Assessment of the Potential of Trichoderma spp. Strains Native to Bagua (Amazonas, Peru) in the Biocontrol of Frosty Pod Rot (Moniliophthora roreri)

 ,
, 
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Acquisition of Microorganisms
2.2. In vitro Experiments
- PP = parasitism (%);
- TG = Trichoderma growth;
- N = number of samples taken from each replicate.
- PA = antibiosis (%);
- RG = radial growth of non-confronted M. roreri (mm);
- RGT = radial growth of M. roreri-confronted Trichoderma (mm).
- PA = potential antagonism;
- TM = Trichoderma mycoparasitism against M. roreri (%);
- TA = Trichoderma antibiosis against M. roreri (%).
2.3. Field Experiments
- I: Incidence (%);
- DP: Number of damaged pods;
- TP: Total pods.
- E = efficiency (%);
- FIWoT = % of final incidence without application of Trichoderma spp.;
- FIWT = % of final incidence with application of Trichoderma spp.
2.4. Statistical Analyses
3. Results and Discussion
3.1. In Vitro Mycoparasitism of Trichoderma spp. Against FPR
3.2. In Vitro Antibiosis of Trichoderma spp. Against FPR
3.3. In Vitro Potential Antagonism of Trichoderma spp. Against FPR
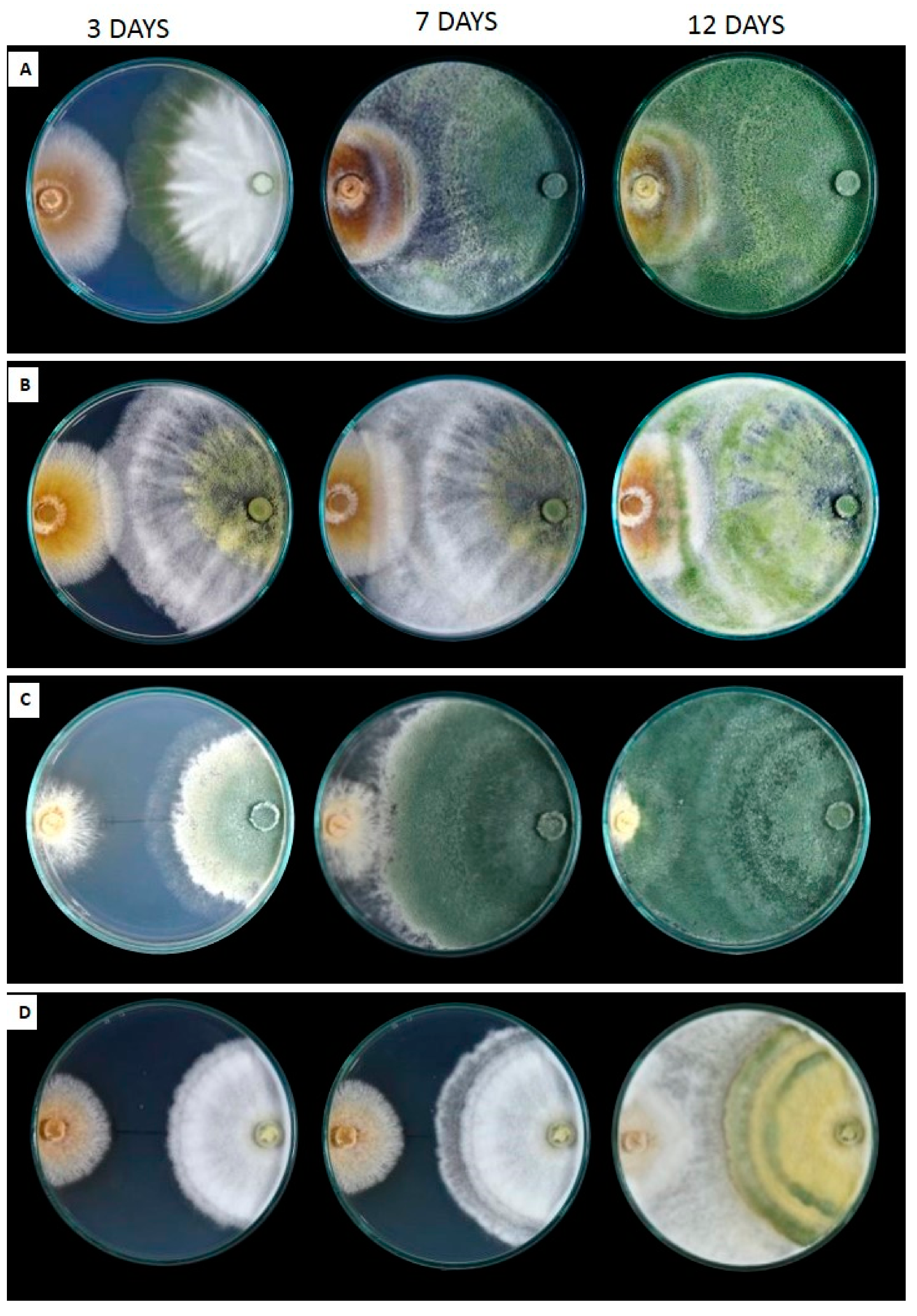
3.4. Field Responses of Trichoderma spp. Against FPR
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
Appendix A



References
- INEI. IV Censo Nacional Agropecuario; Punto&Grafía: Lima, Peru, 2013. [Google Scholar]
- Ministerio de Agricultura y Riego. Análisis de La Cadena Productiva Del Cacao, Con Enfoque En Los Pequeños Productores de Limitado Acceso Al Mercado; DGPA–DEEIA: Lima, Peru, 2018. [Google Scholar]
- Phillips-Mora, W.; Ortiz, C.F.; Aime, M.C. Fifty years of Frosty Pod Rot in Central America: Chronology of its spread and impact from Panama to Mexico. In Proceedings of the 15th International Cocoa Research Conference, San Jose, Costa Rica, 9–14 October 2006; Volume I, pp. 1039–1047. [Google Scholar]
- Díaz-Valderrama, J.R.; Leiva-Espinoza, S.T.; Aime, C. The history of cacao and its diseases in the Americas. Phytopathology 2020, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Griffith, G.W.; Nicholson, J.; Nenninger, A.; Birch, R.N.; Hedger, J.N. Witches’ brooms and frosty pods: Two major pathogens of cacao. N. Z. J. Bot. 2003, 41, 423–435. [Google Scholar] [CrossRef]
- Torres de la Cruz, M.; Ortiz García, C.F.; Téliz Ortiz, D.; Mora Aguilera, A.; Nava Díaz, C. Temporal progress and integrated management of frosty pod rot (Moniliophthora roreri) of cocoa in Tabasco, Mexico. J. Plant Pathol. 2011, 93, 31–36. [Google Scholar] [CrossRef]
- Enríquez, G. Cacao Orgánico: Guía Para Los Productores Ecuatorianos; Instituto Nacional Autónomo de Investigaciones Agropecuarias: Quito, Ecuador, 2004. [Google Scholar]
- Soberanis, W.; Ríos, R.; Arévalo, E.; Zúñiga, L.; Cabezas, O.; Krauss, U. Increased frequency of phytosanitary pod removal in cacao (Theobroma cacao) increases yield economically in eastern Peru. Crop Prot. 1999, 18, 677–685. [Google Scholar] [CrossRef]
- Bateman, R.P.; Hidalgo, E.; García, J.; Arroyo, C.; Ten Hoopen, G.M.; Adonijah, V.; Krauss, U. Application of chemical and biological agents for the management of frosty pod rot (Moniliophthora roreri) in Costa Rican cocoa (Theobroma cacao). Ann. Appl. Biol. 2005, 147, 129–138. [Google Scholar] [CrossRef]
- Gidoin, C.; Avelino, J.; Deheuvels, O.; Cilas, C.; Bieng, M.A.N. Shade tree spatial structure and pod production explain frosty pod rot intensity in cacao agroforests, Costa Rica. Phytopathology 2014, 104, 275–281. [Google Scholar] [CrossRef] [Green Version]
- Ortíz-García, C.F.; Torres-de la Cruz, M.; del Hernández-Mateo, S.C. Comparación de dos sistemas de manejo del cultivo del cacao, en presencia de Moniliophthora roreri, En México. Rev. Fitotec. Mex. 2015, 38, 191–196. [Google Scholar] [CrossRef]
- Leandro-Muñoz, M.E.; Tixier, P.; Germon, A.; Rakotobe, V.; Phillips-Mora, W.; Maximova, S.; Avelino, J. Effects of microclimatic variables on the symptoms and signs onset of Moniliophthora roreri, causal agent of moniliophthora pod rot in cacao. PLoS ONE 2017, 12, e0184638. [Google Scholar] [CrossRef] [Green Version]
- Hidalgo, E.; Bateman, R.; Krauss, U.; Ten Hoopen, M.; Martínez, A. A Field investigation into delivery systems for agents to control Moniliophthora roreri. Eur. J. Plant Pathol. 2003, 109, 953–961. [Google Scholar] [CrossRef]
- Phillips-Mora, W.; Castillo, J.; Krauss, U.; Rodríguez, E.; Wilkinson, M.J. Evaluation of cacao (Theobroma cacao) clones against seven colombian isolates of Moniliophthora roreri from four pathogen genetic groups. Plant Pathol. 2005, 54, 483–490. [Google Scholar] [CrossRef]
- Krauss, U.; Soberanis, W. Rehabilitation of diseased cacao fields in Peru through shade regulation and timing of biocontrol measures. Agrofor. Syst. 2001, 53, 179–184. [Google Scholar] [CrossRef]
- Crozier, J.; Arroyo, C.; Morales, H.; Melnick, R.L.; Strem, M.D.; Vinyard, B.T.; Collins, R.; Holmes, K.A.; Bailey, B.A. The influence of formulation on Trichoderma biological activity and frosty pod rot management in Theobroma cacao. Plant Pathol. 2015, 64, 1385–1395. [Google Scholar] [CrossRef]
- Bailey, B.A.; Bae, H.; Strem, M.D.; Crozier, J.; Thomas, S.E.; Samuels, G.J.; Vinyard, B.T.; Holmes, K.A. Antibiosis, mycoparasitism, and colonization success for endophytic Trichoderma isolates with biological control potential in Theobroma cacao. Biol. Control 2008, 46, 24–35. [Google Scholar] [CrossRef]
- Evans, H.C.; Holmes, K.A.; Thomas, S.E. Endophytes and mycoparasites associated with an indigenous forest tree, Theobroma gileri, in ecuador and a preliminary assessment of their potential as biocontrol agents of cocoa diseases. Mycol. Prog. 2003, 2, 149–160. [Google Scholar] [CrossRef]
- Krauss, U.; Martijn ten Hoopen, G.; Hidalgo, E.; Martínez, A.; Stirrup, T.; Arroyo, C.; García, J.; Palacios, M. The effect of cane molasses amendment on biocontrol of frosty pod rot (Moniliophthora roreri) and black pod (Phytophthora spp.) of cocoa (Theobroma cacao) in Panama. Biol. Control 2006, 39, 232–239. [Google Scholar] [CrossRef]
- Reyes-Figueroa, O.; Ortiz-García, C.F.; De La Cruz, M.T.; Lagunes-Espinoza, L.D.C.; Valdovinos-Ponce, G. Especies de Trichoderma del agroecosistema cacao con potencial de biocontrol sobre Moniliophthora roreri. Rev. Chapingo Ser. Ciencias. y del Ambient. 2016, 22, 149–163. [Google Scholar] [CrossRef]
- Infante, D.; Martínez, B.; González, N.; Reyes, Y. Mecanismos de acción de Trichoderma frente a hongos fitopatógenos. Rev. Protección Veg. 2009, 24, 14–21. [Google Scholar]
- Nelson, P.E.; Toussoun, T.A.; Marasas, W.F.O. Fusarium Species: An Illustrated Manual for Identification; Pennsylvania State University Press: University Park, PA, USA, 1983. [Google Scholar]
- Rifai, M.A. A Revision of the genus Trichoderma. Mycol. Pap. 1969, 116, 1–56. [Google Scholar]
- Samuels, G.J. Trichoderma: Systematics, the sexual state, and ecology. Phytopathology 2006, 96, 195–206. [Google Scholar] [CrossRef] [Green Version]
- Schuster, A.; Schmoll, M. Biology and biotechnology of Trichoderma. Appl. Microbiol. Biotechnol. 2010, 87, 787–799. [Google Scholar] [CrossRef] [Green Version]
- Torres-De La Cruz, M.; Ortiz-García, C.F.; Bautista-Muñoz, C.; Ramírez-Pool, J.A.; Ávalos-Contreras, N.; Cappello-García, S.; De La Cruz-Pérez, A. Diversidad de Trichoderma en el agroecosistema cacao del estado de Tabasco, México. Rev. Mex. Biodivers. 2015, 86, 947–961. [Google Scholar] [CrossRef]
- Estrada, V.; Vélez, A.; López, J. Estandarización de una metodología para obtener cultivos monospóricos del hongo Beauveria bassiana. Cenicafé 1997, 48, 59–65. [Google Scholar]
- Chaverri, P.; Castlebury, L.A.; Overton, B.E.; Samuels, G.J. Hypocrea/Trichoderma: Species with conidiophores elongations and green conidia. Mycologia 2003, 95, 1100–1140. [Google Scholar] [CrossRef]
- Gams, W.; Bissett, J. Morphology and Identification of Trichoderma. In Trichoderma and Gliocladium; Kubicek, C.P., Hamman, G.F., Eds.; Taylor and Francis: London, UK, 2002; pp. 3–34. [Google Scholar]
- Kraus, G.F.; Druzhinina, I.; Gams, W.; Bissett, J.; Zafari, D.; Szakacs, G.; Koptchinski, A.; Prillinger, H.; Zare, R.; Kubicek, C.P. Trichoderma brevicompactum Sp. Nov. Mycologia 2004, 96, 1059. [Google Scholar] [CrossRef] [PubMed]
- Park, M.S.; Bae, K.S.; Yu, S.H. Two new species of Trichoderma associated with green mold of oyster mushroom cultivation in Korea. Mycobiology 2006, 34, 111. [Google Scholar] [CrossRef] [Green Version]
- Samuels, G.J.; Dodd, S.L.; Lu, B.; Petrini, O.; Schroers, H.; Druzhinina, I.S. The Trichoderma koningii aggregate species. Stud. Mycol. 2006, 56, 67–133. [Google Scholar] [CrossRef] [Green Version]
- Samuels, G.J.; Lieckfeldt, E.; Nirenberg, H.I. Trichoderma asperellum, a new species with waited conidia, and redescription of T. viride. Sydowia 1999, 51, 71–88. [Google Scholar]
- Holmes, K.A.; Schroers, H.; Thomas, S.E.; Evans, H.C.; Samuels, J. Taxonomy and biocontrol potential of a new species of Trichoderma from the Amazon Basin of South America. Mycol. Prog. 2004, 3, 199–210. [Google Scholar] [CrossRef]
- Abbott, W.S. A Method of Computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Hebbar, K.P.; Lumsden, R.D. Formulation and Fermentation of Biocontrol Agents of Cocoa Fungal Pathogens: Example of Trichoderma Species. In Research Methodology in Biocontrol of Plant Diseases with Special Reference to Fungal Diseases of Cocoa; Krauss, U., Hebbar, P., Eds.; CATIE: Turrialba, Costa Rica, 1999; pp. 63–68. [Google Scholar]
- Krauss, U.; Soberanis, W. Effect of fertilization and biocontrol application frequency on cocoa pod diseases. Biol. Control 2002, 24, 82–89. [Google Scholar] [CrossRef]
- Sivasithamparam, K.; Ghisalberti, L. Secondary metabolism in Trichoderma and Gliocladium. In Trichoderma and Gliocladium; Harman, G.E., Kubicek, C.P., Eds.; Taylor and Francis Group: London, UK, 1998; pp. 139–191. [Google Scholar]
- Howell, C.R. Mechanisms employed by Trichoderma species in the biological control of plant diseases: The History and evolution of current concepts. Plant Dis. 2003, 87, 4–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinale, F.; Marra, R.; Scala, F.; Ghisalberti, E.L.; Lorito, M.; Sivasithamparam, K. Major secondary metabolites produced by two commercial Trichoderma strains active against different phytopathogens. Lett. Appl. Microbiol. 2006, 43, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Monte, E. Understanding Trichoderma: Between biotechnology and microbial ecology. Int. Microbiol. 2001, 4, 1–4. [Google Scholar] [CrossRef]
- Krauss, U.; Hoopen, M.; Hidalgo, E.; Martínez, A.; Arroyo, C.; García, J.; Portuguez, A.; Sánchez, V. Manejo integrado de la moniliasis (Moniliophthora roreri) del cacao (Theobroma cacao) en Talamanca, Costa Rica. Agrofor. Am. 2003, 10, 52–58. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
| District | UTM Coordinates | Altitude (m.a.s.l.) | Strains |
|---|---|---|---|
| Copallín | 787460/ 9371158 | 894 | CP19-1; CP19-2; CP19-3 |
| 787010/ 9371871 | 931 | CP20-1; CP20-2; CP20-3 | |
| 786828/ 9373275 | 873 | CP22-1; CP22-2; CP22-3 | |
| 786287/ 9373896 | 962 | CP23-1; CP23-2; CP23-3; CP23-4; CP23-5 | |
| 786771/ 9371113 | 820 | CP24-1; CP24-2; CP24-3; CP24-4; CP24-5; CP24-6; CP24-7; CP24-8 | |
| 787395/ 9369923 | 785 | CP43-1; CP43-2; CP43-3; CP43-4; CP43-5 | |
| 787524/ 9369984 | 801 | CP44-1 | |
| 787575/ 9370291 | 825 | CP46-1 | |
| 787663/ 9370405 | 835 | CP49-1; CP49-2 | |
| 787574/ 9370503 | 846 | CP50-1 | |
| 787682/ 9370679 | 850 | CP51-1; CP51-2; CP51-3 | |
| 787695/ 9370824 | 855 | CP52-1; CP52-2 | |
| 787375/ 9371358 | 905 | CP53-1; CP53-2; CP53-3 | |
| 787315/ 9371220 | 898 | CP54-1 | |
| 787414/ 9371086 | 890 | CP55-1; CP55-2; CP55-3; CP55-4 | |
| 787465/ 9371110 | 894 | CP56-1 | |
| 787440/ 9371333 | 913 | CP57-1; CP57-2; CP57-3; CP57-4 | |
| 787398/ 9371399 | 915 | CP59-1; CP59-2 | |
| La Peca | 781248/ 9377461 | 657 | CP14-1; CP14-2; CP14-3; CP14-4; CP14-5; CP14-6; CP14-7 |
| 782400/ 9382298 | 1001 | CP39-1; CP39-2; CP39-3; CP39-4 | |
| 781206/ 9377452 | 647 | CP15-1; CP15-2 | |
| 785913/ 9373121 | 1063 | CP16-1; CP16-2; CP16-3; CP16-4; CP16-5; CP16-6; CP16-7; CP16-8; CP16-9; CP16-10; CP16-11; CP16-12; CP16-13; CP16-14 | |
| 783389/9380375 | 887 | CP17-1; CP17-2; CP17-3; CP17-4; CP17-5; CP17-6; CP17-7 | |
| 782893/ 9380404 | 897 | CP18-1; CP18-2; CP18-3 | |
| 787237/ 9376536 | 1111 | CP25-1 | |
| 786802/ 9377593 | 1137 | CP27-1 | |
| 785718/ 9377204 | 1026 | CP28-1; CP28-2; CP28-3; CP28-4; CP28-5 | |
| La Peca | 785213/ 9377128 | 1011 | CP29-1; CP29-2; CP29-3 |
| 785501/ 9377143 | 1033 | CP30-1 | |
| 782448/ 9377142 | 737 | CP32-1; CP32-2; CP32-3 | |
| 782412/ 9377183 | 731 | CP33-1; CP33-2; CP33-3 | |
| 782353/ 9377313 | 714 | CP34-1; CP34-2; CP34-3; CP34-4 | |
| 782410/ 9377234 | 724 | CP35-1; CP35-2; CP35-3 | |
| 783488/ 9377494 | 764 | CP36-1; CP36-2; CP36-3 | |
| 783926/ 9380799 | 958 | CP37-1; CP37-2 | |
| 783122/ 9381573 | 981 | CP38-1; CP38-2 | |
| 786176/ 9378005 | 1011 | CP42-1; CP42-2; CP42-3; CP42-4; CP42-5 | |
| Aramango | 783111/ 9399062 | 515 | CP1-1; CP1-2; CP1-3; CP1-4; CP1-5; CP1-6 |
| 786053/ 9398435 | 874 | CP61-1; CP61-2; CP61-3; CP61-4 | |
| 786017/ 9398414 | 891 | CP62-1; CP62-2 | |
| 786355/ 9398229 | 949 | CP63-1; CP63-2 | |
| 785646/ 9398424 | 859 | CP64-1 | |
| Imaza | 794828/ 9437021 | 315 | CP4-1; CP4-2; CP4-3; CP4-4; CP4-5 |
| 793717/ 9421156 | 341 | CP6-1; CP6-2; CP6-3; CP6-4; CP6-5; CP6-6; CP6-7 | |
| 800851/ 9427309 | 284 | CP7-1; CP7-2; CP7-3; CP7-4; CP7-5; CP7-6; CP7-7 | |
| 801104/ 9427855 | 286 | CP9-1; CP9-2; CP9-5 | |
| 800750/ 9427725 | 279 | CP8-1; CP8-2; CP8-3; CP8-4 | |
| 800999/ 9427257 | 284 | CP10-1; CP10-2; CP10-3; CP10-4; CP10-5; CP10-6 | |
| 794520/ 9422160 | 320 | CP11-1; CP11-2; CP11-3; CP11-4; CP11-5; CP11-6; CP11-7; CP11-8; CP11-9 | |
| 800705/ 9427717 | 278 | CP12-1; CP12-2; CP12-3; CP12-4; CP12-5 | |
| 801103/ 9427282 | 286 | CP13-1; CP13-2; CP13-3; CP13-4; CP13-5; CP13-6 | |
| 800945/ 9427291 | 285 | CP31-1; CP31-2; CP31-3; CP31-4; CP31-5 | |
| 794808/ 9436976 | 314 | CP5-1; CP5-2 |
| Strain | Mycoparasitism | Strain | Mycoparasitism | Strain | Mycoparasitism | Strain | Mycoparasitism |
|---|---|---|---|---|---|---|---|
| CP18-2 | 100 ± 0a | CP32-2 | 92 ± 4.9a | CP14-4 | 84 ± 9.8b | CP7-5 | 80 ± 6.32b |
| CP18-3 | 100 ± 0a | CP31-1 | 92 ± 8a | CP20-3 | 84 ± 7.48b | CP7-3 | 80 ± 11b |
| CP24-6 | 100 ± 0a | CP19-3 | 92 ± 8a | CP14-7 | 84 ± 7.48b | CP7-6 | 80 ± 8.94b |
| CP25-1 | 100 ± 0a | CP36-1 | 92 ± 4.9a | CP23-4 | 84 ± 7.48b | CP57-3 | 80 ± 11b |
| CP17-7 | 100 ± 0a | CP35-2 | 92 ± 4.9a | CP23-3 | 84 ± 7.48b | CP57-2 | 80 ± 8.94b |
| CP17-1 | 100 ± 0a | CP34-3 | 92 ± 4.9a | CP23-2 | 84 ± 7.48b | CP22-2 | 80 ± 6.32b |
| CP17-2 | 100 ± 0a | CP34-2 | 92 ± 4.9a | CP16-8 | 84 ± 7.48b | CP64-1 | 80 ± 11b |
| CP17-4 | 100 ± 0a | CP12-5 | 92 ± 4.9a | CP16-7 | 84 ± 7.48b | CP8-3 | 80 ± 8.94b |
| CP17-5 | 100 ± 0a | CP16-3 | 92 ± 8a | CP16-6 | 84 ± 7.48b | CP19-1 | 80 ± 12.6b |
| CP28-1 | 100 ± 0a | CP61-3 | 92 ± 4.9a | CP20-1 | 84 ± 11.7b | CP7-1 | 80 ± 8.94b |
| CP37-2 | 100 ± 0a | CP1-4 | 92 ± 3.74a | CP19-2 | 84 ± 4b | CP6-7 | 80 ± 8.94b |
| CP42-4 | 100 ± 0a | CP1-6 | 92 ± 4.9a | CP12-2 | 84 ± 7.48b | CP52-2 | 80 ± 12.6b |
| CP43-3 | 100 ± 0a | CP4-1 | 92 ± 4.9a | CP13-5 | 84 ± 4b | CP23-1 | 80 ± 8.94b |
| CP53-2 | 100 ± 0a | CP61-4 | 92 ± 4.9a | CP11-3 | 84 ± 11.2b | CP57-4 | 76 ± 9.8b |
| CP33-2 | 100 ± 0a | CP62-2 | 92 ± 4.9a | CP13-6 | 84 ± 4b | CP8-2 | 76 ± 7.48b |
| CP28-3 | 100 ± 0a | CP12-4 | 92 ± 4.9a | CP13-4 | 84 ± 7.48b | CP11-5 | 76 ± 9.8b |
| CP28-5 | 100 ± 0a | CP42-3 | 92 ± 4.9a | CP1-1 | 84 ± 9.8b | CP11-4 | 76 ± 7.48b |
| CP29-1 | 100 ± 0a | CP24-7 | 88 ± 7.35a | CP12-3 | 84 ± 7.48b | CP11-2 | 76 ± 11.7b |
| CP32-1 | 100 ± 0a | CP8-4 | 88 ± 8a | CP24-8 | 84 ± 9.8b | CP63-1 | 76 ± 11.7b |
| CP13-1 | 100 ± 0a | CP14-6 | 88 ± 8a | CP24-3 | 84 ± 7.48b | CP6-2 | 76 ± 11.7b |
| CP10-4 | 100 ± 0a | CP24-5 | 88 ± 8a | CP24-2 | 84 ± 9.8b | CP31-2 | 76 ± 7.48b |
| CP16-5 | 100 ± 0a | CP5-2 | 88 ± 4.9a | CP6-4 | 84 ± 7.48b | CP31-3 | 76 ± 7.48b |
| CP10-3 | 100 ± 0a | CP1-3 | 88 ± 8a | CP4-4 | 84 ± 7.48b | CP1-5 | 76 ± 9.8b |
| CP16-14 | 100 ± 0a | CP6-1 | 88 ± 8a | CP55-3 | 84 ± 7.48b | CP31-4 | 76 ± 7.48b |
| CP14-5 | 100 ± 0a | CP31-5 | 88 ± 8a | CP52-1 | 84 ± 4b | CP49-2 | 76 ± 7.48b |
| CP27-1 | 98 ± 2a | CP9-1 | 88 ± 4.9a | CP7-2 | 84 ± 7.48b | CP59-2 | 76 ± 9.8b |
| CP33-3 | 96 ± 4a | CP57-1 | 88 ± 8a | CP38-2 | 82 ± 11.1b | CP55-4 | 76 ± 7.48b |
| CP32-3 | 96 ± 4a | CP59-1 | 88 ± 8a | CP16-2 | 80 ± 8.94b | CP56-1 | 76 ± 11.7b |
| CP38-1 | 96 ± 4a | CP61-1 | 88 ± 5.83a | CP13-3 | 80 ± 8.94b | CP55-1 | 76 ± 7.48b |
| CP36-2 | 96 ± 4a | CP24-1 | 88 ± 8a | CP43-5 | 80 ± 8.94b | CP24-4 | 72 ± 10.2b |
| CP16-13 | 96 ± 4a | CP42-1 | 88 ± 12a | CP16-1 | 80 ± 6.32b | CP11-8 | 72 ± 12b |
| CP16-11 | 96 ± 4a | CP37-1 | 88 ± 8a | CP5-1 | 80 ± 8.94b | CP51-2 | 72 ± 10.2b |
| CP22-1 | 96 ± 4a | CP39-4 | 88 ± 8a | CP10-1 | 80 ± 11b | CP53-3 | 72 ± 8b |
| CP11-7 | 96 ± 4a | CP16-10 | 88 ± 8a | CP53-1 | 80 ± 8.94b | CP62-1 | 72 ± 10.2b |
| CP29-2 | 96 ± 4a | CP11-6 | 88 ± 4.9a | CP46-1 | 80 ± 8.94b | CP6-5 | 72 ± 10.2b |
| CP28-4 | 96 ± 4a | CP17-6 | 88 ± 8a | CP15-1 | 80 ± 8.94b | CP6-3 | 68 ± 8b |
| CP17-3 | 96 ± 4a | CP16-12 | 88 ± 8a | CP50-1 | 80 ± 12.6b | CP8-1 | 68 ± 13.6b |
| CP12-1 | 96 ± 4a | CP6-6 | 88 ± 4.9a | CP42-2 | 80 ± 11b | CP23-5 | 68 ± 8b |
| CP30-1 | 96 ± 4a | CP35-1 | 88 ± 4.9a | CP4-2 | 80 ± 8.94b | CP63-2 | 68 ± 4.9b |
| CP29-3 | 96 ± 4a | CP34-4 | 88 ± 8a | CP1-2 | 80 ± 6.32b | CP43-1 | 68 ± 12b |
| CP14-3 | 96 ± 4a | CP35-3 | 88 ± 8a | CP39-3 | 80 ± 11b | CP51-1 | 68 ± 10.2b |
| CP16-9 | 96 ± 4a | CP4-5 | 88 ± 8a | CP11-9 | 80 ± 6.32b | CP20-2 | 64 ± 11.7b |
| CP28-2 | 96 ± 4a | CP22-3 | 88 ± 8a | CP43-2 | 80 ± 6.32b | CP43-4 | 64 ± 11.7b |
| CP9-5 | 92 ± 4.9a | CP15-2 | 86 ± 6.78a | CP16-4 | 80 ± 8.94b | CP49-1 | 64 ± 13.3b |
| CP9-2 | 92 ± 4.9a | CP4-3 | 84 ± 4b | CP42-5 | 80 ± 11b | CP11-1 | 56 ± 4c |
| CP39-2 | 92 ± 4.9a | CP36-3 | 84 ± 7.48b | CP14-1 | 80 ± 8.94b | CP44-1 | 48 ± 15c |
| CP18-1 | 92 ± 8a | CP61-2 | 84 ± 7.48b | CP55-2 | 80 ± 6.32b | CP51-3 | 44 ± 7.48c |
| CP34-1 | 92 ± 4.9a | CP10-2 | 84 ± 11.7b | CP54-1 | 80 ± 11b | CP13-2 | 36 ± 11.7c |
| CP10-5 | 92 ± 4.9a | CP39-1 | 84 ± 11.7b | CP14-2 | 80 ± 6.32b | CP7-4 | 32 ± 13.6c |
| CP33-1 | 92 ± 4.9a | CP10-6 | 84 ± 11.7b | CP7-7 | 80 ± 11b |
| Strain | Antibiosis | Strain | Antibiosis | Strain | Antibiosis | Strain | Antibiosis |
|---|---|---|---|---|---|---|---|
| CP7-1 | 57.92 ± 4.43a | CP15-2 | 52.91 ± 1.57 ª | CP35-3 | 51.367 ± 0.327a | CP50-1 | 48.47 ± 2.37 ª |
| CP24-6 | 57.27 ± 1.47a | CP16-5 | 52.907 ± 0.341 ª | CP31-2 | 51.32 ± 2.56a | CP62-1 | 48.44 ± 3.44 ª |
| CP23-1 | 56.69 ± 4.99a | CP8-3 | 52.88 ± 2.12 ª | CP33-3 | 51.25 ± 1.3a | CP9-2 | 48.34 ± 1.82 ª |
| CP35-1 | 56.12 ± 2.38a | CP39-4 | 52.87 ± 1.43 ª | CP59-1 | 51.2 ± 2.53a | CP53-3 | 48.33 ± 2.14 ª |
| CP13-4 | 56.03 ± 3.28a | CP16-2 | 52.77 ± 1.77 ª | CP16-11 | 51.166 ± 0.7a | CP55-3 | 48.31 ± 1.21 ª |
| CP38-2 | 55.843 ± 0.522a | CP35-2 | 52.76 ± 2.58 ª | CP6-2 | 51.087 ± 0.418a | CP4-1 | 48.2 ± 1.66 ª |
| CP11-4 | 55.77 ± 1.55a | CP7-5 | 52.75 ± 1.62 ª | CP7-6 | 51.07 ± 1.85a | CP17-1 | 48.17 ± 1.74 ª |
| CP14-7 | 55.765 ± 0.49a | CP7-4 | 52.71 ± 1.67a | CP28-4 | 51.02 ± 1.12a | CP53-1 | 48.12 ± 1.97 ª |
| CP10-6 | 55.46 ± 2.74a | CP17-3 | 52.66 ± 1.06a | CP13-2 | 50.886 ± 0.594a | CP10-1 | 47.934 ± 0.909a |
| CP10-3 | 55.361 ± 0.684a | CP1-6 | 52.6 ± 1.29a | CP11-9 | 50.885 ± 0.879a | CP62-2 | 47.71 ± 2.46 ª |
| CP24-7 | 55.182 ± 0.249a | CP6-3 | 52.584 ± 0.551a | CP14-3 | 50.88 ± 1.99a | CP56-1 | 47.59 ± 1.8 ª |
| CP36-3 | 54.94 ± 3.08a | CP8-2 | 52.58 ± 1.67a | CP22-2 | 50.86 ± 1.73a | CP31-3 | 47.39 ± 2.05 ª |
| CP61-1 | 54.836 ± 0.551a | CP39-1 | 52.477 ± 0.995a | CP34-3 | 50.81 ± 3.33a | CP1-2 | 46.92 ± 8.09b |
| CP16-7 | 54.8 ± 1.18a | CP38-1 | 52.43 ± 1.25a | CP5-2 | 50.81 ± 1.27a | CP20-2 | 46.85 ± 2.56b |
| CP42-2 | 54.76 ± 1.89a | CP4-3 | 52.423 ± 0.383a | CP32-3 | 50.727 ± 0.828a | CP55-1 | 46.74 ± 2.64b |
| CP31-4 | 54.68 ± 2.29a | CP16-10 | 52.39 ± 1.47a | CP6-1 | 50.697 ± 0.473a | CP57-2 | 46.25 ± 1.62b |
| CP19-1 | 54.43 ± 1.26a | CP16-1 | 52.364 ± 0.914a | CP8-4 | 50.67 ± 2.22a | CP23-3 | 46.12 ± 3.44b |
| CP61-3 | 54.28 ± 1.51a | CP24-3 | 52.35 ± 1.42a | CP29-1 | 50.62 ± 2.12a | CP1-3 | 45.95 ± 3.73b |
| CP15-1 | 54.173 ± 0.567a | CP6-5 | 52.26 ± 1.25a | CP18-3 | 50.599 ± 0.598a | CP52-2 | 45.91 ± 2.83b |
| CP43-4 | 54.15 ± 1.58a | CP37-2 | 52.26 ± 1.44a | CP12-1 | 50.49 ± 1.36a | CP51-1 | 45.63 ± 2.14b |
| CP61-2 | 54.14 ± 1.4a | CP17-4 | 52.222 ± 0.803a | CP63-1 | 50.48 ± 2.25a | CP52-1 | 45.46 ± 1.22b |
| CP64-1 | 54.122 ± 0.831a | CP5-1 | 52.221 ± 0.886a | CP9-1 | 50.3 ± 1.26a | CP51-3 | 45.3 ± 1.65b |
| CP42-4 | 54.03 ± 2.42a | CP13-3 | 52.22 ± 1.82a | CP33-1 | 50.208 ± 0.898a | CP55-2 | 45.18 ± 1.69b |
| CP42-1 | 54.01 ± 2.5a | CP16-4 | 52.22 ± 1a | CP29-3 | 50.201 ± 0.861a | CP43-5 | 45.09 ± 2.95b |
| CP11-7 | 53.99 ± 1.18a | CP30-1 | 52.197 ± 0.682a | CP34-2 | 50.15 ± 0.925a | CP23-2 | 44.92 ± 2.91b |
| CP36-2 | 53.98 ± 1.03a | CP18-1 | 52.191 ± 0.933a | CP43-3 | 50.04 ± 6.41a | CP20-3 | 44.7 ± 3.47b |
| CP51-2 | 53.96 ± 1.13a | CP37-1 | 52.15 ± 2.04a | CP16-14 | 50.02 ± 1.05a | CP4-2 | 44.47 ± 2.22b |
| CP16-3 | 53.886 ± 0.825a | CP29-2 | 52.144 ± 0.917a | CP11-5 | 49.983 ± 0.636a | CP22-1 | 44.2 ± 3.4b |
| CP42-3 | 53.88 ± 2.2a | CP10-2 | 52.13 ± 3.86a | CP46-1 | 49.98 ± 4.72a | CP59-2 | 43.82 ± 1.92b |
| CP39-3 | 53.78 ± 2.2a | CP31-5 | 52.12 ± 1.02a | CP49-2 | 49.97 ± 2.9a | CP43-2 | 43.33 ± 2.3b |
| CP11-2 | 53.65 ± 1.54a | CP11-6 | 52.083 ± 0.768a | CP6-6 | 49.9 ± 1.31a | CP1-4 | 43.035 ± 0.929b |
| CP16-13 | 53.593 ± 0.595a | CP10-5 | 52.051 ± 0.913a | CP19-3 | 49.821 ± 0.736a | CP57-4 | 42.96 ± 2.56b |
| CP28-1 | 53.59 ± 1.95a | CP34-1 | 52.03 ± 1.14a | CP13-6 | 49.82 ± 1.88a | CP22-3 | 42.89 ± 1.17b |
| CP16-6 | 53.59 ± 1.92a | CP33-2 | 52.008 ± 0.622a | CP25-1 | 49.733 ± 0.668a | CP23-4 | 42.6 ± 2.51b |
| CP36-1 | 53.57 ± 2.03a | CP32-2 | 51.961 ± 0.704a | CP28-5 | 49.669 ± 0.902a | CP4-5 | 42.48 ± 3.98b |
| CP13-1 | 53.45 ± 1.55a | CP13-5 | 51.91 ± 1.3a | CP18-2 | 49.65 ± 2.49a | CP24-2 | 42.36 ± 1.7b |
| CP24-1 | 53.33 ± 3.08a | CP31-1 | 51.889 ± 0.591a | CP55-4 | 49.65 ± 1.35a | CP23-5 | 41.91 ± 2.8c |
| CP63-2 | 53.27 ± 1.52a | CP6-4 | 51.79 ± 0.595a | CP11-8 | 49.65 ± 1.32a | CP57-1 | 41.52 ± 1.26c |
| CP14-4 | 53.234 ± 0.976a | CP28-3 | 51.75 ± 1.12a | CP9-5 | 49.637 ± 0.989a | CP57-3 | 41.51 ± 2.27c |
| CP61-4 | 53.2 ± 1.76a | CP7-2 | 51.71 ± 1.49a | CP7-7 | 49.38 ± 1.37a | CP49-1 | 41.37 ± 3.13c |
| CP7-3 | 53.15 ± 2.52a | CP14-6 | 51.67 ± 1.57a | CP39-2 | 49.37 ± 1.05a | CP19-2 | 41.15 ± 3.22c |
| CP42-5 | 53.121 ± 0.785a | CP17-7 | 51.665 ± 0.747a | CP1-5 | 49.31 ± 1.83a | CP24-5 | 40.99 ± 2.13c |
| CP14-5 | 53.099 ± 0.342a | CP34-4 | 51.659 ± 0.533a | CP8-1 | 49.07 ± 1.03a | CP1-1 | 40.41 ± 3.47c |
| CP16-8 | 53.044 ± 0.928a | CP24-4 | 51.57 ± 1.33a | CP12-2 | 49.058 ± 0.869a | CP4-4 | 40.34 ± 1.59c |
| CP28-2 | 53.013 ± 0.52a | CP16-12 | 51.53 ± 1.32a | CP54-1 | 48.95 ± 1.15a | CP43-1 | 39.4 ± 3.42c |
| CP12-4 | 53.011 ± 0.451a | CP10-4 | 51.506 ± 0.953a | CP44-1 | 48.72 ± 2.87a | CP53-2 | 38.206 ± 0.85c |
| CP17-2 | 53.01 ± 0.682a | CP16-9 | 51.48 ± 1.07a | CP6-7 | 48.71 ± 0.867a | CP27-1 | 36.13 ± 1.16d |
| CP20-1 | 52.99 ± 1.06a | CP12-5 | 51.456 ± 0.645a | CP11-1 | 48.69 ± 1.91ª | CP24-8 | 35.16 ± 5.7d |
| CP32-1 | 52.946 ± 0.834a | CP14-2 | 51.449 ± 0.962a | CP17-5 | 48.678 ± 0.897ª | CP11-3 | 33.36 ± 1.13d |
| CP14-1 | 52.93 ± 1.42a | CP17-6 | 51.425 ± 0.637a | CP12-3 | 48.54 ± 1.25ª |
| Strain | Potential Antagonism | Strain | Potential Antagonism | Strain | Potential Antagonism | Strain | Potential Antagonism |
|---|---|---|---|---|---|---|---|
| CP24-6 | 78.64 ± 0.735a | CP10-5 | 72.03 ± 2.75a | CP39-1 | 68.24 ± 5.53 ª | CP53-1 | 64.06 ± 5.16b |
| CP10-3 | 77.68 ± 0.342a | CP34-1 | 72.01 ± 2.25a | CP4-3 | 68.21 ± 1.87 ª | CP10-1 | 63.97 ± 5.88b |
| CP42-4 | 77.01 ± 1.21a | CP32-2 | 71.98 ± 2.71a | CP24-3 | 68.17 ± 3.17 ª | CP31-2 | 63.66 ± 3.92b |
| CP28-1 | 76.79 ± 0.962a | CP31-1 | 71.94 ± 4.02a | CP10-2 | 68.06 ± 6.99 ª | CP6-2 | 63.54 ± 5.9b |
| CP13-1 | 76.72 ± 0.773a | CP12-5 | 71.73 ± 2.58a | CP13-5 | 67.95 ± 1.94 ª | CP1-2 | 63.46 ± 5.24b |
| CP14-5 | 76.55 ± 0.171a | CP24-7 | 71.59 ± 3.71a | CP6-4 | 67.89 ± 3.49 ª | CP23-4 | 63.3 ± 3.14b |
| CP17-2 | 76.51 ± 0.341a | CP61-1 | 71.42 ± 2.94a | CP7-2 | 67.85 ± 3.56 ª | CP63-1 | 63.24 ± 5.62b |
| CP32-1 | 76.47 ± 0.417a | CP34-3 | 71.4 ± 2.54a | CP1-4 | 67.52 ± 1.78 ª | CP24-2 | 63.18 ± 4.23b |
| CP16-5 | 76.45 ± 0.787a | CP33-1 | 71.1 ± 2.69a | CP42-2 | 67.38 ± 5.42 ª | CP57-2 | 63.13 ± 4.66b |
| CP37-2 | 76.13 ± 0.719a | CP34-2 | 71.08 ± 2.76a | CP19-1 | 67.21 ± 6.5 ª | CP11-5 | 62.99 ± 5.01b |
| CP17-4 | 76.11 ± 0.502a | CP42-1 | 71 ± 6.99a | CP15-1 | 67.09 ± 4.64a | CP49-2 | 62.98 ± 2.91b |
| CP33-2 | 76 ± 0.311a | CP19-3 | 70.91 ± 3.92a | CP27-1 | 67.07 ± 1.12a | CP51-2 | 62.98 ± 4.86b |
| CP28-3 | 75.87 ± 0.561a | CP9-5 | 70.82 ± 2.49a | CP64-1 | 67.06 ± 5.81a | CP52-2 | 62.95 ± 6.37b |
| CP17-7 | 75.83 ± 0.373a | CP39-2 | 70.68 ± 2.84a | CP1-3 | 66.97 ± 3.14a | CP55-4 | 62.82 ± 3.58b |
| CP10-4 | 75.75 ± 0.476a | CP24-1 | 70.67 ± 4.1a | CP13-6 | 66.91 ± 2.07a | CP1-5 | 62.65 ± 5.66b |
| CP29-1 | 75.31 ± 1.06a | CP39-4 | 70.43 ± 4.6a | CP39-3 | 66.89 ± 5.63a | CP55-2 | 62.59 ± 3.89b |
| CP18-3 | 75.3 ± 0.299a | CP16-10 | 70.19 ± 3.36a | CP7-3 | 66.57 ± 6.19b | CP19-2 | 62.58 ± 2.68b |
| CP43-3 | 75.02 ± 3.21a | CP9-2 | 70.17 ± 2.19a | CP42-5 | 66.56 ± 5.81b | CP43-5 | 62.54 ± 4.73b |
| CP16-14 | 75.01 ± 0.523a | CP22-1 | 70.1 ± 1.67a | CP12-2 | 66.53 ± 3.83b | CP4-2 | 62.23 ± 4.79b |
| CP11-7 | 75 ± 2.28a | CP4-1 | 70.1 ± 2.95a | CP14-1 | 66.47 ± 4.62b | CP1-1 | 62.2 ± 5.69b |
| CP36-2 | 74.99 ± 1.84a | CP37-1 | 70.07 ± 4.39a | CP8-3 | 66.44 ± 5.03b | CP4-4 | 62.17 ± 4.4b |
| CP25-1 | 74.87 ± 0.334a | CP31-5 | 70.06 ± 3.94a | CP16-2 | 66.39 ± 4.5b | CP6-5 | 62.13 ± 4.59b |
| CP28-5 | 74.83 ± 0.451a | CP11-6 | 70.04 ± 2.37a | CP7-5 | 66.37 ± 2.49b | CP56-1 | 61.79 ± 5.82b |
| CP18-2 | 74.82 ± 0.662a | CP13-4 | 70.02 ± 4.79a | CP12-3 | 66.27 ± 3.17b | CP24-4 | 61.78 ± 4.97b |
| CP16-13 | 74.8 ± 2.26a | CP14-7 | 69.88 ± 3.76a | CP16-1 | 66.18 ± 2.93b | CP31-3 | 61.69 ± 4.61b |
| CP28-2 | 74.51 ± 2.18a | CP62-2 | 69.86 ± 1.81a | CP55-3 | 66.15 ± 3.64b | CP43-2 | 61.67 ± 3.41b |
| CP17-5 | 74.34 ± 0.449a | CP14-6 | 69.84 ± 3.5a | CP5-1 | 66.11 ± 4.45b | CP55-1 | 61.37 ± 4.66b |
| CP17-3 | 74.33 ± 1.88a | CP34-4 | 69.83 ± 3.93a | CP13-3 | 66.11 ± 4.36b | CP11-8 | 60.82 ± 5.51b |
| CP38-1 | 74.22 ± 2.38a | CP16-12 | 69.77 ± 3.73a | CP16-4 | 66.11 ± 4.67b | CP57-3 | 60.76 ± 5.8b |
| CP30-1 | 74.1 ± 1.82a | CP10-6 | 69.73 ± 5.65a | CP11-4 | 65.89 ± 4.23b | CP63-2 | 60.63 ± 2.37b |
| CP17-1 | 74.09 ± 0.87a | CP17-6 | 69.71 ± 4.01a | CP14-2 | 65.72 ± 2.78b | CP6-3 | 60.29 ± 3.89b |
| CP29-2 | 74.07 ± 1.79a | CP35-3 | 69.68 ± 4.1a | CP7-6 | 65.54 ± 4.52b | CP62-1 | 60.22 ± 5.94b |
| CP16-9 | 73.74 ± 2.23a | CP59-1 | 69.6 ± 3.54a | CP22-3 | 65.44 ± 4.01b | CP53-3 | 60.16 ± 3.63b |
| CP33-3 | 73.63 ± 1.51a | CP36-3 | 69.47 ± 3.81a | CP11-9 | 65.44 ± 2.79b | CP59-2 | 59.91 ± 4.15b |
| CP16-11 | 73.58 ± 2.01a | CP15-2 | 69.45 ± 3.51a | CP22-2 | 65.43 ± 3.54b | CP24-8 | 59.58 ± 5.66b |
| CP28-4 | 73.51 ± 1.91a | CP5-2 | 69.4 ± 2.96a | CP31-4 | 65.34 ± 4.17b | CP57-4 | 59.48 ± 5.75b |
| CP14-3 | 73.44 ± 2.08a | CP16-7 | 69.4 ± 3.91a | CP4-5 | 65.24 ± 4.79b | CP43-4 | 59.08 ± 5.17b |
| CP32-3 | 73.36 ± 2.28a | CP6-1 | 69.35 ± 4a | CP23-3 | 65.06 ± 4.56b | CP11-3 | 58.68 ± 5.86b |
| CP12-1 | 73.24 ± 2.52a | CP8-4 | 69.33 ± 2.94a | CP46-1 | 64.99 ± 5.96b | CP8-1 | 58.54 ± 6.95b |
| CP61-3 | 73.14 ± 2.98a | CP9-1 | 69.15 ± 2.33a | CP11-2 | 64.83 ± 6.22b | CP51-1 | 56.82 ± 4.66b |
| CP29-3 | 73.1 ± 1.89a | CP53-2 | 69.1 ± 0.425a | CP57-1 | 64.76 ± 3.42b | CP20-2 | 55.42 ± 6.47c |
| CP16-3 | 72.94 ± 4.33a | CP61-2 | 69.07 ± 3.43a | CP52-1 | 64.73 ± 1.75b | CP23-5 | 54.95 ± 2.67c |
| CP42-3 | 72.94 ± 3.34a | CP7-1 | 68.96 ± 5.09a | CP7-7 | 64.69 ± 6.1b | CP43-1 | 53.7 ± 6.33c |
| CP36-1 | 72.79 ± 1.68a | CP6-6 | 68.95 ± 2.53a | CP24-5 | 64.49 ± 3.82b | CP49-1 | 52.68 ± 7.04c |
| CP61-4 | 72.6 ± 3a | CP38-2 | 68.92 ± 5.5a | CP54-1 | 64.47 ± 5.37b | CP11-1 | 52.35 ± 2.15c |
| CP12-4 | 72.51 ± 2.42a | CP16-6 | 68.79 ± 3.67a | CP23-2 | 64.46 ± 2.47b | CP44-1 | 48.36 ± 7.06c |
| CP35-2 | 72.38 ± 1.38a | CP14-4 | 68.62 ± 4.62a | CP6-7 | 64.35 ± 4.32b | CP51-3 | 44.65 ± 4.49c |
| CP1-6 | 72.3 ± 2.65a | CP16-8 | 68.52 ± 4.02a | CP20-3 | 64.35 ± 4.14b | CP13-2 | 43.44 ± 5.63c |
| CP18-1 | 72.1 ± 4.19a | CP20-1 | 68.49 ± 6.03a | CP8-2 | 64.29 ± 3.73b | CP7-4 | 42.36 ± 6.69c |
| CP35-1 | 72.06 ± 3.15a | CP23-1 | 68.35 ± 6.81a | CP50-1 | 64.23 ± 6.69b |
| Site | Treatment | Incidence y (%) | Severity (%) | Efficiency (%) | Yield (kg/ha) |
|---|---|---|---|---|---|
| Copallín | Control | 15.2a | 52.5a | --- z | 826.39b |
| CP10-3 | 4.05c | 30.43b | 71.9a | 1095.0ab | |
| CP53-2 | 8.05bc | 30.6b | 44.53bc | 952.94ab | |
| CP24-6 | 5.62bc | 27.36b | 61.42ab | 1115.0a | |
| CP38-2 | 8.77b | 35.1b | 38.99c | 969.94ab | |
| F = 13.52 p = 0.0005 | F = 11.04 p = 0.001 | F = 14.29 p = 0.0014 | F = 3.97 p = 0.035 | ||
| La Peca | Control | 20.32a | 51.43a | --- | 449.33b |
| CP10-3 | 10.32b | 32.05b | 49.23a | 866.0a | |
| CP53-2 | 9.94b | 33.35b | 51.16a | 873.33 ª | |
| CP24-6 | 10.96b | 28.63b | 45.88a | 787.0a | |
| CP38-2 | 10.58b | 34.10b | 47.79a | 823.67 ª | |
| F = 28.47 p < 0.0001 | F = 31.17 p < 0.0001 | F = 0.5 p = 0.69 | F = 10.57 p = 0.0013 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leiva, S.; Oliva, M.; Hernández, E.; Chuquibala, B.; Rubio, K.; García, F.; Torres de la Cruz, M. Assessment of the Potential of Trichoderma spp. Strains Native to Bagua (Amazonas, Peru) in the Biocontrol of Frosty Pod Rot (Moniliophthora roreri). Agronomy 2020, 10, 1376. https://doi.org/10.3390/agronomy10091376
Leiva S, Oliva M, Hernández E, Chuquibala B, Rubio K, García F, Torres de la Cruz M. Assessment of the Potential of Trichoderma spp. Strains Native to Bagua (Amazonas, Peru) in the Biocontrol of Frosty Pod Rot (Moniliophthora roreri). Agronomy. 2020; 10(9):1376. https://doi.org/10.3390/agronomy10091376
Chicago/Turabian StyleLeiva, Santos, Manuel Oliva, Elgar Hernández, Beimer Chuquibala, Karol Rubio, Flor García, and Magdiel Torres de la Cruz. 2020. "Assessment of the Potential of Trichoderma spp. Strains Native to Bagua (Amazonas, Peru) in the Biocontrol of Frosty Pod Rot (Moniliophthora roreri)" Agronomy 10, no. 9: 1376. https://doi.org/10.3390/agronomy10091376