

Abstract
Normal aging is associated with a variable decline in cognitive functions. Among these, executive function, decision-making, and working memory are primarily associated with the prefrontal cortex. Although a number of studies have examined the structural substrates of cognitive decline associated with aging within this cortical area, their functional correlates remain poorly understood. To fill this gap, we aimed to identify functional synaptic substrates of age-associated frontal-dependent deficits in layer 2/3 pyramidal neurons of medial prefrontal cortex of 3-, 9-, and ≥23-month-old Fischer 344 rats. We combined, in the same animals, novelty recognition and exploratory behavioral tasks with assessment of structural and functional aspects of prefrontal synaptic properties. We found that subsets of aged animals displayed stereotyped exploratory behavior or memory deficits. Despite an age-dependent dendritic spine loss, patch-clamp recording of synaptic activity revealed an increase in miniature EPSC frequency restricted to aged animals with preserved exploratory behavior. In contrast, we found a strong positive relationship between miniature IPSC frequency and the occurrence of both stereotyped exploratory behavior and novelty-related memory deficits. The enhanced miniature inhibitory tone was accompanied by a deficit in activity-driven inhibition, also suggesting an impaired dynamic range for modulation of inhibition in the aged, cognitively impaired animals. Together, our data indicate that differential changes in the balance of inhibitory to excitatory synaptic tone may underlie distinct trajectories in the evolution of cognitive performance during aging.
Introduction
With global aging of the population in Western countries, age-related cognitive disorders are a major problem of public health. Hence, numerous research groups have investigated the processes implicated in the pathogenesis of those age-related illnesses. One of the main objectives of research on brain aging in humans is to isolate biomarkers or neurobiological mechanisms implicated in the cognitive and behavioral degradations associated with senescence (Persson et al., 2006; Raji et al., 2009). During aging, a decline in multiple domains of cognitive functions, including short- and long-term memory, psychomotor speed, attention, and executive function, can be observed. Moreover, aging is often accompanied by the development of depression, anxiety, and associated neuropsychiatric symptoms (Lyketsos et al., 2002).
During the last decades, the vast majority of animal studies addressing the consequences of aging on brain function focused on aspects related to memory, particularly spatial memory (Rosenzweig and Barnes, 2003). Numerous studies identified AMPA or NMDA receptor-mediated dysfunction and changes in LTP or LTD properties within several memory-related brain areas, notably the hippocampus and associated temporal areas (Burke and Barnes, 2010). Yet, besides these memory deficits, executive function, decision-making, and working memory also decline in an age-dependent manner (Hedden and Gabrieli, 2004). The prefrontal cortex appears as a critical brain area to study these age-related cognitive declines (Cummings, 1995; Fuster, 2000, 2001). Although a number of studies have addressed age-dependent structural changes, few relate behavioral assessments with single-cell morphometric analyses in the aging prefrontal cortex, especially in rodents. In particular, a study relating these parameters to functional changes in synaptic activity is still lacking. Consequently, the fate of the synaptic homeostasis in aging remains primarily unknown. To fill this gap, we adapted behavioral tests to correlate the animal performances, within different age groups (3, 6–9, and 23–27 months), with indexes of the structural and functional characteristics of synaptic inputs onto layer II/III prefrontal pyramidal neurons. We provide evidence of a functional synaptic imbalance toward inhibitory activity as a substrate of cognitive impairment in the aged cohort while, in contrast, an increase in excitatory synaptic tone is observed in aged unimpaired animals. The balance between basal inhibitory and excitatory synaptic tone may thus distinguish successful from unsuccessful cognitive aging.
Materials and Methods
Animals.
Data presented in this study were obtained from a total of 81 male Fischer 344 rats divided into three groups of age: young (3 months old; N = 10), adult (7.1 ± 1.4 month old; N = 19), and aged (24.2 ± 1.6 month old; N = 52). Animals were obtained from the National Institute on Aging (Bethesda, DC) rodent colony. Animals were maintained on an inverted cycle (light on at 9:00 P.M., light off at 9:00 A.M.) and on ad libitum food and water diet. Animals were allowed to acclimate to their new environment for at least 1 month before starting electrophysiological or behavioral experiments. Animals were removed from the study if presenting any evidence of tumors, illness, or general weakness. All procedures were performed according to the guidelines of the Canadian Council on Animal Care and were approved by the Animal Care Committee of Laval University.
Behavioral testing.
Before testing, animals were transferred from their housing facilities to the testing room once per day for 3 d for at least 1 h, to allow them to acclimate to the room. All experiments were conducted under low-intensity, red-light illumination to minimize visual perceptions from the rats to minimize differences in visual accuracy between the age groups. To avoid any interference caused by proximity of the experimenters, the latter were not present in the room during the testing. All sessions took place between 2:00 and 5:00 P.M. during the dark phase of the light/dark cycle, were recorded with a digital camera, and were stored on a portable personal computer for subsequent analysis. Animals were killed at least 1 week after the last behavioral test to minimize influence of the test itself on the cellular parameters measured.
Modified hole-board task.
To quantify motor and directed exploratory behavior, we modified the classical hole-board task and apparatus (File and Wardill, 1975a, b). The modified hole-board (mHB) apparatus used was a 50 × 50 × 30 cm arena made of a black Plexiglas floor with a hole (3 cm diameter) in each corner. All sessions were subsequently analyzed with an automated tracking system (Any-Maze; Stoelting). Rats were placed individually in the center of the modified hole-board apparatus and allowed 10 min of free exploration of the novel environment. Each rat was tested once. The standard measures were distance traveled, maximum speed reached by the animal, number of head entries in each corner, and average distance from the border (i.e., cumulative sum divided by the total duration of the test). The relative exploration for each corner was established by dividing the number of head entries by the sum of head entries in the four corners. Finally, the corners were ordered from the highest (preferential corner) to the lowest; the preferential corner was then chosen as an index of corner exploration (ICE) for each rat, ranging from 25% (no preferential corner) to 100% (only one corner explored).
Novel object recognition task.
Animals were placed into an adapted test arena for object recognition. The apparatus consisted of a triangular start zone (30 × 30 × 40 cm) separated from a rectangular test zone (40 × 25 cm) by a movable barrier.
The novel object recognition task, for two sample objects within the same environment, was used to study memory after 1 h delays in young, adult, and aged rats. Each session was recorded with a digital camera. First, two identical objects were placed in the chamber as described previously (Bevins and Besheer, 2006). After a 5 min exploration of the objects, the rat was returned to its home cage for 1 h. To test for object recognition, a new copy of the familiar object and a novel object were placed in the chamber and the rat was again placed in the chamber for 5 min to explore the objects. Object recognition was measured in a single trial. A novel object recognition (NOR) index was computed as follows: novel object interaction time/total interaction time with both objects.
Slice preparation and electrophysiology.
Rats were deeply anesthetized with ketamine and xylazine and decapitated. The brain was quickly removed (<60 s) and placed in ice-cold solution containing (in mm) 210 sucrose, 3.0 KCl, 0.75 CaCl2, 3.0 MgSO4 1.0 NaH2PO4, 26 NaHCO3, and 10 glucose, saturated with 95% O2 and 5% CO2. Coronal slices of prefrontal cortex, including the prelimbic area, were cut at 400 μm and kept in artificial CSF (ACSF) containing (in mm) 124 NaCl, 3.0 KCl, 1.5 CaCl2, 1.3 MgSO4 1.0 NaH2PO4, 26 NaHCO3, and 20 glucose, gassed with 95% O2-5% CO2 at room temperature. Slices were allowed to recover for at least 1 h before any recording. A slice was transferred to a submerge-type chamber exposed to ACSF flowing at a rate of 2–3 ml/min. Recordings were performed between 32°C and 34°C.
Whole-cell patch-clamp recording.
Patch pipettes were pulled from borosilicate glass capillaries (World Precision Instruments) and filled with an intracellular solution (pH 7.2; 280–290 mOsm) composed of 100 mm Cs gluconate, 5 mm CsCl, 10 mm HEPES, 2 mm MgCl2, 1 mm CaCl2, 11 mm BAPTA, 4 mm ATP, 0. mm 4 GTP, and 0.25% Neurobiotin (Vector Laboratories). The junction potential of the pipette was corrected by subtracting 11.4 mV before recording. A Multiclamp 700B amplifier (Molecular Devices) was used for the recording. The access resistance was monitored throughout each experiment, and only recordings with stable access and holding current for at least 3 min were used. Mean holding current was not different across age groups (young, −195 ± 46 pA; adult, −150 ± 35 pA; aged, −173 ± 27 pA). Access resistance was not compensated. At the beginning of each recording, three 100-ms-long hyperpolarizing pulses were used to measure the input resistance of the neuron. The extracellular concentration of potassium was raised to 5 mm to increase the frequency of PSCs. After completion of excitatory and inhibitory spontaneous activity recordings, tetrodotoxin (TTX; 1 mm; Sigma/RBI) was also added to the ACSF to block voltage-gated sodium channels and isolate action potential-independent mEPSCs and mIPSCs. All data were acquired and analyzed using pCLAMP (Clampex 9.2 and Clampfit 9.2; Molecular Devices). Data were digitized at 10 kHz and filtered at 1 kHz for subsequent analyses. For EPSC and IPSC analyses, a template was created in Clampfit 9.2 (Clements and Bekkers, 1997).
Immunohistochemistry.
Freshly fixed 400-μm-thick slices taken from 4% PFA were used to inject neurons with Lucifer yellow as described previously (Labrakakis et al., 2009). Briefly, pyramidal neurons in layer II/III of the prelimbic area cortex were impaled with a micropipette containing 1–2% Lucifer yellow (Invitrogen) in 0.2 m PB and injected at 0.5–2 nA for approximately 5–10 min to fill the dendritic tree until spines on apical tuft dendrites became clearly visible. For revelation, slices were rinsed in 0.1 m PB three times for 10 min and incubated overnight at 4°C in a solution of 0.1 m PB, 5% sucrose, and 0.1% Triton. On the second day, three 10 min washes in 0.1 m PB and 0.1% Triton were performed before incubating in a quenching solution (0.1 m PB, 0.1% Triton, 0.3% H2O2) 2 h at room temperature. Slices were washed three times for 10 min in 0.1 m PB and 0.1% Triton. After 2 h at room temperature in a preblocking solution (0.1 m PB, 0.1% Triton, 10% NGS), slices were incubated overnight at 4°C with a rabbit anti-Lucifer yellow primary antibody (1:10,000; Sigma-Aldrich) in 0.1 m PB, 0.1% Triton, 5% NGS. On the third day, slices were incubated in a goat anti-rabbit biotinylated secondary antibody (1:500; Vector Laboratories) for 2 h at room temperature, in 0.1 m PB, after three 10 min washes each in 0.1 m PB. Slices were finally rinsed with three other 10 min washes in 0.1 m PB. The reaction solution was made from an ABC kit (Vectastain; Vector Laboratories). Sections were washed again three times for 10 min in 0.1 m PB. A 0.025% DAB–nickel solution (Sigma-Aldrich) was filtered and applied to slices for 15 min. Then, 0.006% H2O2 was added to the well, and slices were removed from the solution a few seconds after and washed several times in 0.1 m PB to stop the reaction. We thus obtained long-term photostable labeling of dendritic spines and fine neuronal processes (up to the 13th branch order).
Morphometric analysis.
Three to five neurons (3.73 ± 0.79, mean ± SD) per animal were reconstructed in 3D using a DM4000 microscope (Leica) equipped with a 40× objective (1.3 NA), a motorized stage, a video camera system, and Neurolucida morphometry software (MBF Bioscience) by an experimenter blind to the behavioral performance. Only well isolated, single-labeled neurons were used for morphometric analysis, and neurons with blunted apical dendrites were discarded from the quantification. The total length of the apical and basal dendrites was calculated with NeuroExplorer software (MicroBrightField). Sholl analyses (in 10 μm radial increments) were conducted to estimate changes in dendritic tree complexity.
For spine density analysis, three to five neurons (3.67 ± 0.72, mean ± SD) were imaged per animal. Transmitted z-stacks (z-step, 0.4 μm) were acquired with an Axioimager microscope (Zeiss), Plan-Apochromat 63×/1.40 oil-immersion objective, Axiocam camera (16-bit depth, grayscale), and Axiovision software (Zeiss), yielding an image with pixel dimensions of 0.102 × 0.102 × 0.4 μm. Settings for lamp intensity, diaphragm opening size, condenser aperture, and alignment were optimized initially to achieve proper Köhler illumination and held constant throughout the study to ensure that all images were digitized under the same illumination conditions. To be included in the analyses, segments from basal dendritic branches had to satisfy the following criteria: (1) be either parallel to or at acute angles relative to the coronal surface of the section to allow unambiguous identification of spines; and (2) segments had no overlap with other branches that would obscure visualization of spines, avoid the first and last 10% of dendritic segments, and avoid large fluctuations in the dendritic diameter (Dumitriu et al., 2010, 2011; Bloss et al., 2011). For semiautomated spine density analysis, grayscale images were first reversed. With this approach, we were able to use the No Neighbor 2D fluorescence deconvolution algorithm (MetaMorph; Molecular Devices) to remove out-of-focus information (Holmes and O'Connor, 2000). The z-stack images were then imported and analyzed using NeuronStudio software (Rodriguez et al., 2008). Our primary interest was on spine density and the relative change in proportion of spine subclass. Three-dimensional automated NeuronStudio analysis included spine density (number of spines per unit of dendritic length of the segment) and spine type (e.g., mushroom, thin, stubby) (Bloss et al., 2011). Each segment was visually inspected to manually add or remove missing/false-positive spines. Spines were classified as thin or mushroom type if the ratio of their maximum head diameter-to-maximum neck diameter was >1.1. Otherwise, spines were classified as “other,” presumably stubby spines. Because stubby spines were classified based on the absence of a neck and that they can be confounded with a hump on the dendrite (Holmes and O'Connor, 2000; Dumitriu et al., 2010), we decided to restrict our analysis to spines with a clear head, or with a clear increase in signal (i.e., thin and mushroom spines). No further analysis was performed on the other spines (presumably stubby spines). Of the spines that met our criterion, those with a maximum head diameter <0.35 μm and a neck ratio >2.3 were classified as thin spines, and those with maximum spine-head diameters >0.35 μm were classified as mushroom spines (Bloss et al., 2011). To minimize bias, all analyses were done blind to the behavioral performances of animals. Four to 30 (mean, 21.2) 10-μm-long basal dendritic segments (mean, 10.6 μm) were acquired per neuron. Based on these restrictive criteria, ∼11,000 spines were analyzed. Dendritic spine data were collapsed across all dendritic distances (i.e., overall analysis) and first averaged per neuron and then for each animal.
Analysis and statistics.
Electrophysiological and morphological data were analyzed before any behavioral analyses were performed so that the experimenters were blind to the cognitive status of the animal. Cognitive indexes were computed automatically, freeing the measurements from experimenter biases. Finally, normality of data was tested with a Shapiro-Wilk test. Multiple comparisons were performed with the pairwise Sidak-Holm test. Correlations were tested with the Pearson or Spearman product moment correlation coefficient. Data were considered statistically significant at p < 0.05. Results are given as mean ± SEM throughout the text, where n is the number of cells and N is the number of animals.
Results
Lack of change in synaptic activity in the different age groups
To first assess the effect of aging on synaptic activity, we performed, in the prelimbic area of the medial prefrontal cortex, whole-cell patch-clamp recordings of layer II/III pyramidal neurons from young, adult, and aged rats. Neither mean input resistances (young, 94 ± 17.7 MΩ; adult, 102 ± 13.3 MΩ; aged, 151 ± 33.9 MΩ) nor access resistance differed between groups (young, 18.6 ± 0.9 MΩ; adult, 17.3 ± 1.9 MΩ; aged, 16.3 ± 0.65 MΩ). To isolate EPSCs, recordings were performed in voltage-clamp mode while maintaining the membrane potential at the reversal potential for GABAA receptor-mediated PSCs (−60 mV), then to isolate IPSCs, membrane potential was maintained at the reversal potential for ionotropic glutamate receptor-mediated PSCs (0 mV). We first recorded action potential-dependent synaptic activity, and then by adding TTX we isolated action potential-independent spontaneous synaptic activity. As a group, aged rats did not differ from other groups regarding the mean amplitude or frequency of sEPSCs, sIPSCs (Fig. 1A,B), and mEPSCs or mIPSCs (Fig. 2A,B). These results demonstrate so far that, when taken as groups, the different age cohorts showed a comparable level of synaptic activity impinging onto layer II/III pyramidal neurons in the prefrontal cortex.

Lack of change in synaptic properties during aging in the prefrontal cortex. Representative traces of spontaneous synaptic events (sEPSCs, A; sIPSCs, B) in layer 2/3 pyramidal cells obtained from young (blue), adult (gray), and aged (black) rats are shown. No significant changes were observed in the mean frequency or mean amplitude of sEPSCs or sIPSCs. The recording condition is represented in the inset.

Lack of change in miniature synaptic activity during aging in the prefrontal cortex. Representative traces of miniature synaptic events (mEPSCs, A; mIPSCs, B) recorded in layer 2/3 pyramidal cells in the presence of TTX in the same group of rats as in Figure 1, A and B, are shown. No significant changes were observed in the mean frequency or mean amplitude of mEPSCs or mIPSCs (p > 0.05, Sidak-Holm pairwise multiple comparisons; n = 14–55 cells from N = 9–32 rats). The recording condition is represented in the inset.
A subset of aged rat displayed stereotyped exploratory behavior
As described in several studies, the aging process leads to heterogeneous behavioral and cognitive declines in the aged population with major individual differences, allowing segregation of animals into aged impaired versus unimpaired individuals (Fischer et al., 1987; Gage et al., 1988, 1989). To take into account such factors, a series of exploratory and novelty-based behavioral tests were also conducted on a subset of animals before electrophysiological recordings.
To quantify their locomotor performance and directed exploratory behavior, animals were tested in a mHB apparatus (see Materials and Methods for details), with a hole inserted in the floor of each corner. To minimize stress induced during experiments, all tests were conducted under red-light illumination during the dark phase of the light/dark cycle. This was done to reduce the anxiogenic aspect of the procedure, allowing us to study locomotor and exploratory behaviors of the animals independently of emotional factors. Surprisingly, the analysis of animal displacements revealed two main types of exploratory behavior in the aged population. Some aged rats showed excessive exploration in one corner, i.e., an increase in the number of head entries, vertical exploration and hole exploration in the corner zone, consequently neglecting the other corners, characteristic of a stereotyped exploratory behavior. However, this behavior was not generalized to the entire aged population, as some aged rats explored the new environment in a more equilibrated way such as did the young and adult groups (Fig. 3A). We created an index of corner exploration to quantify the pattern of exploration for each rat. The relative exploration for each corner was established by dividing the number of head entries by the sum of head entries in the four corners. For each rat, the corner with the highest relative exploration index (preferential corner) was chosen and labeled as the ICE, theoretically ranging from 25% (no preferential corner) to 100% (only one corner explored).

A subset of aged rats displayed stereotyped exploratory behavior. A, Representative paths for young (Y, blue), adult (Ad, gray), Aged-GE (brown), and Aged-BE (green) rats in the modified hole-board arena diagram (inset) and the distribution of ICE obtained for each group. In the young and adult groups, the distribution of ICE was well fit with single Gaussian functions, whereas the aged group was better fit with a double Gaussian function. We thus isolated two aged subgroups, an Aged-GE group with ICE comparable with that of younger groups and an Aged-BE group with higher ICE indicative of a stereotyped exploratory behavior. The Aged-BE animals were defined as those with an ICE > mean of young + 2 × SD. B, Average score of ICE of young, adult, aged good explorer, and aged bad explorer groups. C, The total distance traveled during the 10 min test in the hole-board arena was significantly lower for Aged-BE. D, The Aged-BE did not display thigmotaxis as shown by the lack of differences in the average distance from the arena border. E, We did not observe a significant difference in the maximum speed of displacement during the test. Data shown are means ± SEM. *p < 0.05; ***p < 0.001, Sidak-Holm pairwise multiple comparisons.
The distributions of the ICE of young and adult groups were well fitted with a single Gaussian function, whereas the distribution of ICE for the aged group was best fitted with a double Gaussian function (Fig. 3A). Based on their ICE, two populations of animals could thus be distinguished in the aged group: (1) an aged group, labeled as the aged good-explorer group (Aged-GE), displayed a directed exploratory behavior similar to that of the young and adult groups; and (2) aged animals with a significantly greater ICE, labeled as the aged bad-explorer group (Aged-BE), henceforth defined as having an ICE deviating by >2 SD above the peak of the distribution observed in the young group. Consequently, the Aged-BE group displayed a significant alteration in their exploratory behavior (Fig. 3B).
The total distance traveled was not significantly different between adults and Aged-GE rats but was significantly lower for Aged-BE rats, consistent with the development of stereotyped exploratory behavior, where the animals preferentially explore the same hole (Fig. 3C). This phenomenon was not explained by an increase in anxiety or thigmotaxis, as demonstrated by the lack of significant difference between the four groups in the average distance traveled from the apparatus border (Fig. 3D) (Simon et al., 1994; Hines et al., 2008). It was also not explained by reduced locomotion performance, as all groups reached similar maximum speed of displacement during the test (Fig. 3E).
A subset of aged rats displayed frontal-related memory deficits
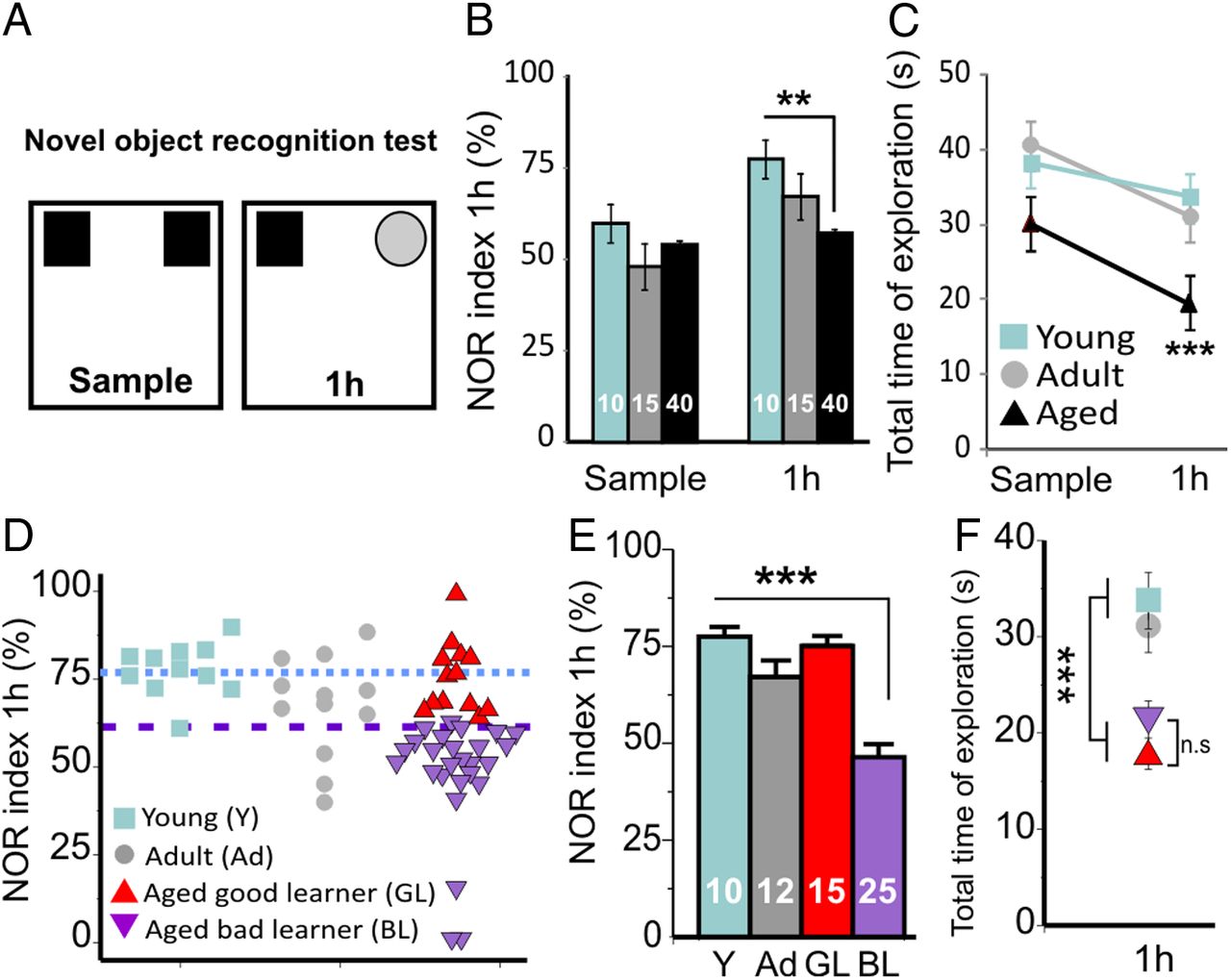
Given the role of the frontal cortex in both directed exploratory behavior and novelty-based, nonspatial memory (Apergis-Schoute et al., 2006), animals were assessed using a NOR test at 1 h retention delay (test phase). This paradigm is known to be dependent on medial prefrontal cortex integrity (Akirav and Maroun, 2006). At the sample phase of the NOR test, no difference was observed between the young, adult, and aged groups, confirming that there was no bias in preference for one side of the apparatus. The total time of exploration was also not statistically different between the three groups at the sample phase (p = 0.12), ruling out any differences in motivation. In contrast, after a 1 h delay, the aged group showed significantly poorer performance than the young and adult groups (Fig. 4B). We also found a decrease in the total exploration time for aged rats during the test phase when compared with the young and adult groups (Fig. 4C).

Novelty-based memory deficits occur in a subgroup of aged rats. A, Mnemonic performances were assessed by a NOR test. Data are presented as the ratio of time spent exploring the new object at 1 h (NOR index). B, During the training phase (sample phase), the rats explored to the same extent the two identical objects. At 1 h, the aged group explored less the new object compared with young rats, confirming a disruption in the memorization process. C, Aged animals explored for a significantly shorter time the two objects at 1 h but for the same duration during the sample phase. D, E, We isolated as Aged-BL (BL) animals with a NOR index lower than that of the young group (Y, blue dotted line) by 2 × SD (purple dotted line). F, The aged group explored less the objects; however, there was no significant difference between the Aged-GL (GL) and Aged-BL (BL) in total time of exploration. **p < 0.01; ***p < 0.001, Sidak-Holm pairwise multiple comparisons.
Following similar logic than for the mHB analysis, the aged group was divided into two subgroups of rats based on their respective performance in the NOR task. To this end, the aged group was divided into two subgroups: a “good learner” (Aged-GL) subgroup, with a NOR index comparable to that in young and adult rats, and a “bad learner” (Aged-BL) group, defined as having a NOR index deviating by > 2 SD above the mean value obtained in the young group (Fig. 4D–F).
The lower total exploration time found in the aged group (Fig. 4C) may reflect poorer motivation in this age group. Thus, the lower NOR index in this group may simply be a consequence of this lower level of motivation. To control for this, we compared the exploration index in the two subgroups of aged animals (Aged-GL and Aged-BL). The total exploration time at 1 h was not significantly different between the Aged-GL and Aged-BL groups, indicating that the lower NOR index in the Aged-BL reflects poorer memory performance and not decreased motivation (Fig. 4F).
Stratification of the aged groups unmasks a correlation between prefrontal synaptic activity and exploratory behavioral performance
We next wanted to investigate whether these behavioral changes echo changes at the synaptic level.
Next we analyzed the synaptic activity after stratification of the aged group into Aged-GE and Aged-BE based on the performance of animals tested in the mHB task (Figs. 5, 6). Whereas no major changes were observed regarding action potential-dependent synaptic activity (Fig. 5), our analysis revealed a significant increase in mean mEPSC frequency for the Aged-GE group (Fig. 6B) when compared with the young and Aged-BE groups. In contrast, the Aged-BE displayed a drastic increased in mean mIPSC frequency (Fig. 6C; p < 0.001). The mean amplitude of events was not changed (Fig. 6B,C). Plotting the frequency of synaptic events against the performance in the mHB for each rat revealed a significant correlation between ICE and mEPSC frequency within the aged group as well as between ICE and mIPSC frequency across age groups and within the aged group (Fig. 6D,E). From our data, such changes seemed to be independent from changes in neuronal integrative properties; indeed, there was no correlation between neuronal input resistance and synaptic activity or behavioral performance, neither across age nor within the aged group (p > 0.05, one-way ANOVA).
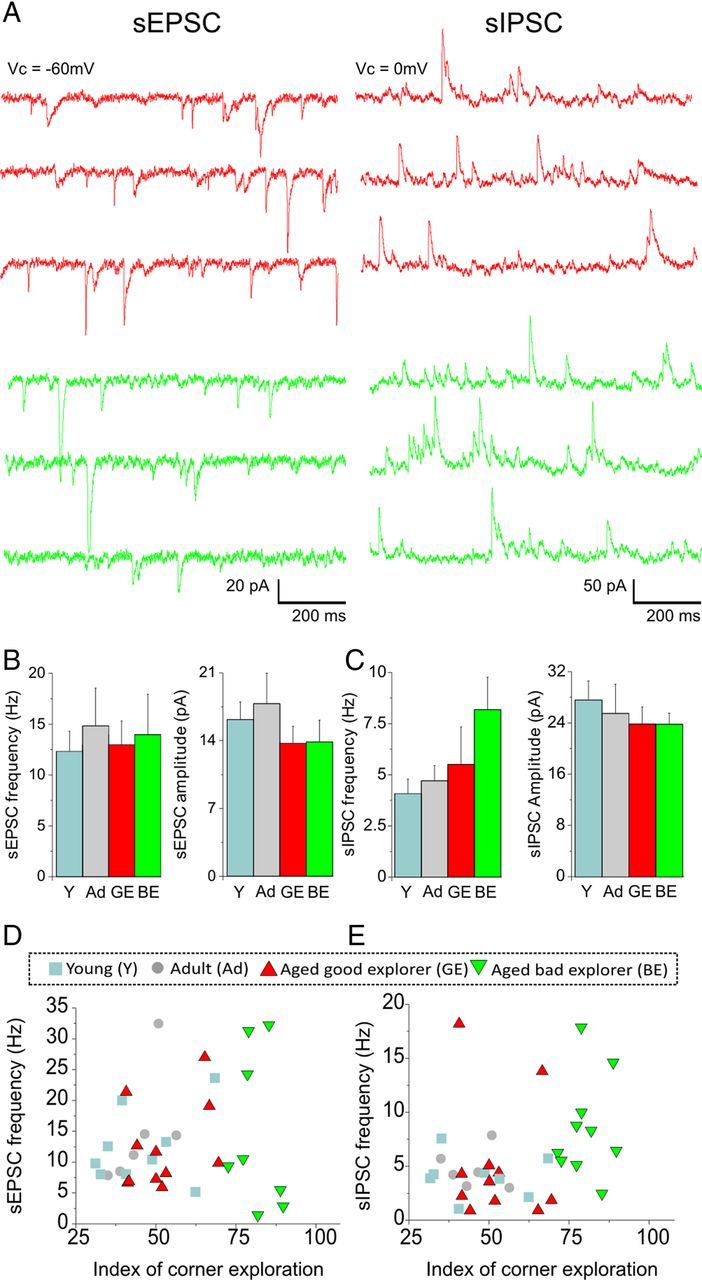
Exploratory behavior is not associated with spontaneous synaptic activity. A, Representative traces of sEPSCs and sIPSCs recorded in layer 2/3 prefrontal pyramidal neurons of Aged-GE (red) and Aged-BE (green) rats. B, There were no changes in sEPSC frequency in layer 2/3 pyramidal neurons from the Aged-GE group when compared with the young (Y) and Aged-BE groups. There was also no change in mean sEPSC amplitude (N = 6–10 rats per group). C, No changes in sIPSC frequency in Aged-BE rats compared with the other groups. There was also no change in mean sIPSC amplitude (N = 6–9 rats per group). D, To evaluate the relationship between excitatory synaptic activity and behavioral performance, the ICE of each rat was plotted against the sEPSC frequency. No significant correlation could be established across the four age groups or within the aged group. E, No relationship between the ICE and the sIPSC frequency across the four groups or within the aged group. Data shown are means ± SEM; Sidak-Holm pairwise multiple comparisons.

Exploratory behavior is associated with specific change in the balance of action potential-independent synaptic transmission in pyramidal neurons of the medial prefrontal cortex. A, Representative traces of mIPSC and mEPSC recorded in layer 2/3 prefrontal pyramidal neurons of Aged-GE (red) and Aged-BE (green) rats. B, We observed a significant increase in mEPSC frequency in layer 2/3 pyramidal neurons from the Aged-GE group when compared with the young (Y) and Aged-BE groups. There was no change in mean mEPSC amplitude (N = 4–8 rats per group). C, On the other hand, our analysis revealed a significant increase in mIPSC frequency in Aged-BE rats compared with the other groups. There was no change in mean mIPSC amplitude (N = 6–9 rats per group). D, To evaluate the relationship between excitatory synaptic activity and behavioral performance, the ICE of each rat was plotted against the mEPSC frequency. No significant correlation could be established across the four age groups; however, we observed a significant correlation within the aged group (p < 0.05). E, A linear positive correlation was established between the ICE and the mIPSC frequency across the four groups (gray text; p < 0.001; r = 0.63) as well as within the aged group (red text and red line; p < 0.05; r = 0.49). Data shown are means ± SEM. *p < 0.05; ***p < 0.001, Sidak-Holm pairwise multiple comparisons.
Aged rats with novelty-based memory impairments also display an imbalance in prefrontal synaptic activity toward inhibition
In line with the above results, stratification of aged animals into good and bad learners revealed an inverse correlation between mnemonic performance and inhibitory synaptic activity impinging onto prefrontal pyramidal neurons (Figs. 7, 8). Again, although no changes in action potential-dependent synaptic activity were seen (Fig. 7), we found that Aged-BL at 1 h displayed a drastic enhancement in GABAA input activity compared with Aged-GL and other age groups (Fig. 8C). In addition, we found, for each rat, a significant correlation between mIPSC frequency and NOR index at 1 h (Fig. 8E) independent of changes in their mean amplitude (Fig. 8B,C). On the other hand, no correlation with mEPSC frequency could be established (Fig. 8C,D). These results also suggest an association between the balance of inhibitory-to-excitatory synaptic activity in the prefrontal cortex and novelty-based, nonspatial memory performance in aged animals.

Memory deficits are not correlated with spontaneous synaptic activity. A, Representative traces of sEPSC and sIPSC recorded in layer II/III prefrontal pyramidal neurons of Aged-GL (red) and Aged-BL (purple) rats. B, No change in sEPSC frequency or amplitude across age groups (N = 6–12 rats per group). C, No changes in sIPSC frequency or amplitude was observed across age groups (N = 6–11 rats per group). Y, Young; Ad, adult. D and E, No correlation between excitatory or inhibitory synaptic activity and the mnemonic performance at 1 h could be established across the four age groups or within the aged group. Sidak-Holm pairwise multiple comparisons were conducted.

Memory deficits are significantly correlated with an imbalance in synaptic activity toward inhibition. A, Representative traces of mIPSC and mEPSC recorded in layer II/III prefrontal pyramidal neurons of Aged-GL (red) and Aged-BL (purple) rats. B, No change in the mean excitatory synaptic frequency or amplitude across age groups (N = 4–8 rats per group). C, In contrast, a significant increase in mIPSC frequency was observed in the Aged-BL group (N = 6–10 rats per group. Y, Young; Ad, adult. D, No correlation between excitatory synaptic activity and memory performances. E, A significant correlation between the inhibitory synaptic activity and the mnemonic performance at 1 h could be established across the four age groups (p < 0.05; r = −0.38) as well as within the aged group (p < 0.05; r = −0.52). Data shown are means ± SEM. ***p < 0.001, Sidak-Holm pairwise multiple comparisons.
Age-dependent changes in dendritic spine density
Changes in the geometry of dendritic tree and spine density are important correlates of aging and cognitive deficits (Luebke and Rosene, 2003; Dumitriu et al., 2010; Luebke et al., 2010; Hara et al., 2012). Therefore, we tested whether any modification in the dendritic tree could underlie changes in the synaptic activity and could thus contribute to the distinct behavioral trajectories occurring in the aged group. To achieve this, we performed single-cell intracellular labeling with Neurobiotin during patch-clamp recording and single-cell Lucifer yellow injection in freshly fixed brain slices (Buhl and Lübke, 1989; Wong et al., 2000; Wong et al., 2006; Cordero-Erausquin et al., 2009; Labrakakis et al., 2009). A total of 56 neurons, with typical morphological features of pyramidal neurons, from 15 animals (young, n = 25 cells from 7 rats; aged, n = 31 cells from 8 rats; Fig. 9A) were analyzed. The morphometric analyses did not reveal significant correlations between the length of dendrite and the performance in the novel object recognition task or the exploratory behavior (Fig. 9B).

Age-dependent changes in dendritic spine density. A, Three examples of pyramidal neurons from layer II/III drawn after injection of Lucifer yellow and followed by anti-Lucifer yellow immunodetection conjugated with DAB-Ni-based reaction. B, No correlation between the dendritic length and behavioral performance. C, Illustration of the methods used to analyze spine density of layer II/III pyramidal neurons: the grayscale image scale was reversed and deconvolved, and a semiautomated analysis of dendritic spine density was performed, yielding a separation of thin and mushroom spines. The other spines, presumably stubby spines, were discarded from analyses. D, Significant age-dependent spine loss. E, Scatter plots of thin spine densities versus exploratory or novel object recognition performance. The thin spine density appears to be predictive of behavioral performance across all groups of age but not within the aged group. F, No relation between mushroom spines density and behavioral performance. **p < 0.01 Student's t test.
We next focused on the evolution of spine density (Fig. 9C) and its role in age-dependent cognitive impairments. Consistent with previous reports (Peters et al., 2008; Dumitriu et al., 2010; Luebke et al., 2010; Bloss et al., 2011; Hara et al., 2012) in nonhuman primate as well as in rodent prefrontal cortex, we found an age-related significant reduction in spine density in layer II/III pyramidal neurons (34% decrease from 0.94 ± 0.17 spines/μm in the young group to 0.62 ± 0.12 in the aged group, mean ± SD; p < 0.001, Student's t test; Fig. 9D). As reported previously (Dumitriu et al., 2010), this change in synaptic density was mainly driven by a loss of thin spines (40% decrease from 0.62 ± 0.10 in the young group to 0.37 ± 0.06 in the aged group; p < 0.001, Student's t test). In contrast, there was no significant age-related change in mushroom spine density (21.8% decrease from 0.32 ± 0.07 in the young group to 0.25 ± 0.06 in the aged group; p = 0.09, Student's t test).
None of the changes in total spine density observed were correlated with the behavioral performance (neither across all groups of age nor within the aged group; data not shown). Yet recent reports indicate that specific depletion of thin spines appear to be a major feature of aging (Dumitriu et al., 2010). We thus evaluated the respective density of thin and mushroom spines for each cell and collapsed for each rat. A significant correlation was found across all groups of age between thin spine density and memory performances (Fig. 9E). However, none of these relationships could be drawn within the aged group. There was no relationship between mushroom spine density and cognitive performance (Fig. 9F). Together, these results rule out the idea that morphological postsynaptic changes are sufficient to explain the enhanced excitatory synaptic activity observed in the Aged-GE group.
Stereotyped exploratory behavior and novelty-based memory deficits are correlated
The above findings implicate an enhancement in inhibitory synaptic activity impinging onto prefrontal pyramidal neurons as a substrate of the cognitive deficits observed in a subgroup of aged animals. This inference is consistent with the finding that the NOR index was correlated with the ICE across all age groups (Fig. 10).

Exploratory behavior correlates with memory performance and inhibitory synaptic activity. A, We found a significant linear correlation between the performances of the animals in the NOR test (NOR index) and the modified hole-board arena test (ICE) (red line; p < 0.01; r = −0.38). The black dotted lines represent the cutoff limits between the impaired and unimpaired status for each test. The majority of animals displayed optimal behavioral performance (yellow region), whereas a subset of aged animals was impaired on both tests (gray region) B, Contour map where frequency (in hertz) of mIPSCs is plotted using color coding, superimposed on the values plotted in A. Animals with the most severe behavioral impairments also displayed an enhanced GABAA receptor-mediated synaptic activity (bottom right quadrant), whereas, at the opposite end, animals with a lower inhibitory synaptic activity displayed the best performance during the behavioral tests (top left quadrant).
Discussion
The present study reports three major findings. First, besides memory deficits, a subset of aged rats display stereotyped exploratory behavior strongly related to the balance in synaptic tone in medial prefrontal cortex. Second, an increase in inhibitory synaptic tone is a common substrate of multiple aging-related cognitive declines. Third, most of the correlates of behavioral performance within the age group seem to be presynaptic.
Aged rats with exploratory or memory deficits displayed an imbalance in background synaptic activity mainly driven by an increase in GABAA receptor-mediated miniature activity. A similar age-related increase in inhibitory tone has been evidenced in studies of rodent and nonhuman primate prefrontal cortex (Abdulla et al., 1995; Griffith and Murchison, 1995; Luebke et al., 2004; Dickstein et al., 2007) Nevertheless, none of these studies was able to establish a direct correlation between these synaptic changes and the degree of age-related, frontal-dependent cognitive impairment. Such discrepancies may rely on both species differences between rodent and primates (Luebke et al., 2010; Morrison and Baxter, 2012) and some experimental design differences (e.g., K+- vs Cs+-based solution), which may affect detection of distal synaptic events. Besides these studies in the frontal cortex, recent reports showed that changes in inhibitory synapse physiology or integrity in temporal structures could underlie some age-dependent memory deficits (Wong et al., 2006; Rissman et al., 2007; Majdi et al., 2009). As such, reducing inhibitory GABAA function by pharmacological intervention has been shown to improve learning ability in aging rodents (Fernandez et al., 2007; Yoshiike et al., 2008). On the other hand, infusion of GABA within the frontal cortex has been shown to be detrimental for normal learning and memory (Meneses et al., 1993; Kleschevnikov et al., 2004; Fernandez et al., 2007), thus reinforcing the crucial role of this cortical area in several memory-dependent tasks.
Interestingly, we observed an increase in AMPA receptor-mediated basal synaptic activity in aged rats with preserved exploratory behavior. Consistent with this observation, several studies reported that pharmacological activation of AMPA receptor induces cognitive performance enhancements in frontal-dependent tasks, such as the delayed nonmatch to sample task, the Y-maze test (Hampson et al., 1998; Bloss et al., 2008), object recognition (Lebrun et al., 2000), or contextual serial discrimination tasks (Beracochea et al., 2007). The increase in glutamatergic tone that we observed in successfully aged animals (i.e., without stereotyped exploratory behavior) is also in line with a recent report demonstrating overactivation of glutamatergic-dependent pathways in successful cognitive aging (Ménard and Quirion, 2012). Moreover, several studies in human reported greater activation in frontal cortex related to better cognitive performance among older participants compared with younger individuals, thus supporting the notion of increased brain activity in successful aging (for review, see Eyler et al., 2011). Nevertheless, no change in excitatory tone was observed in aged good-learner animals. Such discrepancy with the enhanced excitatory activity recorded in aged good explorers may relate to the behavioral domain investigated. Indeed, stereotyped behaviors, as quantified in the present study, have remained virtually not investigated in studies of aging animals (Lalonde and Badescu, 1995; Lalonde and Strazielle, 2009). In contrast, these behaviors have been investigated extensively in animal models of neuropsychiatric disorders (Hines et al., 2008; Kocerha et al., 2009; Chao et al., 2010). Several lines of evidence point to an imbalance in excitatory/inhibitory E/I inputs in prefrontal cortex as a major contributor to psychiatric disorders in humans (for review, see Lewis et al., 2012) as well as abnormal stereotypic behaviors in animal models (Dani et al., 2005; Lisé and El-Husseini, 2006; Hines et al., 2008; Chao et al., 2010). Consistent with this, we found an imbalance in synaptic activity similar to that observed in animal models of neuropsychiatric disease with similar behavioral abnormalities to those observed in our study (Lisé and El-Husseini, 2006; Keith and El-Husseini, 2008; Südhof, 2008). Moreover, it has been shown that stereotyped behavior that result from drug sensitization can be blocked by GABAA antagonists or induced by GABAA agonists (Karler et al., 1995, 1997), thus reinforcing a link between alterations in GABAA transmission in the frontal cortex and appearance of stereotyped behaviors. Together, our findings substantiate the idea that during aging, differential directions in basal synaptic E/I ratio underlie differential cognitive trajectories. As such, whereas an imbalance toward inhibitory tone is detrimental, an imbalance toward excitatory tone may compensate for an age-dependent spine loss and thus preserve optimal exploratory behavior.
Our morphological analyses combined with electrophysiological recordings also demonstrate that despite an age-dependent loss in dendritic spines, there was no significant decrease in the frequency of mEPSCs. On the contrary, we found an increase in mEPSC frequency in cognitively preserved aged rats; thus, a loss of synapses did not necessarily translate into observable functional deficits. Indeed, considering that most of the lost spines are thin spines (Dumitriu et al., 2010; Bloss et al., 2011), i.e., likely silent or weakly contributing to synaptic inputs (Kasai et al., 2003), their loss may not be reflected in significant changes recorded at the levels of the soma. Yet, our results may also indicate that presynaptic mechanisms compensate for the loss of postsynaptic structure, maintaining synaptic homeostasis (Paradis et al., 2001; Turrigiano and Nelson, 2004).
In contrast to the changes observed in mPSC frequency, no change in sEPSC or sIPSC frequency was found in slices from aged rats. Of particular interest, however, is the finding that the frequency of mIPSC in the aged cognitively impaired groups was comparable to that of sIPSCs. This indicates a collapse in the action potential-dependent (or activity-driven) inhibitory tone (i.e., difference between sIPSC and mIPSC frequency) concurrent with the enhanced basal, action potential-independent inhibitory tone. This results in a loss of dynamic range along which inhibition can be modulated combined with an increase in synaptic noise, specifically in the aged, cognitively impaired groups. The net effect may be an impaired capability to adjust network excitability by modulating inhibitory neuron activity, which could be a significant substrate of cognitive impairment. Indeed, the signal-to-noise ratio is an important feature of information processing and deficiency in neuromodulation is suspected to result in unstable cortical representation in aging frontal cortex (Li et al., 2001; Bories et al., 2012). The same phenomenon is not true for the enhanced frequency of mEPSC observed in the aged good explorer group, because in the latter case, a wide dynamic range remained: that is, the frequency of sEPSCs was still threefold larger than that of mEPSCs. The enhanced excitatory tone may still be beneficial in contrast to the enhanced inhibitory tone. These observations call for additional modeling studies to explore the role of excitatory versus inhibitory synaptic activity in stabilizing cortical network activity in aging.
While a number of studies have examined age-related memory deficits (Rosenzweig and Barnes, 2003; Burke and Barnes, 2006; Disterhoft and Oh, 2007), few studies have focused on other cognitive deficits associated with aging, such as psychiatric-like symptoms. Consequently, the neurobiological substrates of such age-related abnormalities have never been clearly identified. We thus focused on this age-related behavioral alteration, given that this could represent part of more complex behavioral alterations appearing in senile dementia. Our findings of similarities between abnormal behavior and changes in E/I ratio displayed by a subset of aged rats and those observed in models of psychiatric disorders thus warrant future investigations into psychiatric symptoms associated with aging in animals. Our results also call for additional studies on presynaptic release of GABA from cortical interneuron in cognitively characterized aged animals to elucidate the biological mechanisms underlying the age-dependent increase in inhibitory tone. Finally, several studies have reported an imbalance in synaptic activity in animal models of Alzheimer's disease at an earlier time point than those studied here (Palop et al., 2007; Sun et al., 2009; Palop and Mucke, 2010; Verret et al., 2012). It thus remains to be investigated whether or not the latter phenotype of Alzheimer's disease represents an early manifestation of the imbalance in synaptic activity we observed in our model of dementia associated with normal aging and whether or not they share a common feature at the synaptic level.
Footnotes
This work was supported by National Institute on Aging Grant R01 AG020529 and Canadian Institutes of Health Research Grant MOP12942 to Y.D.K. C.B. was supported by a studentship from the Centre de Recherche sur le Cerveau, le Comportement et la Neuropsychiatrie. M.J.G. and Y.D.K. are, respectively, Chercheur Boursier and Chercheur National of the Fonds de la Recherche en Santé du Québec.
- Correspondence should be addressed to Dr. Yves De Koninck, 2601, de la Canardière, Québec, QC, Canada G1J 2G3. yves.dekoninck{at}crulrg.ulaval.ca





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}