

Abstract
Forest management and disturbances are among the main drivers of changes in forest dynamics in temperate ecosystems. To promote and maintain forest multifunctionality and species persistence in the landscape, it is critical that the interactions between these factors and forest biodiversity are disentangled. Still, the relationships between disturbances and forest management are poorly understood and may hinder an adequate planning of management and conservation actions in these forests. Here we address this issue via a coupled ecological-economic modeling system under different climate change scenarios. We employed data from a large-scale field-based research in southwestern Germany, in combination with a climate-sensitive forest growth model. Thereby, we quantified changes in multiple biodiversity indicators (including richness of birds, bats and flying insect orders) and tree microhabitats (TreMs) in the face of disturbance and management interventions. Our results show that windstorms may cause economic damage in managed forests, but at the same time improve biodiversity indicators in these areas. Salvage logging, however, may dampen these positive impacts for the majority of indicators considered. Moreover, management solutions targeting at wind risk mitigation may be detrimental to some taxa (e.g. forest birds) but still performed better than the business-as-usual management, in terms of the biodiversity indicators applied. We conclude that considering disturbance impacts on forest dynamics will be key to maintain the sustainability in the use of forest resources and support species persistence in temperate forest landscapes.
Export citation and abstract BibTeX RIS

Original content from this work may be used under the terms of the Creative Commons Attribution 4.0 license. Any further distribution of this work must maintain attribution to the author(s) and the title of the work, journal citation and DOI.
1. Introduction
Biodiversity loss remains a central issue in the management of natural resources. Human activities have led to substantial modifications in habitat quality and availability, affecting extinction rates worldwide (Ceballos et al 2015). This biodiversity decline has triggered the implementation of conservation programs, which cascaded to forest management practices. For example, the expansion of protected forest areas and implementation of retention forestry are being adopted in central Europe to mitigate negative impacts of management on forest biota (Gustafsson et al 2020). These conservation policies, however, often disregard the interactions between forest management, climate change and natural disturbances, and their influence on biodiversity. This gap needs to be urgently addressed, to identify adaptive management solutions and build resilient landscapes in the future. In central Europe, the increase in disturbance activity due to climate change will likely be one of the main drivers of future changes in forest dynamics (Seidl et al 2017). In this context, sound conservation policies need to include the interactions between natural disturbances and forest biodiversity.
Windstorms are the main disturbance agents in European forests (Hanewinkel et al 2011). For example, the windstorm Lothar caused a damage of almost 200 million m3 of wood in Central Europe (Hanewinkel et al 2011). Forest management and natural disturbances also strongly affect resource availability for forest taxa and socio-economic aspects of forest management, due to their influence on various ecosystem goods and services, such as climate regulation, protective function, wood production and forest profitability (Thom and Seidl 2016, Sebald et al 2019). The harmonization of economic goals and the maintenance of habitats for multiple forest taxa require adequate planning tools, which resolve potential trade-offs and produces the highest possible benefits under climate and disturbance uncertainty. These tools may provide a set of optimal benchmark solutions that can be adapted by decision-makers to local contexts, according to their preferences for different ecosystem services.
The impacts and interactions between climate, disturbances and forest taxa are, however, nontrivial and multifaceted. While the increase in deadwood amounts related to wind disturbances may be beneficial for some taxa, the loss of old growth habitat types is likely to be detrimental to other organisms. For example, the creation of gaps on the forest cover promotes early successional stages in affected areas and modifies structural diversity (Larue et al 2019; Swanson et al 2011). Such habitats are scarce in continuous cover forestry systems of central Europe. Hilmers et al (2018) investigated the responses of several taxonomic groups to different forest successional stages in Europe. The authors report a U-shaped temporal pattern on the response of producers, first-order consumers and saprotrophs. Conversely, fungi displayed an inverse U-shaped response. Hence, wind disturbances may act synergistically or trade-off with different forest taxa due to the novel stand structures created, including lying deadwood, snags and modifications in carbon stocks of forest stands.
The effects of windstorms on climate mitigation potential and carbon storage have been addressed for temperate forests in Europe (e.g. Seidl et al 2014, Reyer et al 2017). Numerous studies have investigated the economic impacts of disturbances, including windstorms, drought, fires and pest outbreaks on the economic profitability of forest management (Brunette et al 2015, Müller et al 2019, Paul et al 2019). However, the interactions between disturbance events and forest biodiversity are understudied (e.g. Thom et al 2017). Here we aimed to disentangle the connections between disturbance events, forest management and forest biodiversity, and to provide insights on the consequences of forest management decisions to multiple biodiversity indicators and forest profitability. We investigate the following research questions:
- What are the effects of stand structure on multiple biodiversity indicators?
- What are the interactions between windstorms, salvage logging and biodiversity indicators?
- What are the economic impacts of wind disturbances and the optimal management strategies to cope with risk and simultaneously maintain habitat for multiple taxa?
To answer the research questions, we combined the responses of the forest growth model Sibyla (Fabrika 2005) with the empirical wind disturbance model developed by Albrecht et al (2012). We carried out a multi-taxon analysis, assessing species richness in relation to novel forest structural composition resulting from windstorm events, using a Monte Carlo simulation approach. We assessed the impacts of disturbance events on the economic output of forest management via the net present value (NPV) and changes in biodiversity indicators based on the richness of birds, bats and flying insects, and additionally the richness of tree microhabitats (TreMs). Finally, we identified robust management solutions (portfolios) to balance economic and conservation objectives in a temperate forest landscape.
2. Material and methods
To identify the impacts of wind disturbances and forest management on biodiversity indicators and forest profitability, we applied a coupled ecological-economic approach. Initially, we established the responses of biodiversity indicators, including the richness of forest birds, bats, flying insect orders (referred to as insect richness in the remainder of the manuscript) and TreMs, to environmental (temperature and altitude) and forest stand's structural parameters via generalized linear mixed effects model (GLMM) and applying Bayesian inference. In a second step, we simulated forest dynamics under alternative management options and climate change scenarios using a climate sensitive forest growth model, capable to endogenously mimic the effects of wind disturbance events. To assess the impacts of windstorms on the biodiversity indicators, we quantified changes in the stand structural parameters and applied the models developed in the first step to predict changes in richness. Finally, we used a robust optimization framework to find management solutions, referred to as management portfolios in the remainder of the manuscript. These solutions aimed to balance biodiversity conservation and forest profitability under four alternative preference scenarios: baseline management (business-as-usual), risk mitigation (minimization of economic damage caused by windstorms), balanced solution (maximization of biodiversity indicators while maintaining forest profitability) and biodiversity maximization (maximization of biodiversity indicators and no requirements on forest profitability).
2.1. Data
We employed data from a field-based research composed by 109 plots in the Southern Black Forest- Germany, covering a forest connectivity (<50%, 50%–75% and >75%) and a forest structure (<5 habitat trees/ha, 5–15 habitat trees/ha and >15 habitat trees/ha) gradient. The inventory recorded tree identity and diameter at breast height (DBH) of all trees, the height of a representative 7% of the trees and the volume of standing and lying deadwood. This dataset was used initialize the forest growth model. The richness of forest birds, orders of flying insects, tree microhabitats and bat activity were assessed in the same plots. Birds seen or heard in the plots were recorded in three surveys during the spring of 2017. Bats were recorded via ultrasonic sound recorders and flying insects were collected with modified window traps in the same period (see Knuff et al 2019 for details). Finally, TreMs were evaluated and recorded based on a detailed catalogue for TreMs classification by Larrieu et al 2018 (for details see supplementary 1 (available online at http://stacks.iop.org/ERL/15/0940a3/mmedia) and Storch et al 2020).
2.2. Forest simulations
The forest simulations were carried out using the semi-empirical growth model Sibyla (Fabrika 2005), in a 50-year simulation period (2016–2065). Sibyla is a distance-dependent individual tree model that simulates forest stand dynamics at tree, stand and landscape level. The model includes the description of growth, mortality and detailed management options. Forest growth is dependent on multiple climatic variables (atmospheric CO2- and N2O- concentration, average temperature, temperature range, precipitation) that inform a potential growth modifier. This is subsequently combined with a tree competition index and a tree vitality index to yield the actual increment.
We simulated four thinning intensities, applying thinning from above (see supplementary 2), namely the business-as-usual (BAU) intensity (calculated as the volume removal in percentage of the standing stock), increase or decrease in thinning intensity and no thinning. These thinning schemes were tailored to the main species in the study region, according to the German National Forest Inventory data (https://bwi.info). Each thinning intensity, referred to as management regimes in the remainder of the manuscript, was driven by three climate change scenarios given by the HadGEM2-ES global climate model, forced by the representative concentration pathways 4.5, 6.0 and 8.5 (https://www.isimip.org).
We retrieved from the forest simulations (for each thinning intensity and climate change scenario) the list of trees in the plots at each time step of the simulation period. These information included tree growth, biomass, the list of trees remaining in the plot, trees removed by management, dead trees due to natural mortality and dead trees due to windstorms. Moreover, the list of trees also included information regarding the species identity, DBH, height, total volume and volume per wood assortment class.
We included deadwood dynamics in the simulations via a post-processing routine, considering deadwood input from natural mortality and windstorms. Both sources of deadwood were assigned to the lying deadwood and snag pools. For deadwood arising from natural mortality, we differentiated between standing and lying deadwood, according to the proportions retrieved from the German National Forest Inventory in the past 10 years (https://bwi.info). For deadwood created by windstorms, a critical slenderness coefficient was applied (height/DBH ratio) as a decision criterion for stem breakage or uprooting. Uprooted trees were allocated to the lying deadwood pool, whereas broken trees were allocated to the snag pool (standing portion of the stem) and lying deadwood pool (broken portion of the stem). Finally, we computed at each period the actual lying and standing deadwood using the decay models developed by Meyer et al (2009) (details in supplementary 2).
2.3. Wind disturbance
We used the wind damage model proposed by Albrecht et al (2012) to estimate the probability of total stand damage (equation (1)), using the species and dominant height as predictors. We set the other predictors in the original model to its mean value and adding the respective constant to the intercept of the model. Subsequently, we randomly assigned total damage events (equal to 75% of the standing stock) across the simulation plots based on the calculated probability. We calculated the mortality of partially damage stands using equation (2), for each plot and species. Finally, the wind damage probability was included in the forest growth simulations using a Monte Carlo approach endogenous to the growth model Sibyla (considering ten repetitions), for each climate scenario and thinning intensity.
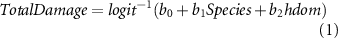
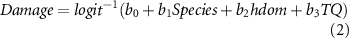
Where:  : probability of total damage;
: probability of total damage;  : amount of damage (in %) for partially damaged stands;
: amount of damage (in %) for partially damaged stands;  : predictor's coefficients retrieved from Albrecht et al (2012);
: predictor's coefficients retrieved from Albrecht et al (2012);  : tree species group;
: tree species group;  : dominant height (mean height of 100 largest trees in a stand);
: dominant height (mean height of 100 largest trees in a stand);  : thinning quotient of past 10 years.
: thinning quotient of past 10 years.
2.4. Biodiversity indicators
We fitted empirical models for each biodiversity indicator against stand predictors affected by forest management and windstorm events, as well as environmental proxies (ranges are given in supplementary 1). We used the data collected in the field surveys to regress species richness of birds, bats and insect orders, as well as the richness of TreMs, against the stand height, basal area, density, deadwood volume, number of snags, share of conifers in the stand (in terms of the standing volume), altitude and mean temperature during the vegetation season. We applied GLMMs, with the plots as random effects, due to potential spatial autocorrelation among them. All indicators of stand's structural complexity were assumed to potentially influence all biodiversity indicators, either through the direct availability of resource and roosting opportunity or by influencing related forest taxa.
To calibrate the models, we applied Bayesian inference, employing the R package brms (Bürkner 2017). We used a Poisson likelihood to estimate the richness of all indicators (equation (3)) and uniform priors in the interval [−10, 10] for all predictors, which were normalized to avoid scaling issues. We ran three chains for each model with 20 000 iterations and discarding the initial 10 000. Finally, environmental parameters were included in the analysis via the average temperature in the vegetation season (May to September) and the Altitude. The former is important to manifest the effects of climate change on species richness and the latter to express the variation in forest types and management systems applied in the study region. To predict the biodiversity indicators, we tracked the stand structural parameters (used as predictors) at each time step from the forest simulations described in section 2.2. Moreover, we assumed no change in species composition, i.e. no immigration or emigration beyond the study site, as well as no change in the tree species composition in forest management (which would affect TreMs).
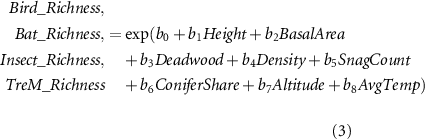
Where:  : model's coefficients;
: model's coefficients;  : tree microhabitat richness (N/ha);
: tree microhabitat richness (N/ha);  : bird richness(N/ha);
: bird richness(N/ha);  : approximated bat richness(N/ha);
: approximated bat richness(N/ha);  : insect order richness(N/ha);
: insect order richness(N/ha);  : mean plot height (m);
: mean plot height (m);  : plot basal area (m2/ha);
: plot basal area (m2/ha);  : lying deadwood volume (m3/ha);
: lying deadwood volume (m3/ha);  : plot density (N/ha);
: plot density (N/ha);  : number of snags (N/ha);
: number of snags (N/ha);  : volume share of conifers (%);
: volume share of conifers (%);  : plot altitude (m a.s.l.);
: plot altitude (m a.s.l.);  : mean temperature in the vegetation season in 2017 and 2018 (°C).
: mean temperature in the vegetation season in 2017 and 2018 (°C).
2.5. Economic impacts and robust management solutions
Harmonizing wood production with the provision of other ecosystem services remains an important challenge in the management of forest resources (Piponiot et al 2019). To address this issue and account for potential different preferences related to forest profitability and biodiversity conservation goals of multiple agents or stakeholders, we developed alternative preference scenarios (table 1). Here we adopted a normative approach to the forest planning problem and aimed to derive alternative management responses that optimally balance economic and conservation objectives. These solutions may serve as a benchmark for decision-makers, which can adapt the framework proposed to local contexts and individual preferences.
Table 1. Preference scenarios used to build the optimization models.
| Preference scenario | Code | Objective function | Description |
|---|---|---|---|
| A | Baseline | Maximize NPV | Baseline solution for the strategic forest planning problem, aiming to maximize forest profitability (NPV), disregarding management impacts on biodiversity and windstorm risk. |
| B | Risk mitigation | Maximize worst-case NPV | Solutions that safeguard profitability in the face of windstorm risk, economic uncertainty and climate change. The model had the aim to maximize profitability in the worst-case scenarios of the NPV distribution, thus mitigating the impacts of windstorms and other sources of uncertainty. |
| C | Balanced | Maximize biodiversity indicators | Solutions that promote forest biodiversity under windstorm risk and climate change, while maintaining the economic feasibility of forest management (enforcing a positive NPV in the worst-case scenarios). |
| D | Maximize biodiversity | Maximize biodiversity indicators | Solutions that promote forest biodiversity under windstorm risk and climate change with no constraints on forest profitability. |
To find these optimal responses in each preference scenario, we employed robust optimization models that simultaneously take into account economic and ecological indicators of forest management under windstorm risk, climate change and economic parameters uncertainty (interest rate and wood price). To this end, we applied a similar approach to Augustynczik et al (2018) and used a Bernstein approximation scheme to construct Bernstein uncertainty sets (Ben-Tal et al 2009). This is a more general form of ellipsoidal uncertainty sets, with the advantage of offering a same level of protection with reduced conservativeness. Thereby, we developed robust counterparts of the planning problems, finding solutions that guaranteed the performance of the objectives in worst-case scenarios, in the face of parameter fluctuations (details in supplementary 3).
For the economic analysis, we computed the NPV of forest management in the 50-years simulation period (details in supplementary 4). We considered the standing stock at the beginning of the simulation period as the initial investment and the returns were given by discounted thinning revenues and the value of the standing stock at the end of the simulation period. Based on the economic output of forest management and the corresponding impact on the biodiversity indicators considered here, we defined the optimal management portfolio considering four preference scenarios (table 1).
Uncertainty related to climate change (3 RCPs), windstorm risk (30 realizations), economic parameters (Monte Carlo draw of interest rate and wood prices) and on the estimated species richness (via a posterior sampling) were included in the optimization models for policies B to D. For the uncertainty in species and TreM richness, we sampled parameter values for all predictors (equation (3)) from their posterior distribution considering one standard deviation from the mean values. For the interest rate uncertainty, we sampled uniformly from the interval [0.005, 0.01] and for wood price we applied a variation of up to plus and minus 20% of the current price for each assortment class and species. Furthermore, we created Compromise Programming models for scenarios C and D, to balance the various biodiversity indicators (details in supplementary 3 and supplementary 5).
3. Results
3.1. Biodiversity indicators
The richness of forest birds was most responsive to environmental parameters, namely the average temperature during vegetation season and altitude. Conversely, richness of bats and TreMs were more responsive to stand structure and thus to forest management interventions (figure 1). Finally, moderate effects of both environment and stand structure were observed for insect order richness, with altitude, basal area and stand height having the strongest effects on this indicator.
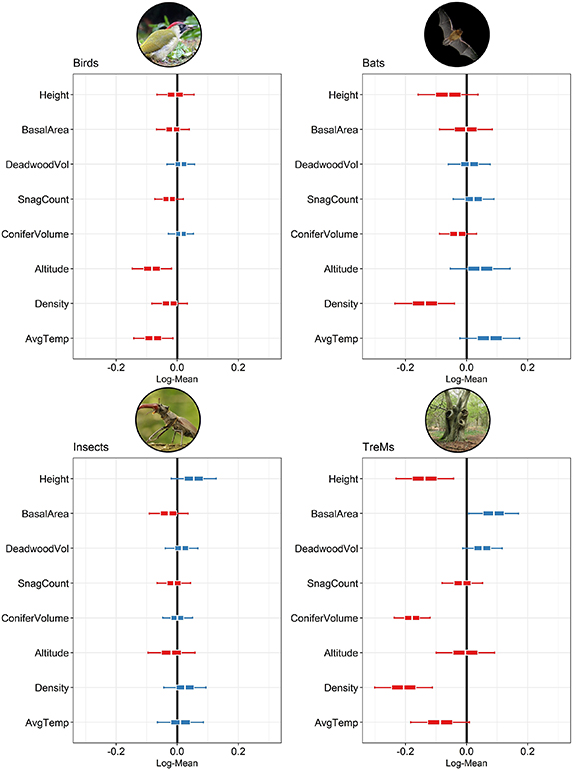
Figure 1. Posterior distribution of models' predictors (plot height, basal area, lying deadwood volume, number of snags, conifer volume share, altitude, density and average temperature in the vegetation season respectively). The red color indicates a negative correlation, whereas blue color indicates positive correlation with the mean parameter value.
Download figure:
Standard image High-resolution imageThe increase in average temperature during the vegetation season positively affected the richness of bats and insect orders, while an increase in this parameter caused a reduction in the richness of bird species and TreMs. A reduced richness of birds and insects was recorded in higher altitudes, whereas bats showed the opposite trend. The relationships between plot altitude and species richness might be mediated by the different species composition and climatic parameters, e.g. an increased share of conifers and lower stand height.
Tree density and plot height were important parameters for the evaluation of bat richness, which improved in sparse stands. Conifer-dominated stands displayed lower bat richness, whereas lying deadwood volume positively affected this taxon. TreMs responded positively to stands with higher basal area and lower height, accompanied by a negative impact of stand density, i.e. TreM di versity was increased in stands with larger trees. Conversely, bird richness was weakly affected by management parameters.
We highlight that the direction of the biodiversity indicators' responses to the structural parameters of forest stands showed a considerable degree of uncertainty. The posterior distributions for several structural characteristics exhibited positive probability density over positive and negative values. Additionally, the explained variance of the fitted models ranged from 0.19 for birds to 0.48 for TreMs (for details on the models' diagnostics see supplementary 1).
3.2. Disturbance-biodiversity interactions
All biodiversity indicators analyzed in this study showed a positive response to the occurrence of wind disturbances (figure 2). The richness of forest birds increased up to 4% due to the creation of deadwood and reduction in basal area, compared to the baseline case (no disturbances). The occurrence of disturbances had also a positive impact on the predicted richness of insects and bats (5% and 7% increase in richness at the end of the planning horizon, respectively), due to the reduction in density, basal area and increase in deadwood amounts. The results also indicate that these patterns may be transient for some biodiversity indicators, due to the decay of deadwood over time. Increased deadwood volume and a reduction in the basal area benefited insect order richness, increasing this biodiversity indicator in damaged stands. These patterns were less pronounced in managed plots, since windstorm events led to lower amounts of deadwood and snag creation in these scenarios.
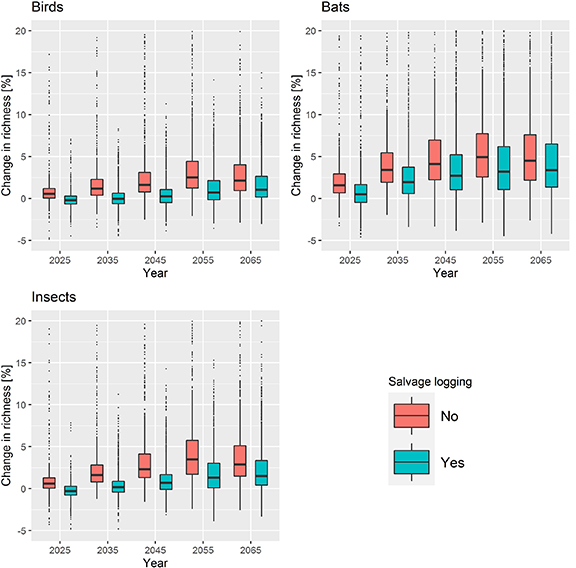
Figure 2. Impacts of windstorm damage and salvage logging on the richness of forest taxa considering the no management strategy, in comparison with the baseline management. The figure shows only the plots and simulation runs where the windstorm damage was at least 15 m3 ha−1 during the simulation period and considering no management interventions.
Download figure:
Standard image High-resolution imageManagement had important implications to windstorm damage and economic outcomes. Increased thinning intensity led, in general, to lower economic damage (details in supplementary 5). Salvage logging buffered the impacts of disturbances, bringing the biodiversity indicators closer to the baseline scenario (0% change in richness) (figure 2). For bat richness, however, the positive influence of disturbances were largely maintained as basal area and density, important parameters for this indicator (see figure 1), were unaffected by salvage logging. Moreover, revenues generated by this management intervention enabled a marginal recovery of economic value from windfelled trees (108 to 2803 EUR ha−1, depending on the management scenario).
3.3. Optimal management strategies under windstorm risk and climate change
The application of different preference scenarios resulted in a major divergence in the optimal portfolios obtained. In general, scenarios aiming at forest profitability (A and B) predominantly applied increased and BAU thinning intensities, whereas the opposite pattern occurred in biodiversity-oriented scenarios (C and D), corresponding to a 33%, 36%, 30% and 21% removal of the standing stock in each period for preference scenarios A to D, respectively. Moreover, a strong management diversification was promoted in scenarios including risk, especially in the balanced solution (C) (figure 3).
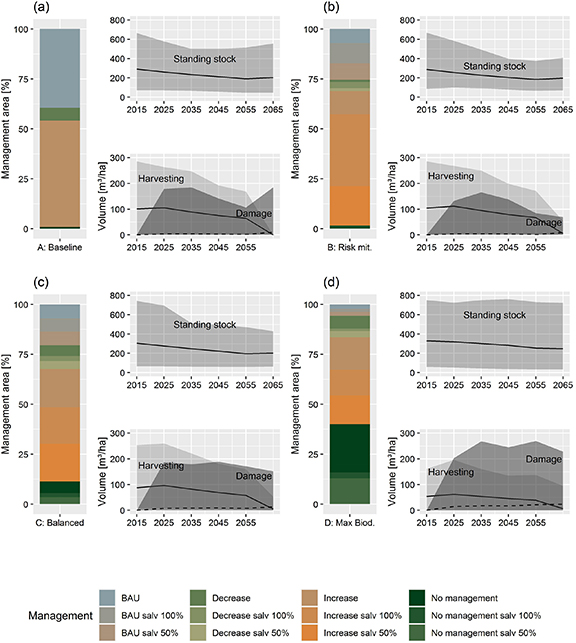
Figure 3. Optimal management portfolios for the preference scenarios A to D and respective development of the standing stock, harvesting volume (solid line) and windstorm damage (dashed line), including the averages and the range of the plots as the shaded areas. Panel 3(a) shows the results for the baseline scenario, 3(b) for the risk mitigation, 3(c) for the balanced solution and 3(d) for biodiversity maximization. BAU stands for business-as-usual thinning intensity, salv 100% refers to full salvaging of windfelled trees and salv 50% to partial salvaging of windfelled trees.
Download figure:
Standard image High-resolution imageApart from the impacts on forest biodiversity, the application of different management portfolios led to a variation in forest profitability under windstorm risk. In the face of uncertain conditions, preference scenario (B) (risk mitigation) outperformed the baseline preference (A), with higher mean NPV (2000 and 1680 EUR ha−1 EUR/ha, respectively) and VaR (1642 and 985 EUR ha−1, respectively). When windstorm damage was taken into account, a reduction in standing stock occurred during the simulation period for scenarios focused on profitability, especially during the initial periods of the planning horizon (right panels in figure 3). This was accompanied by an increase in the harvesting volume, reducing the exposure of these plots to windstorm damage.
The biodiversity indicators were also sensitive to the optimal portfolios applied, i.e. the optimal solutions of preference scenarios (A) to (D) (figure 4). Bird richness decreased up to 8%, depending on the preference scenario. Increases in bat richness ranged from 17%–20% and up to 5% for insects. TreM richness reduced up to 6% in scenarios (A) and (B). As expected, the balanced (C) and biodiversity maximization preference scenarios (D) showed the best outcomes for all indicators. Additionally, the risk mitigation scenario (B) displayed slightly better biodiversity indicator values than the baseline.

{kind=link}
{kind=link}
{kind=link}
Figure 4. Changes in richness along the simulation period [%] of different forest taxa and tree microhabitats (TreMs), considering different preference scenarios: A: baseline, B: risk mitigation, C: balanced and D: maximize biodiversity.
Download figure:
Standard image High-resolution image{kind=link}
4. Discussion
4.1. Biodiversity responses to stand structure and climate drivers
The biodiversity indicators investigated in this study displayed opposing responses to several stand structural parameters and environmental drivers, with forest bats and insects benefitting from higher temperatures, whereas bird richness was negatively correlated with this parameter. Similarly, disturbance events benefitted indicators with positive response to decreased basal area and increased deadwood availability (e.g. birds and bats). Thom et al (2017) also reported diverging responses from forest taxa to climate change impacts on a temperate forest landscape in Austria. A positive influence of climate change to the richness of ground vegetation, Hemiptera, Hymenoptera, Mollusca, Symphyta and Syrphidae was observed. Negative responses, in terms of species richness, were detected for Araneae, Carabidae and saproxylic beetles. In agreement with this study, the authors found a positive influence of disturbance events on the richness of all taxa considered.
Our results show mixed effects of climate change on the indicators studied. Rebelo et al (2010) also report increasing species richness of bats in temperate European forests in the period 2050–2060 using species distribution models. However, a reduction towards the end of the century in more extreme climate scenarios can be expected. Birds may also benefit from increased temperatures, given their ecological adaptability (Stephens et al 2016). Bale et al (2002) and Robinet and Roques (2010) indicate that increased temperatures in temperate regions may benefit some insect groups, resulting in more generations within a year, due to faster development times. Positive responses of bat and TreM richness can be expected after windstorm events, due to the creation of gaps (increasing foraging opportunity for forest bats) and microhabitats related to tree damage (e.g. broken braches and crowns) (Loeb and O'Keefe 2011; Thorn et al 2016).
4.2. Economic impacts of windstorm events and salvage logging
Salvage logging marginally counterbalanced the economic losses caused by windstorms. Apart from the recovery of economic value, salvage logging has the goal of mitigation bark beetle outbreaks after wind disturbances (Leverkus et al 2018). Such outbreaks are likely to become more frequent under climate change, due to the positive impacts of increased summer temperatures on the development rates of insects (e.g. Seidl and Rammer 2017).
The application of salvage logging remains controversial, however, due to insufficient evidence on the effectivity of this management action to control bark beetle outbreaks and the negative effects on biodiversity (Dobor, et al., 2020). Salvage logging dampened the positive impact of disturbances on deadwood-dependent biodiversity indicators in our analysis. Lindenmayer and Noss (2006) identify this trade-off between economic goals and biodiversity generated by salvage logging and indicate that this practice leads to alterations in the structural complexity and removal of legacies, negatively affecting the taxa associated with these structures. Thus, salvage logging must be conducted in a sensible manner, optimally planning the allocation and intensity of wood removals.
4.3. Robust management solutions and risk mitigation effect
The inclusion of windstorm risk and economic uncertainty in the optimization model required a strong management diversification (in terms of thinning intensities applied). Such patterns are ubiquitous in the literature, since the risk related to wood prices and disturbances can be thereby diluted (e.g. Paul et al 2019, Knoke et al 2020a). We considered solely the modification of thinning intensity to cope with these risks in the optimization model. Forests in the study area are majorly managed under a continuous-cover system, and in the time period considered in this study, thinning intensity is a determinant factor for forest profitability and risk exposure. Nevertheless, for other management systems and longer planning horizons, further options are available to support forest resilience. For example, the promotion of mixed stands and modification of rotation lengths may reduce vulnerability and exposure to biotic and abiotic risks (Díaz-Yáñez, et al 2019; Neuner et al 2015, Knoke et al 2020a). Such options may be seamlessly implemented in the framework proposed here, e.g. by creating additional management strategies as combinations of thinning schemes, rotation lengths and species composition (e.g. Augustynczik et al 2020).
Typically, the consideration of windstorm risk in forest planning problems demands an increase in the share of early harvestings and reduction of the standing stock of forest stands (e.g. Amacher, et al 2009; Augustynczik et al 2018). This modification of stand structure may cascade to the availability of habitats for different forest taxa, raising concerns on the impact of such management interventions on forest biodiversity. The results obtained in this study indicate that the risk-mitigation scenario led to poorer outcomes, in terms of species richness, compared to the biodiversity maximization or balanced scenarios. Nevertheless, it still performed better than the baseline management for the mature continuous-cover systems studied here.
We used the NPV as an economic decision criterion in the optimization models and considered a range of different preference scenarios to describe agents' priorities in the management of forest resources (scenarios (A) to (D)). Hence, our results may provide benchmark solutions for decision-makers, which can adapt these to local contexts. It is important to consider, however, that the NPV maximization assumes risk neutrality or a marginal project in the agents' portfolio, and different streams of revenues may generate different utility levels (see Knoke et al 2020b). Furthermore, the optimization model developed here may deviate from individual preferences and attitudes towards conservation and profitability goals. Although it is impractical to derive a single solution that is optimal for all preferences and stakeholders simultaneously, such standard solutions may shed light on the optimal response under various preferences combinations and provide general management guidelines. We highlight that the development of richer decision models may provide additional insights on optimal forest management strategies, e.g. considering utility maximization models tailored to local preferences, welfare maximization models and the inclusion of decision-making biases in the decision process (e.g. Dhami 2016, Augustynczik et al 2020, Knoke et al 2020b).
Furthermore, additional robustness metrics may be applied to balance different management goals and biodiversity indicators, and warrant attention in future studies. For example, Knoke et al (2020b) proposes a framework for multiple criteria robust optimization for forest planning, using ellipsoidal uncertainty sets and minimizing deviations of parameters of interest in relation to their reference scenarios. Similarly, the framework of Robust Decision-Making is well suited to address problems of this nature, allowing to search and select robust solutions that meet multiple criteria across a wide range of scenarios or minimizing the regret related to alternative management solutions (e.g. Kasprzyk et al 2013).
4.4. Limitations
We addressed here the impacts of environmental drivers and disturbances on indicators related to multiple forest taxa and TreMs. Although these biodiversity indicators span over a variety of habitat requirements, this is not an exhaustive list of forest taxa and the inclusion of additional indicators is warranted. Moreover, the responses of the different indicators analyzed here was based on (log)linear empirical relationships to stand structure and environmental parameters. Some of the parameters included in our model might diverge from this assumption and display an optimal response (e.g. to temperature). Nevertheless, given a reasonable distance from the optimal condition, linear approximations are able to adequately capture the indicators' responses. We also did not include potential interactions among different indicators. In particular, the abundance of insects may define resource availability for several bat and bird species, and an increase in its population due to increased disturbance activity or warmer climate may enhance the responses of other taxa.
We highlight that the balanced and biodiversity maximization scenarios ((C) and (D), respectively) applied a same weight to the different biodiversity indicators and could, therefore, trade the loss of one indicator by a gain in the other, if preferences for indicators change. Different biodiversity indicators may have different sensitivity towards management interventions, and some respond more to climate drivers than to stand structure. Thus, additional requirements aiming to limit losses related to specific indicators may be included in our framework via the addition of constraints to limit this behavior.
5. Conclusions
Forest management and disturbance events are key drivers of forest dynamics in temperate forests and display complex interactions with species richness. This demands the inclusion of disturbance risk in the planning of management and conservation actions. In this sense, the simultaneous promotion of species persistence and provision of ecosystem services require an integrated and spatially explicit management approach, which balances the maintenance of habitat for multiple forest taxa with production goals. For example, by focusing harvestings and salvage logging where it has least trade-off with biodiversity conservation. Additionally, an expansion of forest reserves may be applied in combination with increased wood utilization in areas with higher forest productivity. Such management diversification schemes, in combination with a dynamic approach to conservation practices that allows the integration of new information as it becomes available, will play a key role on the preservation of ecosystem functioning under environmental change and disturbance drivers.
Acknowledgments
We acknowledge the funding of this research to the German Research Foundation (DFG), ConFoBi project (number GRK 2123). The article processing charge was funded by the Baden-Württemberg Ministry of Science, Research and Art and the University of Freiburg in the funding programme Open Access Publishing.
Data availability statement
The data that support the findings of this study are available upon reasonable request from the authors.