
Potential Current and Future Distribution of the Long-Whiskered Owlet (Xenoglaux loweryi) in Amazonas and San Martin, NW Peru

 ,
,  ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
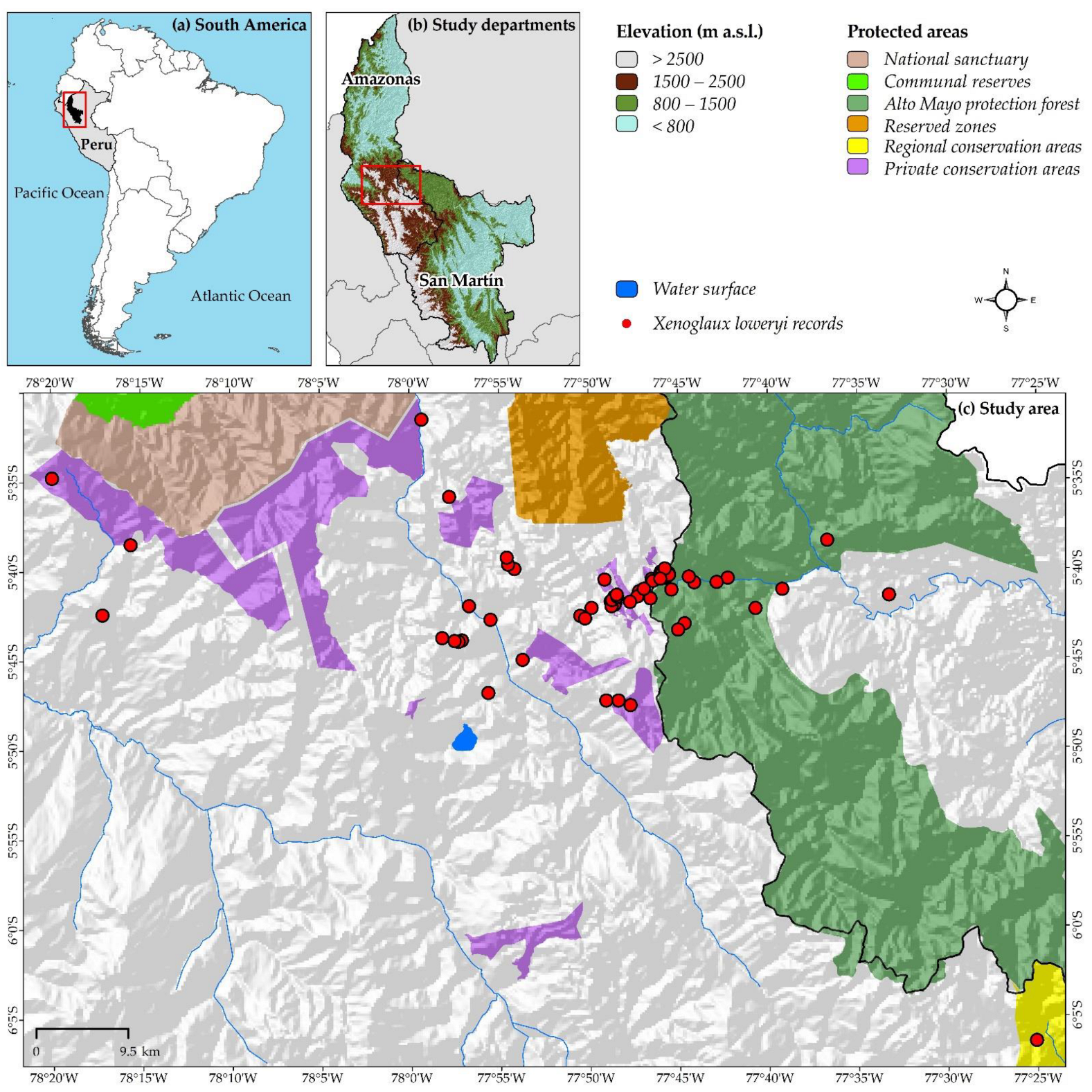
2.1. Study Area
2.2. Georeferenced Records of X. loweryi
2.3. Environmental Variables
2.4. Selection of Environmental Variables
2.5. Modelling under Current and Future Climate Change Scenarios
2.6. Assessment of Habitat Range Change under Current, Future and IUCN Geographic Boundaries
2.7. Changes in Habitat Suitability under Both Climate Change and Human Disturbance Combined
2.8. Identifying Habitat Changes and Priority Areas for Research and Conservation
3. Results
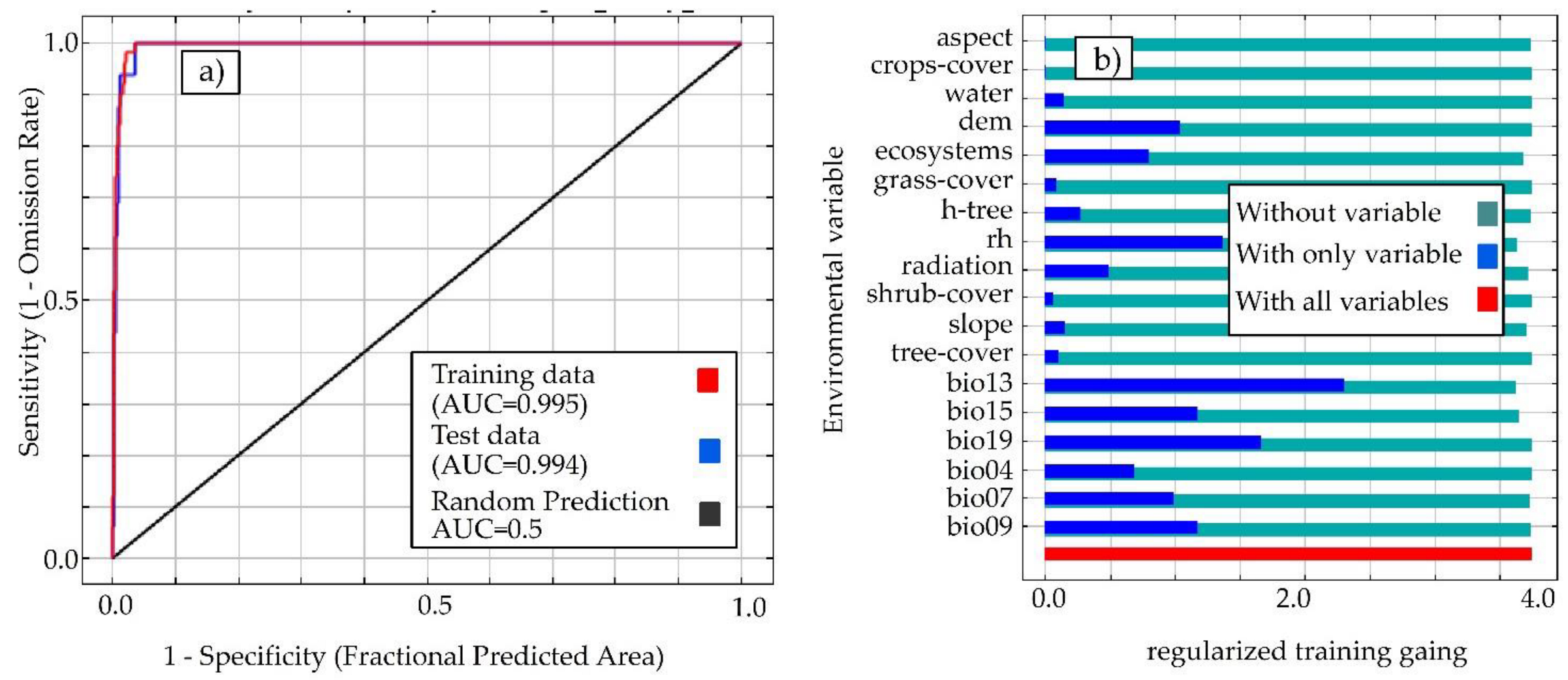
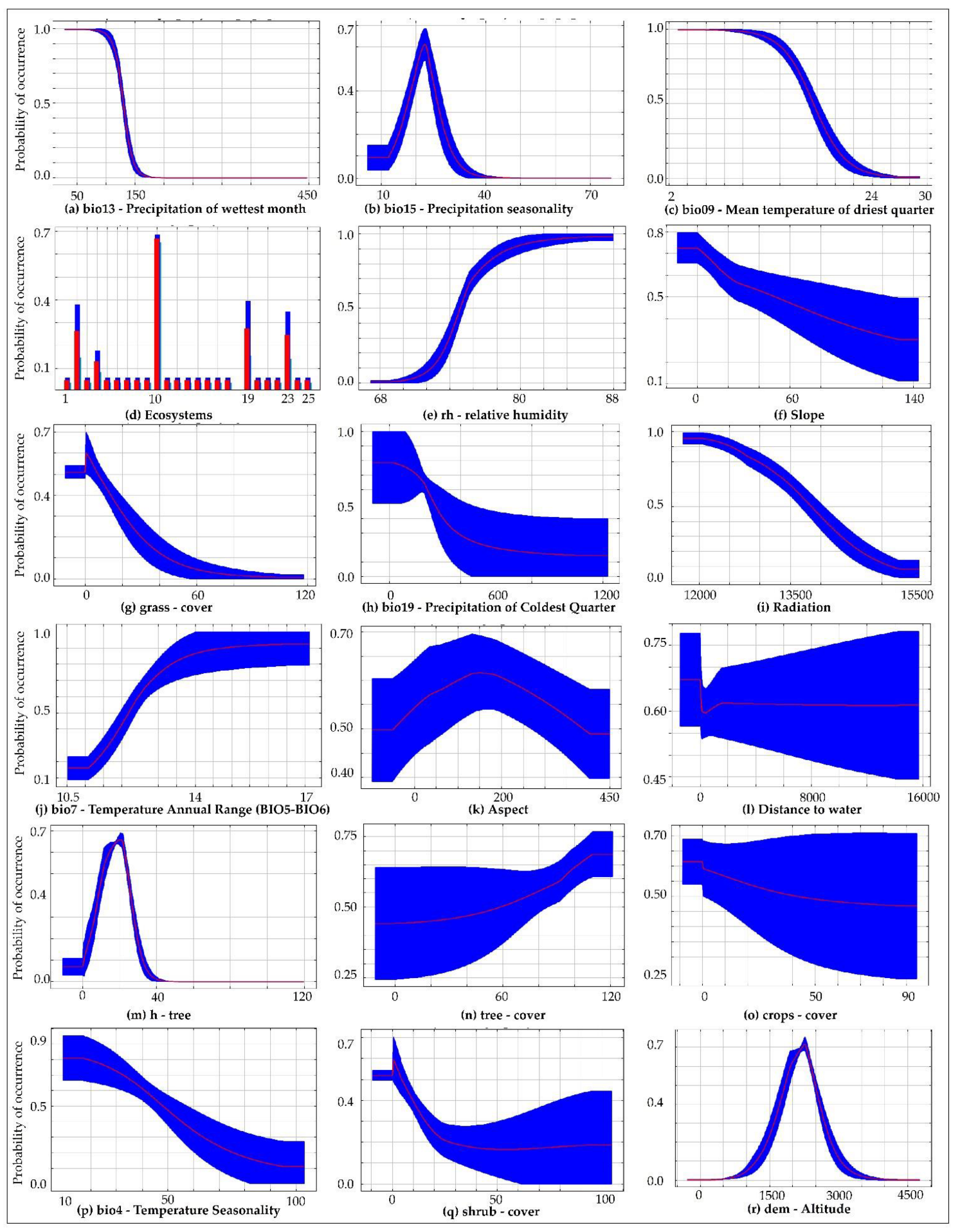
3.1. Model Performance and Contributions of Environmental Variables
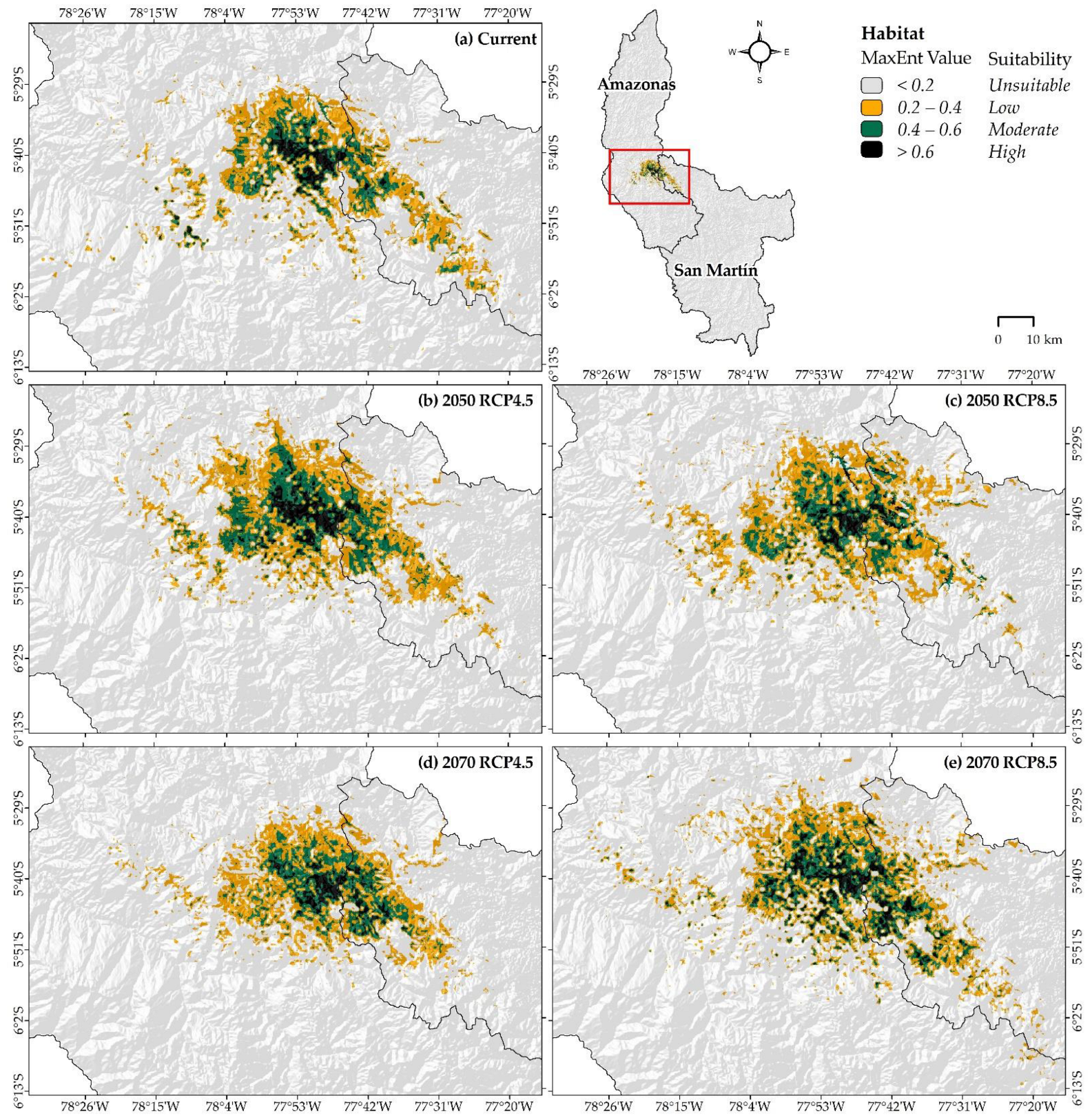
3.2. Potential Distribution under Current and Climate Change Scenarios of X. loweryi
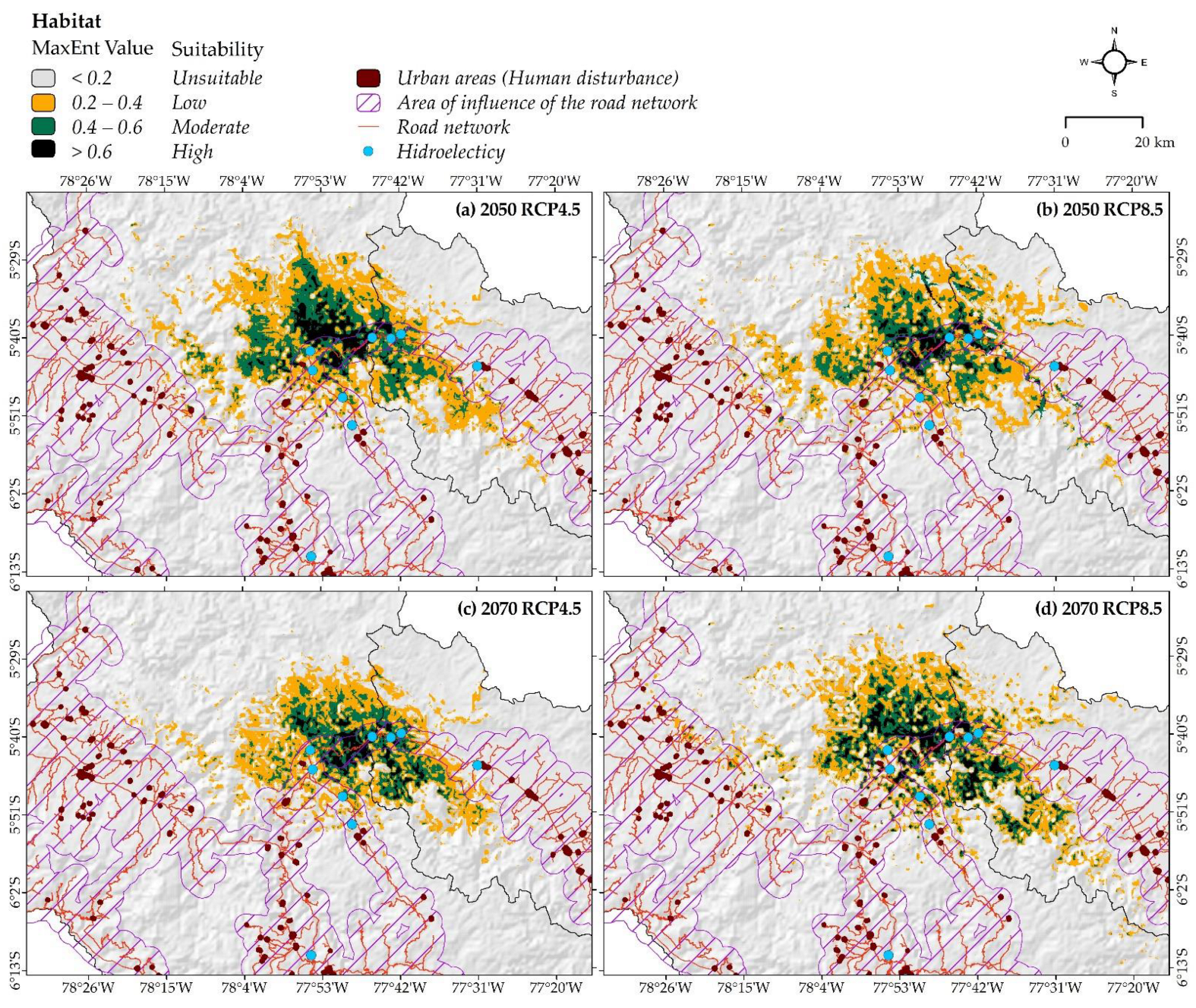
3.3. Changes in Habitat Suitability under Both Climate Change and Human Disturbance Combined
3.4. Stability and Habitat Loss of X. loweryi in Protected Areas
4. Discussion
4.1. Environmental Variables in the Distribution of X. loweryi
4.2. Current Distribution, under Future Climate Change Scenarios and Human Disturbance
4.3. Conservation Priority Areas for X. loweryi
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Neill, J.P.O.; Graves, G.R. A new genus and species of owl (Aves: Strigidae) from Peru. Auk 1977, 94, 409–416. [Google Scholar]
- Angulo Pratolongo, F.; Palomino Condori, W.C.; Arnal-Delgado, H.; Aucca Chutas, C.; Uchofen Mena, Ó. Análisis de distribución de aves de alta prioridad de conservación e identificación de propuestas de áreas para su conservación. In Asociación Ecosistemas Andinos; American Bird Conservancy: Cusco, Perú, 2008; pp. 1–149. [Google Scholar]
- MINAM Listado de Especies CITES Peruanas de Fauna Silvestre; MINAM (Ed.) Ministerio del Ambiente: Lima, Perú, 2018; Volume 1, pp. 1–135.
- Brinkhuizen, D.M.; Shackelford, D. The Long-whiskered Owlets Xenoglaux loweryi of Abra Patricia. Neotrop. Bird. Mag. 2012, 10, 39–46. [Google Scholar]
- Lane, D.F.; Angulo, F. The distribution, natural history, and status of the Long-whiskered Owlet (Xenoglaux loweryi). Wilson J. Ornithol. 2018, 130, 650–657. [Google Scholar] [CrossRef]
- Alarcón, A.; Shanee, S.; Huaman, G.; Shanee, N. Nota sobre la dieta de la Lechucita Bigotona, Xenoglaux loweryi en Yambrasbamba, Amazonas. Rev. Peru. Biol. 2016, 23, 335–338. [Google Scholar] [CrossRef]
- BirdLife International Species Factsheet: Xenoglaux Loweryi. Available online: http://datazone.birdlife.org/species/factsheet/long-whiskered-owlet-xenoglaux-loweryi (accessed on 4 December 2020).
- Young, B.E. Distribución de las Especies Endémicas en la Vertiente Oriental de los Andes en Perú y Bolivia; Wust Ediciones; Nature Serve: Arlington, VA, USA, 2007; ISBN 0971105359. [Google Scholar]
- BirdLife International White-Throated Screech-Owl (Megascops albogularis)—BirdLife Species Factsheet. Available online: http://datazone.birdlife.org/species/factsheet/white-throated-screech-owl-megascops-albogularis (accessed on 23 May 2022).
- BirdLife International Cinnamon Screech-Owl (Megascops petersoni)—BirdLife Species Factsheet. Available online: http://datazone.birdlife.org/species/factsheet/cinnamon-screech-owl-megascops-petersoni (accessed on 23 May 2022).
- BirdLife International Band-Bellied Owl (Pulsatrix melanota) - BirdLife Species Factsheet. Available online: http://datazone.birdlife.org/species/factsheet/band-bellied-owl-pulsatrix-melanota (accessed on 23 May 2022).
- Kouba, M.; Bartoš, L.; Bartošová, J.; Hongisto, K.; Korpimäki, E. Interactive influences of fluctuations of main food resources and climate change on long-term population decline of Tengmalm’s owls in the boreal forest. Sci. Rep. 2020, 10, 1–14. [Google Scholar] [CrossRef]
- Brambilla, M.; Scridel, D.; Bazzi, G.; Ilahiane, L.; Iemma, A.; Pedrini, P.; Bassi, E.; Bionda, R.; Marchesi, L.; Genero, F.; et al. Species interactions and climate change: How the disruption of species co-occurrence will impact on an avian forest guild. Glob. Chang. Biol. 2020, 26, 1212–1224. [Google Scholar] [CrossRef]
- SERFOR. Libro Rojo de la Fauna Silvestre Amenazada del Perú; Primera, Ed.; SERFOR: Lima, Perú, 2018; ISBN 9786124690822.
- Oliva, M.; Maicelo, J.L.; Bardales, W. Propiedades fisicoquímicas del suelo en diferentes estadios de la agricultura migratoria en el Área de Conservación Privada “Palmeras de Ocol”, distrito de Molinopampa, provincia de Chachapoyas (departamento de Amazonas). Rev. Investig. Agroproducción SustenTable 2017, 1, 9–21. [Google Scholar] [CrossRef]
- Shanee, N.; Shanee, S. Land Trafficking, Migration, and Conservation in the “No-Man’s Land” of Northeastern Peru. Trop. Conserv. Sci. 2016, 9, 1940082916682957. [Google Scholar] [CrossRef] [Green Version]
- Dourojeanni, M.J. Ocupación Humana Y Áreas Protegidas De La Amazonia Del Perú. Ecol. Apl. 2014, 13, 225. [Google Scholar] [CrossRef]
- Van Nieuwenhuyse, D.; Genot, J.; Johnson, D.H. The little owl: Conservation, ecology and behavior of Athene noctua. Choice Rev. Online 2009, 46, 46–4441. [Google Scholar] [CrossRef]
- Carrara, E.; Arroyo-Rodríguez, V.; Vega-Rivera, J.H.; Schondube, J.E.; de Freitas, S.M.; Fahrig, L. Impact of landscape composition and configuration on forest specialist and generalist bird species in the fragmented Lacandona rainforest, Mexico. Biol. Conserv. 2015, 184, 117–126. [Google Scholar] [CrossRef]
- Reino, L.; Triviño, M.; Beja, P.; Araújo, M.B.; Figueira, R.; Segurado, P. Modelling landscape constraints on farmland bird species range shifts under climate change. Sci. Total Environ. 2018, 625, 1596–1605. [Google Scholar] [CrossRef] [PubMed]
- BirdLife International Xenoglaux loweryi. The IUCN Red List of Threatened Species. e.T22689320A180768478. Available online: http://datazone.birdlife.org/species/factsheet/long-whiskered-owlet-xenoglaux-loweryi/refs (accessed on 9 February 2021).
- Warren, R.; Vanderwal, J.; Price, J.; Welbergen, J.A.; Atkinson, I.; Ramirez-Villegas, J.; Osborn, T.J.; Jarvis, A.; Shoo, L.P.; Williams, S.E.; et al. Quantifying the benefit of early climate change mitigation in avoiding biodiversity loss. Nat. Clim. Chang. 2013, 3, 678–682. [Google Scholar] [CrossRef] [Green Version]
- Singh, H.; Kumar, N.; Kumar, M.; Singh, R. Modelling habitat suitability of western tragopan (Tragopan melanocephalus) a range-restricted vulnerable bird species of the Himalayan region, in response to climate change. Clim. Risk Manag. 2020, 29, 100241. [Google Scholar] [CrossRef]
- Şekercioĝlu, Ç.H.; Primack, R.B.; Wormworth, J. The effects of climate change on tropical birds. Biol. Conserv. 2012, 148, 1–18. [Google Scholar] [CrossRef]
- Meza, G.M.; Castillo, E.B.; Guzmán, C.T.; Cotrina Sánchez, D.A.; Guzman Valqui, B.K.; Oliva, M.; Bandopadhyay, S.; López, R.S.; Rojas Briceño, N.B. Predictive modelling of current and future potential distribution of the spectacled bear (Tremarctos ornatus) in Amazonas, northeast Peru. Animals 2020, 10, 1816. [Google Scholar] [CrossRef]
- Ariel, L.; Salas, E.; Seamster, V.A.; Boykin, K.G.; Harings, N.M.; Dixon, K.W. Modeling the impacts of climate change on Species of Concern (birds) in South Central U.S. based on bioclimatic variables. AIMS Environ. Sci. 2017, 4, 358–385. [Google Scholar] [CrossRef]
- Sheykhi Ilanloo, S.; Ebrahimi, E.; Valizadegan, N.; Ashrafi, S.; Rezaei, H.R.; Yousefi, M. Little owl (Athene noctua) around human settlements and agricultural lands: Conservation and management enlightenments. Acta Ecol. Sin. 2020, 40, 347–352. [Google Scholar] [CrossRef]
- Phillips, S.J.; Andersonb, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Int. J. Glob. Environ. Issues 2006, 6, 231–252. [Google Scholar] [CrossRef] [Green Version]
- Guisan, A.; Thuiller, W. Predicting species distribution: Offering more than simple habitat models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef]
- Merow, C.; Smith, M.J.; Silander, J.A. A practical guide to MaxEnt for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Anderson, R.P.; Dudík, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Graham, C.H.; Manion, G.; Wisz, M.S.; Zimmermann, N.E.; et al. Novel methods improve prediction of species’ distributions from occurrence data. Eur. J. Biochem. 2006, 29, 129–151. [Google Scholar] [CrossRef] [Green Version]
- Zhu, B.; Wang, B.; Zou, B.; Xu, Y.; Yang, B.; Yang, N.; Ran, J. Assessment of habitat suitability of a high-mountain Galliform species, buff-throated partridge (Tetraophasis szechenyii). Glob. Ecol. Conserv. 2020, 24, e01230. [Google Scholar] [CrossRef]
- Mudereri, B.T.; Mukanga, C.; Mupfiga, E.T.; Gwatirisa, C.; Kimathi, E.; Chitata, T. Analysis of potentially suitable habitat within migration connections of an intra-African migrant-the Blue Swallow (Hirundo atrocaerulea). Ecol. Inform. 2020, 57, 101082. [Google Scholar] [CrossRef]
- Jose, V.S.; Nameer, P.O. The expanding distribution of the Indian Peafowl (Pavo cristatus) as an indicator of changing climate in Kerala, southern India: A modelling study using MaxEnt. Ecol. Indic. 2020, 110, 105930. [Google Scholar] [CrossRef]
- Wang, G.; Wang, C.; Guo, Z.; Dai, L.; Wu, Y.; Liu, H.; Li, Y.; Chen, H.; Zhang, Y.; Zhao, Y.; et al. Integrating Maxent model and landscape ecology theory for studying spatiotemporal dynamics of habitat: Suggestions for conservation of endangered Red-crowned crane. Ecol. Indic. 2020, 116, 106472. [Google Scholar] [CrossRef]
- Li, P.; Zhu, W.; Xie, Z.; Qiao, K. Integration of multiple climate models to predict range shifts and identify management priorities of the endangered Taxus wallichiana in the Himalaya–Hengduan Mountain region. J. For. Res. 2020, 31, 2255–2272. [Google Scholar] [CrossRef] [Green Version]
- Vargas, J. Clima, Informe Temático. In Proyecto Zonificación Ecológica y Económica del Departamento de Amazonas, Convenio Entre el IIAP y el Gobierno Regional de Amazonas; GRA: Iquitos, Perú, 2010; ISBN 9788578110796. [Google Scholar]
- Villegas Mori, E.M.; Pérez Vilca, W.; Altamirano, O. Resultados de la Zonificación Ecológica y Económica del Departamento de San Martín; Primera, Ed.; Gobierno Regional de San Martín: Lima, Perú, 2009; Volume 1982.
- Vargas, J. Clima del Departamento de San Martín. In Proyecto de Zonificación Ecológica y Económica, Convenio Entre el Instituto de Investigaciones de la Amazonía Peruana y el Gobierno Regional de San Martín; GRSM: Iquitos, Perú, 2007. [Google Scholar]
- GBIF.org GBIF Occurrence Download. Available online: https://www.gbif.org/occurrence/download/0025112-200613084148143 (accessed on 20 July 2020).
- Deb, J.C.; Phinn, S.; Butt, N.; McAlpine, C.A. Climatic-Induced Shifts in the Distribution of Teak (Tectona grandis) in Tropical Asia: Implications for Forest Management and Planning. Environ. Manag. 2017, 60, 422–435. [Google Scholar] [CrossRef]
- Boria, R.A.; Olson, L.E.; Goodman, S.M.; Anderson, R.P. Spatial filtering to reduce sampling bias can improve the performance of ecological niche models. Ecol. Modell. 2014, 275, 73–77. [Google Scholar] [CrossRef]
- de Oliveira-Silva, A.E.; Piratelli, A.J.; Zurell, D.; da Silva, F.R. Vegetation cover restricts habitat suitability predictions of endemic Brazilian Atlantic Forest birds. Perspect. Ecol. Conserv. 2021, 20, 1–8. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- IPCC Climate Change 2014: Synthesis Report. In Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneve, Switzerland, 2014; ISBN 9789291691432.
- Meehl, G.A.; Washington, W.M.; Arblaster, J.M.; Hu, A.; Teng, H.; Tebaldi, C.; Sanderson, B.N.; Lamarque, J.F.; Conley, A.; Strand, W.G.; et al. Climate system response to external forcings and climate change projections in CCSM4. J. Clim. 2012, 25, 3661–3683. [Google Scholar] [CrossRef]
- Gent, P.R.; Danabasoglu, G.; Donner, L.J.; Holland, M.M.; Hunke, E.C.; Jayne, S.R.; Lawrence, D.M.; Neale, R.B.; Rasch, P.J.; Vertenstein, M.; et al. The community climate system model version 4. J. Clim. 2011, 24, 4973–4991. [Google Scholar] [CrossRef]
- van Vuuren, D.P.; Edmonds, J.; Kainuma, M.; Riahi, K.; Thomson, A.; Hibbard, K.; Hurtt, G.C.; Kram, T.; Krey, V.; Lamarque, J.F.; et al. The representative concentration pathways: An overview. Clim. Chang. 2011, 109, 5–31. [Google Scholar] [CrossRef]
- Farr, T.; Rosen, P.A.; Caro, E.; Crippen, R.; Duren, R.; Hensley, S.; Kobrick, M.; Paller, M.; Rodriguez, E.; Roth, L.; et al. Shuttle Radar Topography Mission: Mission to map the world. Rev. Geophys. 2007, 45, 3–5. [Google Scholar] [CrossRef]
- MINEDU Descarga de Información Espacial del MED. Available online: http://sigmed.minedu.gob.pe/descargas/ (accessed on 5 December 2020).
- Buchhorn, M.; Smets, B.; Bertels, L.; Roo, B.; Lesiv, M.; Tsendbazar, N.-E.; Herold, M.; Fritz, S. Copernicus Global Land Service: Land Cover 100m: Collection 3: Epoch 2015: Globe. VITO 2020, 580265. [Google Scholar] [CrossRef]
- MINAM INTERCAMBIO de Datos – Geoservidor. Available online: https://geoservidor.minam.gob.pe/recursos/intercambio-de-datos/ (accessed on 5 December 2020).
- Potapov, P.; Li, X.; Hernandez-Serna, A.; Tyukavina, A.; Hansen, M.C.; Kommareddy, A.; Pickens, A.; Turubanova, S.; Tang, H.; Silva, C.E.; et al. Mapping global forest canopy height through integration of GEDI and Landsat data. Remote Sens. Environ. 2020, 253, 112165. [Google Scholar] [CrossRef]
- New, M.; Lister, D.; Hulme, M.; Makin, I. A high-resolution data set of surface climate over global land areas. Clim. Res. 2002, 21, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Varouchakis, E.A. Geostatistics: Mathematical and Statistical Basis; Elsevier Inc.: Amsterdam, The Netherlands, 2019; ISBN 9780128116890. [Google Scholar]
- Tanner, E.P.; Papeş, M.; Elmore, R.D.; Fuhlendorf, S.D. Incorporating abundance information and guiding variable selection for climate-based ensemble forecasting of species’ distributional shifts. PLoS ONE 2017, 12, e0184316. [Google Scholar] [CrossRef] [Green Version]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J.; et al. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Sony, R.K.; Sen, S.; Kumar, S.; Sen, M.; Jayahari, K.M. Niche models inform the effects of climate change on the endangered Nilgiri Tahr (Nilgiritragus hylocrius) populations in the southern Western Ghats, India. Ecol. Eng. 2018, 120, 355–363. [Google Scholar] [CrossRef]
- Fielding, A.H.; Bell, J.F. A review of methods for the assessment of prediction errors in conservation presence/absence models. Environ. Conserv. 1997, 24, 38–49. [Google Scholar] [CrossRef]
- Warren, D.L.; Seifert, S.N. Ecological niche modeling in Maxent: The importance of model complexity and the performance of model selection criteria. Ecol. Appl. 2011, 21, 335–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araujo, M.; Pearson, R.; Thuiller, W.; Erhard, M. Validation of species-climate impact models under climate change. Glob. Chang. Biol. 2005, 11, 1504–1513. [Google Scholar] [CrossRef] [Green Version]
- Manel, S.; Williams, H.C.; Ormerod, S.J. Evaluating presence—Absence models in ecology: The need to account for prevalence. J. Appl. Ecol. 2001, 38, 921–931. [Google Scholar] [CrossRef]
- Barrett, M.A.; Brown, J.L.; Junge, R.E.; Yoder, A.D. Climate change, predictive modeling and lemur health: Assessing impacts of changing climate on health and conservation in Madagascar. Biol. Conserv. 2013, 157, 409–422. [Google Scholar] [CrossRef]
- Ansari, H.M.; Ghoddousi, A. Water availability limits brown bear distribution at the southern edge of its global range. Ursus 2018, 29, 13–24. [Google Scholar] [CrossRef]
- Zhang, J.; Jiang, F.; Li, G.; Qin, W.; Li, S.; Gao, H.; Cai, Z.; Lin, G.; Zhang, T. Maxent modeling for predicting the spatial distribution of three raptors in the Sanjiangyuan National Park, China. Ecol. Evol. 2019, 9, 6643–6654. [Google Scholar] [CrossRef] [Green Version]
- Zhen, J.; Wang, X.; Meng, Q.; Song, J.; Liao, Y.; Xiang, B.; Guo, H.; Liu, C.; Yang, R.; Luo, L. Fine-scale evaluation of giant panda habitats and countermeasures against the future impacts of climate change and human disturbance (2015–2050): A case study in Ya’an, China. Sustainability 2018, 10, 1081. [Google Scholar] [CrossRef] [Green Version]
- MTC Ministerio de Transportes y Comunicaciones: Transporte Terrestre por Carretera. Available online: https://portal.mtc.gob.pe/estadisticas/descarga.html (accessed on 6 December 2020).
- ANP, S.G. Visor de las Áreas Naturales Protegidas. Available online: http://geo.sernanp.gob.pe/visorsernanp/ (accessed on 6 December 2020).
- MINAM Mapa Nacional de Ecosistemas del Perú: Memoria Descriptiva; Dirección General de Ordenamiento Territorial Ambiental; MINAM: Lima, Perú, 2018.
- All, U.T.C. Thermal Effects of Radiation and Wind on a Small Bird and Implications for Microsite Selection. Ecology 2016, 77, 2228–2236. [Google Scholar]
- Vié, J.-C.; Hilton-Taylor, C.; Stuart, S.N. Wildlife in a Changing World—An Analysis of the 2008 IUCN Red List of Threatened Species; IUCN: Gland, Switzerland, 2009; ISBN 9782831710631. [Google Scholar]
- Peterson, A.T.; Sánchez-Cordero, V.; Soberón, J.; Bartley, J.; Buddemeier, R.W.; Navarro-Sigüenza, A.G. Effects of global climate change on geographic distributions of Mexican Cracidae. Ecol. Modell. 2001, 144, 21–30. [Google Scholar] [CrossRef]
- Rojas Briceño, N.B.; Barboza Castillo, E.; Maicelo Quintana, J.L.; Oliva Cruz, S.M.; Salas López, R. Deforestación en la Amazonía peruana: Índices de cambios de cobertura y uso del suelo basado en SIG. Boletín la Asoc. Geógrafos Españoles 2019, 81, 1–34. [Google Scholar] [CrossRef]
- Mendoza Chichipe, M.E.; Salas López, R.; Barboza Castillo, E. Análisis multitemporal de la deforestación usando la clasificación basada en objetos, distrito de Leymebamba (Perú). Rev. Investig. Para El Desarro. SustenTable 2017, 3, 67–76. [Google Scholar] [CrossRef] [Green Version]
- Salas López, R.; Barboza Castillo, E.; Oliva Cruz, M. Dinámica multitemporal de índices de deforestación en el distrito de Florida, departamento de Amazonas, Perú. Indes 2016, 2, 1–9. [Google Scholar] [CrossRef]
- Arroyave Maya, M.; Gómez, C.; Gutiérrez, M.; Múnera, D.; Zapata, P.; Vergara, I.; Andrade, L.; Ramos, K. Impactos de las carreteras sobre la fauna silvestre y sus principales medidas de manejo. Rev. EIA 2006, 5, 45–57. [Google Scholar] [CrossRef]
- Morelli, F.; Rodríguez, R.A.; Benedetti, Y.; Delgado, J.D. Avian roadkills occur regardless of bird evolutionary uniqueness across Europe. Transp. Res. Part D Transp. Environ. 2020, 87, 102531. [Google Scholar] [CrossRef]
- Figueroa, C.J.P.; Ruíz, R.D.C.L.; Guadarrama, E.M.; Leal, J.D.D.V.; Méndez, J.S.; Zayas, E.E.M.; Campillo, L.M.G.; Ruíz, L.J.R.; Hernández, Y.S.C.; Ruíz, F.S.Z. Un asesino a sueldo: El impacto de las carreteras en la fauna silvestre. Kuxulkab’ 2014, 20. [Google Scholar]
- Pérez-García, J.M.; DeVault, T.L.; Botella, F.; Sánchez-Zapata, J.A. Using risk prediction models and species sensitivity maps for large-scale identification of infrastructure-related wildlife protection areas: The case of bird electrocution. Biol. Conserv. 2017, 210, 334–342. [Google Scholar] [CrossRef] [Green Version]
- Monteferri, B. Áreas de Conservación Privada en el Perú: Avances y Propuestas a 20 Años de su Creación; Sociedad Peruana de Derecho Ambiental: Lima, Peru, 2019; ISBN 9786124261442. [Google Scholar]
- Aquino, R.; Encarnación, F. Proyecto Zonificación Ecológica y Económica del Departamento de Amazonas, Convenio Entre el IIAP y el Gobierno Regional de Amazonas; GRSM: Iquitos, Perú, 2010; ISBN 9788578110796. [Google Scholar]
- Juhani Mikkola, H. Diversity of the Owl Species in the Amazon Region. In Ecosystem and Biodiversity of Amazonia; InTech Open: London, UK, 2021; pp. 107–119. [Google Scholar] [CrossRef]
- Salinas-Rodríguez, M.M.; Sajama, M.J.; Gutiérrez-Ortega, J.S.; Ortega-Baes, P.; Estrada-Castillón, A.E. Identification of endemic vascular plant species hotspots and the effectiveness of the protected areas for their conservation in Sierra Madre Oriental, Mexico. J. Nat. Conserv. 2018, 46, 6–27. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Habitat Suitability | Current (km2) | 2050 1 | 2070 (%) 1, 2 | ||
|---|---|---|---|---|---|
| RCP 4.5 | RCP 8.5 | RCP 4.5 | RCP 8.5 | ||
| High | 140.85 | 186.32 | 133.95 | 126.28 | 234.19 |
| 32.3 | −4.9 | −10.3 (−32.2) | 66.3 (74.8) | ||
| Moderate | 416.88 | 553.93 | 461.82 | 404.81 | 529.89 |
| 32.9 | 10.8 | −2.9 (−26.9) | 27.1 (14.7) | ||
| Low | 1048.79 | 1216.11 | 1218.45 | 990.19 | 1184.01 |
| 16.0 | 16.2 | −5.6 (−18.6) | 12.9 (−2.8) | ||
| Total | 1606.52 | 1956.36 | 1814.22 | 1521.28 | 1948.09 |
| 21.8 | 12.9 | −5.3 (−22.2) | 21.3 (7.4) | ||
| Habitat Suitability | Current IUCN (km2) | 2050 1 | 2070 (%) 1, 2 | ||
|---|---|---|---|---|---|
| RCP 4.5 | RCP 8.5 | RCP 4.5 | RCP 8.5 | ||
| High | 121.39 | 139.73 | 92.41 | 111.89 | 173.13 |
| 15.1 | −23.9 | −7.8 (−19.9) | 42.6 (87.3) | ||
| Moderate | 248.09 | 305.20 | 253.31 | 250.25 | 289.86 |
| 23.0 | 2.1 | 0.9 (−18.0) | 16.8 (14.4) | ||
| Low | 396.40 | 422.30 | 437.45 | 433.38 | 396.28 |
| 6.5 | 10.4 | 9.3 (2.6) | 0.0 (−9.4) | ||
| Total | 765.88 | 867.22 | 783.18 | 795.52 | 859.27 |
| 13.2 | 2.3 | 3.9 (−8.3) | 12.2 (9.7) | ||
| Habitat suitability | Current (km2) | 2050 1 | 2070 (%) 1, 2 | ||
|---|---|---|---|---|---|
| RCP 4.5 | RCP 8.5 | RCP 4.5 | RCP 8.5 | ||
| High | 140.85 | 70.78 | 77.02 | 64.54 | 72.43 |
| −49.7 | −45.3 | −54.2 (−8.8) | −48.6 (−6.0) | ||
| Moderate | 416.88 | 112.77 | 108.38 | 108.30 | 121.63 |
| −72.9 | −74.0 | −74.0 (−4.0) | −70.8 (12.2) | ||
| Low | 1048.79 | 244.51 | 219.47 | 213.22 | 248.08 |
| −76.7 | −79.1 | −79.7 (−12.8) | −76.3 (13.0) | ||
| Total | 1606.52 | 428.06 | 404.87 | 386.06 | 442.14 |
| −73.4 | −74.8 | −76.0 (−9.8) | −72.5 (9.2) | ||
| NPA Modalities | IUCN | Current (km2) | 2050 1 | 2070 1 | ||
|---|---|---|---|---|---|---|
| RCP 4.5 | RCP 8.5 | RCP 4.5 | RCP 8.5 | |||
| National Park | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | |
| National Sanctuary | 82.12 | 6.39 | 52.92 | 5.73 | 1.87 | 35.62 |
| 5 (20.9) | 0.4 (1.6) | 2.7 (13.5) | 0.3 (1.5) | 0.1 (0.5) | 1.8 (9.1) | |
| Communal Reserve | 0.00 | 2.79 | 4.72 | 1.15 | 0.01 | 3.15 |
| 0 (0) | 0.2 (0.2) | 0.2 (0.4) | 0.1 (0.1) | 0 (0) | 0.2 (0.3) | |
| Alto Mayo protected forest | 568.20 | 418.86 | 447.51 | 517.98 | 423.09 | 567.15 |
| 34.9 (31.2) | 26.1 (23) | 22.9 (24.6) | 28.6 (28.5) | 27.8 (23.2) | 29.1 (31.2) | |
| Reserved Zones | 0.10 | 117.19 | 174.33 | 164.66 | 117.31 | 180.58 |
| 0 (0) | 7.3 (2.7) | 8.9 (4) | 9.1 (3.8) | 7.7 (2.7) | 9.3 (4.2) | |
| Regional Conservation Areas | 33.97 | 0.14 | 0.00 | 0.00 | 0.01 | 4.04 |
| 2.1 (0.8) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0.2 (0.1) | |
| Private Conservation Areas | 271.72 | 105.39 | 159.84 | 120.46 | 107.45 | 124.91 |
| 16.7 (17.3) | 6.6 (6.7) | 8.2 (10.2) | 6.6 (7.7) | 7.1 (6.8) | 6.4 (7.9) | |
| Total | 956.11 | 650.76 | 839.31 | 809.99 | 649.74 | 915.44 |
| 58.8 (3.1) | 40.5 (2.1) | 42.9 (2.8) | 44.6 (2.7) | 42.7 (2.1) | 47 (3) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meza Mori, G.; Rojas-Briceño, N.B.; Cotrina Sánchez, A.; Oliva-Cruz, M.; Olivera Tarifeño, C.M.; Hoyos Cerna, M.Y.; Ramos Sandoval, J.D.; Torres Guzmán, C. Potential Current and Future Distribution of the Long-Whiskered Owlet (Xenoglaux loweryi) in Amazonas and San Martin, NW Peru. Animals 2022, 12, 1794. https://doi.org/10.3390/ani12141794
Meza Mori G, Rojas-Briceño NB, Cotrina Sánchez A, Oliva-Cruz M, Olivera Tarifeño CM, Hoyos Cerna MY, Ramos Sandoval JD, Torres Guzmán C. Potential Current and Future Distribution of the Long-Whiskered Owlet (Xenoglaux loweryi) in Amazonas and San Martin, NW Peru. Animals. 2022; 12(14):1794. https://doi.org/10.3390/ani12141794
Chicago/Turabian StyleMeza Mori, Gerson, Nilton B. Rojas-Briceño, Alexander Cotrina Sánchez, Manuel Oliva-Cruz, Christian M. Olivera Tarifeño, Marlon Y. Hoyos Cerna, Jhonny D. Ramos Sandoval, and Cristóbal Torres Guzmán. 2022. "Potential Current and Future Distribution of the Long-Whiskered Owlet (Xenoglaux loweryi) in Amazonas and San Martin, NW Peru" Animals 12, no. 14: 1794. https://doi.org/10.3390/ani12141794