitry, a set of subcortical areas that comprise the subthalamic nucleus (STN), the external and internal parts of the globus pallidus (GPe, GPi), the striatum, and the SNc. These nuclei interact with cortical areas via the thalamus, thus giving rise to several parallel BG-thalamo-cortical loops (Fig. 2). The loss of dopaminergic cells in the SNc reduces dopamine levels, leading to an abnormal BG-thalamo-cortical network activity, which in turn impacts motor cortical areas involved in the generation of movement, and prefrontal areas involved in cognitive functions. Given that dopamine is also a key element in reward-based learning, these anatomical substrates explain how the symptoms of PD can affect both the motor and cognitive domains111,112. Here, we review the different neuromodulation targets and strategies that have been investigated to treat both motor and cognitive symptoms of PD.

a Anatomical locations of the BG, cortical and thalamic areas involved in the pathophysiology of PD. DBS targeting either the STN or the GPi (lightning bolts) is one of the main treatments for motor symptoms of PD. b The loss of dopaminergic cells within the SNc in PD induces dysfunction of the direct and hyperdirect pathways of the cortico-BG-thalamocortical circuit, which affects both motor and cognitive functions, and can be partially restored using DBS of the STN or GPi.
Conventional DBS of STN or GPi for motor symptoms
The two main strategies for treating motor symptoms associated with PD are dopaminergic medication (levodopa) and DBS of the STN113,114 or GPi115, which leads in both cases to an improvement in motor symptoms and a reduction in the required levodopa medication116. Conventional DBS protocols, which employ frequencies in the high-gamma range (100–185 Hz), improve bradykinesia, rigidity and tremors, but are poorly effective for axial motor symptoms such as freezing of gait and swallowing. Instead, these symptoms may be better controlled by low-frequency DBS, delivered at frequencies in the lower gamma range (60–80 Hz)117,118.
Unfortunately, conventional DBS for PD brings little to no improvement, or sometimes even a worsening, of cognitive functions119,120,121. In particular, a meta-analysis of 41 articles gathering a total of 1622 patients reported that DBS of the STN worsens psychomotor speed, memory, semantic fluency, phenomic fluency and general cognitive functions119. In another study, patients who had long-term follow-up (3.6–10.5 years post-surgery) showed an overall cognitive decline and conversion from MCI to dementia, indicating that conventional DBS of the STN did not stop the cognitive decline observed in PD120. Although the literature on the non-motor effects of DBS of the GPi is more scarce, the cognitive outcomes seem to be slightly better when targeting GPi rather than STN116,122. Importantly, a recent study in 91 PD patients with bilateral DBS of the STN has shown that the precise location within the STN itself could also have an impact on the non-motor effects, including mood/apathy, attention/memory, and sleep/fatigue, which would explain the large variability observed across patients122.
The nucleus basalis of Meynert (NBM): a potential target for cognitive impairment
In addition to the loss of dopaminergic cells in the SNc, PD patients with cognitive impairment or dementia present a loss of cholinergic output from a basal forebrain nucleus called the NBM123. This observation has motivated several studies based on DBS of the NBM for improving cognitive function in PDD and other conditions, such as AD and dementia with Lewy bodies (DLB)124,125,126,127,128,129,130,131,132.
In an encouraging case study from 2009, a patient with PDD was implanted with four leads, placed bilaterally in the STN and NBM to treat both motor and cognitive symptoms125. This patient showed an improvement in cognitive functions (attention, concentration, alertness, drive and spontaneity) and changes in apraxia, when DBS was delivered simultaneously to the NBM (at 20 Hz) and STN leads, but not when stimulation was applied only to the STN125,126. This case study triggered a series of small-scale (six patients each) randomized, double-blind sham-controlled trials of NBM DBS at 20 Hz in patients with AD127,133, PD128, or DLB130,134. Although these studies validated safety and feasibility, they were rather inconclusive in terms of efficacy on cognition. Interestingly, the main outcome was an improvement in neuropsychiatric symptoms in some patients with PD or DLB, especially a reduction in visual hallucinations128,130.
Recently, simultaneous DBS of the NBM together with either the STN or GPi has been revisited, to treat both motor and cognitive symptoms of PD as in the original study from 2009. A case report published in 2019 showed the feasibility to target both the GPi and NBM unilaterally with a single lead, which had its most distal contact in the NBM and its second-most distal contact in the GPi129. Furthermore, current IPGs allow to stimulate simultaneously with different frequencies on different contacts, which was used to deliver pulses at 20 Hz and 130 Hz, respectively, in the NBM and GPi. This study showed a partial improvement in cognitive functions 3 months after the combined stimulation, which warrants further investigation.
Theta DBS for cognitive impairment in PD
Motor and cognitive symptoms of PD may be treated by targeting specific anatomical structures with DBS, but also by applying different stimulation frequencies to a given structure, which may in turn recruit different network dynamics. In particular, there is ample evidence for the role of theta oscillations (4–12 Hz) in cognitive processes. This observation has led several groups to compare the cognitive effects of STN DBS in the theta range (at either 5 or 10 Hz) with more standard protocols in the high-gamma range (130 Hz)135,136,137,138.
A first study in 2006 reported that 10-Hz stimulation of the STN significantly improved verbal fluency compared to standard 130-Hz stimulation in 12 patients with PD135. There was also a non-significant trend towards improvement when compared to the absence of stimulation. A later study in 15 subjects showed that 5-Hz DBS of the STN significantly improved performance on verbal processing speed and response inhibition through color-word interference, when compared to no stimulation and 130-Hz DBS136. Recently, two additional studies revisited these concepts. One clinical trial in 12 patients systematically assessed verbal fluency for episodic and non-episodic categories, color-word interference, and random number generation, during DBS of the STN at 130 Hz, at 10 Hz, or in the absence of stimulation137. The results indicated a significant improvement specifically for the episodic verbal fluency task when theta DBS was delivered, an interesting finding given the prominence of this rhythm in episodic memory. Finally, the latest study investigated the impact of stimulation location (ventral vs dorsal part of the STN) and frequency (personalized theta vs 130 Hz) on verbal fluency in nine patients138. Theta DBS was delivered at frequencies between 4 and 8 Hz depending on the subject (mean 5.7 Hz). Importantly, it was found that theta DBS of the dorsal part of the STN yielded significantly better cognitive outcomes than no stimulation or gamma stimulation of either the dorsal or ventral part of the STN.
Neural oscillations underlying cognition in health and memory disorders
In the previous section, we reviewed how neuromodulation strategies originally designed for motor symptoms could be adapted to cognitive symptoms in the context of PD, by either targeting different anatomical targets or tapping into different networks based on the stimulation frequency. Hereafter, we focus on the role of neuronal oscillations in cognitive processes in general, and memory in particular, with the intent to develop neuromodulation strategies for neuronal disorders that are primarily cognitive by nature, such as memory impairments and AD.
Neural oscillations in normal cognitive functions
Cognitive functions are associated with neural oscillations in specific frequency ranges affecting higher order neocortical areas, called association cortices, and other limbic or subcortical structures such as the hippocampus, especially in the theta (4–12 Hz) and gamma (~40 Hz) ranges139. These oscillations usually affect simultaneously multiple brain structures, which belong to a common large-scale brain network subserving a given cognitive function140,141. It is thought that long-range phase synchronization in the low frequencies (especially theta) allows remote areas to communicate with each other142,143,144. Moreover, coupling between oscillations in two different frequency bands, known as cross-frequency coupling (CFC), allows these long-range interactions to influence local gamma oscillations and information processing. The most widely studied example is the coupling between the phase of theta oscillations and the amplitude of gamma oscillations in limbic circuits145,146,147,148,149,150,151,152. Together, long-range phase synchronization and both local and long-range CFC allow complex interactions between remote areas belonging to the same large-scale brain network153. Understanding these interactions, how they are affected in cognitive disorders, and how to restore them using neuromodulation, is therefore critical for the development of future therapies15,154.
Among all cognitive processes, learning and memory have received considerable research attention because of their pivotal role in our daily lives. We will focus here on the circuit underlying episodic memory (Fig. 3), which is a form of long-term memory underlying the learning and recollection of past events. Episodic memory involves bidirectional interactions between the hippocampus and other cortical areas, including the prefrontal cortex (PFC)155,156. These interactions are partly mediated by the entorhinal cortex (EC), which acts as a hub between the hippocampus and these cortical areas. The dorsolateral part of the PFC (dlPFC) is thought to be mostly associated with working memory, or the maintenance of information (e.g., from sensory cortices) for a short time period before being used by other cognitive processes. The medial PFC (mPFC) is involved in memory consolidation and provides contextual information during episodic memory retrieval. In addition to the connections mediated by the EC, there is a direct projection from the ventral/anterior hippocampus to mPFC, which may be associated with this role in processing contextual information.

a Limbic and cortical structures involved in cognition, in particular episodic memory. These structures are part of a common large-scale brain network. b Schematic representation of the connections between these structures, including intra-hippocampal circuitry (red dashed box), the original circuit of Papez (open arrowheads), and additional connections between them (filled arrowheads). The NBM provides widespread cholinergic innervation to neocortical areas and has therefore received attention as a neuromodulation target for cognition. The fornix has been the main chronic DBS target in AD patients. Other areas (hippocampus, EC, neocortical areas) have been mainly targeted in acute studies in epileptic patients.
In summary, there is strong evidence that functional interactions between the hippocampus and PFC involve theta oscillations in both structures, theta phase synchronization, and theta-gamma CFC142,143,144,145,146,147,148,149,150,151,152. Recent evidence in NHPs shows that other oscillations such as alpha and beta in the hippocampus and PFC are also involved in associative learning157. Overall, these oscillations seem to be not just an epiphenomenon, but to play a causative role in efficient memory encoding and retrieval6.
Alteration of network connectivity and neural oscillations in AD and memory disorders
Memory disorders occur when a part of this episodic memory circuit gets disrupted. This finding has been supported by a large body of evidence showing structural and functional disconnections between nodes of the large-scale episodic memory network in neurodegenerative diseases158,159, post-traumatic amnesia160 and other forms of memory disorders161. For example, the EC is one of the first areas to be affected in the early stages of AD, even before the first symptoms are diagnosed, and plays a key role in the communication between neocortical areas and the hippocampus162. In post-traumatic amnesia, mechanical damage to the long-range white matter tracts connecting these structures also mediates the associated deficits160.
This structural and functional disconnection results in an alteration of neural oscillations and impacts cognitive processes. Such alterations in theta and gamma rhythms and their CFC have been observed in AD and temporal lobe epilepsy161. A long history of EEG studies in patients with AD has shown an increase in low-frequency delta and theta rhythms, and a decrease in faster alpha and beta rhythms163. Some studies have also reported either a decrease164 or an increase165,166 in gamma oscillations, a change in the modulation of theta during a working memory task in subjects with AD or MCI167, and either a reduced165 or a larger168,169 theta-gamma CFC. Overall, the observed changes are not always consistent from one clinical study to another and may depend on the recording condition (resting-state, steady-state evoked potentials, or during a cognitive task), on interindividual variability, and on potential compensatory mechanisms that cloud the interpretation of these reports.
Despite the variability and relative scarcity of human studies on this topic, the changes in theta and gamma rhythms, as well as their CFC, have also been thoroughly investigated in animal models of AD. The results in individual theta and gamma rhythms were not quite conclusive in these models or did not mirror the observations in patients161, but a reduced theta-gamma CFC was consistently observed in several transgenic mouse models of AD159,170,171,172. Only one study has found an opposite trend in a rat model of hyperglycemia, which is a risk factor for AD173. Although more work is needed in different animal models and at different timepoints during the evolution of each phenotype, theta-gamma CFC appears as the most reproducible oscillatory alteration both in animal models and in humans with AD. This observation is quite compelling given the role of CFC in non-pathophysiological learning and memory processes.
Neuromodulation for AD and memory disorders
The observation that neural oscillations play a key role in cognitive processes and are disrupted in a range of memory disorders, including AD, leads us to the enticing hypothesis that restoring these oscillations and their couplings might rescue the associated symptoms15,154. In turn, chronic restoration of these oscillations might trigger neuroplasticity mechanisms, slow down the progression of neurodegenerative processes, or even lead to long-lasting improvements174. Below, we review the main intracranial neuromodulation studies that have aimed at improving cognition and memory in patients with AD chronically implanted with DBS electrodes, or in epileptic patients who underwent acute intracranial monitoring prior to resective surgery (Table 1).
DBS of the NBM in AD
Historically, the NBM was the first DBS target to be investigated as a potential symptomatic treatment of AD. As discussed previously, this structure has been targeted in PDD, AD and DLB to increase the release of acetylcholine, which led to improved neuropsychiatric outcomes in some patients131. In the context of AD, an original case study from 1985 showed increased glucose metabolic activity in the ipsilateral temporal and parietal lobes but no effect on cognition or memory124. More recently, bilateral DBS of the NBM led to stable or improved cognitive function 12 months after surgery in four out of six patients with mild to moderate AD127. In a follow-up study, the same procedure was applied to two patients at an earlier phase of the disease133. In both of them, overall cognitive function was improved or stable during the first year of DBS and either returned to baseline or degraded during the following year, while memory function was stable over these 2 years. Additionally, clock agnosia completely disappeared in one patient. Although inconclusive due to the small number of subjects, these results motivate future work and possible refinements to modulate cholinergic brain networks in AD. For example, intermittent stimulation of the NBM (20 s every minute) has been shown to improve working memory in NHPs, whereas continuous stimulation impaired memory175.
DBS of the fornix in AD and epileptic patients
The fornix is a bundle of fibers that contains both the main output tract of the hippocampus and some afferent fibers from areas such as the basal forebrain. Because its integrity is critical to episodic memory, it appears as a target of choice for modulating memory function. In 2008, Lozano and colleagues made the serendipitous observation that DBS of the fornix evoked vivid recollection of memories in a patient treated with hypothalamic DBS for obesity176. This prompted them to perform a phase I safety and feasibility study of fornix DBS in six patients with mild AD177. DBS was delivered continuously, with stimulation amplitudes lower than those used to evoke vivid memories. Positron Emission Tomography (PET) revealed that glucose metabolism increased after both 1 month and 1 year of stimulation, especially in the temporal and parietal cortices. Some patients showed stable or improved cognitive or memory functions at six and/or 12 months post-surgery, but the results were not consistent across subjects and varied with the type of assessment. Similar results were later reported in one patient with mild AD by an independent group178.
In a subsequent report, the authors investigated the volume change of the hippocampus, mammillary bodies, and fornix following 1 year of stimulation in the same six patients179. Unlike the expected hippocampal atrophy normally seen in AD patients, an increased hippocampal volume was observed bilaterally in two of the patients, including the one who showed improved cognitive performance after one year. By contrast, bilateral hippocampal volume increase was never observed among the 25 matched subjects from the AD Neuroimaging Initiative database179.
In a follow-up double-blinded phase II clinical trial, DBS was applied bilaterally to the fornix of 42 patients with mild AD180,181. Half of the patients received DBS immediately after implantation, while the other half only received DBS after 1 year of sham stimulation (i.e. with the stimulator turned off). Fornix DBS significantly increased cerebral glucose metabolism after 6 months in the group receiving stimulation, but it did not reveal any statistically significant effect on cognitive functions. Post-hoc analyses indicated a non-significant trend towards cognitive benefits in older patients (>65-year old), which may be due to the fact that younger patients are usually affected by more malignant forms of the disease.
In all the above studies, stimulation was applied at 130 Hz as commonly used for PD. However, several animal studies have investigated other frequencies and patterns of fornix DBS, in particular theta and theta-burst stimulation182,183. Theta-burst stimulation consists of high-frequency bursts (typically 4–20 pulses at 100–500 Hz), nested within a low-frequency envelope in the theta range (4–8 Hz). It is optimal for inducing Long-Term Potentiation (LTP) in the hippocampus in vitro184, and superior to both conventional high-frequency stimulation (130 Hz) and low-frequency theta stimulation for improving learning and memory in a rat pharmacological model of amnesia182 and a rat model of traumatic brain injury183. Based on these results, one clinical study tested the immediate effects on memory of theta-burst DBS of the fornix in four epileptic patients185. They showed an immediate and reversible improvement in a visuospatial memory task in all four patients, but inconsistent results in other tasks related to verbal memory or naming.
DBS of the entorhinal area and other limbic structures in epileptic patients
Because of its major role in learning and memory, the hippocampus appears as an obvious neuromodulation target for memory enhancement. However, direct electrical stimulation of the hippocampus was shown to impair memory processes in early reports from 1985 that used amplitudes above the threshold to elicit epileptiform afterdischarges186,187. More recently, a similar impairment was also shown with stimulation amplitudes below the afterdischarge threshold188,189.
Instead of the hippocampus itself, its afferents from the EC could also be stimulated. In a first pivotal study in seven epileptic patients, Suthana and colleagues showed that DBS of the EC during spatial memory encoding improved subsequent retrieval at the group level190. The same stimulation applied to the hippocampus did not reveal any significant effect. However, in an attempt to replicate these results, Jacobs and colleagues found that DBS of either the EC or the hippocampus during spatial and verbal memory encoding actually disrupted subsequent retrieval191, probably due to differences in the exact DBS location between the two studies. In follow-up work, theta-burst microstimulation, known to optimally induce hippocampal LTP, was delivered to the right EC through thin microelectrodes during memory encoding, resulting in improved performance on a face recognition task192. The same group also compared the effects of stimulating the grey versus white matter of the entorhinal area, the lateralization of these effects, and the impact of electrode size (macro- versus microelectrode) in 22 epileptic patients193. Importantly, they identified the right entorhinal white matter (so-called angular bundle) as the optimal stimulation site to improve memory encoding. By contrast, electrode size was not a significant predictor of memory performance.
Further research is still needed to ask whether these results are robust across subjects and experimental conditions. For example, one study showed improvement in memory encoding during direct hippocampal stimulation, and this discrepancy was attributed to differences in the memory tasks194. Memory enhancement or disruption could also be achieved by stimulating other targets within the limbic circuit, such as the amygdala195 or the posterior cingulate cortex196. Finally, other approaches such as multisite synchronized197 and/or closed-loop stimulation198 within the limbic circuit could be used to refine these protocols, as detailed in the last paragraph of this section.
Direct cortical stimulation of neocortical areas in epileptic patients
In parallel with DBS of limbic structures, the effects of neocortical stimulation on memory performance have also been investigated. The earliest report that temporal lobe stimulation can evoke visual and auditory experience of past memories dates back to Penfield and Perot in 1963199. Recently, a series of collaborative studies led by Worrell and Kahana, respectively, at the Mayo Clinic and the University of Pennsylvania revisited this concept200,201,202,203.
Kucewicz and colleagues investigated the effects of electrical stimulation of four structures (hippocampus, parahippocampal neocortex, PFC and temporal cortex) in 22 patients with epilepsy (each structure was stimulated in only a subset of the patients)200,201. Stimulation was delivered during the encoding phase of a verbal memory task using depth electrodes and subdural ECoG grids. They found that only stimulation of the lateral temporal cortex was able to enhance subsequent retrieval, which was observed both at the group level and in two of the four tested participants200. Electrophysiologically, word presentation induced high-gamma activity (62-118 Hz) in several areas. Stimulation caused an increase of this induced activity in the lateral temporal cortex, which was associated with memory enhancement, but a decrease in the other brain areas. Therefore, high-gamma activity could be a reliable biomarker for predicting stimulation effects201.
Additionally, Ezzyat and colleagues trained machine learning algorithms to discriminate between trials of good and bad encoding based on neural activity preceding the stimulation period202. They showed that stimulation delivered during trials of bad encoding improved memory performance, while stimulation during good encoding trials decreased performance. This dependency on the encoding state was deemed to account for the variability observed in the effects of stimulation across patients and trials.
Closed-loop and multi-site approaches for intracranial neuromodulation of memory
All results described so far employed open-loop stimulation delivered either continuously or applied during memory encoding to improve subsequent retrieval. Such protocols do not take into account the ongoing activity of the implanted brain areas, which is influenced not only by the stimulation but also by intrinsic changes in neural activity. To mitigate this limitation, the patterns and parameters of stimulation could be adjusted in real-time based on the current neural state, a strategy called closed-loop stimulation198,203,204.
A closed-loop approach was demonstrated by Ezzyat and colleagues following their discovery that stimulation effects depend on the ongoing neural state. Using multivariate classifiers able to classify trials with good or bad encoding202, they delivered stimulation to the lateral temporal cortex only when a trial of bad encoding was detected203. This protocol improved retrieval in a more consistent way than open-loop stimulation.
To further refine these techniques, neurostimulation could also be applied to multiple sites, corresponding to different nodes of the distributed memory network, with the appropriate phase relationship to mimic physiological processes. In this spirit, Alagapan and colleagues applied network-targeted stimulation, which was delivered at two cortical locations (frontal and parietal) either in-phase or in anti-phase, within the theta or alpha range depending on the participant205. Tested in three epileptic patients during a working memory task, this strategy affected phase synchronizations within the targeted network and improved working memory.
Finally, the most versatile neurotechnologies should probably include both aspects: closed-loop and multisite stimulation. A memory neuroprosthesis based on these concepts was pioneered by Berger and colleagues in rats206, later tested in NHPs207, and recently in epileptic patients198,208. The authors extracted neuronal signals from microelectrodes implanted into the CA1 and CA3 fields of the hippocampus to build a multi-input multi-output non-linear model able to predict the spiking activity of CA1 neurons based on the activity of CA3 neurons. This model was then used to control spatiotemporal patterns of electrical stimulation delivered to the CA1 field based on real-time recordings from CA3 neurons. Stimulation applied during the encoding phase of a working memory task led to improved performance. Importantly, the effect in epileptic patients was more pronounced in those having a prior history of brain injury and memory impairments, demonstrating the suitability of this approach for memory disorders208. This conceptually very interesting approach would be suitable when memory impairments originate specifically from dysfunction within the hippocampus, but does not target the overall large-scale brain networks involved in episodic memory.
Outlook: towards large-scale memory neuroprostheses
The need for novel neurostimulation strategies
Considering that episodic memory relies on a distributed network of brain areas that interact via long-range connections through phase synchronization mechanisms and phase-amplitude CFC, we hypothesize that a physiological stimulation paradigm for memory enhancement or restoration should include the following key features:
• Spatial specificity: different areas (hippocampus, EC, PFC, etc.) should be targeted independently by the stimulation.
• Spectral specificity: each area should be stimulated with a particular frequency or a set of superimposed frequencies (e.g. theta to promote long-range interactions and gamma for local information processing).
• Temporal specificity: this aspect can be divided into two different timescales. On a long timescale (on the order of hundreds of milliseconds to seconds), the pattern of brain activation should depend on the task being carried out (e.g. encoding or retrieval) and potentially on the stage within this task. On a short timescale (tens of milliseconds), the phase of each stimulation pulse within each area should be carefully adjusted so that different areas are stimulated with the appropriate phase relationship (in-phase, out-of-phase, etc.).
These different features require a technological framework for spatially, temporally, and spectrally patterned neuromodulation of the large-scale episodic memory network to improve memory encoding and retrieval (Fig. 4), which critically depends on two key elements:

Future neuroprosthetic systems for memory restoration may benefit from distributed multi-electrode implants targeting large-scale brain networks, combining depth electrodes, ECoG grids and potentially novel electrode technologies for combined recordings and stimulation. Field potentials measured by these implants would then be sent to computing devices via high-channel-count recording systems. These computing devices would extract in real-time specific spectral features of these raw neural signals, such as the instantaneous power, phase and phase-amplitude coupling of theta and gamma oscillations in hippocampal structures. These features would then be used as control signals in closed-loop algorithms that deliver spatially, temporally and spectrally patterned neurostimulation protocols to enhance the neural features of successful memory processes (encoding or retrieval). These complex stimulation patterns would next be applied to the distributed neural implants via high-channel-count neurostimulators. Neurostimulation would finally affect neuronal activity through mechanisms that depend on the geometry of local fibers, which can be explained or theoretically predicted by computational models of neurostimulation. Schematics of high-channel-count recording and stimulation systems were adapted with permission from Blackrock Neurotech.
• the ability to simultaneously record and stimulate the large-scale episodic memory network through novel neural implants that target distributed brain areas, for example through intracerebral depth macroelectrodes classically used for stereo-encephalography (stereo-EEG), and subdural ECoG implants.
• the development of novel neurostimulation strategies that promote physiological oscillations throughout this distributed network, which require patterned stimulation protocols (such as theta burst) and the ability to trigger them (responsive stimulation) or adjust their stimulation parameters in real-time based on ongoing brain activity (adaptive or closed-loop stimulation).
Novel electrode technologies: opportunities and limitations
Both depth macroelectrodes and ECoG grids used in epileptic patients take their roots back to the middle of the 20th century209,210,211. Although clinically validated, these adopted neurotechnologies suffer from several impediments, namely the relatively small number of channels and large recording/stimulation volumes, which may limit the spatial selectivity required for the fine manipulation of cognitive and memory functions. Cognitive neuroprostheses might therefore benefit from more refined electrode technologies that can target both deep and cortical structures in large-scale brain networks at multiple spatial scales9.
For example, recent advances in micro- and nano-fabrication technologies have led to the development of silicon probes, which enable the simultaneous recording of tens and even thousands of neurons deep inside the brain212,213,214,215,216,217,218,219,220,221,222,223. These ultra-thin shanked probes (cross section: 20–120 μm, length: up to 20 mm) allow simultaneous recordings from up to 1356 sites224. Some probes (Neurotech Alliance, E-beam, Silicon microprobes, etc.) are designed with multiple shanks for greater cortical and subcortical area coverage. For single-shank probes such as Neuropixels, larger brain areas can be targeted using multiple probes, a strategy that allowed to record from approximately 30,000 neurons located in 42 regions in the mouse brain224,225.
The use of such high-density multi-electrode and multi-shank probes presents both advantages and challenges. Compared to larger conventional electrodes, these much smaller silicon probes have reduced neural tract damage. However, the ultra-small cross-sectional area and stiffness may lead to electrode drift due to blood flow, and make breakages frequent212,215. Like conventional electrodes, silicon probes also show inflammatory response and tissue scarring that deteriorates the quality of signals over time212, but recordings up to 3 months are feasible226. To mitigate tissue scarring and electrode drifts, a dissolvable shaft has also been developed212. Notably, tissue response was not observed until about 1 year for some probes216,217,218, while for others the duration was as short as 2–3 weeks226.
Most of these studies have been focused on rodents, with only a few reports in NHPs223,227 and humans undergoing intraoperative neurosurgical procedures219,228. Challenges associated with these early clinical demonstrations include probe breakage and recording of fewer neurons, due to the short experimental duration (<1 h) and the time necessary for neuronal activity to stabilize. An additional limitation is that Neuropixels probes are currently not suitable for neurostimulation224. Despite these current challenges, they represent an attractive option for the next-generation neuroprostheses targeting large-scale brain networks.
The use of NHPs in cognitive neuroprosthetic research
To allow the translation of cognitive neuroprosthetic technologies into clinical practice, and given the regulatory constraints on the development of novel medical devices, we believe that large animal models are required. Because of their similarity in anatomy, functional connections and default-mode network topology with the human brain229,230, NHPs appear as the best species for investigating large-scale network dynamics and developing invasive neuromodulation strategies that should eventually be used in humans.
Like humans, NHPs develop task-specific cognitive impairments associated with normal ageing, in particular in the fields of visuospatial or spatial working memory, which depend on the PFC alone or in interaction with hippocampal-temporal structures231,232,233. These deficits mainly mimic normal cognitive ageing and are associated with amyloid-beta senile plaques. In addition to natural ageing, memory deficits can be reversibly induced by pharmacological agents such as the anticholinergic drug scopolamine, which has been shown to impair working memory in both rhesus and cynomolgus macaques during various tasks234,235,236.
Until recently, studies had failed to find other signs of AD, such as tau pathology, neurofibrillary tangles, or behavioral symptoms in most NHP species237,238, except in the mouse lemur Microcebus murinus239. However, this picture has recently changed, with several groups finding evidence of tau pathology similar to human AD in some (but not all) of aged (more than 20 years) African green monkeys240,241, rhesus242,243 and cynomolgus macaques244. To artificially replicate the pathophysiological features of AD in NHP, several strategies have also been tested such as the induction of insulin deficiency using streptozotocin245,246, the intraventricular injection of synthetic amyloid-beta oligomers247, or the injection into the EC of a viral vector expressing a double tau mutation248. Finally, the advent of genetic engineering tools in NHPs has recently led to the generation of marmoset monkeys carrying mutations in the PSEN1 gene, which is involved in familial forms of AD249,250. However, none of these NHP models of AD has been validated at a behavioral level, which is essential for testing the efficacy of cognitive neuroprostheses.
Computational models of neurostimulation in the context of memory processes
Computational modeling has been extensively used to better understand the effects of electrical neurostimulation, ranging from DBS in PD, to PNS, SCS, cochlear implants, and retinal prostheses (e.g.251). However, in the context of memory, even state-of-the-art computational models are limited in their ability to integrate hippocampal oscillations, memory processes, and the effects of electrical stimulation on local circuits and distant connected areas in large-scale brain networks. From a neuroengineering viewpoint, there is therefore an urgent need to develop novel biologically realistic computational models that guide electrode design, placement and stimulation protocols to enhance the efficacy of memory neuroprostheses252,253.
Computational models of theta-gamma neural oscillations and memory processes
Neural oscillations such as theta, gamma and their CFC have been modeled at different levels of resolution, from abstract neural masses to more biophysically realistic conductance-based models. Neural masses comprising two interconnected populations of excitatory and inhibitory neurons can produce gamma oscillations in the presence of an external input provided at a theta frequency, hence leading the theta-nested gamma oscillations254,255. Such oscillations also emerge from a conductance-based model of the hippocampal formation containing single-compartment excitatory and inhibitory neurons in the EC, dentate gyrus, CA3 and CA1 fields of the hippocampus, still in the presence of an external theta input256. Theta-gamma CFC also appears in a conductance-based multicompartment model of neocortex, which includes two cortical layers, a hypercolumn and minicolumn structure, and a built-in attractor network that represents the reactivation of specific cell assemblies during memory retrieval257,258.
Linking these oscillations with memory processes represents another major challenge. One such attempt explored the biophysical mechanisms of memory encoding and retrieval in a multicompartment model of the hippocampal CA1 field, demonstrating that the modulation of dendritic and somatic inhibition by the theta rhythm influences LTP and cell output259. Furthermore, another multicompartment study of the hippocampal CA3 showed that cholinergic deprivation leads to a slowing of gamma oscillations produced by the network, which can potentially affect the ability to store and recall information accurately260. These works provide the necessary leverage to investigate how neural oscillations and memory processes are altered in neurological disorders such as AD.
Computational models of extracellular electrical neurostimulation
Most models of electrical stimulation have focused on the mechanisms underlying DBS in motor disorders such as PD (e.g. refs. 261,262,263,264), often approximating stimulation as an intracellular current that enters into the soma. However, the effects of extracellular electrical stimulation are much more complex, as illustrated by a series of modeling studies that date back to the 80s265,266,267,268,269. McNeal and Rattay initially introduced cable models to describe axons as a series of capacitance and resistance circuits, similar to the Hodgkin-Huxley formalism but as a cable following the axonal trajectory265,266. They found that the level of activation of nerve compartments depends on the second spatial derivative of the electrical potential along the membrane. Subsequent studies investigated in more detail the effects of tissue properties268, excitation thresholds269 and action potential propagation267. Cable models successfully explained the mechanisms underlying various neuroprostheses251 and reconciled contradictory hypotheses on the effects of DBS in PD270.
Despite the wealth of computational work that involves neurostimulation of peripheral nerves or BG, very few studies have attempted to replicate hippocampal stimulation in a biologically realistic way. A detailed model of the dentate gyrus, which involves granule cells and entorhinal afferents271, was confronted with experimental data during electrical stimulation in rat hippocampal slices252. This study demonstrated the ability to predict the optimal electrode placement to maximize population response within localized structures. However, models that integrate more widespread brain areas in a biologically realistic way are still lacking.
Recently, a neuroimaging and computational pipeline for creating personalized computational models of the spinal cord was developed to predict the effects of spinal cord stimulation protocols on the recruitment of afferent fibers in lumbosacral spinal segments14. This pipeline guided the design of new electrode arrays and their accurate placement during surgery. We envision that a similar approach combining neuroimaging and computational modeling can be used to create personalized models of the hippocampal formation and predict the effects of electrical stimulation at various locations within this structure and with different stimulation parameters and control protocols.
Conclusion
Since their very first prototypes about half a century ago, sensorimotor neuroprostheses have steadily evolved, paralleling progress in neurophysiology and engineering sciences. While some of these technologies are already clinically accepted, others are still making their way through the hurdles of clinical and regulatory validation, from preclinical and pilot studies to multicentric clinical trials. Neuromodulation and neuroprosthetic technologies have also received considerable attention from the scientific community for their potential in treating cognitive disorders. Recent exploratory studies open exciting avenues for the future development of such neuroprosthetic systems that affect neural oscillations throughout large-scale brain networks, palliate the associated deficits and potentially trigger neuroplasticity. We believe that these endeavors will require the elaboration of novel neural implants and neurostimulation protocols, the use of appropriate preclinical models to test them, and will be refined by computational approaches derived from neuroimaging data.
References
Schnitzler, A. & Gross, J. Normal and pathological oscillatory communication in the brain. Nat. Rev. Neurosci. 6, 285–296 (2005).
Uhlhaas, P. J. & Singer, W. Neural synchrony in brain disorders: relevance for cognitive dysfunctions and pathophysiology. Neuron 52, 155–168 (2006).
Nicolo, P. et al. Coherent neural oscillations predict future motor and language improvement after stroke. Brain 138, 3048–3060 (2015).
Pevzner, A., Izadi, A., Lee, D. J., Shahlaie, K. & Gurkoff, G. G. Making waves in the brain: what are oscillations, and why modulating them makes sense for brain injury. Front. Syst. Neurosci. 10, (2016).
Nishimura, Y., Morichika, Y. & Isa, T. A subcortical oscillatory network contributes to recovery of hand dexterity after spinal cord injury. Brain 132, 709–721 (2009).
Herrmann, C. S., Strüber, D., Helfrich, R. F. & Engel, A. K. EEG oscillations: from correlation to causality. Int J. Psychophysiol. 103, 12–21 (2016).
Borton, D., Micera, S., Millán, J., del, R. & Courtine, G. Personalized neuroprosthetics. Sci. Transl. Med. 5, 210rv2 (2013).
Lebedev, M. A. & Nicolelis, M. A. L. Brain-machine interfaces: from basic science to neuroprostheses and neurorehabilitation. Physiol. Rev. 97, 767–837 (2017).
Nurmikko, A. Challenges for large-scale cortical interfaces. Neuron 108, 259–269 (2020).
Editorial. The painstaking pace of bioelectronic interfaces. Nat. Biomed. Eng. 4, 933–934 (2020).
Benabid, A. L. Deep brain stimulation for Parkinson’s disease. Curr. Opin. Neurobiol. 13, 696–706 (2003).
Capogrosso, M. et al. A brain-spine interface alleviating gait deficits after spinal cord injury in primates. Nature 539, 284–288 (2016).
Wagner, F. B. et al. Targeted neurotechnology restores walking in humans with spinal cord injury. Nature 563, 65–71 (2018).
Rowald, A. et al. Activity-dependent spinal cord neuromodulation rapidly restores trunk and leg motor functions after complete paralysis. Nat. Med. 28, 260–271 (2022).
Mankin, E. A. & Fried, I. Modulation of human memory by deep brain stimulation of the entorhinal-hippocampal circuitry. Neuron 106, 218–235 (2020).
Prochazka, A. Motor Neuroprostheses. Compr. Physiol. 9, 127–148 (2018).
Liberson, W. T., Holmquest, H. J., Scot, D. & Dow, M. Functional electrotherapy: stimulation of the peroneal nerve synchronized with the swing phase of the gait of hemiplegic patients. Arch. Phys. Med Rehabil. 42, 101–105 (1961).
Nashold, B. S. et al. Electromicturition in paraplegia: implantation of a spinal neuroprosthesis. Proc. Veterans Adm. Spinal Cord. Inj. Conf. 18, 161–165 (1971).
Wolpaw, J. R. et al. Brain-computer interface technology: a review of the first international meeting. IEEE Trans. Rehabil. Eng. 8, 164–173 (2000).
Taylor, D. M., Tillery, S. I. H. & Schwartz, A. B. Direct cortical control of 3D neuroprosthetic devices. Science 296, 1829–1832 (2002).
Marquez-Chin, C. & Popovic, M. R. Functional electrical stimulation therapy for restoration of motor function after spinal cord injury and stroke: a review. Biomed. Eng. Online 19, 34 (2020).
Peckham, P. H. & Knutson, J. S. Functional electrical stimulation for neuromuscular applications. Annu. Rev. Biomed. Eng. 7, 327–360 (2005).
Popovic, M. R., Popovic, D. B. & Keller, T. Neuroprostheses for grasping. Neurological Res. 24, 443–452 (2002).
Ho, C. H. et al. Functional electrical stimulation and spinal cord injury. Phys. Med Rehabil. Clin. N. Am. 25, 631–654 (2014).
Kapadia, N. et al. A randomized trial of functional electrical stimulation for walking in incomplete spinal cord injury: effects on walking competency. J. Spinal Cord. Med. 37, 511–524 (2014).
Sinacore, D. R., Delitto, A., King, D. S. & Rose, S. J. Type II fiber activation with electrical stimulation: a preliminary report. Phys. Ther. 70, 416–422 (1990).
Trimble, M. H. & Enoka, R. M. Mechanisms underlying the training effects associated with neuromuscular electrical stimulation. Phys. Ther. 71, 273–280 (1991). discussion 280-282.
Heyters, M., Carpentier, A., Duchateau, J. & Hainaut, K. Twitch analysis as an approach to motor unit activation during electrical stimulation. Can. J. Appl Physiol. 19, 451–461 (1994).
Seáñez, I., Capogrosso, M., Minassian, K. & Wagner, F.B. Spinal Cord Stimulation to Enable Leg Motor Control and Walking in People with Spinal Cord Injury. In: Neurorehabilitation Technology. (eds Reinkensmeyer, D.J., Marchal-Crespo, L., Dietz, V.) (Springer, Cham. 2022).
Rattay, F., Minassian, K. & Dimitrijevic, M. R. Epidural electrical stimulation of posterior structures of the human lumbosacral cord: 2. quantitative analysis by computer modeling. Spinal Cord. 38, 473–489 (2000).
Capogrosso, M. A computational model for epidural electrical stimulation of spinal sensorimotor circuits. J. Neurosci. 33, 19326–19340 (2013).
Barolat, G., Myklebust, J. B. & Wenninger, W. Enhancement of voluntary motor function following spinal cord stimulation-case study. Appl Neurophysiol. 49, 307–314 (1986).
Dimitrijevic, M. R., Gerasimenko, Y. & Pinter, M. M. Evidence for a spinal central pattern generator in humans. Ann. N. Y. Acad. Sci. 860, 360–376 (1998).
Minassian, K., Hofstoetter, U. S., Dzeladini, F., Guertin, P. A. & Ijspeert, A. The human central pattern generator for locomotion: does it exist and contribute to walking? Neuroscientist 23, 649–663 (2017).
Jilge, B. et al. Initiating extension of the lower limbs in subjects with complete spinal cord injury by epidural lumbar cord stimulation. Exp. Brain Res. 154, 308–326 (2004).
Minassian, K. et al. Stepping-like movements in humans with complete spinal cord injury induced by epidural stimulation of the lumbar cord: electromyographic study of compound muscle action potentials. Spinal Cord. 42, 401–416 (2004).
Carhart, M. R., He, J., Herman, R., D’Luzansky, S. & Willis, W. T. Epidural spinal-cord stimulation facilitates recovery of functional walking following incomplete spinal-cord injury. IEEE Trans. Neural Syst. Rehabil. Eng. 12, 32–42 (2004).
Herman, R., He, J., D’Luzansky, S., Willis, W. & Dilli, S. Spinal cord stimulation facilitates functional walking in a chronic, incomplete spinal cord injured. Spinal Cord. 40, 65–68 (2002).
Huang, H., He, J., Herman, R. & Carhart, M. R. Modulation effects of epidural spinal cord stimulation on muscle activities during walking. IEEE Trans. Neural Syst. Rehabil. Eng. 14, 14–23 (2006).
Harkema, S. et al. Effect of epidural stimulation of the lumbosacral spinal cord on voluntary movement, standing, and assisted stepping after motor complete paraplegia: a case study. Lancet 377, 1938–1947 (2011).
Angeli, C. A., Edgerton, V. R., Gerasimenko, Y. P. & Harkema, S. J. Altering spinal cord excitability enables voluntary movements after chronic complete paralysis in humans. Brain 137, 1394–1409 (2014).
Rejc, E., Angeli, C. & Harkema, S. Effects of lumbosacral spinal cord epidural stimulation for standing after chronic complete paralysis in humans. PLoS ONE 10, e0133998 (2015).
Grahn, P. J. et al. Enabling task-specific volitional motor functions via spinal cord neuromodulation in a human with paraplegia. Mayo Clin. Proc. 92, 544–554 (2017).
Rejc, E., Angeli, C. A., Atkinson, D. & Harkema, S. J. Motor recovery after activity-based training with spinal cord epidural stimulation in a chronic motor complete paraplegic. Sci. Rep. 7, 13476 (2017).
Angeli, C. A. et al. Recovery of over-ground walking after chronic motor complete spinal cord injury. N. Engl. J. Med. 379, 1244–1250 (2018).
Gill, M. L. et al. Neuromodulation of lumbosacral spinal networks enables independent stepping after complete paraplegia. Nat. Med. 24, 1677–1682 (2018).
Wenger, N. et al. Spatiotemporal neuromodulation therapies engaging muscle synergies improve motor control after spinal cord injury. Nat. Med. 22, 138–145 (2016).
Capogrosso, M. et al. Configuration of electrical spinal cord stimulation through real-time processing of gait kinematics. Nat. Protoc. 13, 2031–2061 (2018).
Kathe, C. et al. The neurons that restore walking after paralysis. Nature 611, 540–547 (2022).
Pesaran, B. et al. Investigating large-scale brain dynamics using field potential recordings: analysis and interpretation. Nat. Neurosci. 21, 903–919 (2018).
Georgopoulos, A. P., Kalaska, J. F., Caminiti, R. & Massey, J. T. On the relations between the direction of two-dimensional arm movements and cell discharge in primate motor cortex. J. Neurosci. 2, 1527–1537 (1982).
Wessberg, J. et al. Real-time prediction of hand trajectory by ensembles of cortical neurons in primates. Nature 408, 361–365 (2000).
Serruya, M. D., Hatsopoulos, N. G., Paninski, L., Fellows, M. R. & Donoghue, J. P. Instant neural control of a movement signal. Nature 416, 141–142 (2002).
Velliste, M., Perel, S., Spalding, M. C., Whitford, A. S. & Schwartz, A. B. Cortical control of a prosthetic arm for self-feeding. Nature 453, 1098–1101 (2008).
Hochberg, L. R. et al. Neuronal ensemble control of prosthetic devices by a human with tetraplegia. Nature 442, 164–171 (2006).
Hochberg, L. R. et al. Reach and grasp by people with tetraplegia using a neurally controlled robotic arm. Nature 485, 372–375 (2012).
Collinger, J. L. et al. High-performance neuroprosthetic control by an individual with tetraplegia. Lancet 381, 557–564 (2013).
Handelman, D. A. et al. Shared control of bimanual robotic limbs with a brain-machine interface for self-feeding. Front. Neurorobot. 16, 918001 (2022).
Willett, F. R., Avansino, D. T., Hochberg, L. R., Henderson, J. M. & Shenoy, K. V. High-performance brain-to-text communication via handwriting. Nature 593, 249–254 (2021).
Vansteensel, M. J. et al. Fully implanted brain-computer interface in a locked-in patient with ALS. N. Engl. J. Med. 375, 2060–2066 (2016).
Benabid, A. L. et al. An exoskeleton controlled by an epidural wireless brain-machine interface in a tetraplegic patient: a proof-of-concept demonstration. Lancet Neurol. 18, 1112–1122 (2019).
Moses, D. A. et al. Neuroprosthesis for decoding speech in a paralyzed person with anarthria. N. Engl. J. Med. 385, 217–227 (2021).
Metzger, S. L. et al. Generalizable spelling using a speech neuroprosthesis in an individual with severe limb and vocal paralysis. Nat. Commun. 13, 6510 (2022).
Fallegger, F. et al. MRI-compatible and conformal electrocorticography grids for translational research. Adv. Sci. 8, 2003761 (2021).
Bouton, C. E. et al. Restoring cortical control of functional movement in a human with quadriplegia. Nature 533, 247–250 (2016).
Ajiboye, A. B. et al. Restoration of reaching and grasping movements through brain-controlled muscle stimulation in a person with tetraplegia: a proof-of-concept demonstration. Lancet 389, 1821–1830 (2017).
Lorach, H., Charvet, G., Bloch, J. & Courtine, G. Brain-spine interfaces to reverse paralysis. Natl Sci. Rev. 9, nwac009 (2022).
Raspopovic, S., Valle, G. & Petrini, F. M. Sensory feedback for limb prostheses in amputees. Nat. Mater. 20, 925–939 (2021).
Clippinger, F. W., Avery, R. & Titus, B. R. A sensory feedback system for an upper-limb amputation prosthesis. Bull. Prosthet. Res. 10–22, 247–258 (1974).
Clippinger, F. W., Seaber, A. V., McElhaney, J. H., Harrelson, J. M. & Maxwell, G. M. Afferent sensory feedback for lower extremity prosthesis. Clin. Orthop. Relat. Res. 169, 202–206 (1982).
Ortiz-Catalan, M., Håkansson, B. & Brånemark, R. An osseointegrated human-machine gateway for long-term sensory feedback and motor control of artificial limbs. Sci. Transl. Med. 6, 257re6 (2014).
Ortiz-Catalan, M., Mastinu, E., Sassu, P., Aszmann, O. & Brånemark, R. Self-contained neuromusculoskeletal arm prostheses. N. Engl. J. Med. 382, 1732–1738 (2020).
Tan, D. W. et al. A neural interface provides long-term stable natural touch perception. Sci. Transl. Med. 6, 257ra138 (2014).
Charkhkar, H. et al. High-density peripheral nerve cuffs restore natural sensation to individuals with lower-limb amputations. J. Neural Eng. 15, 056002 (2018).
Dhillon, G. S. & Horch, K. W. Direct neural sensory feedback and control of a prosthetic arm. IEEE Trans. Neural Syst. Rehabil. Eng. 13, 468–472 (2005).
Rossini, P. M. et al. Double nerve intraneural interface implant on a human amputee for robotic hand control. Clin. Neurophysiol. 121, 777–783 (2010).
Horch, K., Meek, S., Taylor, T. G. & Hutchinson, D. T. Object discrimination with an artificial hand using electrical stimulation of peripheral tactile and proprioceptive pathways with intrafascicular electrodes. IEEE Trans. Neural Syst. Rehabil. Eng. 19, 483–489 (2011).
Raspopovic, S. et al. Restoring natural sensory feedback in real-time bidirectional hand prostheses. Sci. Transl. Med. 6, 222ra19 (2014).
Petrini, F. M. et al. Six-month assessment of a hand prosthesis with intraneural tactile feedback. Ann. Neurol. 85, 137–154 (2019).
Petrini, F. M. et al. Sensory feedback restoration in leg amputees improves walking speed, metabolic cost and phantom pain. Nat. Med. 25, 1356–1363 (2019).
Petrini, F. M. et al. Enhancing functional abilities and cognitive integration of the lower limb prosthesis. Sci. Transl. Med. 11, eaav8939 (2019).
Davis, T. S. et al. Restoring motor control and sensory feedback in people with upper extremity amputations using arrays of 96 microelectrodes implanted in the median and ulnar nerves. J. Neural Eng. 13, 036001 (2016).
Wendelken, S. et al. Restoration of motor control and proprioceptive and cutaneous sensation in humans with prior upper-limb amputation via multiple Utah Slanted Electrode Arrays (USEAs) implanted in residual peripheral arm nerves. J. Neuroeng. Rehabil. 14, 121 (2017).
Penfield, W. & Boldrey, E. Somatic motor and sensory representation in the cerebral cortex of man as studied by electrical stimulation1. Brain 60, 389–443 (1937).
Romo, R., Hernández, A., Zainos, A. & Salinas, E. Somatosensory discrimination based on cortical microstimulation. Nature 392, 387–390 (1998).
Romo, R., Hernández, A., Zainos, A., Brody, C. D. & Lemus, L. Sensing without touching: psychophysical performance based on cortical microstimulation. Neuron 26, 273–278 (2000).
Na, F., W, D., Tl, H., Ma, L. & Ma, N. Primate reaching cued by multichannel spatiotemporal cortical microstimulation. J. Neurosci. 27, 5593–5602 (2007).
Tabot, G. A. et al. Restoring the sense of touch with a prosthetic hand through a brain interface. Proc. Natl Acad. Sci. USA 110, 18279–18284 (2013).
Dadarlat, M. C., O’Doherty, J. E. & Sabes, P. N. A learning-based approach to artificial sensory feedback leads to optimal integration. Nat. Neurosci. 18, 138–144 (2015).
O’Doherty, J. E., Shokur, S., Medina, L. E., Lebedev, M. A. & Nicolelis, M. A. L. Creating a neuroprosthesis for active tactile exploration of textures. Proc. Natl Acad. Sci. USA 116, 21821–21827 (2019).
O’Doherty, J. E. et al. Active tactile exploration using a brain-machine-brain interface. Nature 479, 228–231 (2011).
Klaes, C. et al. A cognitive neuroprosthetic that uses cortical stimulation for somatosensory feedback. J. Neural Eng. 11, 056024 (2014).
Flesher, S. N. et al. Intracortical microstimulation of human somatosensory cortex. Sci. Transl. Med. 8, 361ra141 (2016).
Armenta Salas, M. et al. Proprioceptive and cutaneous sensations in humans elicited by intracortical microstimulation. Elife 7, e32904 (2018).
Hughes, C. L. et al. Perception of microstimulation frequency in human somatosensory cortex. Elife 10, e65128 (2021).
Hughes, C. L. et al. Neural stimulation and recording performance in human sensorimotor cortex over 1500 days. J. Neural Eng. 18, (2021).
Flesher, S. N. et al. A brain-computer interface that evokes tactile sensations improves robotic arm control. Science 372, 831–836 (2021).
Fifer, M. S. et al. Intracortical somatosensory stimulation to elicit fingertip sensations in an individual with spinal cord injury. Neurology 98, e679–e687 (2022).
Johnson, L. A. et al. Direct electrical stimulation of the somatosensory cortex in humans using electrocorticography electrodes: a qualitative and quantitative report. J. Neural Eng. 10, 036021 (2013).
Hiremath, S. V. et al. Human perception of electrical stimulation on the surface of somatosensory cortex. PLoS ONE 12, e0176020 (2017).
Lee, B. et al. Engineering artificial somatosensation through cortical stimulation in humans. Front Syst. Neurosci. 12, 24 (2018).
Kramer, D. R. et al. Technical considerations for generating somatosensation via cortical stimulation in a closed-loop sensory/motor brain-computer interface system in humans. J. Clin. Neurosci. 63, 116–121 (2019).
Valle, G. et al. Biomimetic intraneural sensory feedback enhances sensation naturalness, tactile sensitivity, and manual dexterity in a bidirectional prosthesis. Neuron 100, 37–45.e7 (2018).
Risso, G. et al. Optimal integration of intraneural somatosensory feedback with visual information: a single-case study. Sci. Rep. 9, 7916 (2019).
Rognini, G. et al. Multisensory bionic limb to achieve prosthesis embodiment and reduce distorted phantom limb perceptions. J. Neurol. Neurosurg. Psychiatry 90, 833–836 (2019).
Valle, G. et al. Mechanisms of neuro-robotic prosthesis operation in leg amputees. Sci. Adv. 7, eabd8354 (2021).
Preatoni, G., Valle, G., Petrini, F. M. & Raspopovic, S. Lightening the perceived prosthesis weight with neural embodiment promoted by sensory feedback. Curr. Biol. 31, 1065–1071.e4 (2021).
Benabid, A. L., Pollak, P., Louveau, A., Henry, S. & de Rougemont, J. Combined (thalamotomy and stimulation) stereotactic surgery of the VIM thalamic nucleus for bilateral Parkinson disease. Appl. Neurophysiol. 50, 344–346 (1987).
Aarsland, D. & Kurz, M. W. The epidemiology of dementia associated with Parkinson disease. J. Neurol. Sci. 289, 18–22 (2010).
Shahlaie, K., Sperry, L., Wei, L. & Zhang, L. Current Research on Deep Brain Stimulation and Cognitive Impairment in Parkinson’s Disease. In: Dementia in Parkinson’s Disease - Everything you Need to Know (eds Zhang, L. & Olichney, J.M.) (2021).
Moustafa, A. A. et al. Motor symptoms in Parkinson’s disease: a unified framework. Neurosci. Biobehav. Rev. 68, 727–740 (2016).
Moustafa, A. A. et al. Interrelations between cognitive dysfunction and motor symptoms of Parkinson’s disease: behavioral and neural studies. Rev. Neurosci. 27, 535–548 (2016).
Benazzouz, A., Gross, C., Féger, J., Boraud, T. & Bioulac, B. Reversal of rigidity and improvement in motor performance by subthalamic high-frequency stimulation in MPTP-treated monkeys. Eur. J. Neurosci. 5, 382–389 (1993).
Pollak, P. et al. [Effects of the stimulation of the subthalamic nucleus in Parkinson disease]. Rev. Neurol. (Paris) 149, 175–176 (1993).
Siegfried, J. & Lippitz, B. Bilateral chronic electrostimulation of ventroposterolateral pallidum: a new therapeutic approach for alleviating all parkinsonian symptoms. Neurosurgery 35, 1126–1129 (1994). discussion 1129-1130.
Ramirez-Zamora, A. & Ostrem, J. L. Globus pallidus interna or subthalamic nucleus deep brain stimulation for parkinson disease: a review. JAMA Neurol. 75, 367 (2018).
Baizabal-Carvallo, J. F. & Alonso-Juarez, M. Low-frequency deep brain stimulation for movement disorders. Parkinsonism Relat. Disord. 31, 14–22 (2016).
Jia, F. et al. Deep brain stimulation at variable frequency to improve motor outcomes in Parkinson’s disease. Mov. Disord. Clin. Pr. 5, 538–541 (2018).
Combs, H. L. et al. Cognition and depression following deep brain stimulation of the subthalamic nucleus and globus Pallidus Pars Internus in Parkinson’s disease: A Meta-Analysis. Neuropsychol. Rev. 25, 439–454 (2015).
Gruber, D. et al. Longterm outcome of cognition, affective state, and quality of life following subthalamic deep brain stimulation in Parkinson’s disease. J. Neural Transm. (Vienna) 126, 309–318 (2019).
Kurcova, S. et al. Bilateral subthalamic deep brain stimulation initial impact on nonmotor and motor symptoms in Parkinson’s disease: An open prospective single institution study. Medicine (Baltimore) 97, e9750 (2018).
Petry-Schmelzer, J. N. et al. Non-motor outcomes depend on location of neurostimulation in Parkinson’s disease. Brain 142, 3592–3604 (2019).
Wilson, H., de Natale, E. R. & Politis, M. In Handbook of Clinical Neurology (eds. Swaab, D. F. et al.) Vol. 179, 189–205 (Elsevier, 2021).
Turnbull, I. M., McGeer, P. L., Beattie, L., Calne, D. & Pate, B. Stimulation of the basal nucleus of Meynert in senile dementia of Alzheimer’s type. A preliminary report. Appl. Neurophysiol. 48, 216–221 (1985).
Freund, H.-J. et al. Cognitive functions in a patient with Parkinson-dementia syndrome undergoing deep brain stimulation. Arch. Neurol. 66, 781–785 (2009).
Barnikol, T. T. et al. Changes in apraxia after deep brain stimulation of the nucleus basalis Meynert in a patient with Parkinson dementia syndrome. Mov. Disord. 25, 1519–1520 (2010).
Kuhn, J. et al. Deep brain stimulation of the nucleus basalis of Meynert in Alzheimer’s dementia. Mol. Psychiatry 20, 353–360 (2015).
Gratwicke, J. et al. Bilateral deep brain stimulation of the nucleus basalis of Meynert for Parkinson disease dementia: a randomized clinical trial. JAMA Neurol. 75, 169–178 (2018).
Nombela, C., Lozano, A., Villanueva, C. & Barcia, J. A. Simultaneous stimulation of the globus Pallidus Interna and the Nucleus Basalis of Meynert in the Parkinson-dementia syndrome. Dement Geriatr. Cogn. Disord. 47, 19–28 (2019).
Gratwicke, J. et al. Bilateral nucleus basalis of Meynert deep brain stimulation for dementia with Lewy bodies: a randomised clinical trial. Brain Stimul. 13, 1031–1039 (2020).
Kumbhare, D. et al. Nucleus Basalis of Meynert stimulation for dementia: theoretical and technical considerations. Front. Neurosci. 12, 614 (2018).
Nazmuddin, M., Philippens, I. H. C. H. M. & van Laar, T. Electrical stimulation of the nucleus basalis of meynert: a systematic review of preclinical and clinical data. Sci. Rep. 11, 11751 (2021).
Kuhn, J. et al. Deep brain stimulation of the nucleus basalis of Meynert in early stage of Alzheimer’s dementia. Brain Stimul. 8, 838–839 (2015).
Maltête, D. et al. Nucleus basalis of Meynert stimulation for lewy body dementia: a phase i randomized clinical trial. Neurology 96, e684–e697 (2021).
Wojtecki, L. et al. Frequency-dependent reciprocal modulation of verbal fluency and motor functions in subthalamic deep brain stimulation. Arch. Neurol. 63, 1273–1276 (2006).
Scangos, K. W., Carter, C. S., Gurkoff, G., Zhang, L. & Shahlaie, K. A pilot study of subthalamic theta frequency deep brain stimulation for cognitive dysfunction in Parkinson’s disease. Brain Stimul. 11, 456–458 (2018).
Lam, J. et al. Cognitive effects of theta frequency bilateral subthalamic nucleus stimulation in Parkinson’s disease: a pilot study. Brain Stimul. 14, 230–240 (2021).
Lee, D. J. et al. Acute low frequency dorsal subthalamic nucleus stimulation improves verbal fluency in Parkinson’s disease. Brain Stimul. 14, 754–760 (2021).
Ward, L. M. Synchronous neural oscillations and cognitive processes. Trends Cogn. Sci. 7, 553–559 (2003).
Bressler, S. L. & Menon, V. Large-scale brain networks in cognition: emerging methods and principles. Trends Cogn. Sci. 14, 277–290 (2010).
Siegel, M., Donner, T. H. & Engel, A. K. Spectral fingerprints of large-scale neuronal interactions. Nat. Rev. Neurosci. 13, 121–134 (2012).
Fell, J. & Axmacher, N. The role of phase synchronization in memory processes. Nat. Rev. Neurosci. 12, 105–118 (2011).
Headley, D. B. & Paré, D. Common oscillatory mechanisms across multiple memory systems. NPJ Sci. Learn. 2, 1 (2017).
Gruber, M. J. et al. Theta phase synchronization between the human hippocampus and prefrontal cortex increases during encoding of unexpected information: a case study. J. Cogn. Neurosci. 30, 1646–1656 (2018).
Bragin, A. et al. Gamma (40-100 Hz) oscillation in the hippocampus of the behaving rat. J. Neurosci. 15, 47–60 (1995).
Siapas, A. G., Lubenov, E. V. & Wilson, M. A. Prefrontal phase locking to hippocampal theta oscillations. Neuron 46, 141–151 (2005).
Canolty, R. T. et al. High gamma power is phase-locked to theta oscillations in human neocortex. Science 313, 1626–1628 (2006).
Paz, R., Bauer, E. P. & Paré, D. Theta synchronizes the activity of medial prefrontal neurons during learning. Learn Mem. 15, 524–531 (2008).
Sirota, A. et al. Entrainment of neocortical neurons and gamma oscillations by the hippocampal theta rhythm. Neuron 60, 683–697 (2008).
Colgin, L. L. et al. Frequency of gamma oscillations routes flow of information in the hippocampus. Nature 462, 353–357 (2009).
Tort, A. B. L., Komorowski, R. W., Manns, J. R., Kopell, N. J. & Eichenbaum, H. Theta-gamma coupling increases during the learning of item-context associations. Proc. Natl Acad. Sci. USA 106, 20942–20947 (2009).
Wulff, P. et al. Hippocampal theta rhythm and its coupling with gamma oscillations require fast inhibition onto parvalbumin-positive interneurons. Proc. Natl Acad. Sci. USA 106, 3561–3566 (2009).
Daume, J., Gruber, T., Engel, A. K. & Friese, U. Phase-amplitude coupling and long-range phase synchronization reveal frontotemporal interactions during visual working memory. J. Neurosci. 37, 313–322 (2017).
Salimpour, Y. & Anderson, W. S. Cross-frequency coupling based neuromodulation for treating neurological disorders. Front. Neurosci. 13, 125 (2019).
Preston, A. R. & Eichenbaum, H. Interplay of hippocampus and prefrontal cortex in memory. Curr. Biol. 23, R764–R773 (2013).
Rolls, E. T. The storage and recall of memories in the hippocampo-cortical system. Cell Tissue Res. 373, 577–604 (2018).
Brincat, S. L. & Miller, E. K. Frequency-specific hippocampal-prefrontal interactions during associative learning. Nat. Neurosci. 18, 576–581 (2015).
Li, M., Long, C. & Yang, L. Hippocampal-prefrontal circuit and disrupted functional connectivity in psychiatric and neurodegenerative disorders. Biomed. Res Int 2015, 810548 (2015).
Ahnaou, A. et al. Emergence of early alterations in network oscillations and functional connectivity in a tau seeding mouse model of Alzheimer’s disease pathology. Sci. Rep. 7, 14189 (2017).
De Simoni, S. et al. Disconnection between the default mode network and medial temporal lobes in post-traumatic amnesia. Brain 139, 3137–3150 (2016).
Kitchigina, V. F. Alterations of coherent theta and gamma network oscillations as an early biomarker of temporal lobe epilepsy and Alzheimer’s disease. Front Integr. Neurosci. 12, 36 (2018).
Hyman, B. T., Van Hoesen, G. W., Damasio, A. R. & Barnes, C. L. Alzheimer’s disease: cell-specific pathology isolates the hippocampal formation. Science 225, 1168–1170 (1984).
Jeong, J. EEG dynamics in patients with Alzheimer’s disease. Clin. Neurophysiol. 115, 1490–1505 (2004).
Herrmann, C. S. & Demiralp, T. Human EEG gamma oscillations in neuropsychiatric disorders. Clin. Neurophysiol. 116, 2719–2733 (2005).
Wang, J. et al. Enhanced gamma activity and cross-frequency interaction of resting-state electroencephalographic oscillations in patients with Alzheimer’s disease. Front. Aging Neurosci. 9, 243 (2017).
van Deursen, J. A., Vuurman, E. F. P. M., Verhey, F. R. J., Van Kranen-Mastenbroek, V. H. J. M. & Riedel, W. J. Increased EEG gamma band activity in Alzheimer’s disease and mild cognitive impairment. J. Neural Transm. (Vienna) 115, 1301–1311 (2008).
Goodman, M. S. et al. Changes in theta but not alpha modulation are associated with impairment in working memory in Alzheimer’s disease and mild cognitive impairment. J. Alzheimers Dis. 68, 1085–1094 (2019).
Musaeus, C. S., Nielsen, M. S., Musaeus, J. S. & Høgh, P. Electroencephalographic cross-frequency coupling as a sign of disease progression in patients with mild cognitive impairment: a pilot study. Front. Neurosci. 14, 790 (2020).
Goodman, M. S. et al. Theta-gamma coupling and working memory in Alzheimer’s dementia and mild cognitive impairment. Front. Aging Neurosci. 10, 101 (2018).
Goutagny, R. et al. Alterations in hippocampal network oscillations and theta-gamma coupling arise before Aβ overproduction in a mouse model of Alzheimer’s disease. Eur. J. Neurosci. 37, 1896–1902 (2013).
Etter, G. et al. Optogenetic gamma stimulation rescues memory impairments in an Alzheimer’s disease mouse model. Nat. Commun. 10, 5322 (2019).
Ittner, A. A., Gladbach, A., Bertz, J., Suh, L. S. & Ittner, L. M. p38 MAP kinase-mediated NMDA receptor-dependent suppression of hippocampal hypersynchronicity in a mouse model of Alzheimer’s disease. Acta Neuropathol. Commun. 2, 149 (2014).
Wirt, R. A. et al. Altered theta rhythm and hippocampal-cortical interactions underlie working memory deficits in a hyperglycemia risk factor model of Alzheimer’s disease. Commun. Biol. 4, 1036 (2021).
Rajji, T. K. Impaired brain plasticity as a potential therapeutic target for treatment and prevention of dementia. Expert Opin. Therapeutic Targets 23, 21–28 (2019).
Liu, R. et al. Intermittent stimulation of the nucleus basalis of Meynert improves working memory in adult monkeys. Curr. Biol. 27, 2640–2646.e4 (2017).
Hamani, C. et al. Memory enhancement induced by hypothalamic/fornix deep brain stimulation. Ann. Neurol. 63, 119–123 (2008).
Laxton, A. W. et al. A phase I trial of deep brain stimulation of memory circuits in Alzheimer’s disease. Ann. Neurol. 68, 521–534 (2010).
Fontaine, D. et al. Symptomatic treatment of memory decline in Alzheimer’s disease by deep brain stimulation: a feasibility study. J. Alzheimers Dis. 34, 315–323 (2013).
Sankar, T. et al. Deep brain stimulation influences brain structure in Alzheimer’s disease. Brain Stimul. 8, 645–654 (2015).
Lozano, A. M. et al. A phase II study of fornix deep brain stimulation in mild Alzheimer’s disease. J. Alzheimers Dis. 54, 777–787 (2016).
Leoutsakos, J.-M. S. et al. Deep brain stimulation targeting the fornix for mild Alzheimer Dementia (the advance trial): a two year follow-up including results of delayed activation. J. Alzheimers Dis. 64, 597–606 (2018).
Shirvalkar, P. R., Rapp, P. R. & Shapiro, M. L. Bidirectional changes to hippocampal theta-gamma comodulation predict memory for recent spatial episodes. Proc. Natl Acad. Sci. USA 107, 7054–7059 (2010).
Sweet, J. A., Eakin, K. C., Munyon, C. N. & Miller, J. P. Improved learning and memory with theta-burst stimulation of the fornix in rat model of traumatic brain injury. Hippocampus 24, 1592–1600 (2014).
Larson, J., Wong, D. & Lynch, G. Patterned stimulation at the theta frequency is optimal for the induction of hippocampal long-term potentiation. Brain Res. 368, 347–350 (1986).
Miller, J. P. et al. Visual-spatial memory may be enhanced with theta burst deep brain stimulation of the fornix: a preliminary investigation with four cases. Brain 138, 1833–1842 (2015).
Halgren, E. & Wilson, C. L. Recall deficits produced by afterdischarges in the human hippocampal formation and amygdala. Electroencephalogr. Clin. Neurophysiol. 61, 375–380 (1985).
Halgren, E., Wilson, C. L. & Stapleton, J. M. Human medial temporal-lobe stimulation disrupts both formation and retrieval of recent memories. Brain Cogn. 4, 287–295 (1985).
Coleshill, S. G. et al. Material-specific recognition memory deficits elicited by unilateral hippocampal electrical stimulation. J. Neurosci. 24, 1612–1616 (2004).
Lacruz, M. E. et al. Single pulse electrical stimulation of the hippocampus is sufficient to impair human episodic memory. Neuroscience 170, 623–632 (2010).
Suthana, N. et al. Memory enhancement and deep-brain stimulation of the entorhinal area. N. Engl. J. Med. 366, 502–510 (2012).
Jacobs, J. et al. Direct electrical stimulation of the human entorhinal region and hippocampus impairs memory. Neuron 92, 983–990 (2016).
Titiz, A. S. et al. Theta-burst microstimulation in the human entorhinal area improves memory specificity. Elife 6, (2017).
Mankin, E. A. et al. Stimulation of the right entorhinal white matter enhances visual memory encoding in humans. Brain Stimul. 14, 131–140 (2021).
Jun, S., Kim, J. S. & Chung, C. K. Direct stimulation of human hippocampus during verbal associative encoding enhances subsequent memory recollection. Front Hum. Neurosci. 13, 23 (2019).
Inman, C. S. et al. Direct electrical stimulation of the amygdala enhances declarative memory in humans. Proc. Natl Acad. Sci. USA 115, 98–103 (2018).
Natu, V. S. et al. Stimulation of the posterior cingulate cortex impairs episodic memory encoding. J. Neurosci. 39, 7173–7182 (2019).
Fell, J. et al. Memory modulation by weak synchronous deep brain stimulation: a pilot study. Brain Stimulation 6, 270–273 (2013).
Hampson, R. E. et al. Developing a hippocampal neural prosthetic to facilitate human memory encoding and recall. J. Neural Eng. 15, 036014 (2018).
Penfield, W. & Perot, P. The brain’s record of auditory and visual experience. a final summary and discussion. Brain 86, 595–696 (1963).
Kucewicz, M. T. et al. Evidence for verbal memory enhancement with electrical brain stimulation in the lateral temporal cortex. Brain 141, 971–978 (2018).
Kucewicz, M. T. et al. Electrical stimulation modulates high γ activity and human memory performance. eNeuro 5, 0369–17.2018 (2018).
Ezzyat, Y. et al. Direct brain stimulation modulates encoding states and memory performance in humans. Curr. Biol. 27, 1251–1258 (2017).
Ezzyat, Y. et al. Closed-loop stimulation of temporal cortex rescues functional networks and improves memory. Nat. Commun. 9, 365 (2018).
Senova, S., Chaillet, A. & Lozano, A. M. Fornical closed-loop stimulation for Alzheimer’s disease. Trends Neurosci. 41, 418–428 (2018).
Alagapan, S. et al. Network-targeted, multi-site direct cortical stimulation enhances working memory by modulating phase lag of low-frequency oscillations. Cell Rep. 29, 2590–2598.e4 (2019).
Berger, T. W. et al. A cortical neural prosthesis for restoring and enhancing memory. J. Neural Eng. 8, 046017 (2011).
Deadwyler, S. A. et al. A cognitive prosthesis for memory facilitation by closed-loop functional ensemble stimulation of hippocampal neurons in primate brain. Exp. Neurol. 287, 452–460 (2017).
Roeder, B. M. et al. Patterned hippocampal stimulation facilitates memory in patients with a history of head impact and/or brain injury. Front Hum. Neurosci. 16, 933401 (2022).
Hayne, R., Meyers, R. & Knott, J. R. Characteristics of electrical activity of human corpus striatum and neighboring structures. J. Neurophysiol. 12, 185–195 (1949).
Jasper, H. H. Electrical signs of epileptic discharge. Electroencephalogr. Clin. Neurophysiol. 1, 11–18 (1949).
Reif, P. S., Strzelczyk, A. & Rosenow, F. The history of invasive EEG evaluation in epilepsy patients. Seizure 41, 191–195 (2016).
Ceyssens, F. et al. Chronic neural recording with probes of subcellular cross-section using 0.06 mm2 dissolving microneedles as insertion device. Sens. Actuators B: Chem. 284, 369–376 (2019).
Chauvière, L. et al. In vivo recording quality of mechanically decoupled floating versus skull-fixed silicon-based neural probes. Front. Neurosci. 13, 464 (2019).
Dutta, B. et al. The Neuropixels probe: A CMOS based integrated microsystems platform for neuroscience and brain-computer interfaces. In 2019 IEEE International Electron Devices Meeting (IEDM) 10.1.1–10.1.4 (IEEE, 2019).
Egert, D. et al. Cellular-scale silicon probes for high-density, precisely localized neurophysiology. J. Neurophysiol. 124, 1578–1587 (2020).
Hong, G. & Lieber, C. M. Novel electrode technologies for neural recordings. Nat. Rev. Neurosci. 20, 330–345 (2019).
Jun, J. J. et al. Fully integrated silicon probes for high-density recording of neural activity. Nature 551, 232–236 (2017).
Klein, L. et al. High-density electrophysiological recordings in macaque using a chronically implanted 128-channel passive silicon probe. J. Neural Eng. 17, 026036 (2020).
Paulk, A. C. et al. Large-scale neural recordings with single neuron resolution using Neuropixels probes in human cortex. Nat. Neurosci. 25, 252–263 (2022).
Shobe, J. L., Claar, L. D., Parhami, S., Bakhurin, K. I. & Masmanidis, S. C. Brain activity mapping at multiple scales with silicon microprobes containing 1,024 electrodes. J. Neurophysiol. 114, 2043–2052 (2015).
Ulyanova, A. V. et al. Multichannel silicon probes for awake hippocampal recordings in large animals. Front. Neurosci. 13, 397 (2019).
Yang, L., Lee, K., Villagracia, J. & Masmanidis, S. C. Open source silicon microprobes for high throughput neural recording. J. Neural Eng. 17, 016036 (2020).
Zhang, S. et al. Real-time simultaneous recording of electrophysiological activities and dopamine overflow in the deep brain nuclei of a non-human primate with Parkinson’s disease using nano-based microelectrode arrays. Microsyst. Nanoeng. 4, 17070 (2018).
Steinmetz, N. A., Koch, C., Harris, K. D. & Carandini, M. Challenges and opportunities for large-scale electrophysiology with Neuropixels probes. Curr. Opin. Neurobiol. 50, 92–100 (2018).
Steinmetz, N. A., Zatka-Haas, P., Carandini, M. & Harris, K. D. Distributed coding of choice, action and engagement across the mouse brain. Nature 576, 266–273 (2019).
Fiáth, R. et al. Long-term recording performance and biocompatibility of chronically implanted cylindrically-shaped, polymer-based neural interfaces. Biomed. Eng./Biomedizinische Tech. 63, 301–315 (2018).
Trepka, E. B., Zhu, S., Xia, R., Chen, X. & Moore, T. Functional interactions among neurons within single columns of macaque V1. Elife 11, e79322 (2022).
Chung, J. E. et al. High-density single-unit human cortical recordings using the Neuropixels probe. Neuron 110, 2409–2421.e3 (2022).
Nakahara, K., Adachi, Y., Osada, T. & Miyashita, Y. Exploring the neural basis of cognition: multi-modal links between human fMRI and macaque neurophysiology. Trends Cogn. Sci. 11, 84–92 (2007).
Hutchison, R. M. & Everling, S. Monkey in the middle: why non-human primates are needed to bridge the gap in resting-state investigations. Front. Neuroanat. 6, 29 (2012).
Rapp, P. R. & Amaral, D. G. Evidence for task-dependent memory dysfunction in the aged monkey. J. Neurosci. 9, 3568–3576 (1989).
Herndon, J. G., Moss, M. B., Rosene, D. L. & Killiany, R. J. Patterns of cognitive decline in aged rhesus monkeys. Behav. Brain Res. 87, 25–34 (1997).
Nagahara, A. H., Bernot, T. & Tuszynski, M. H. Age-related cognitive deficits in rhesus monkeys mirror human deficits on an automated test battery. Neurobiol. Aging 31, 1020–1031 (2010).
Rupniak, N. M., Samson, N. A., Steventon, M. J. & Iversen, S. D. Induction of cognitive impairment by scopolamine and noncholinergic agents in rhesus monkeys. Life Sci. 48, 893–899 (1991).
Buccafusco, J. J. et al. The scopolamine-reversal paradigm in rats and monkeys: the importance of computer-assisted operant-conditioning memory tasks for screening drug candidates. Psychopharmacology 199, 481–494 (2008).
Zhou, X. et al. Cholinergic modulation of working memory activity in primate prefrontal cortex. J. Neurophysiol. 106, 2180–2188 (2011).
Heuer, E., Rosen, R. F., Cintron, A. & Walker, L. C. Nonhuman primate models of Alzheimer-like cerebral proteopathy. Curr. Pharm. Des. 18, 1159–1169 (2012).
Finch, C. E. & Austad, S. N. Commentary: is Alzheimer’s disease uniquely human? Neurobiol. Aging 36, 553–555 (2015).
Bons, N., Rieger, F., Prudhomme, D., Fisher, A. & Krause, K.-H. Microcebus murinus: a useful primate model for human cerebral aging and Alzheimer’s disease? Genes Brain Behav. 5, 120–130 (2006).
Cramer, P. E. et al. Aging African green monkeys manifest transcriptional, pathological, and cognitive hallmarks of human Alzheimer’s disease. Neurobiol. Aging 64, 92–106 (2018).
Latimer, C. S. et al. A nonhuman primate model of early Alzheimer’s disease pathologic change: Implications for disease pathogenesis. Alzheimers Dement. 15, 93–105 (2019).
Paspalas, C. D. et al. The aged rhesus macaque manifests Braak stage III/IV Alzheimer’s-like pathology. Alzheimers Dement. 14, 680–691 (2018).
Arnsten, A. F. T. et al. Alzheimer’s-like pathology in aging rhesus macaques: unique opportunity to study the etiology and treatment of Alzheimer’s disease. Proc. Natl Acad. Sci. USA 116, 26230–26238 (2019).
Oikawa, N., Kimura, N. & Yanagisawa, K. Alzheimer-type tau pathology in advanced aged nonhuman primate brains harboring substantial amyloid deposition. Brain Res. 1315, 137–149 (2010).
Heo, J.-H. et al. Spatial distribution of glucose hypometabolism induced by intracerebroventricular streptozotocin in monkeys. J. Alzheimers Dis. 25, 517–523 (2011).
Park, S.-J. et al. Quantitative expression analysis of APP pathway and tau phosphorylation-related genes in the ICV STZ-induced non-human primate model of sporadic Alzheimer’s disease. Int J. Mol. Sci. 16, 2386–2402 (2015).
Forny-Germano, L. et al. Alzheimer’s disease-like pathology induced by amyloid-β oligomers in nonhuman primates. J. Neurosci. 34, 13629–13643 (2014).
Beckman, D. et al. A novel tau-based rhesus monkey model of Alzheimer’s pathogenesis. Alzheimers Dement. 17, 933–945 (2021).
Sato, K. et al. A non-human primate model of familial Alzheimer’s disease. https://doi.org/10.1101/2020.08.24.264259 (2020).
Rizzo, S. J. S., Homanics, G. E., Park, J. E., Silva, A. C. & Strick, P. L. Establishing the marmoset as a non-human primate model of Alzheimer’s disease. Alzheimer’s Dement. 17, e049952 (2021).
Rattay, F. et al. Mechanisms of electrical stimulation with neural prostheses. Neuromodulation 6, 42–56 (2003).
Bingham, C. S. et al. Model-based analysis of electrode placement and pulse amplitude for hippocampal stimulation. IEEE Trans. Biomed. Eng. 65, 2278–2289 (2018).
Cutsuridis, V. Memory prosthesis: is it time for a deep neuromimetic computing approach? Front. Neurosci. 13, 667 (2019).
Onslow, A. C. E., Jones, M. W. & Bogacz, R. A canonical circuit for generating phase-amplitude coupling. PLoS ONE 9, e102591 (2014).
Segneri, M., Bi, H., Olmi, S. & Torcini, A. Theta-nested gamma oscillations in next generation neural mass models. Front. Comput. Neurosci. 14, 47 (2020).
Aussel, A., Buhry, L., Tyvaert, L. & Ranta, R. A detailed anatomical and mathematical model of the hippocampal formation for the generation of sharp-wave ripples and theta-nested gamma oscillations. J. Comput. Neurosci. 45, 207–221 (2018).
Lundqvist, M., Rehn, M., Djurfeldt, M. & Lansner, A. Attractor dynamics in a modular network model of neocortex. Network 17, 253–276 (2006).
Herman, P. A., Lundqvist, M. & Lansner, A. Nested theta to gamma oscillations and precise spatiotemporal firing during memory retrieval in a simulated attractor network. Brain Res. 1536, 68–87 (2013).
Cutsuridis, V., Cobb, S. & Graham, B. P. Encoding and retrieval in a model of the hippocampal CA1 microcircuit. Hippocampus 20, 423–446 (2010).
Menschik, E. D. & Finkel, L. H. Neuromodulatory control of hippocampal function: towards a model of Alzheimer’s disease. Artif. Intell. Med. 13, 99–121 (1998).
Rubin, J. E. & Terman, D. High frequency stimulation of the subthalamic nucleus eliminates pathological thalamic rhythmicity in a computational model. J. Comput. Neurosci. 16, 211–235 (2004).
Pirini, M., Rocchi, L., Sensi, M. & Chiari, L. A computational modelling approach to investigate different targets in deep brain stimulation for Parkinson’s disease. J. Comput. Neurosci. 26, 91–107 (2009).
Kumar, A., Cardanobile, S., Rotter, S. & Aertsen, A. The role of inhibition in generating and controlling Parkinson’s disease oscillations in the Basal Ganglia. Front. Syst. Neurosci. 5, 86 (2011).
Mina, F., Benquet, P., Pasnicu, A., Biraben, A. & Wendling, F. Modulation of epileptic activity by deep brain stimulation: a model-based study of frequency-dependent effects. Front. Comput. Neurosci. 7, 94 (2013).
McNeal, D. R. Analysis of a model for excitation of myelinated nerve. IEEE Trans. Biomed. Eng. 23, 329–337 (1976).
Rattay, F. Analysis of models for external stimulation of axons. IEEE Trans. Biomed. Eng. BME- 33, 974–977 (1986).
Rattay, F. The basic mechanism for the electrical stimulation of the nervous system. Neuroscience 89, 335–346 (1999).
Grill, W. M. Modeling the effects of electric fields on nerve fibers: influence of tissue electrical properties. IEEE Trans. Biomed. Eng. 46, 918–928 (1999).
Warman, E. N., Grill, W. M. & Durand, D. Modeling the effects of electric fields on nerve fibers: determination of excitation thresholds. IEEE Trans. Biomed. Eng. 39, 1244–1254 (1992).
McIntyre, C. C., Grill, W. M., Sherman, D. L. & Thakor, N. V. Cellular effects of deep brain stimulation: model-based analysis of activation and inhibition. J. Neurophysiol. 91, 1457–1469 (2004).
Hendrickson, P. J., Yu, G. J., Song, D. & Berger, T. W. A million-plus neuron model of the hippocampal dentate gyrus: critical role for topography in determining spatiotemporal network dynamics. IEEE Trans. Biomed. Eng. 63, 199–209 (2016).
Acknowledgements
This work was supported by the Bordeaux Neurocampus junior chair programme co-funded by the Région Nouvelle-Aquitaine and the University of Bordeaux Initiative of Excellence (IdEx) (F.B.W., Neurocampus junior chair), and the European Union’s Horizon Europe research and innovation programme under European Research Council (ERC) Starting Grant agreement No. 101040391 (F.B.W., MEMOPROSTHETICS). Views and opinions expressed are however those of the author(s) only and do not necessarily reflect those of the European Union or the European Research Council. Neither the European Union nor the granting authority can be held responsible for them.
Author information
Authors and Affiliations
Contributions
All authors reviewed the literature and wrote individual sections of the manuscript. F.W. integrated the different parts together. All authors contributed to the figures/table and edited the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Communications Biology thanks Vassilis Cutsuridis, Amir Alimohammad and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Primary Handling Editor: Manuel Breuer.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Gupta, A., Vardalakis, N. & Wagner, F.B. Neuroprosthetics: from sensorimotor to cognitive disorders. Commun Biol 6, 14 (2023). https://doi.org/10.1038/s42003-022-04390-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s42003-022-04390-w
This article is cited by
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
