
Ovarian Follicular Dynamics and Its Functional Significance in Relation with Follicle Deviation, Vaginal Cytology, and Hormone Profiles in Llamas (Lama glama)
 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Experimental Animals
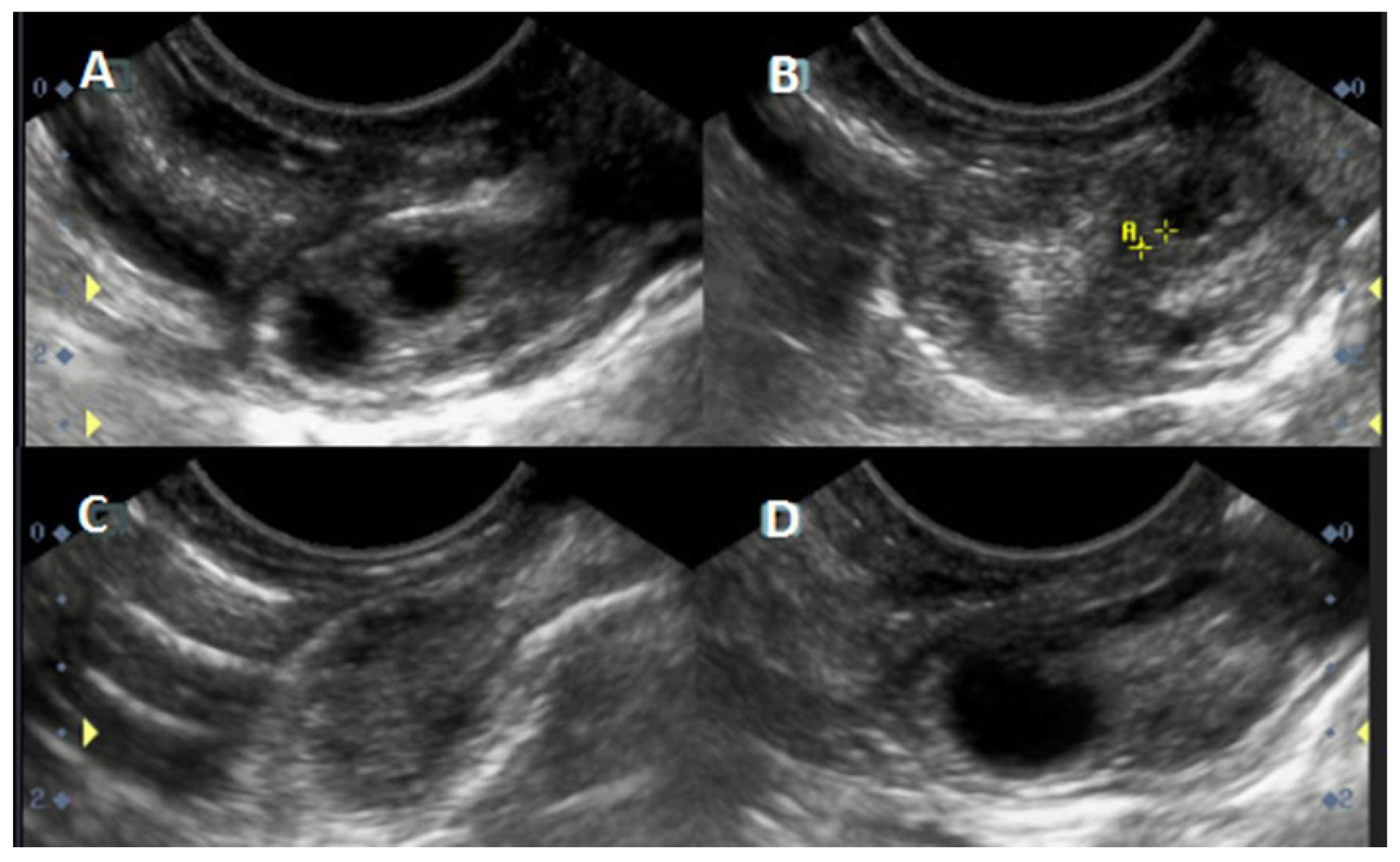
2.3. Preparation Protocol and Ultrasonographic Procedure
2.4. Follicular Ablation and Recruitment
2.5. Characterization of Follicular Deviation
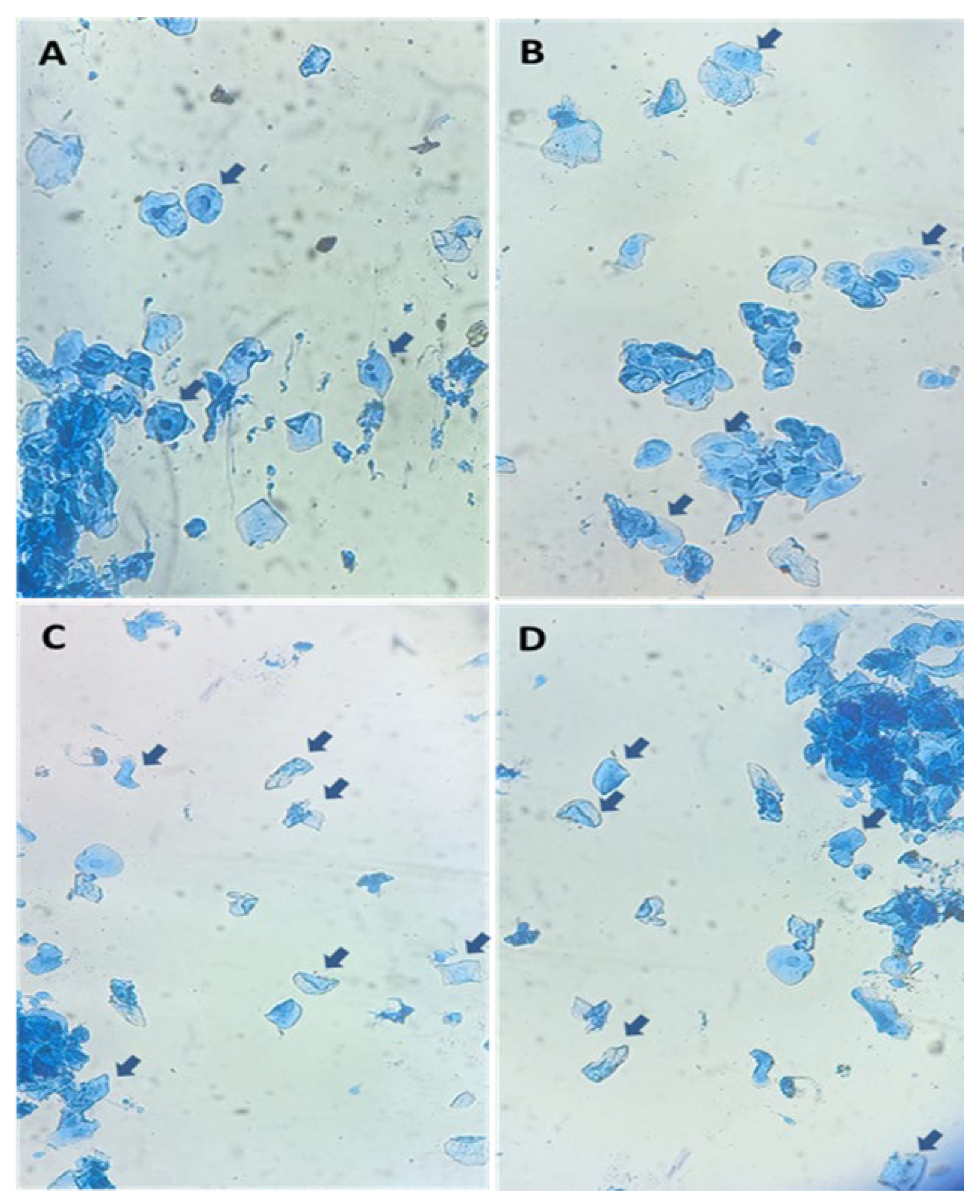
2.6. Vaginal Cytology Evaluation
2.7. Blood Sampling and Endocrine Profile Assessment by Enzyme-Linked Immunosorbent (ELISA) Assay
2.8. Statistical Analysis
3. Results
3.1. Follicular Wave: General Characteristics
3.2. Follicular Diameter (Single DF vs. Codominance): General Characteristics
3.3. Follicular Diameter (Single DF vs. Codominance) during Pre- and Post-Follicular Deviation
3.4. Follicular Growth Profiles (Single DF vs. Codominance) during Pre- and Post-Follicular Deviation
3.5. Vaginal Cytological Profiles and Follicular Dynamics
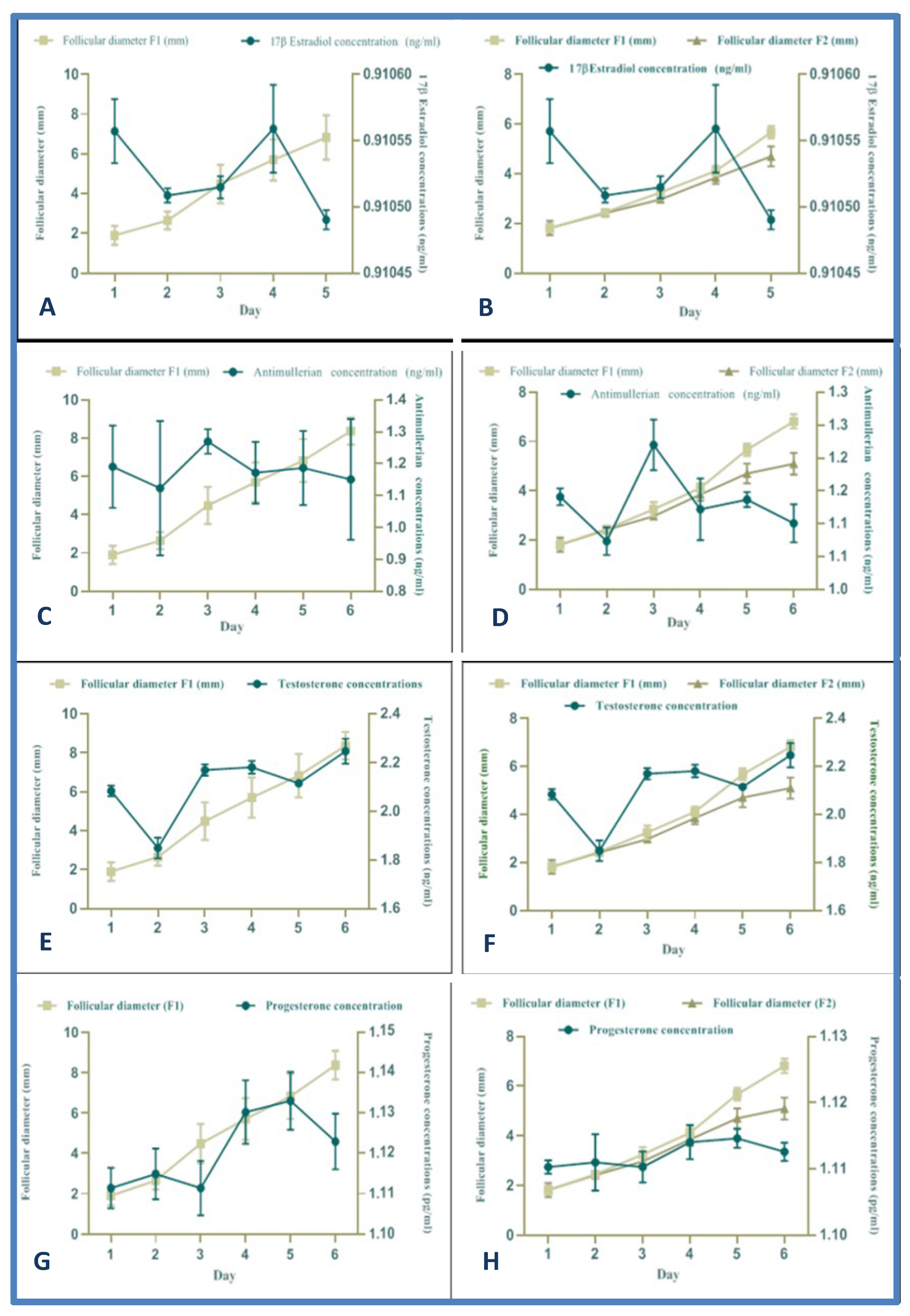
3.6. Chronological Hormone Levels during the Follicular Deviation (Single DF vs. Codominance)
3.7. Hormone Profiles during the Follicular Deviation Process
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Adams, G.P.; Sumar, J.; Ginther, O.J. Effects of Lactational and Reproductive Status on Ovarian Follicular Waves in Llamas (Lama glama). J. Reprod. Fertil. 1990, 90, 535–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaughan, J. Ovarian Function in South American Camelids (Alpacas, Llamas, Vicunas, Guanacos). Anim. Reprod. Sci. 2011, 124, 237–243. [Google Scholar] [CrossRef]
- Cavilla, M.V.; Bianchi, C.P.; Maistruarena, C.; Aba, M.A. Ultrasonographic and Endocrine Characterization of Follicular Waves in Llamas with a Special Reference to the Overlapping Phenomenon during Successive Waves. Reprod. Domest. Anim. 2013, 48, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, J.L.; Macmillan, K.L.; D’Occhio, M.J. Ovarian Follicular Wave Characteristics in Alpacas. Anim. Reprod. Sci. 2004, 80, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Gallelli, M.F.; Bianchi, C.; Zampini, E.; Aba, M.; Gambarotta, M.; Miragaya, M. Plasma IGF1 and 17β-Estradiol Concentrations During the Follicular Wave in Llamas. Front. Vet. Sci. 2020, 7, 555261. [Google Scholar] [CrossRef]
- Ginther, O.J.; Beg, M.A.; Donadeu, F.X.; Bergfelt, D.R. Mechanism of Follicle Deviation in Monovular Farm Species. Anim. Reprod. Sci. 2003, 78, 239–257. [Google Scholar] [CrossRef]
- Gimenes, L.U.; Carvalho, N.A.T.; Sá Filho, M.F.; Vannucci, F.S.; Torres-Júnior, J.R.S.; Ayres, H.; Ferreira, R.M.; Trinca, L.A.; Sartorelli, E.S.; Barros, C.M.; et al. Ultrasonographic and Endocrine Aspects of Follicle Deviation, and Acquisition of Ovulatory Capacity in Buffalo (Bubalus bubalis) Heifers. Anim. Reprod. Sci. 2011, 123, 175–179. [Google Scholar] [CrossRef] [Green Version]
- Ginther, O.J. Switching of Follicle Destiny so That the Second Largest Follicle Becomes Dominant in Monovulatory Species. Theriogenology 2021, 171, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Manjunatha, B.M.; Al-Bulushi, S.; Pratap, N. Ultrasonographic Characterization of Follicle Deviation in Follicular Waves with Single Dominant and Codominant Follicles in Dromedary Camels (Camelus dromedarius). Reprod. Domest. Anim. 2014, 49, 239–242. [Google Scholar] [CrossRef]
- Vaughan, J.L.; Tibary, A. Reproduction in Female South American Camelids: A Review and Clinical Observations. Small Rumin. Res. 2006, 61, 259–281. [Google Scholar] [CrossRef]
- Tibary, A. Monitoring and Controlling Follicular Activity in Camelids. Theriogenology 2018, 109, 22–30. [Google Scholar] [CrossRef]
- Tibary, A.; Anouassi, A.; Khatir, H. Update on Reproductive Biotechnologies in Small Ruminants and Camelids. Theriogenology 2005, 64, 618–638. [Google Scholar] [CrossRef]
- Ratto, M.H.; Silva, M.E.; Huanca, W.; Huanca, T.; Adams, G.P. Induction of Superovulation in South American Camelids. Anim Reprod Sci 2013, 136, 164–169. [Google Scholar] [CrossRef]
- Sumar, J.B. Embryo Transfer in Domestic South American Camelids. Anim. Reprod. Sci. 2013, 136, 170–177. [Google Scholar] [CrossRef]
- Adams, G.P.; Ratto, M.H.; Carrasco, R.A. Natural and Controlled Ovulation in South American Camelids. Anim. Reprod. 2018, 15, 996–1002. [Google Scholar] [CrossRef]
- Siregar, T.N.; Melia, J.; Rohaya; Thasmi, C.N.; Masyitha, D.; Wahyuni, S.; Rosa, J.; Nurhafni; Panjaitan, B.; Herrialfian. Determining Proportion of Exfoliative Vaginal Cell during Various Stages of Estrus Cycle Using Vaginal Cytology Techniques in Aceh Cattle. Vet. Med. Int. 2016, 2016, 3976125. [Google Scholar] [CrossRef] [Green Version]
- England, G.C.W.; Friedrichs, K.R. Cytology of the Male and Female Genital Tracts. In Manual of Diagnostic Cytology of the Dog and Cat; Wiley-Blackwell: West Sussex, UK, 2014; pp. 213–229. [Google Scholar] [CrossRef]
- Haji, M.; Ahmed, F.; Lalrintluanga, K.; Talukdar, D.; Doley, P.; Behera, S.; Sarma, K. The Role of Estrogen and Progesterone Hormone on Vaginal Cytology in Bitch. Int. J. Livest. Res. 2018, 8, 241. [Google Scholar] [CrossRef]
- Sharma, M.; Sharma, N. Vaginal Cytology: An Historical Perspective on Its Diagnostic Use. Adv. Anim. Vet. Sci. 2016, 4, 283–288. [Google Scholar] [CrossRef] [Green Version]
- dos Santos, A.C.; Viana, D.C.; Oliveira, G.B.; Silva, R.S.; Oliveira, M.F.; Assis-Neto, A.C. Follicular Development and Morphological Changes in the Vaginal Epithelium during the Estrous Cycle of Galea Spixii. Microsc. Res. Tech. 2017, 80, 167–176. [Google Scholar] [CrossRef]
- Bravo, P.W.; Fowler, M.E.; Stabenfeldt, G.H.; Lasley, B.L. Ovarian Follicular Dynamics in the Llama. Biol. Reprod. 1990, 43, 579–585. [Google Scholar] [CrossRef] [Green Version]
- Miragaya, M.H.; Aba, M.A.; Capdevielle, E.F.; Ferrer, M.S.; Chaves, M.G.; Rutter, B.; Agüero, A. Follicular Activity and Hormonal Secretory Profile in Vicuna (Vicugna vicugna). Theriogenology 2004, 61, 663–671. [Google Scholar] [CrossRef]
- Manjunatha, B.M.; Pratap, N.; Al-Bulushi, S.; Hago, B.E. Characterization of Ovarian Follicular Dynamics in Dromedary Camels (Camelus dromedarius). Theriogenology 2012, 78, 965–973. [Google Scholar] [CrossRef]
- Ratto, M.H.; Singh, J.; Huanca, W.; Adams, G.P. Ovarian Follicular Wave Synchronization and Pregnancy Rate after Fixed-Time Natural Mating in Llamas. Theriogenology 2003, 60, 1645–1656. [Google Scholar] [CrossRef]
- Adams, G.P.; Ratto, M.H.; Huanca, W.; Singh, J. Ovulation-Inducing Factor in the Seminal Plasma of Alpacas and Llamas. Biol. Reprod. 2005, 73, 452–457. [Google Scholar] [CrossRef] [Green Version]
- Bedenk, J.; Vrtačnik-Bokal, E.; Virant-Klun, I. The Role of Anti-Müllerian Hormone (AMH) in Ovarian Disease and Infertility. J. Assist. Reprod. Genet. 2020, 37, 89–100. [Google Scholar] [CrossRef]
- Van Saun, R.J. Nutritional Requirements and Assessing Nutritional Status in Camelids. Vet. Clin. N. Am.-Food Anim. Pract. 2009, 25, 265–279. [Google Scholar] [CrossRef] [PubMed]
- Ratto, M.; Gomez, C.; Berland, M.; Adams, G.P. Effect of Ovarian Superstimulation on COC Collection and Maturation in Alpacas. Anim. Reprod. Sci. 2007, 97, 246–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huanca, W.F.; Hilari, F.Y.; Villanueva, J.C.; Uchuari, M.; Huanca, W.; Huanca, W.F.; Hilari, F.Y.; Villanueva, J.C.; Uchuari, M.; Huanca, W. 100 Use of Seminal Plasma, Human Chorionic Gonadotropin, and Follicular Ablation on the Interval to Follicular Wave Emergency and Dominant Follicle in Alpacas (Vicugna pacos). Reprod. Fertil. Dev. 2018, 30, 189–190. [Google Scholar] [CrossRef]
- Silva, N.H.; Huanca, W.F.M.; Medina, G.S.; Huanca, W.L. Application Effect of Seminal Plasma, GnRH and Follicular Ablation on the Ovarian Dynamic in Llamas (Lama glama). Rev. Investig. Vet. Peru 2020, 31, e19033. [Google Scholar] [CrossRef]
- Tilaye, D.; Tefera, Y.; Tamrat, D.; Gemechu, W.; Alemayehu, L. Effect of Follicular Ablation and Gonadotropin Priming on the Recovery and Quality of Oocytes in Boran Cows. Int. J. Vet. Sci. Res. 2021, 7, 138–143. [Google Scholar] [CrossRef]
- Goodman, A.L.; Hodgen, G.D. The Ovarian Triad of the Primate Menstrual Cycle. Recent Prog. Horm. Res. 1983, 39, 1–73. [Google Scholar] [CrossRef]
- Jill Colloton, G.G. Practical Atlas of Ruminant and Camelid Reproductive Ultrasonography; Wiley-Blackwell: Ames, IA, USA, 2009. [Google Scholar] [CrossRef]
- Feder, F.H.; González, H.; Arias, P. Comparative Histological Study of the Female Reproductive System in the Llama (Lama guanicoe glama). II. Oviduct, Uterus, Cervix, Vagina. Anat. Histol. Embryol. 1999, 28, 139–142. [Google Scholar] [CrossRef]
- Montes, G.S.; Luque, E.H. Effects of Ovarian Steroids on Vaginal Smears in the Rat. Acta Anat. 1988, 133, 192–199. [Google Scholar] [CrossRef] [PubMed]
- R Core Team—European Environment Agency. 2020. Available online: https://www.eea.europa.eu/data-and-maps/indicators/oxygen-consuming-substances-in-rivers/r-development-core-team-2006 (accessed on 18 October 2022).
- Riveros, J.L.; Schuler, G.; Bonacic, C.; Hoffmann, B.; Chaves, M.G.; Urquieta, B. Ovarian Follicular Dynamics and Hormonal Secretory Profiles in Guanacos (Lama guanicoe). Anim. Reprod. Sci. 2010, 119, 63–67. [Google Scholar] [CrossRef]
- Wiltbank, M.C.; Fricke, P.M.; Sangsritavong, S.; Sartori, R.; Ginther, O.J. Mechanisms That Prevent and Produce Double Ovulations in Dairy Cattle. J. Dairy Sci. 2000, 83, 2998–3007. [Google Scholar] [CrossRef]
- Kulick, L.J.; Bergfelt, D.R.; Kot, K.; Ginther, O.J. Follicle Selection in Cattle: Follicle Deviation and Codominance within Sequential Waves. Biol. Reprod. 2001, 65, 839–846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ginther, O.J.; Bergfelt, D.R.; Kulick, L.J.; Kot, K. Selection of the Dominant Follicle in Cattle: Role of Two-Way Functional Coupling between Follicle-Stimulating Hormone and the Follicles. Biol. Reprod. 2000, 62, 920–927. [Google Scholar] [CrossRef] [Green Version]
- Sartori, R.; Fricke, P.M.; Ferreira, J.C.P.; Ginther, O.J.; Wiltbank, M.C. Follicular Deviation and Acquisition of Ovulatory Capacity in Bovine Follicles. Biol. Reprod. 2001, 65, 1403–1409. [Google Scholar] [CrossRef]
- Ginther, O.J.; Dangudubiyyam, S.V. Effect of Number of 6-Mm Predeviation Follicles and Intraovarian Patterns on Right-Side Ovulation in Heifers. Anim. Reprod. Sci. 2019, 202, 73–79. [Google Scholar] [CrossRef]
- Joel Pacheco, C. Caracterización de La Citología Exfoliativa Vaginal En Alpacas (Vicugna pacos). Rev. Investig. Vet. Perú 2017, 28, 886–893. [Google Scholar] [CrossRef] [Green Version]
- Majama, Y.B.; Mshelia, G.D.; Ahmed, Y.; Abba, Y.; Kwari, H.D. Exfoliative Cytology of the Uterus and Vagina during the Follicular and Luteal Phases in One-Humped Camel (Camelus dromedarius). Sokoto J. Vet. Sci. 2018, 16, 33. [Google Scholar] [CrossRef] [Green Version]
- Chaves, M.G.; Aba, M.; Agüero, A.; Egey, J.; Berestin, V.; Rutter, B. Ovarian Follicular Wave Pattern and the Effect of Exogenous Progesterone on Follicular Activity in Non-Mated Llamas. Anim. Reprod. Sci. 2002, 69, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Cancino, A.K.; Rebuffi, G.; Alberio, Y. Control of Ovarian Follicular Dynamics in Early Postpartum Llama (Lama glama). Arch. Zootec 2011, 60, 23–32. [Google Scholar] [CrossRef] [Green Version]
- Perez, G.U.; Pari, P.D.; Gutierrez, Y.F.; Málaga, A.J.; Luque, M.N.; Rojas, E.R.; Pérez, D.M.; Perez, G.U.; Pari, P.D.; Gutierrez, Y.F.; et al. Comparación Ultrasonográfica Transvaginal y Transrectal de La Dinámica Folicular En Ondas Sucesivas de Llamas (Lama glama). Rev. Investig. Vet. Perú 2021, 32, e19504. [Google Scholar] [CrossRef]
- Kanca, H.; Karakas, K.; Dalgic, M.A.; Salar, S.; Izgur, H. Vaginal Cytology after Induction of Ovulation in the Queen: Comparison of Postoestrus and Dioestrus. Aust. Vet. J. 2014, 92, 65–70. [Google Scholar] [CrossRef]
- Post, K. Canine Vaginal Cytology during the Estrous Cycle. Can. Vet. J. 1985, 26, 101–104. [Google Scholar]
- Reckers, F.; Klopfleisch, R.; Belik, V.; Arlt, S. Canine Vaginal Cytology: A Revised Definition of Exfoliated Vaginal Cells. Front. Vet. Sci. 2022, 9, 834031. [Google Scholar] [CrossRef]
- Tennant, K. General Principles of Cytological Interpretation. In Manual of Diagnostic Cytology of the Dog and Cat; Wiley-Blackwell: West Sussex, UK, 2014; pp. 17–32. [Google Scholar] [CrossRef]
- Nelson, L.R.; Bulun, S.E. Estrogen Production and Action. J. Am. Acad. Dermatol. 2001, 45, S116–S124. [Google Scholar] [CrossRef]
- Díaz, P.U.; Rey, F.; Gareis, N.C.; Notaro, U.S.; Matiller, V.; Belotti, E.M.; Stassi, A.F.; Salvetti, N.R.; Ortega, H.H. Altered Expression of Anti-Müllerian Hormone during the Early Stage of Bovine Persistent Ovarian Follicles. J. Comp. Pathol. 2018, 158, 22–31. [Google Scholar] [CrossRef]
- DI Clemente, N.; Racine, C.; Pierre, A.; Taieb, J. Anti-Müllerian Hormone in Female Reproduction. Endocr. Rev. 2021, 42, 753–782. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Day | Follicular Diameter (Single DF) | Follicular Diameter (Codominance) | |
|---|---|---|---|
| F1 | F2 | ||
| 1 | 1.90 ± 0.28 a | 1.82 ± 0.17 a | 1.82 ± 0.28 a |
| 2 | 2.65 ± 0.05 a | 2.45 ± 0.03 a | 2.42 ± 0.05 a |
| 3 | 4.49 ± 0.11 ab | 3.25 ± 0.28 ab | 2.97 ± 0.11 ab |
| 4 | 5.70 ± 0.25 b | 4.12 ± 0.2 ab | 3.85 ± 0.25 ab |
| 5 | 6.83 ± 0.40 b | 5.67 ± 0.25 b | 4.70 ± 0.40 ab |
| 6 | 8.37 ± 0.44 b | 6.82 ± 0.28 b | 5.10 ± 0.44 b |
| Follicular Structure | Single Dominant Follicle | Codominance | ||
|---|---|---|---|---|
| Pre-Deviation | Post-Deviation (Day 5) | Pre-Deviation | Post-Deviation (Day 5) | |
| F1 | 2.21 ± 0.89 | 6.01 ± 0.73 a | 2.65 ± 0.93 | 6.65 ± 2.61 a |
| F2 | 2.24 ± 0.89 | 2.05 ± 0.52 b | 2.25 ± 0.67 | 5.59 ± 2.17 a |
| F3 | 2.37 ± 0.66 | 2.93 ± 1.02 b | ||
| F4 | 2.32 ± 0.57 | 2.84 ± 0.91 b | ||
| p value | 0.9339 | ˂0.0001 | 0.635 | ˂0.000001 |
| Follicular Structure | Single Dominant Follicle | Codominance | ||
|---|---|---|---|---|
| Pre-Deviation | Post-Deviation (Day 5) | Pre-Deviation | Post-Deviation (Day 5) | |
| F1 | 0.66 ± 0.37 | 0.65 ± 0.31 a | 0.52 ± 0.42 | 0.58 ± 0.34 a |
| F2 | 0.44 ± 0.32 | −0.01 ± 0.29 b | 0.36 ± 0.35 | 0.64 ± 0.67 a |
| F3 | 0.35 ± 0.35 | −0.22 ± 0.45 b | ||
| F4 | 0.24 ± 0.39 | −0.24 ± 0.49 b | ||
| p value | 0.05 | ˂0.000001 | 0.286 | ˂0.000001 |
| Follicular Phase | Single Dominant Follicle | Codominance | ||||
|---|---|---|---|---|---|---|
| Cell Type | ||||||
| Parabasal (%) | Intermediate (%) | Superficial (%) | Parabasal (%) | Intermediate (%) | Superficial (%) | |
| Growth (~7 mm) | 20.12 ± 1.93 a | 42.06 ± 6.37 b | 41.25 ± 3.21 b | 24.12 ± 2.81 a | 39.06 ± 8.22 b | 42.25 ± 5.11 b |
| Dominance (~12 mm) | 20.33 ± 1.09 a | 41.99 ± 7.12 b | 43.00 ± 4.96 b | 23.33 ± 2.00 a | 37.99 ± 9.06 b | 45.00 ± 8.36 b |
| Regression (~10 mm) | 18.02 ± 0.90 a | 44.89 ± 8.03 b | 39.11 ± 6.10 b | 23.02 ± 1.92 a | 38.89 ± 8.53 b | 40.11 ± 7.64 b |
| Day | Hormone Concentration | |||||||
|---|---|---|---|---|---|---|---|---|
| Testosterone (ng/mL) | Anti-Mullerian (ng/mL) | 17 β Estradiol (ng/mL) | Progesterone (pg/mL) | |||||
| Single DF | Codominance | Single DF | Codominance | Single DF | Codominance | Single DF | Codominance | |
| 1 | 2.080 ± 0.020 a | 2.078 ± 0.020 a | 1.210 ± 0.010 | 1.190 ± 0.020 | 0.9105 ± 0.000 | 0.9105 ± 0.000 | 1.110 ± 0.001 | 1.110 ± 0.001 |
| 2 | 1.848 ± 0.040 b | 1.845 ± 0.040 b | 1.110 ± 0.020 | 1.120 ± 0.010 | 0.9104 ± 0.000 | 0.9105 ± 0.000 | 1.119 ± 0.001 | 1.111 ± 0.001 |
| 3 | 2.156 ± 0.020 a | 2.183 ± 0.030 a | 1.260 ± 0.040 | 1.290 ± 0.030 | 0.9105 ± 0.000 | 0.9105 ± 0.000 | 1.110 ± 0.001 | 1.110 ± 0.001 |
| 4 | 2.175 ± 0.020 a | 2.182 ± 0.020 a | 1.190 ± 0.050 | 1.180 ± 0.030 | 0.9105 ± 0.000 | 0.9105 ± 0.000 | 1.113 ± 0.001 | 1.114 ± 0.001 |
| 5 | 2.115 ± 0.020 a | 2.117 ± 0.010 a | 1.185 ± 0.010 | 1.194 ± 0.030 | 0.9104 ± 0.000 | 0.9105 ± 0.000 | 1.114 ± 0.001 | 1.115 ± 0.001 |
| 6 | 2.256 ± 0.050 c | 2.211 ± 0.020 c | 1.150 ± 0.030 | 1.150 ± 0.010 | 0.9105 ± 0.000 | 0.9105 ± 0.000 | 1.112 ± 0.001 | 1.112 ± 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perez-Guerra, U.H.; Quispe, Y.M.; Gonzáles, H.I.; Luque, N.; Ruelas, D.A.; Carretero, M.I.; Gutiérrez-Reinoso, M.A.; Pérez-Durand, M.G.; García-Herreros, M. Ovarian Follicular Dynamics and Its Functional Significance in Relation with Follicle Deviation, Vaginal Cytology, and Hormone Profiles in Llamas (Lama glama). Animals 2022, 12, 3299. https://doi.org/10.3390/ani12233299
Perez-Guerra UH, Quispe YM, Gonzáles HI, Luque N, Ruelas DA, Carretero MI, Gutiérrez-Reinoso MA, Pérez-Durand MG, García-Herreros M. Ovarian Follicular Dynamics and Its Functional Significance in Relation with Follicle Deviation, Vaginal Cytology, and Hormone Profiles in Llamas (Lama glama). Animals. 2022; 12(23):3299. https://doi.org/10.3390/ani12233299
Chicago/Turabian StylePerez-Guerra, Uri H., Yesenia M. Quispe, Henry I. Gonzáles, Natalio Luque, Domingo A. Ruelas, María I. Carretero, Miguel A. Gutiérrez-Reinoso, Manuel G. Pérez-Durand, and Manuel García-Herreros. 2022. "Ovarian Follicular Dynamics and Its Functional Significance in Relation with Follicle Deviation, Vaginal Cytology, and Hormone Profiles in Llamas (Lama glama)" Animals 12, no. 23: 3299. https://doi.org/10.3390/ani12233299