
Optimisation and Characterisation of the Protein Hydrolysate of Scallops (Argopecten purpuratus) Visceral By-Products
 ,
,  , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Raw Materials
2.2. Experimental Design for Optimisation of the Degree of Hydrolysis
2.2.1. Enzymatic Hydrolysis Reaction
2.2.2. Design of Experiment (DOE)
2.2.3. Degree of hydrolysis
2.3. Protein Hydrolysate
2.4. Proximal Composition
2.5. Determination of Amino Acid Composition by High-Performance Liquid Chromatography (HPLC)
2.6. Protein Solubility Curve
2.7. Molecular Weight Profile by Exclusion Chromatography (SEC)
2.8. Statistical Analysis
3. Results and Discussions
3.1. Proximal Composition of Scallops Visceral Meal
3.2. Amino Acid Profile of Scallops Visceral Meal
3.3. Protein Solubility Curve of Scallops Visceral Meal
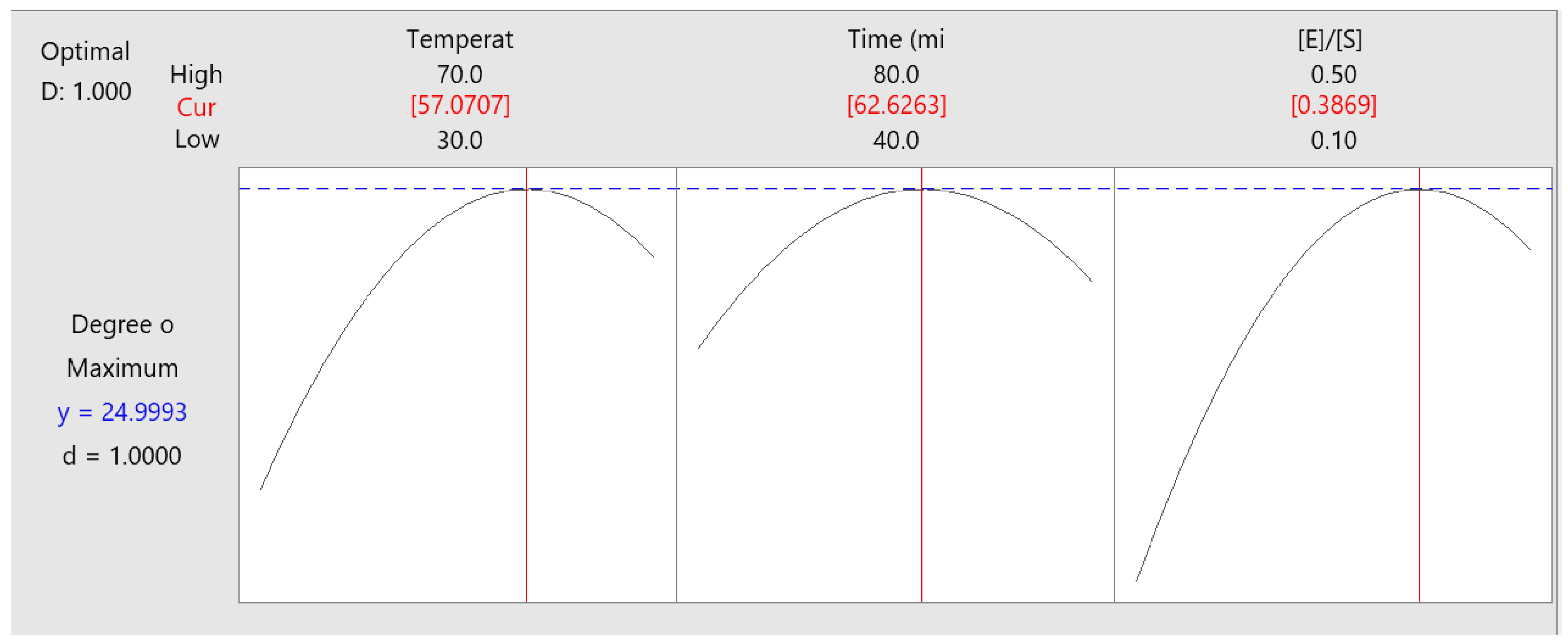
3.4. Optimisation of Enzymatic Hydrolysis
3.5. Characterisation of Protein Hydrolysates
3.6. Amino Acid Profile of Protein Hydrolysates
3.7. Protein Solubility Curve of Hydrolysates
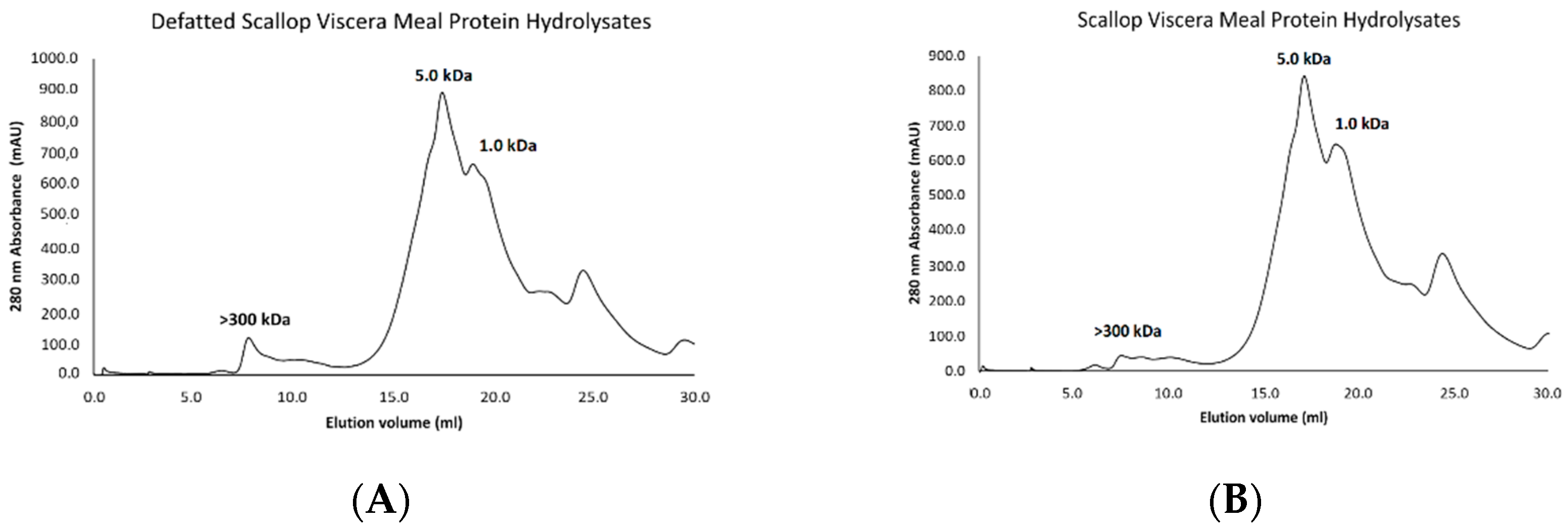
3.8. Molecular Weight Profile of Protein Hydrolysates
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kluger, L.C.; Taylor, M.H.; Wolff, M.; Stotz, W.; Mendo, J.; Lotta Kluger, C.C. From an Open-Access Fishery to a Regulated Aquaculture Business: The Case of the Most Important Latin American Bay Scallop (Argopecten Purpuratus). Rev. Aquac. 2018, 11, 187–203. [Google Scholar] [CrossRef]
- Mendo, J.; Quevedo, I. Estudio de Prospectiva: La Concha de Abanico; Programa Nacional de Innovación en Pesca y Acuicultura: Miraflores, Peru, 2020; p. 137. [Google Scholar]
- Valenzuela, A.; Yánez, C.G.; Golusda, C. EL OSTIÓN DEL NORTE CHILENO (Argopecten Purpuratus), UN ALIMENTO DE ALTO VALOR NUTRICIONAL. Rev. Chil. De Nutr. 2011, 38, 148–155. [Google Scholar] [CrossRef]
- Universidad de Lima De Residuo de Concha de Abanico a Alimento Nutrifuncional. Available online: https://www.ulima.edu.pe/instituto-de-investigacion-cientifica/noticias/de-residuo-de-concha-de-abanico-alimento (accessed on 3 May 2022).
- Colán-Ramos, C.; Gómez-Sánchez, M.; Alcazar-Zamora, J.A.; Aguirre-Velarde, A. Aprovechamiento de Los Residuos Blandos de Concha de Abanico, Argopecten Purpuratus (Lamarck, 1819), Para Producir Harina de Alto Contenido Proteico. Rev. De Investig. Vet. Del Perú 2019, 30, 961–966. [Google Scholar] [CrossRef]
- Shahidi, F.; Ambigaipalan, P. Bioactives From Seafood Processing By-Products. In Encyclopedia of Food Chemistry; Elsevier: Amsterdam, The Netherlands, 2018; pp. 280–288. ISBN 9780128140451. [Google Scholar]
- Gao, R.; Yu, Q.; Shen, Y.; Chu, Q.; Chen, G.; Fen, S.; Yang, M.; Yuan, L.; McClements, D.J.; Sun, Q. Production, Bioactive Properties, and Potential Applications of Fish Protein Hydrolysates: Developments and Challenges. Trends Food Sci. Technol. 2021, 110, 687–699. [Google Scholar] [CrossRef]
- Eghianruwa, Q.A.; Osoniyi, O.R.; Maina, N.; Wachira, S. Bioactive Peptides from Marine Molluscs–A Review. Int. J. Biochem. Res. Rev. 2019, 27, 1–12. [Google Scholar] [CrossRef]
- Paul, A.A.; Eghianruwa, Q.A.; Oparinde, O.G.; Adesina, A.S.; Osoniyi, O. Enzymatic Protein Hydrolysates, and Ultrafiltered Peptide Fractions from Two Molluscs: Tympanotonus fuscatus Var. Radula (L.) and Pachymelania aurita (M.), with Angiotensin-I-Converting Enzyme Inhibitory and DPPH Radical Scavenging Activities. Int. J. Appl. Basic. Med. Res. 2021, 11, 70. [Google Scholar] [CrossRef]
- Jayaprakash, R.; Perera, C.O. Partial Purification and Characterization of Bioactive Peptides from Cooked New Zealand Green-Lipped Mussel (Perna Canaliculus) Protein Hydrolyzates. Foods 2020, 9, 879. [Google Scholar] [CrossRef]
- Chi, C.F.; Hu, F.Y.; Wang, B.; Li, T.; Ding, G.F. Antioxidant and Anticancer Peptides from the Protein Hydrolysate of Blood Clam (Tegillarca Granosa) Muscle. J. Funct. Foods 2015, 15, 301–313. [Google Scholar] [CrossRef]
- Wu, D.; Wu, C.; Tu, M.; Yu, C.; Du, M. Identification and Analysis of Bioactive Peptides from Scallops (Chlamys Farreri) Protein by Simulated Gastrointestinal Digestion. J. Food Process. Preserv. 2018, 42, e13760. [Google Scholar] [CrossRef]
- Petrova, I.; Tolstorebrov, T.; Magne, E. Production of Fish Protein Hydrolysates Step by Step: Technological Aspects, Equipment Used, Major Energy Costs and Methods of Their Minimizing. Int. Aquat. Res. 2018, 10, 223–241. [Google Scholar] [CrossRef]
- Gao, R.; Shen, Y.; Shu, W.; Bai, F.; Jin, W.; Wang, J.; Yuan, L. Optimization of Enzymatic Conditions of Sturgeon Muscles and Their Anti-Inflammatory Potential. J. Food Qual. 2020, 2020, 9698134. [Google Scholar] [CrossRef]
- Bhaskar, N.; Mahendrakar, N.S. Protein Hydrolysate from Visceral Waste Proteins of Catla (Catla Catla): Optimization of Hydrolysis Conditions for a Commercial Neutral Protease. Bioresour. Technol. 2008, 99, 4105–4111. [Google Scholar] [CrossRef] [PubMed]
- Bui, X.D.; Vo, C.T.; Bui, V.C.; Pham, T.M.; Bui, T.T.H.; Nguyen-Sy, T.; Nguyen, T.D.P.; Chew, K.W.; Mukatova, M.D.; Show, P.L. Optimization of Production Parameters of Fish Protein Hydrolysate from Sarda Orientalis Black Muscle (by-Product) Using Protease Enzyme. Clean Technol. Environ. Policy 2020, 23, 31–40. [Google Scholar] [CrossRef]
- Korkmaz, K.; Tokur, B. Optimization of Hydrolysis Conditions for the Production of Protein Hydrolysates from Fish Wastes Using Response Surface Methodology. Food Biosci. 2022, 45, 101312. [Google Scholar] [CrossRef]
- Taoufik, N.; Elmchaouri, A.; El Mahmoudi, S.; Korili, S.A.; Gil, A. Comparative Analysis Study by Response Surface Methodology and Artificial Neural Network on Salicylic Acid Adsorption Optimization Using Activated Carbon. Env. Nanotechnol. Monit. Manag. 2021, 15, 100448. [Google Scholar] [CrossRef]
- Millán-Linares, M.D.C.; Yust, M.D.M.; Alcaide-Hidalgo, J.M.; Millán, F.; Pedroche, J. Lupine Protein Hydrolysates Inhibit Enzymes Involved in the Inflammatory Pathway. Food Chem. 2014, 151, 141–147. [Google Scholar] [CrossRef]
- Box, G.E.P.; Behnken, D.W. Some New Three Level Designs for the Study of Quantitative Variables. Technometrics 1960, 2, 455–475. [Google Scholar] [CrossRef]
- Adler-Nissen, J. Determination of the Degree of Hydrolysis of Food Protein Hydrolysates by Trinitrobenzenesulfonic Acid. J Agric. Food. Chem. 1979, 27, 1256–1262. [Google Scholar] [CrossRef]
- Alaiz, M.; Navarro, J.L.; Girón, J.; Vioque, E. Amino Acid Analysis by High-Performance Liquid Chromatography after Derivatization with Diethyl Ethoxymethylenemalonate. J Chromatogr. A 1992, 591, 181–186. [Google Scholar] [CrossRef]
- Yust, M.M.; Pedroche, J.; Girón-Calle, J.; Vioque, J.; Millán, F.; Alaiz, M. Determination of Tryptophan by High-Performance Liquid Chromatography of Alkaline Hydrolysates with Spectrophotometric Detection. Food Chem. 2004, 85, 317–320. [Google Scholar] [CrossRef]
- la Paz, S.M.D.; Martinez-Lopez, A.; Villanueva-Lazo, A.; Pedroche, J.; Millan, F.; Millan-Linares, M.C. Identification and Characterization of Novel Antioxidant Protein Hydrolysates from Kiwicha (Amaranthus caudatus L.). Antioxidants 2021, 10, 645. [Google Scholar] [CrossRef]
- Benitez-Hernández, A.; Jiménez-Bárcenas, S.P.L.; Sánchez-Gutiérrez, E.Y.; Pérez-Urbiola, J.C.; Tovar-Ramírez, D.; Palacios, E.; Civera-Cerecedo, R. Use of Marine By-Product Meals in Diets for Juvenile Longfin Yellowtail Seriola Rivoliana. Aquac. Nutr. 2018, 24, 562–570. [Google Scholar] [CrossRef]
- Lúquez-Pérez, L.D.R.; Hleap-Zapata, J.I. Viabilidad Del Uso de Harina de Residuos Pesqueros de La Ciénaga de Zapatosa En La Alimentación de Pollos de Engorde. Rev. U.D.C.A Actual. Divulg. Científica 2020, 23. [Google Scholar] [CrossRef]
- Tabakaeva, O.V.; Tabakaev, A.V.; Piekoszewski, W. Nutritional Composition and Total Collagen Content of Two Commercially Important Edible Bivalve Molluscs from the Sea of Japan Coast. J. Food Sci. Technol. 2018, 55, 4877–4886. [Google Scholar] [CrossRef]
- Xing, R.E.; Yang, H.Y.; Wang, X.Q.; Yu, H.H.; Liu, S.; Chen, X.L.; Li, P.C. Effect of Enzymatically Hydrolyzed Scallop Visceral Protein Powder Used as a Replacement of Fish Meal on the Growth Performance, Immune Responses, Intestinal Microbiota and Intestinal Morphology of Broiler Chickens. Livest. Sci. 2018, 207, 15–24. [Google Scholar] [CrossRef]
- Shoveller, A.K.; Stoll, B.; Ball, R.O.; Burrin, D.G. Nutritional and Functional Importance of Intestinal Sulfur Amino Acid Metabolism. J. Nutr. 2005, 135, 1609–1612. [Google Scholar] [CrossRef]
- Surasani, V.K.R. Acid and Alkaline Solubilization (PH Shift) Process: A Better Approach for the Utilization of Fish Processing Waste and by-Products. Environ. Sci. Pollut. Res. 2018, 25, 18345–18363. [Google Scholar] [CrossRef]
- Abdollahi, M.; Undeland, I. Structural, Functional, and Sensorial Properties of Protein Isolate Produced from Salmon, Cod, and Herring by-Products. Food Bioprocess Technol. 2018, 11, 1733–1749. [Google Scholar] [CrossRef]
- Mahdabi, M.; Hosseini Shekarabi, S.P. A Comparative Study on Some Functional and Antioxidant Properties of Kilka Meat, Fishmeal, and Stickwater Protein Hydrolysates. J. Aquat. Food Prod. Technol. 2018, 27, 844–858. [Google Scholar] [CrossRef]
- Sathe, S.K.; Zaffran, V.D.; Gupta, S.; Li, T. Protein Solubilization. J. Am. Oil Chem. Soc. 2018, 95, 883–901. [Google Scholar] [CrossRef]
- Henriques, A.; Vázquez, J.A.; Valcarcel, J.; Mendes, R.; Bandarra, N.M.; Pires, C. Characterization of Protein Hydrolysates from Fish Discards and By-Products from the North-West Spain Fishing Fleet as Potential Sources of Bioactive Peptides. Mar. Drugs 2021, 19, 338. [Google Scholar] [CrossRef] [PubMed]
- Idowu, A.T.; Benjakul, S.; Sinthusamran, S.; Sookchoo, P.; Kishimura, H. Protein Hydrolysate from Salmon Frames: Production, Characteristics and Antioxidative Activity. J. Food Biochem. 2019, 43, e12734. [Google Scholar] [CrossRef] [PubMed]
- Nurdiani, R.; Ramadhan, M.; Prihanto, A.A.; Firdaus, M. Characteristics of Fish Protein Hydrolysate from Mackerel (Scomber japonicus) By- Products. J. Hunan Univ. Nat. Sci. 2022, 49, 75–83. [Google Scholar] [CrossRef]
- Xu, Q.; Gao, F.; Wang, H.; Yang, H. Quality Indices as Potential Markers Indicating the Origin of Cultured Scallop (Argopecten irradians) in the North China Sea. J. Shellfish Res. 2015, 34, 743–750. [Google Scholar] [CrossRef]
- Zhi, T.; Li, X.; Sadiq, F.A.; Mao, K.; Gao, J.; Mi, S.; Liu, X.; Deng, W.; Chitrakar, B.; Sang, Y. Novel Antioxidant Peptides from Protein Hydrolysates of Scallop (Argopecten Irradians) Mantle Using Enzymatic and Microbial Methods: Preparation, Purification, Identification and Characterization. LWT 2022, 164, 113636. [Google Scholar] [CrossRef]
- Najafian, L.; Babji, A.S. Fractionation and Identification of Novel Antioxidant Peptides from Fermented Fish (Pekasam). J. Food Meas. Charact. 2018, 12, 2174–2183. [Google Scholar] [CrossRef]
- Hwang, H.S.; Winkler-Moser, J.K. Antioxidant Activity of Amino Acids in Soybean Oil at Frying Temperature: Structural Effects and Synergism with Tocopherols. Food Chem. 2017, 221, 1168–1177. [Google Scholar] [CrossRef]
- Karami, Z.; Akbari-adergani, B. Bioactive Food Derived Peptides: A Review on Correlation between Structure of Bioactive Peptides and Their Functional Properties. J. Food Sci. Technol. 2019, 56, 535–547. [Google Scholar] [CrossRef]
- González-Serrano, D.J.; Hadidi, M.; Varcheh, M.; Jelyani, A.Z.; Moreno, A.; Lorenzo, J.M. Bioactive Peptide Fractions from Collagen Hydrolysate of Common Carp Fish Byproduct: Antioxidant and Functional Properties. Antioxidants 2022, 11, 509. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Chamorro, I.; Álvarez-Sánchez, N.; Santos-Sánchez, G.; Pedroche, J.; Fernández-Pachón, M.-S.; Millán, F.; Millán-Linares, C.; Lardone, P.J.; Bejarano, I.; Guerrero, J.M.; et al. Immunomodulatory and Antioxidant Properties of Wheat Gluten Protein Hydrolysates in Human Peripheral Blood Mononuclear Cells. Nutrients 2020, 12, 1673. [Google Scholar] [CrossRef]
- Rezvankhah, A.; Yarmand, M.S.; Ghanbarzadeh, B.; Mirzaee, H. Generation of Bioactive Peptides from Lentil Protein: Degree of Hydrolysis, Antioxidant Activity, Phenol Content, ACE-Inhibitory Activity, Molecular Weight, Sensory, and Functional Properties. J. Food Meas. Charact. 2021, 15, 5021–5035. [Google Scholar] [CrossRef]
- Silvestre, M.P.C.; Morais, H.A.; Silva, V.D.M.; Silva, M.R. Degree of Hydrolysis and Peptide Profile of Whey Proteins Using Pancreatin. Nutrire 2013, 38, 278–290. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Independent Variables | Coded Values | ||
|---|---|---|---|
| −1 | 0 | +1 | |
| Temperature (°C) (X1) | 30 | 50 | 70 |
| Time (min) (X2) | 40 | 60 | 80 |
| Enzyme/substrate level (AU/g protein) (X3) | 0.1 | 0.3 | 0.5 |
| Std Order | Run Order | Pt Type | Blocks | Temperature (°C) | Time (min) | [E]/[S] (AU/g de Protein) | Degree of Hydrolysis (DH%) |
|---|---|---|---|---|---|---|---|
| 6 | 1 | 2 | 1 | 70 | 60 | 0.1 | 15.79 ± 0.07 |
| 11 | 2 | 2 | 1 | 50 | 40 | 0.5 | 18.19 ± 0.09 |
| 5 | 3 | 2 | 1 | 30 | 60 | 0.1 | 9.88 ± 0.21 |
| 3 | 4 | 2 | 1 | 30 | 80 | 0.3 | 14.08 ± 0.19 |
| 12 | 5 | 2 | 1 | 50 | 80 | 0.5 | 21.69 ± 0.29 |
| 4 | 6 | 2 | 1 | 70 | 80 | 0.3 | 21.76 ± 0.09 |
| 8 | 7 | 2 | 1 | 70 | 60 | 0.5 | 21.93 ± 0.2 |
| 14 | 8 | 0 | 1 | 50 | 60 | 0.3 | 23.78 ± 0.29 |
| 10 | 9 | 2 | 1 | 50 | 80 | 0.1 | 17.48 ± 0.12 |
| 13 | 10 | 0 | 1 | 50 | 60 | 0.3 | 23.63 ± 0.12 |
| 15 | 11 | 0 | 1 | 50 | 60 | 0.3 | 24.24 ± 0.15 |
| 1 | 12 | 2 | 1 | 30 | 40 | 0.3 | 17.34 ± 0.21 |
| 9 | 13 | 2 | 1 | 50 | 40 | 0.1 | 14.11 ± 0.15 |
| 2 | 14 | 2 | 1 | 70 | 40 | 0.3 | 20.3 ± 0.35 |
| 7 | 15 | 2 | 1 | 30 | 60 | 0.5 | 20.76 ± 0.05 |
| Determination | SVM | SVMD |
|---|---|---|
| Moisture (%) | 1.75 ± 0.17 | 7.03 ± 0.08 |
| Protein (N × 6.25) (%) | 72.23 ± 0.13 | 73.33 ± 0.95 |
| Fat (%) | 7.54 ± 0.14 | nr |
| Ash (%) | 9.54 ± 0.13 | 9.29 ± 0.97 |
| Fibre (%) | 6.75 ± 0.37 | 7.25 ± 0.37 |
| Amino Acids | SVM | SVMD | FAO % (mg Amino Acid/mg Total of Amino Acids) | ||
|---|---|---|---|---|---|
| mg Amino Acid/g Protein | % (mg Amino Acid/mg Total of Amino Acids) | mg Amino Acid/g Protein | % (mg Amino Acid/mg Total of Amino Acids) | ||
| Asp + Asn | 66.57 ± 3.30 | 10.38 ± 0.55 | 47.74 ± 5.86 | 8.55 ± 1.31 | |
| Glu + Gln | 91.56 ± 2.59 | 14.27 ± 0.40 | 77.91 ± 3.63 | 13.92 ± 1.08 | |
| Ser | 34.85 ± 1.94 | 5.42 ± 0.18 | 38.91 ± 3.00 | 6.92 ± 0.32 | |
| His | 12.22 ± 0.82 | 1.90 ± 0.07 | 11.09 ± 1.31 | 1.97 ± 0.17 | 1.6 |
| Gly | 80.06 ± 5.72 | 12.46 ± 0.59 | 86.25 ± 8.69 | 15.33 ± 1.08 | |
| Thr | 32.45 ± 1.64 | 5.05 ± 0.14 | 29.18 ± 2.33 | 5.19 ± 0.25 | 2.5 |
| Arg | 59.70 ± 3.92 | 9.29 ± 0.39 | 56.96 ± 5.27 | 10.66 ± 0.61 | |
| Ala | 35.45 ± 0.97 | 5.53 ± 0.17 | 34.61 ± 1.23 | 6.18 ± 0.41 | |
| Pro | 5.49 ± 2.91 | 0.85 ± 0.43 | 9.19 ± 2.42 | 1.65 ± 0.48 | |
| Tyr b | 20.33 ± 1.11 | 3.16 ± 0.07 | 20.07 ± 2.08 | 3.57 ± 0.26 | |
| Val | 35.89 ± 1.47 | 5.59 ± 0.06 | 10.65 ± 0.09 | 1.90 ± 0.04 | 4.0 |
| Met a | 18.35 ± 0.72 | 2.86 ± 0.14 | 22.32 ± 2.31 | 3.79 ± 0.30 | 2.3 a |
| Cys a | 6.83 ± 0.76 | 1.07 ± 0.14 | 11.12 ± 1.13 | 1.98 ± 0.14 | |
| Ile | 25.65 ± 1.16 | 3.99 ± 0.09 | 12.80 ± 0.47 | 2.28 ± 0.01 | 3.0 |
| Trp | 4.66 ± 0.00 | 0.73 ± 0.03 | 3.96 ± 0.00 | 0.71 ± 0.02 | 0.7 |
| Leu | 46.63 ± 2.16 | 7.26 ± 0.16 | 38.23 ± 1.55 | 6.81 ± 0.07 | 6.1 |
| Phe b | 30.30 ± 1.75 | 4.72 ± 0.19 | 20.52 ± 2.44 | 3.65 ± 0.32 | 4.1 b |
| Lys | 35.04 ± 1.31 | 5.46 ± 0.04 | 27.76 ± 0.22 | 4.95 ± 0.19 | 4.8 |
| Term | Coef | Se Coef | T-Value | p-Value | VIF |
|---|---|---|---|---|---|
| Constant | 23.881 | 0.448 | 53.28 | 0.000 | nr |
| Temperature (°C) | 2.215 | 0.274 | 8.07 | 0.000 | 1.00 |
| Time (min) | 0.633 | 0.274 | 2.31 | 0.027 | 1.00 |
| [E]/[S] (AU/g protein) | 3.164 | 0.274 | 11.53 | 0.000 | 1.00 |
| Temperature (°C) × Temperature (°C) | −3.144 | 0.404 | −7.78 | 0.000 | 1.01 |
| Time (min) × Time (min) | −2.368 | 0.404 | −5.86 | 0.000 | 1.01 |
| [E]/[S] (AU/g protein) × [E]/[S] (AU/g protein) | −3.647 | 0.404 | −9.03 | 0.000 | 1.01 |
| Temperature (°C) × Time (min) | 1.180 | 0.388 | 3.04 | 0.004 | 1.00 |
| Temperature (°C) × [E]/[S] (UA/g protein) | −1.184 | 0.388 | −3.05 | 0.004 | 1.00 |
| Time (min) × [E]/[S] (UA/g protein) | 0.034 | 0.388 | 0.09 | 0.930 | 1.00 |
| Source | DF | Adj SS | Adj MS | F-Value | p-Value |
|---|---|---|---|---|---|
| Model | 9 | 679.585 | 75.509 | 41.76 | 0.000 |
| Linear | 3 | 367.629 | 122.543 | 67.77 | 0.000 |
| Temperature (°C) | 1 | 117.741 | 117.741 | 65.11 | 0.000 |
| Time (min) | 1 | 9.615 | 9.615 | 5.32 | 0.027 |
| [E]/[S] (UA/g protein) | 1 | 240.272 | 240.272 | 132.88 | 0.000 |
| Square | 3 | 278.412 | 92.804 | 51.32 | 0.000 |
| Temperature (°C) × Temperature (°C) | 1 | 109.502 | 109.502 | 60.56 | 0.000 |
| Time (min) × Time (min) | 1 | 62.116 | 62.116 | 34.35 | 0.000 |
| [E]/[S] (UA/g protein) × [E]/[S] (UA/g protein) | 1 | 147.328 | 147.328 | 81.48 | 0.000 |
| 2-Way Interaction | 3 | 33.544 | 11.181 | 6.18 | 0.002 |
| Temperature (°C) × Time (min) | 1 | 16.715 | 16.715 | 9.24 | 0.004 |
| Temperature (°C) × [E]/[S] (UA/g protein) | 1 | 16.816 | 16.816 | 9.30 | 0.004 |
| Time (min) × [E]/[S] (UA/g protein) | 1 | 0.014 | 0.014 | 0.01 | 0.930 |
| Error | 35 | 63.288 | 1.808 | ||
| Lack-of-Fit | 3 | 61.571 | 20.524 | 382.55 | 0.000 |
| Pure Error | 32 | 1.717 | 0.054 | ||
| Total | 44 | 742.873 |
| Determination | SPHFD | DSPHFD |
|---|---|---|
| Moisture (%) | 3.11 ± 0.44 | 4.54 ± 0.24 |
| Ash (%) | 11.08 ± 0.42 | 9.85 ± 0.25 |
| Protein (Nx6.25) (%) | 69.58 ± 0.36 | 71.67 ± 0.36 |
| Yield | 93.92 | 91.29 |
| Colour | Dark browm | Dark browm |
| Degree of hydrolysis (DH %) | 20.44 ± 1.02 | - |
| Amin Acid | SPHFD | DSPHFD | FAO % (mg Amino Acid/mg Total of Amino Acids) | ||
|---|---|---|---|---|---|
| mg Amino Acid/g Protein | % (mg Amino Acid/mg Total of Amino Acids) | mg Amino Acid/g Protein | % (mg Amino Acid/mg Total of Amino Acids) | ||
| Asp + Asn | 94.43 ± 0.88 | 13.68 ± 0.12 | 97.84 ± 9.03 | 12.74 ± 0.72 | |
| Glu + Gln | 110.18 ± 3.22 | 15.92 ± 0.36 | 117.03 ± 1.61 | 15.29 ± 0.40 | |
| Ser | 35.24 ± 0.20 | 5.09 ± 0.03 | 38.38 ± 1.24 | 5.01 ± 0.04 | |
| His | 12.05 ± 0.05 | 1.74 ± 0.01 | 13.32 ± 0.23 | 1.74 ± 0.04 | 1.6 |
| Gly | 73.32 ± 0.81 | 10.60 ± 0.04 | 79.69 ± 0.60 | 10.42 ± 0.50 | |
| Thr | 31.89 ± 0.45 | 4.61 ± 0.03 | 34.15 ± 0.23 | 4.47 ± 0.19 | 2.5 |
| Arg | 57.83 ± 1.26 | 8.36 ± 0.13 | 62.82 ± 0.51 | 8.22 ± 0.41 | |
| Ala | 41.15 ± 0.44 | 5.95 ± 0.02 | 44.70 ± 0.35 | 5.84 ± 0.20 | |
| Pro | 28.45 ± 0.41 | 4.13 ± 0.01 | 56.06 ± 11.23 | 7.08 ± 1.31 | |
| Tyr b | 20.18 ± 0.47 | 2.92 ± 0.05 | 21.91 ± 0.29 | 2.87 ± 0.16 | |
| Val | 22.75 ± 0.52 | 3.29 ± 0.05 | 23.63 ± 0.21 | 3.09 ± 0.14 | 4.0 |
| Met a | 23.44 ± 0.57 | 3.39 ± 0.05 | 24.71 ± 0.44 | 3.23 ± 0.16 | 2.3 a |
| Cys a | 5.48± 0.05 | 0.79± 0.00 | 6.24 ± 0.17 | 0.81 ± 0.01 | |
| Ile | 26.24 ± 0.70 | 3.79 ± 0.06 | 28.25 ± 0.55 | 3.70 ± 0.23 | 3.0 |
| Trp | 5.70 ± 0.00 | 0.82 ± 0.01 | 5.77 ± 0.00 | 0.76 ± 0.03 | 0.7 |
| Leu | 45.20 ± 0.73 | 6.53 ± 0.06 | 49.04 ± 0.34 | 6.41 ± 0.30 | 6.1 |
| Phe b | 22.14 ± 0.75 | 3.20 ± 0.09 | 23.94 ± 0.22 | 3.13 ± 0.16 | 4.1 b |
| Lys | 36.07 ± 0.63 | 5.21 ± 0.06 | 39.58 ± 0.32 | 5.18 ± 0.25 | 4.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chasquibol, N.; Gonzales, B.F.; Alarcón, R.; Sotelo, A.; Márquez-López, J.C.; Rodríguez-Martin, N.M.; del Carmen Millán-Linares, M.; Millán, F.; Pedroche, J. Optimisation and Characterisation of the Protein Hydrolysate of Scallops (Argopecten purpuratus) Visceral By-Products. Foods 2023, 12, 2003. https://doi.org/10.3390/foods12102003
Chasquibol N, Gonzales BF, Alarcón R, Sotelo A, Márquez-López JC, Rodríguez-Martin NM, del Carmen Millán-Linares M, Millán F, Pedroche J. Optimisation and Characterisation of the Protein Hydrolysate of Scallops (Argopecten purpuratus) Visceral By-Products. Foods. 2023; 12(10):2003. https://doi.org/10.3390/foods12102003
Chicago/Turabian StyleChasquibol, Nancy, Billy Francisco Gonzales, Rafael Alarcón, Axel Sotelo, José Carlos Márquez-López, Noelia M. Rodríguez-Martin, María del Carmen Millán-Linares, Francisco Millán, and Justo Pedroche. 2023. "Optimisation and Characterisation of the Protein Hydrolysate of Scallops (Argopecten purpuratus) Visceral By-Products" Foods 12, no. 10: 2003. https://doi.org/10.3390/foods12102003