
Maytenus octogona Superoxide Scavenging and Anti-Inflammatory Caspase-1 Inhibition Study Using Cyclic Voltammetry and Computational Docking Techniques

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
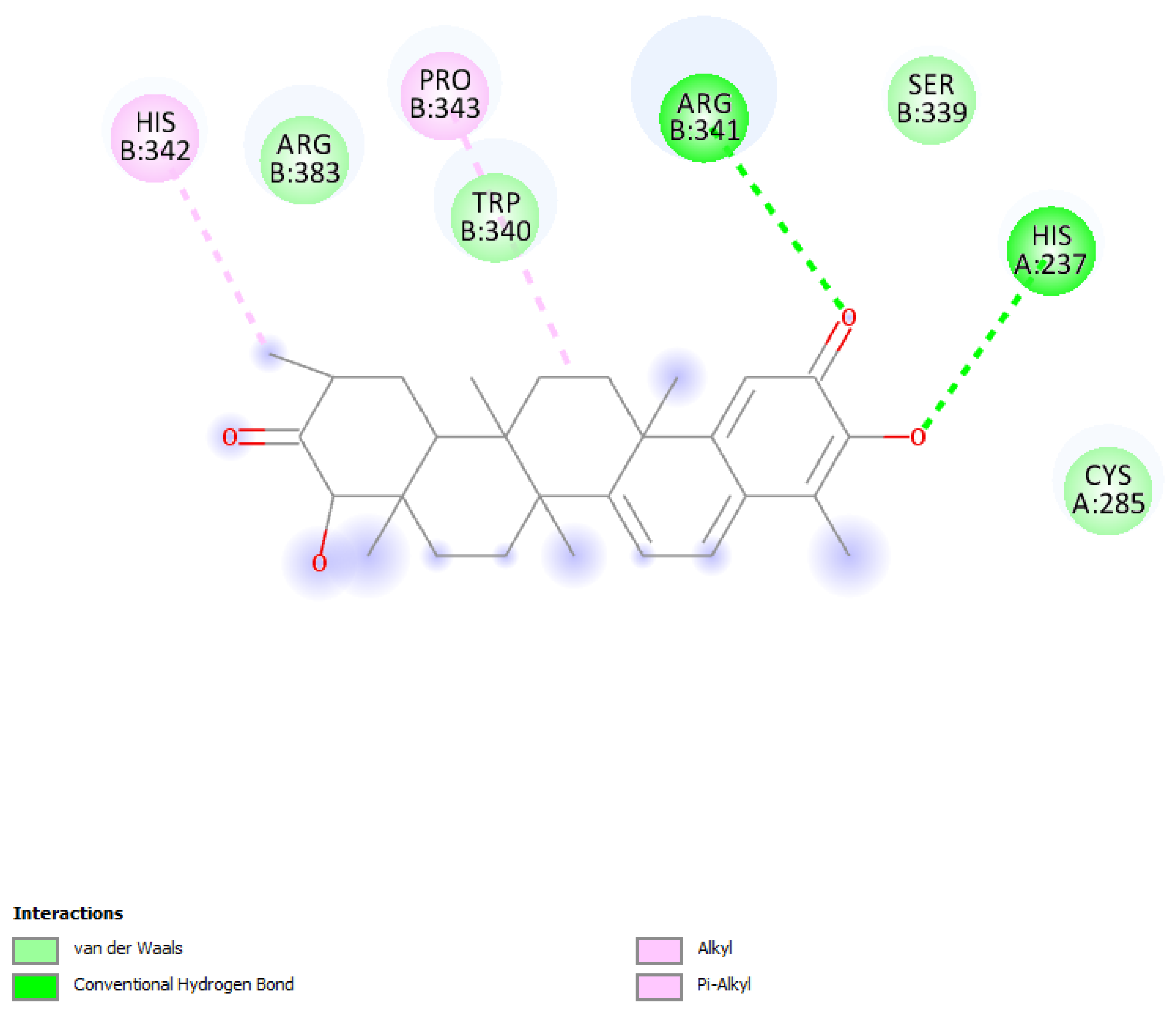
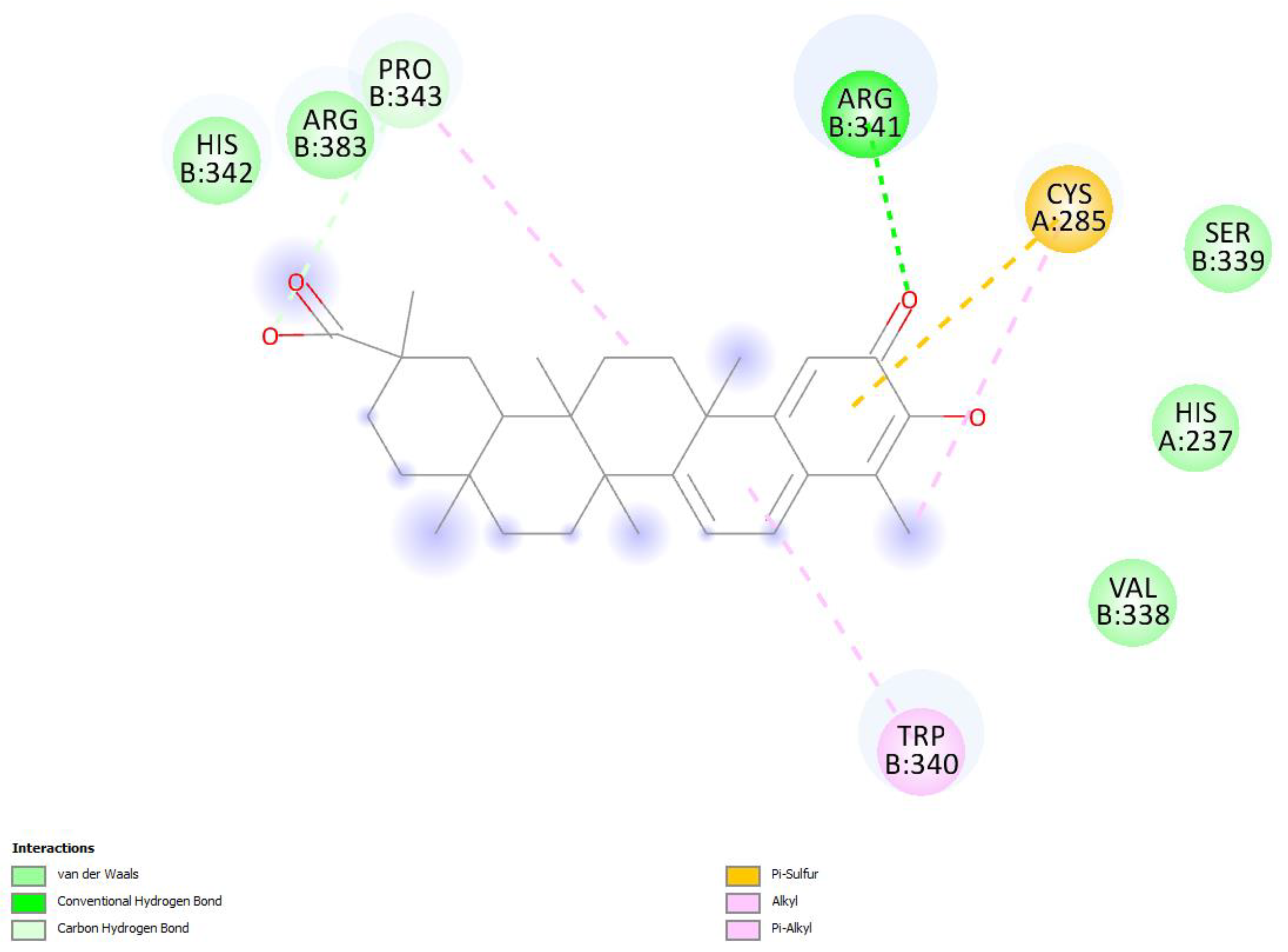
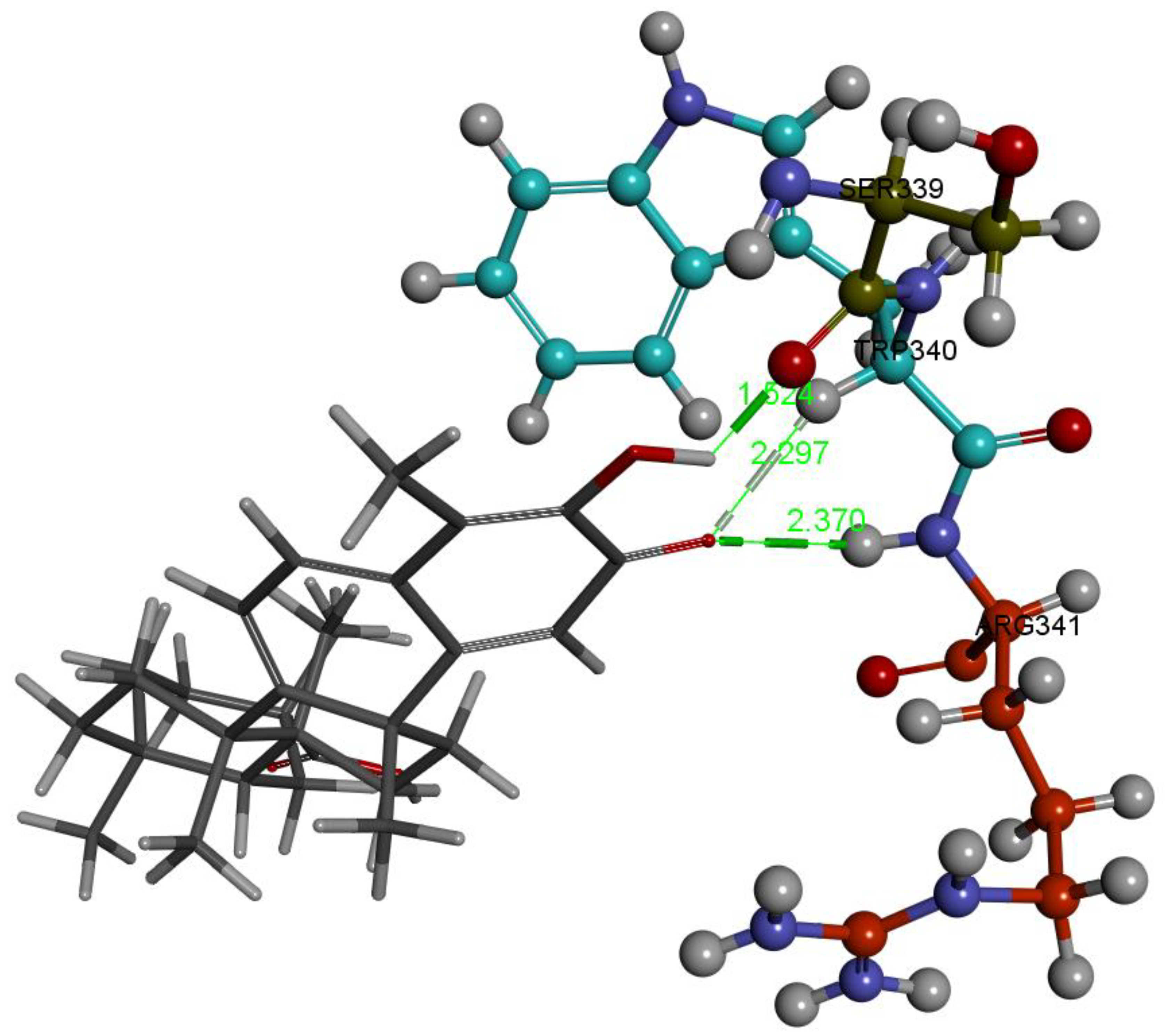
2.1. Caspase-1 Docking
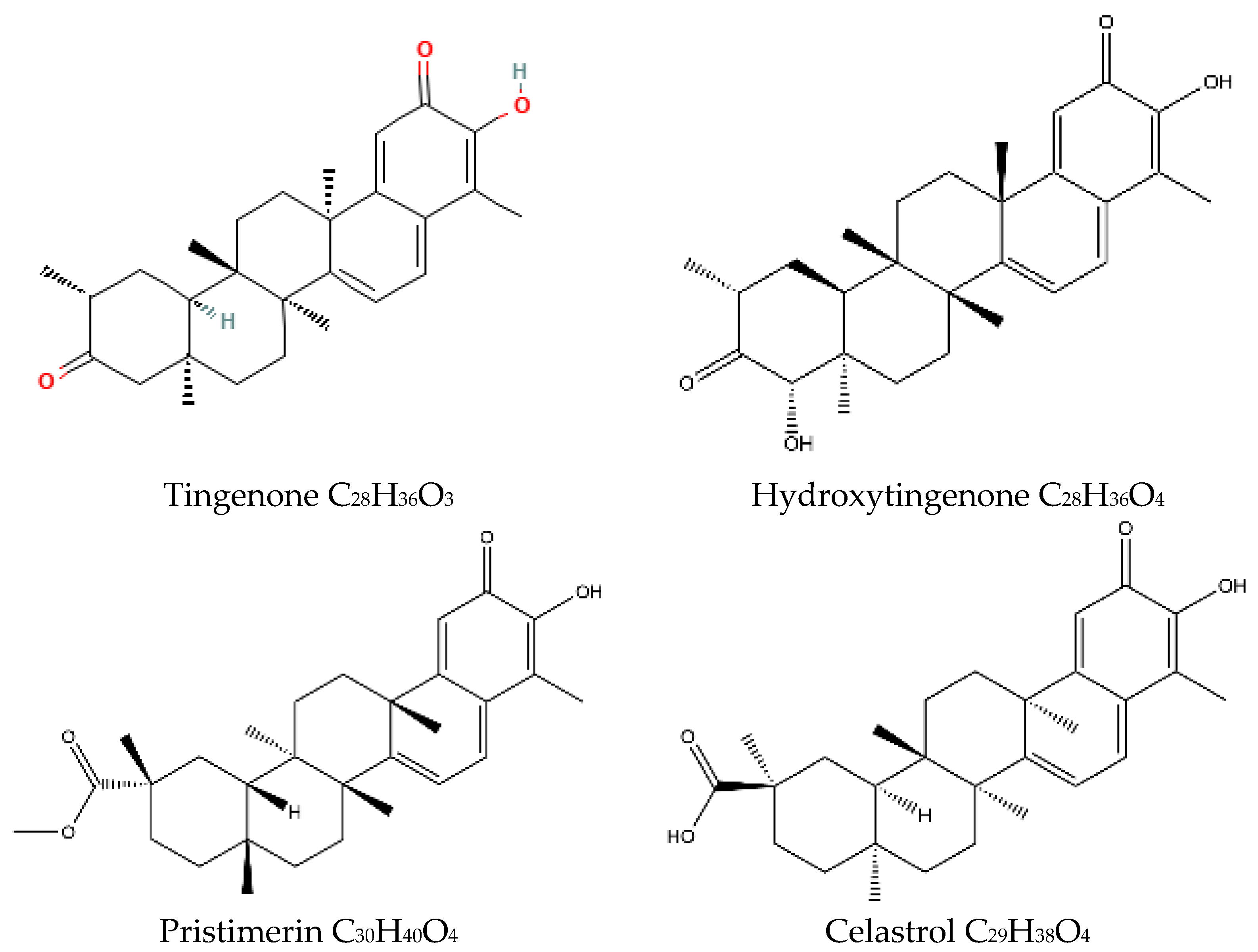
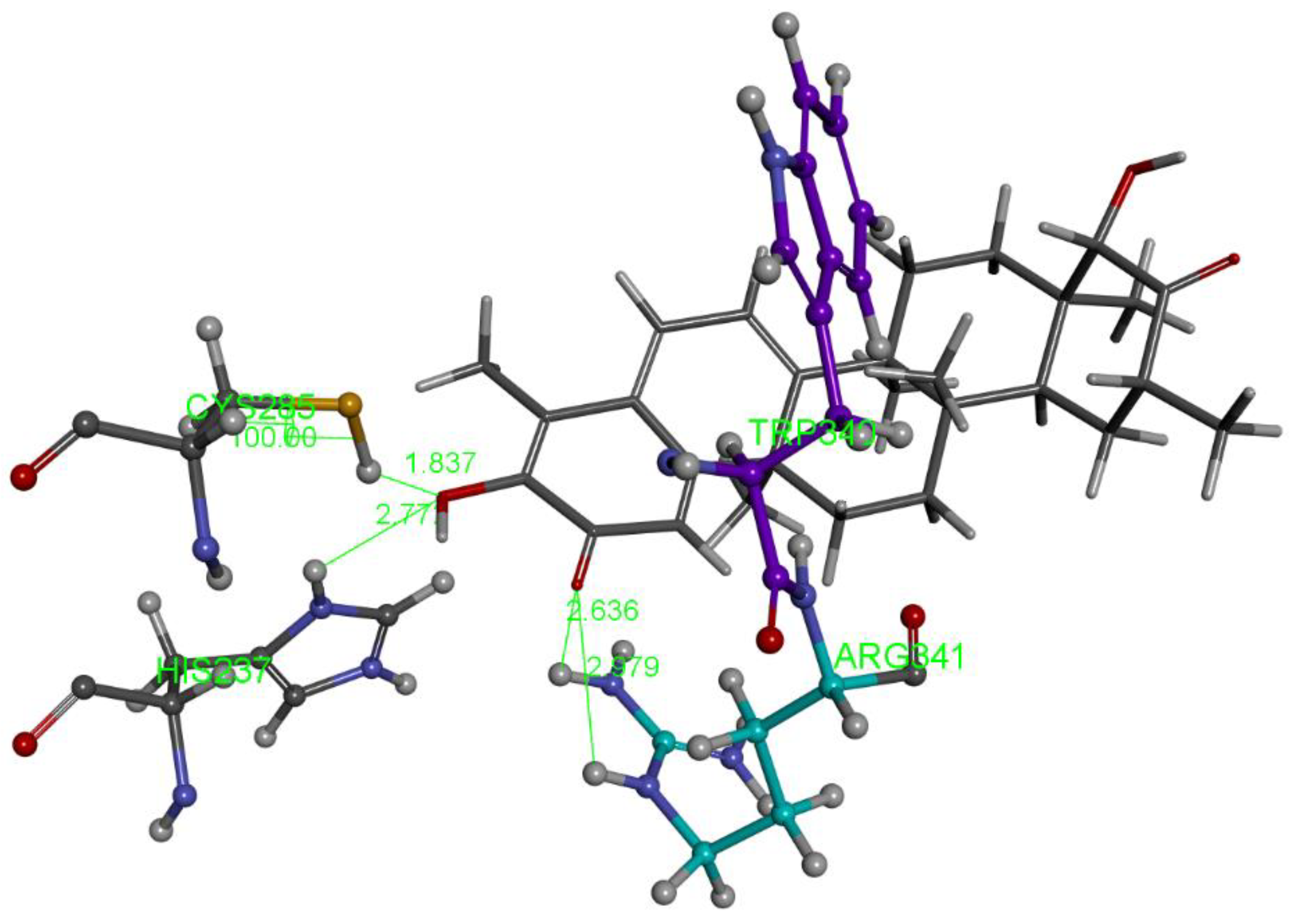
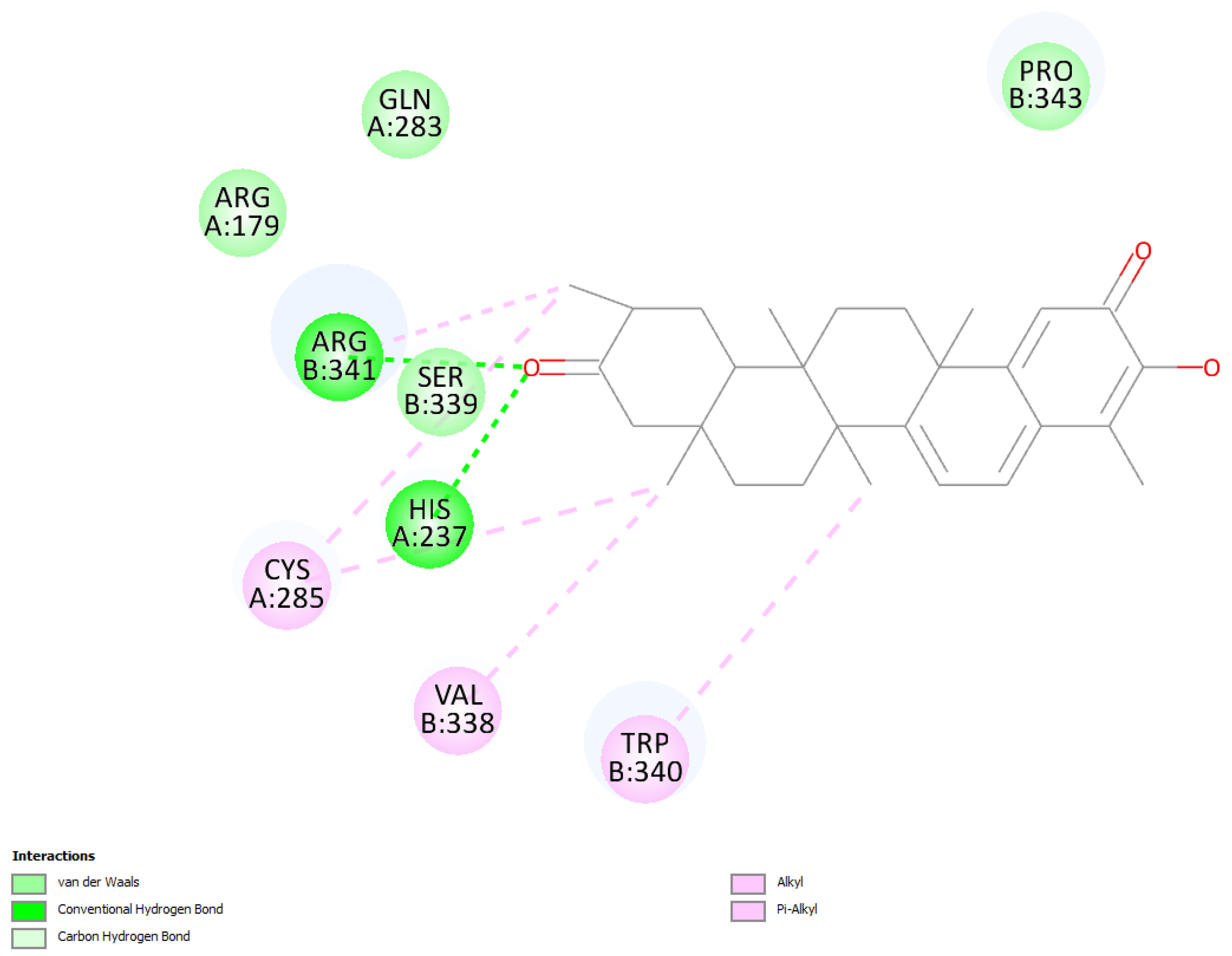
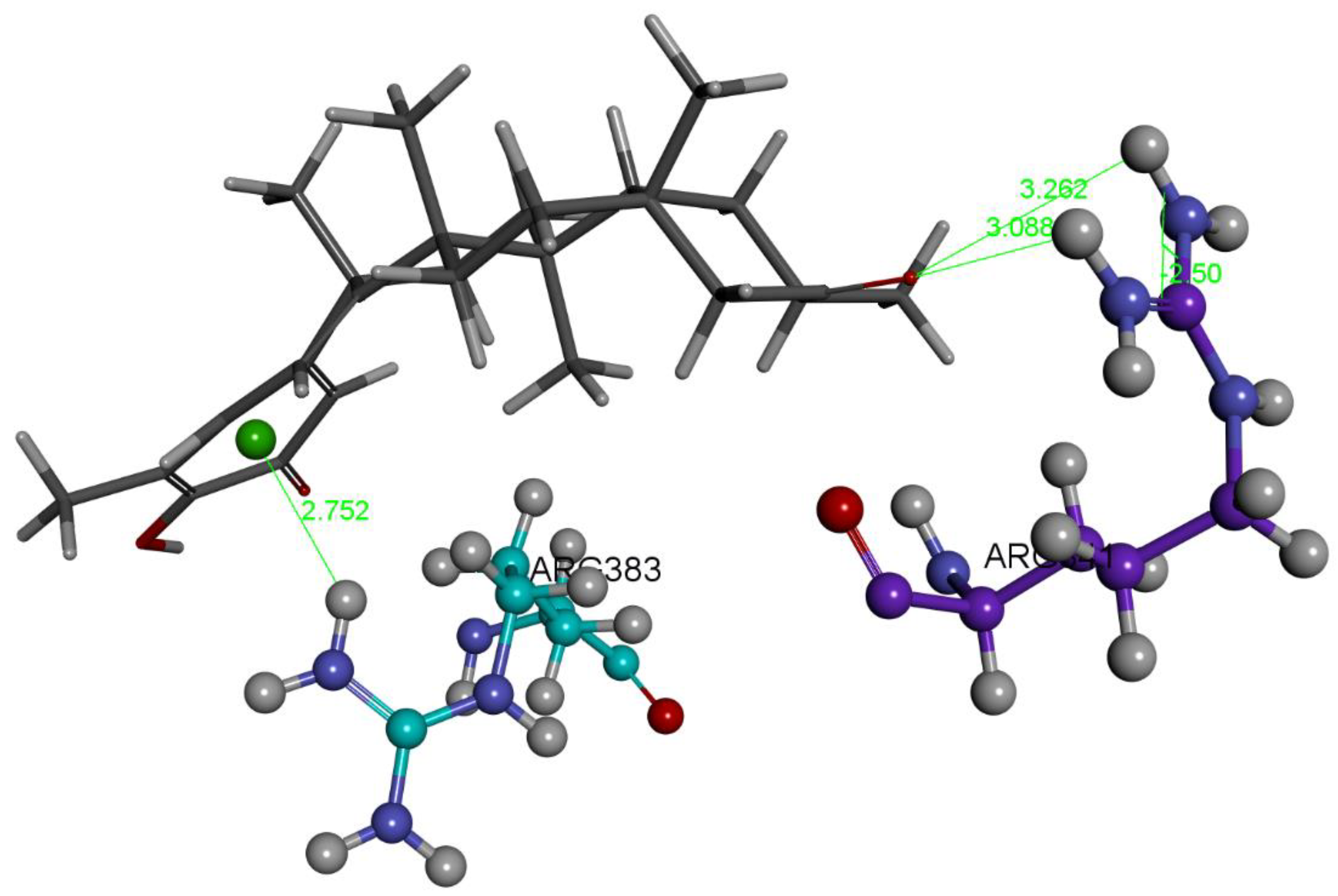
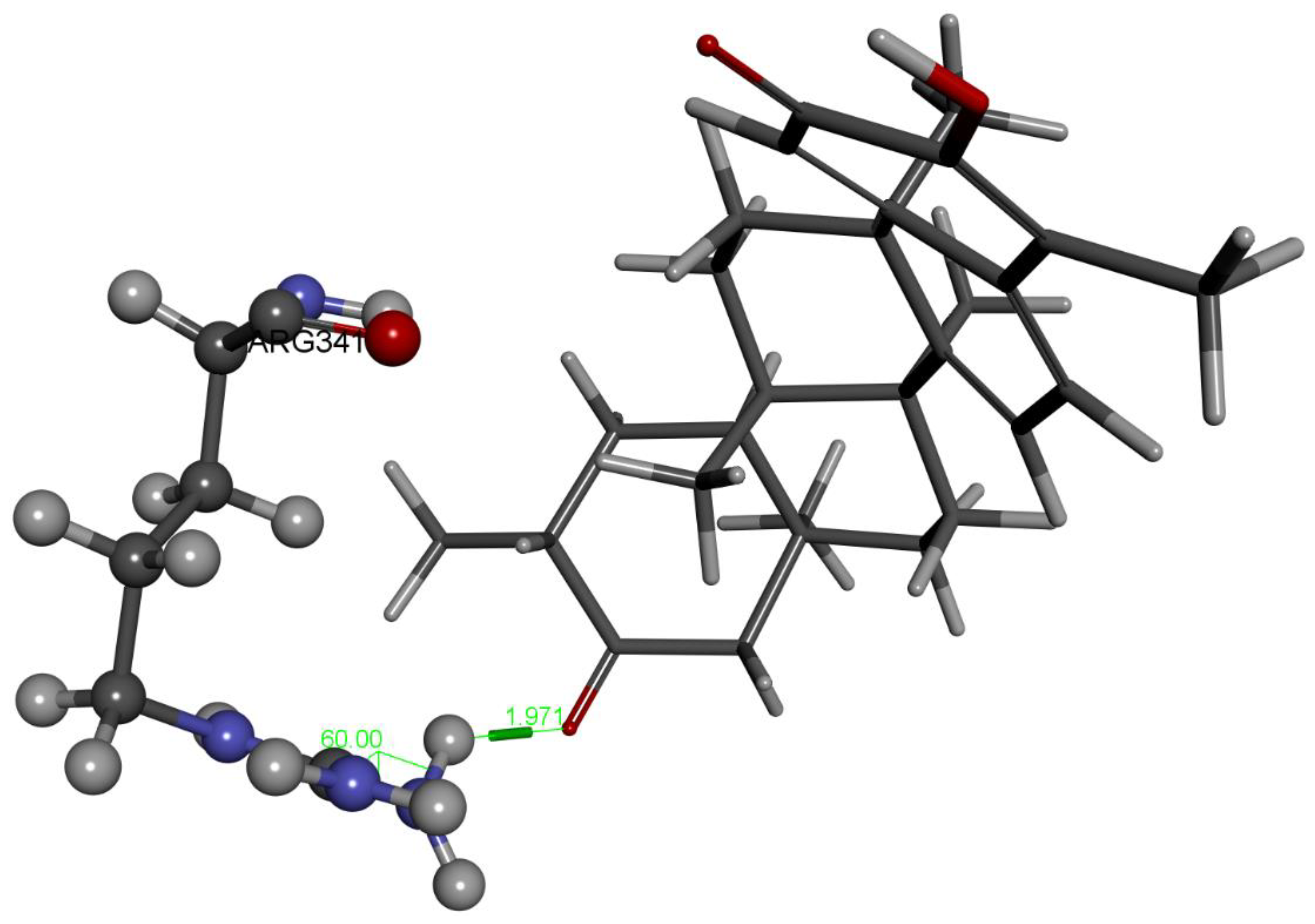
2.1.1. Hydroxytingenone
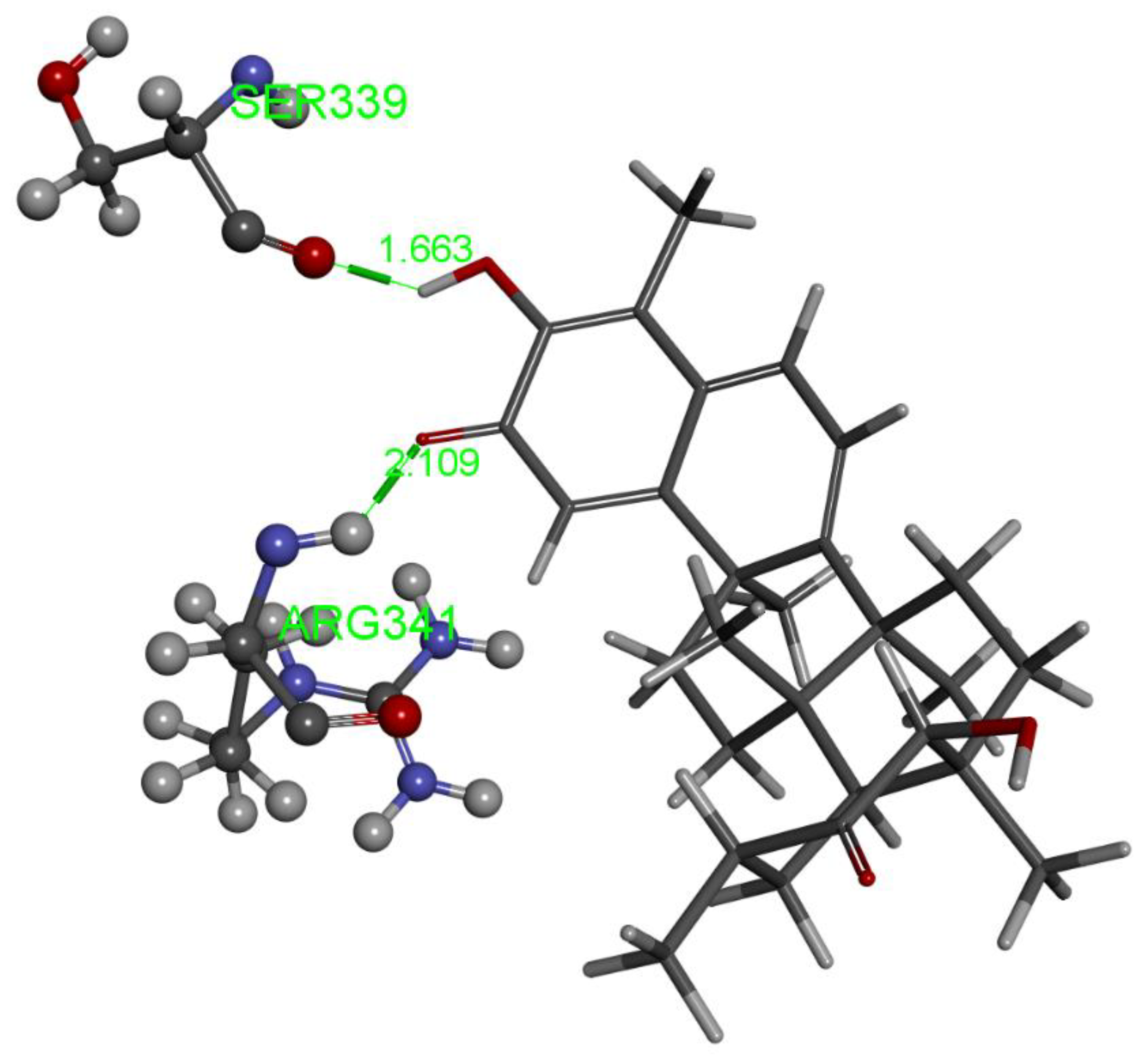
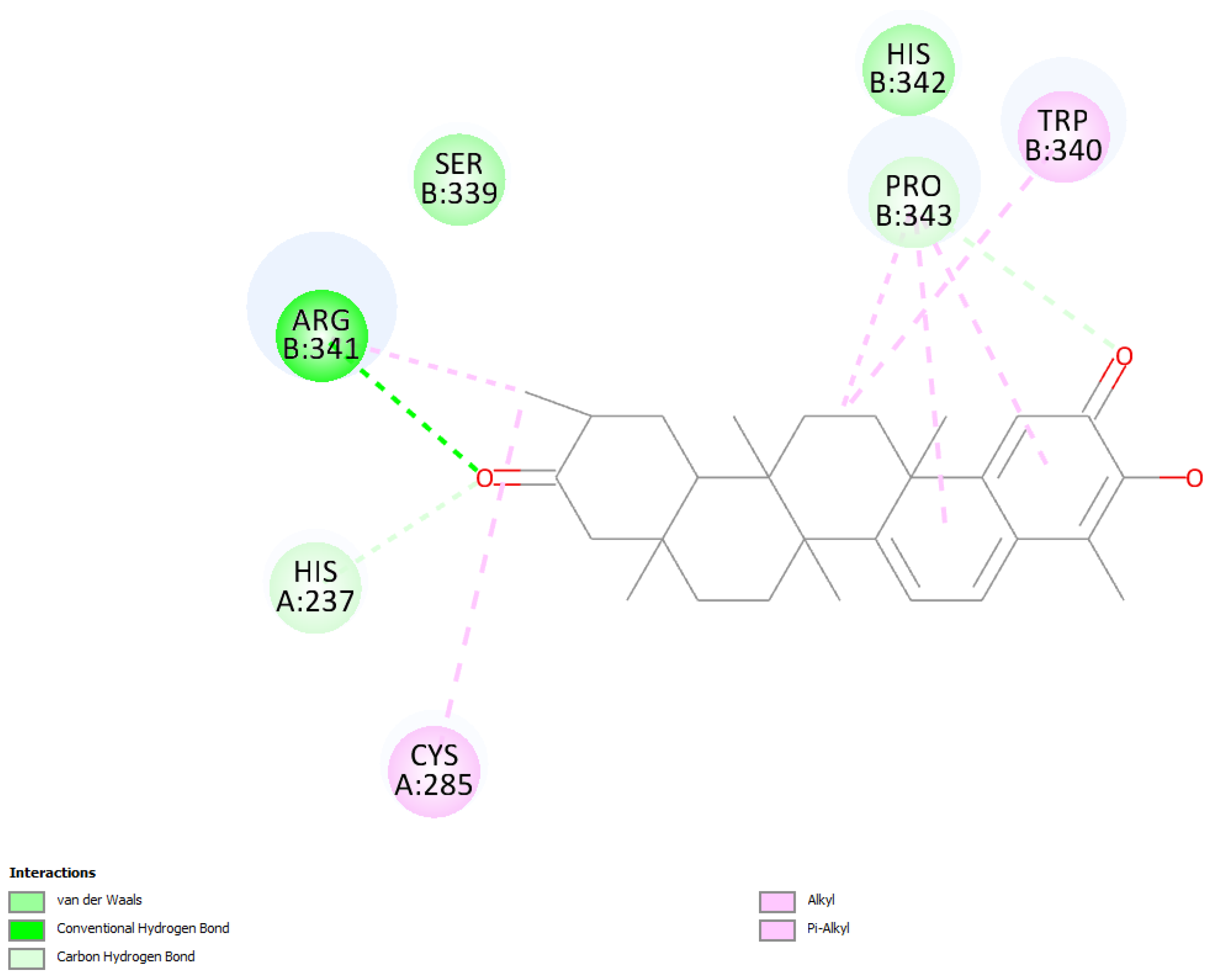
2.1.2. Tingenone
2.1.3. Pristimerin
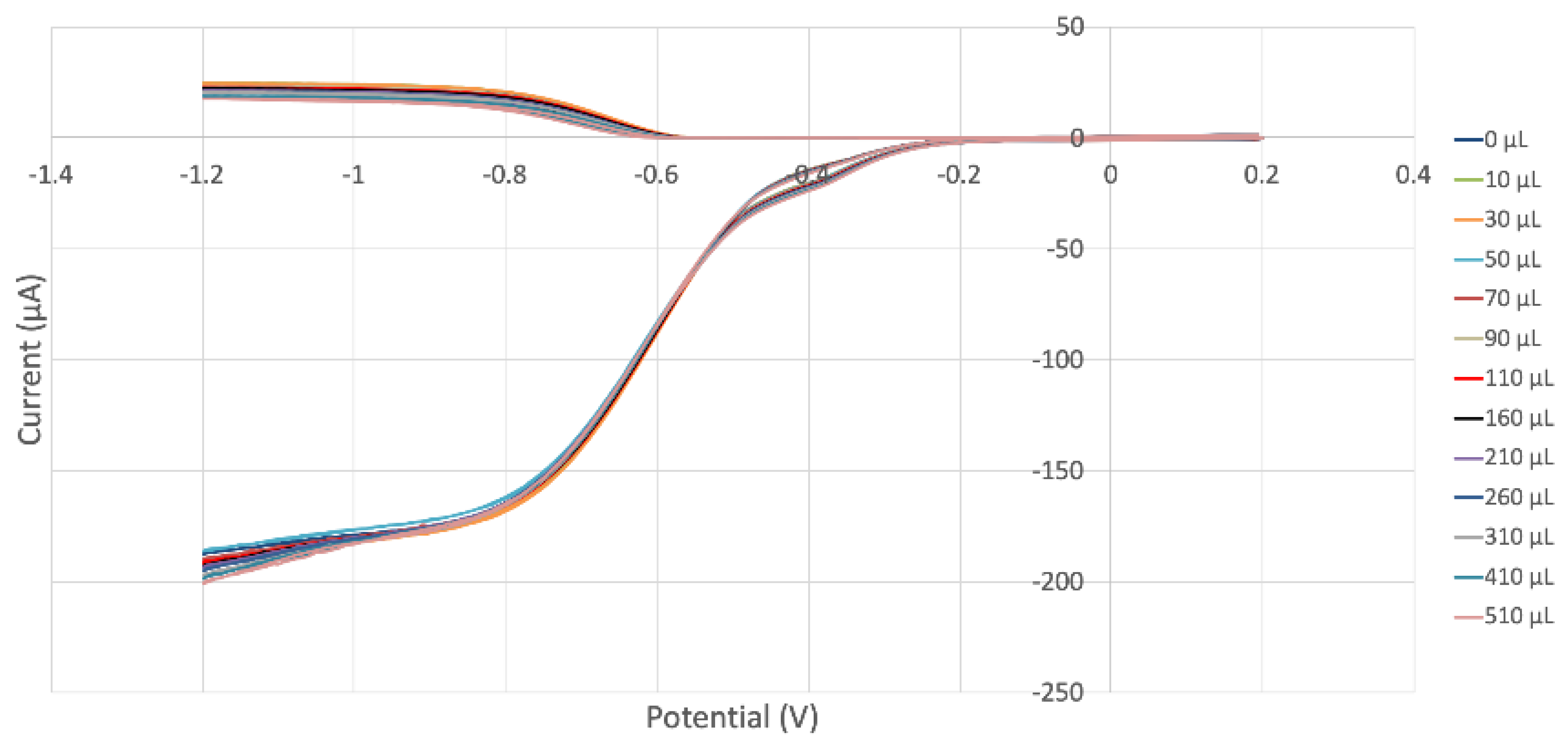
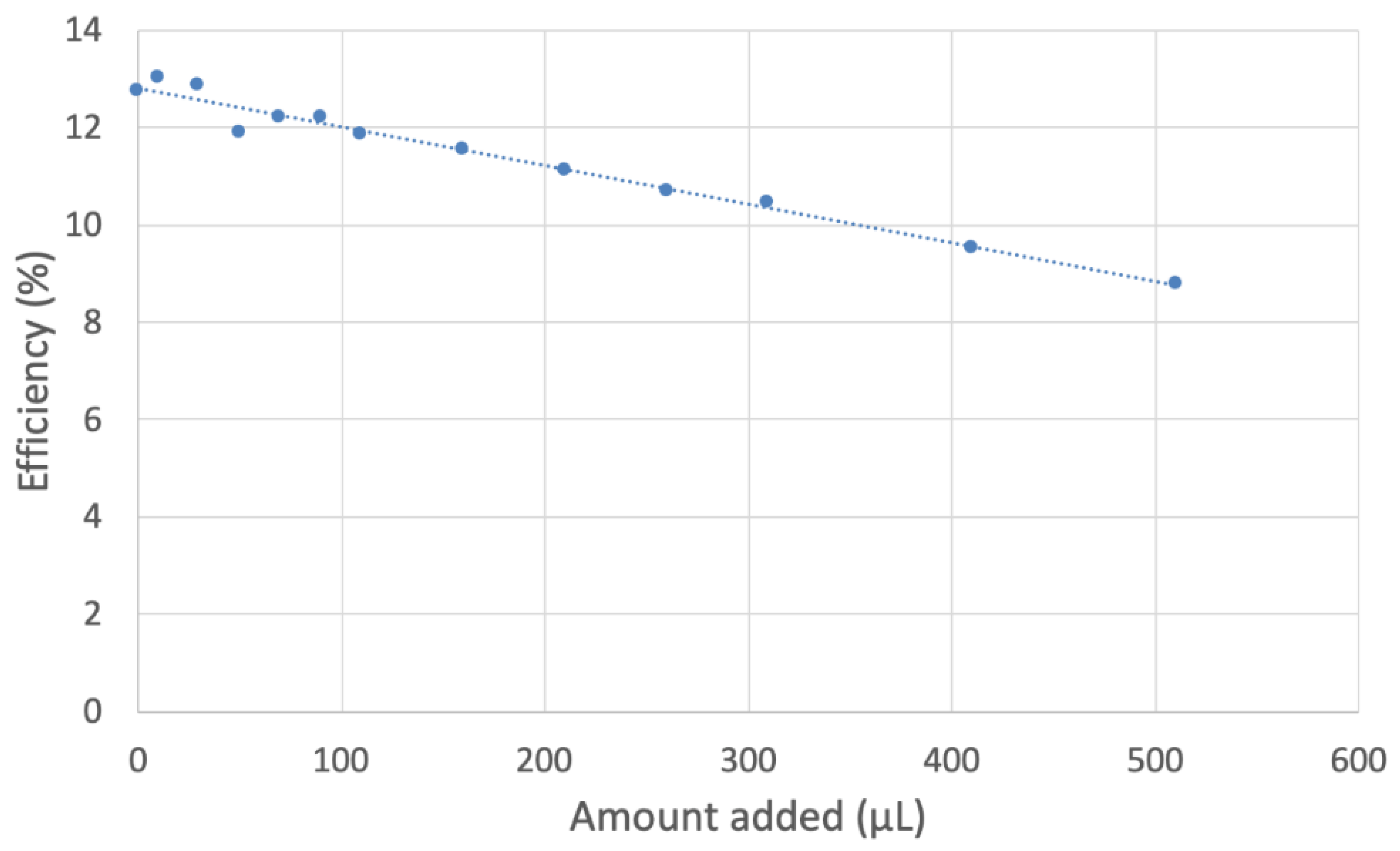
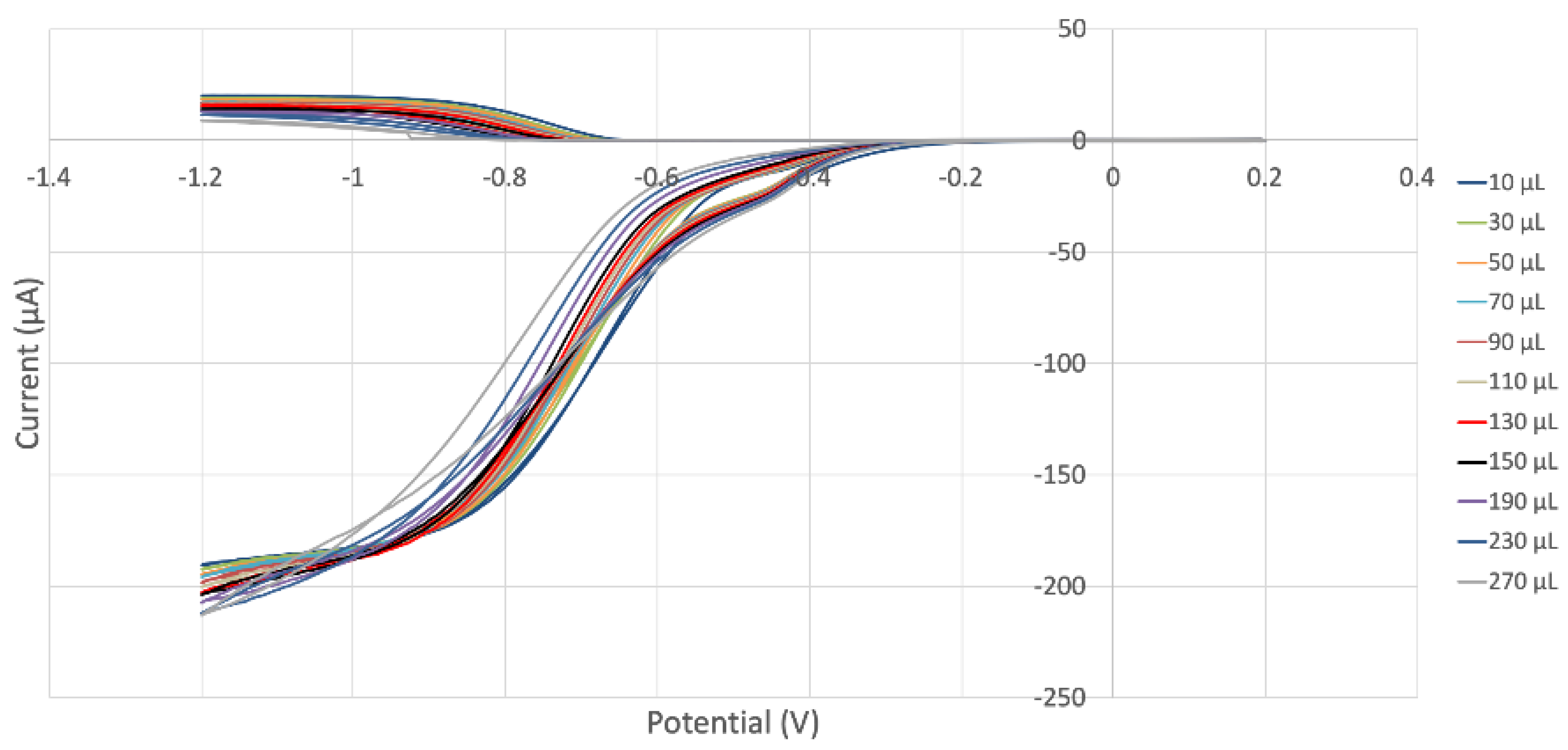
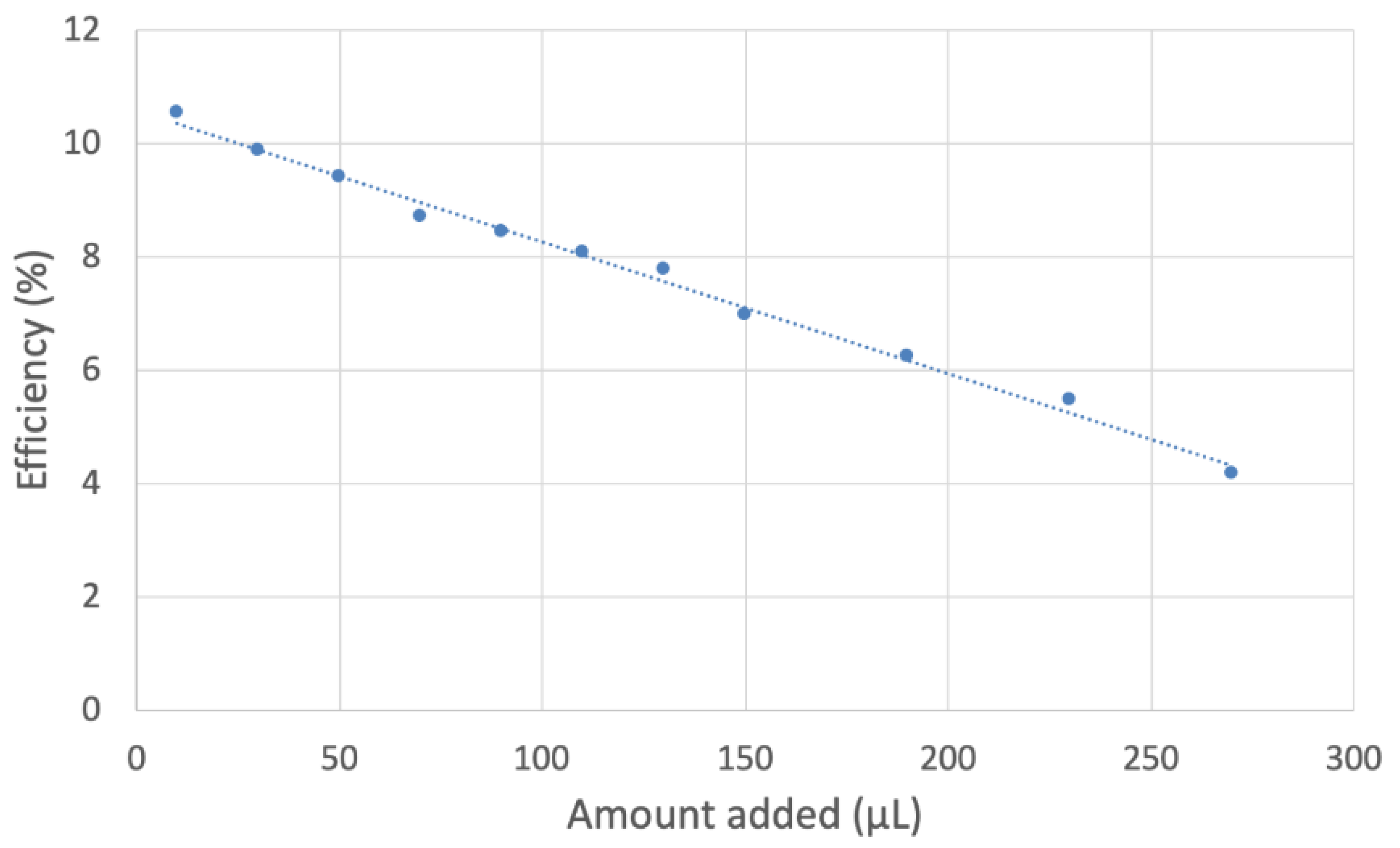
2.2. RRDE
3. Materials and Methods
3.1. Reagents
3.1.1. MOH UNICA (Leaf Extract)
3.1.2. MOR UNICA (Stem Extract)
3.2. Electrochemistry
3.3. Hydrodynamic Voltammetry (RRDE)
3.4. Computational Study
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kesavardhana, S.; Subbarao Malireddi, R.K.; Kanneganti, T.D. Caspases in Cell Death, Inflammation, and Pyroptosis. Annu. Rev. Immunol. 2020, 38, 567. [Google Scholar] [CrossRef] [Green Version]
- Meduri, G.U.; Headley, S.; Kohler, G.; Stentz, F.; Tolley, E.; Umberger, R.; Leeper, K. Persistent elevation of inflammatory cytokines predicts a poor outcome in ARDS. Plasma IL-1 beta and IL-6 levels are consistent and efficient predictors of outcome over time. Chest 1995, 107, 1062. [Google Scholar] [CrossRef] [PubMed]
- Mehta, P.; Mcauley, D.F.; Brown, M.; Sanchez, E.; Tattersall, R.S.; Manson, J.J. COVID-19: Consider cytokine storm syndromes and immunosuppression. Lancet 2020, 395, 1033. [Google Scholar] [CrossRef] [PubMed]
- Nieto-Torres, J.L.; Dediego, M.L.; Verdia-Baguena, C.; Jimenez-Guardeno, J.M.; Regla-Nava, J.A.; Fernandez-Delgado, R.; Castaño-Rodriguez, C.; Alcaraz, A.; Torres, J.; Aguilella, V.M.; et al. Severe acute respiratory syndrome coronavirus envelope protein ion channel activity promotes virus fitness and pathogenesis. PloS Pathog. 2014, 10, e1004077. [Google Scholar] [CrossRef] [Green Version]
- Rogers, A.J.; Guan, J.; Trtchounian, A.; Hunninghake, G.M.; Kaimal, R.; Desai, M.; Kozikowski, L.-A.; DeSouza, L.; Mogan, S.; Liu, K.D.; et al. Association of elevated plasma interleukin-18 level with increased mortality in a clinical trial of statin treatment for acute respiratory distress syndrome. Crit. Care Med. 2019, 47, 1089. [Google Scholar] [CrossRef] [PubMed]
- De Rivero Vaccari, J.C.; Dietrich, W.D.; Keane, R.W.; de Rivero Vaccari, J.P. The Inflammasome in Times of COVID-19. Front. Immunol. 2020, 11, 583373. [Google Scholar] [CrossRef]
- Kigerl, K.A.; de Rivero Vaccari, J.P.; Dietrich, W.D.; Popovich, P.G.; Keane, R.W. Pattern recognition receptors and central nervous system repair. Exp. Neurol. 2014, 258, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamkanfi, M.; Dixit, V.M. Inflammasomes and their roles in health and disease. Annu. Rev. Cell Dev. Biol. 2012, 28, 137–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strowig, T.; Henao-Mejia, J.; Elinav, E.; Flavell, R. Inflammasomes in health and disease. Nature 2012, 481, 278–286. [Google Scholar] [CrossRef]
- Tan, L.Y.; Komarasamy, T.V.; Rmt Balasubramaniam, V. Hyperinflammatory Immune Response and COVID-19: A Double Edged Sword. Front. Immunol. 2021, 12, 742941. [Google Scholar] [CrossRef]
- Caruso, F.; Pedersen, J.Z.; Incerpi, S.; Kaur, S.; Belli, S.; Florea, R.M.; Rossi, M. Mechanism of Caspase-1 Inhibition by Four Anti-inflammatory Drugs Used in COVID-19 Treatment. Int. J. Mol. Sci. 2022, 23, 1849. [Google Scholar] [CrossRef] [PubMed]
- Caruso, F.; Singh, M.; Belli, S.; Berinato, M.; Rossi, M. Interrelated mechanism by which the methide quinone celastrol, obtained from the roots of Tripterygium wilfordii, inhibits main protease 3CLpro of COVID-19 and acts as superoxide radical scavenger. Int. J. Mol. Sci. 2020, 21, 9266. [Google Scholar] [CrossRef]
- Huang, Y.Y.; Chen, L.; Ma, G.X.; Xu, X.D.; Jia, X.G.; Deng, F.S.; Li, X.J.; Yuan, J.Q. A Review on Phytochemicals of the Genus Maytenus and Their Bioactive Studies. Molecules 2021, 26, 4563. [Google Scholar] [CrossRef] [PubMed]
- Niero, R.; de Andrade, S.F.; Cechinel Filho, V. A review of the ethnopharmacology, phytochemistry and pharmacology of plants of the Maytenus genus. Curr. Pharm. Des. 2011, 17, 1851. [Google Scholar] [CrossRef] [PubMed]
- Lock, O.; Perez, E.; Villar, M.; Flores, D.; Rojas, R. Bioactive Compounds from Plants Used in Peruvian Traditional Medicine. Nat. Prod. Commun. 2016, 11, 315–337. [Google Scholar]
- Gonzalez, J.G.; Delle Monache, G.; Delle Monache, F.; Marini-Bettolò, G.B. Chuchuhuasha–a drug used in folk medicine in the Amazonian and Andean areas. A chemical study of Maytenus laevis. J. Ethnopharmacol. 1982, 5, 73. [Google Scholar] [CrossRef]
- Oramas-Royo, S.M.; Chávez, H.; Martín-Rodríguez, P.; Fernández-Pérez, L.; Ravelo, A.G.; Estévez-Braun, A. Cytotoxic triterpenoids from Maytenus retusa. J. Nat. Prod. 2010, 73, 2029. [Google Scholar] [CrossRef]
- Vazdekis, N.E.; Chávez, H.; Estévez-Braun, A.; Ravelo, A.G. Triterpenoids and a lignan from the aerial parts of Maytenus apurimacensis. J. Nat. Prod. 2009, 72, 1045–1048. [Google Scholar] [CrossRef]
- Delgado-Méndez, P.; Herrera, N.; Chávez, H.; Estévez-Braun, A.; Ravelo, A.G.; Cortes, F.; Castanys, S.; Gamarro, F. New terpenoids from Maytenus apurimacensis as MDR reversal agents in the parasite Leishmania. Bioorg. Med. Chem. 2008, 16, 1425–1430. [Google Scholar] [CrossRef]
- Torpocco, V.; Chávez, H.; Estévez-Braun, A.; Ravelo, A.G. New dammarane triterpenes from Maytenus macrocarpa. Chem. Pharm. Bull. 2007, 55, 812–814. [Google Scholar] [CrossRef] [Green Version]
- Available online: https://www.rcsb.org/ (accessed on 25 July 2022).
- Yang, J.; Liu, Z.; Xiao, T.S. Crystal Structure of Caspase-1 in Complex with VX. Available online: https://www.wwpdb.org/pdb?id=pdb_00006pzp (accessed on 25 July 2022).
- Wu, G.; Robertson, D.H.; Brooks, C.L.; Vieth, M.J. Detailed analysis of grid-based molecular docking: A case study of CDOCKER-A CHARMm-based MD docking algorithm. J. Comput. Chem. 2003, 24, 1549. [Google Scholar] [CrossRef] [PubMed]
- Belli, S.; Rossi, M.; Molasky, N.; Middleton, L.; Caldwell, C.; Bartow-McKenney, C.; Duong, M.; Chiu, J.; Gibbs, E.; Caldwell, A.; et al. Effective and novel application of superoxide radical scavenging by natural phenolic antioxidants. Antioxidants 2019, 8, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, M.; Caruso, F.; Kwok, L.; Lee, G.; Caruso, A.; Gionfra, F.; Candelotti, E.; Belli, S.L.; Molasky, N.; Raley-Susman, K.M.; et al. Protection by extra virgin olive oil against oxidative stress in vitro and in vivo. Chemical and biological studies on the health benefits due to a major component of the Mediterranean diet. PLoS ONE 2017, 12, e0189341. [Google Scholar] [CrossRef] [PubMed]
- Sakib, R.; Caruso, F.; Aktar, S.; Belli, S.; Kaur, S.; Hernandez, M.; Rossi, M. Antioxidant Properties of Thymoquinone, Thymohydroquinone and Black Cumin (Nigella sativa L.) Seed Oil: Scavenging of Superoxide Radical Studied Using Cyclic Voltammetry, DFT and Single Crystal X-ray Diffraction. Antioxidants 2023, 12, 607. [Google Scholar] [CrossRef]
- Caruso, F.; Berinato, M.; Hernandez, M.; Belli, S.; Smart, C.; Rossi, M. Antioxidant properties of bee propolis and an important component, galangin, described by X-ray crystal structure, DFTD and hydrodynamic voltammetry. PLoS ONE 2022, 17, e0267624. [Google Scholar] [CrossRef]
- Mayrhofer, K.; Strmcnik, D.; Blizanac, B.; Stamenkovic, V.; Arenz, M.; Markovic, N. Measurement of oxygen reduction activities via the rotating disc electrode method: From Pt model surfaces to carbon-supported high surface area catalysts. Electrochim. Acta 2008, 53, 3181–3188. [Google Scholar] [CrossRef]
- Delley, B.J. From molecules to solids with the DMol3 approach. J. Chem. Phys. 2000, 113, 7756. [Google Scholar] [CrossRef]
- Perdew, J.P.; Chevary, J.A.; Vosko, S.H.; Jackson, K.A.; Pederson, M.R.; Singh, D.J.; Fiolhais, C. Atoms, molecules, solids, and surfaces: Applications of the generalized gradient approximation for exchange and correlation. Phys. Rev. 1992, 46, 6671–6687. [Google Scholar] [CrossRef]
- Becke, A.D. Density-functional exchange-energy approximation with correct asymptotic behavior. Phys. Rev. A 1988, 38, 3098–3100. [Google Scholar] [CrossRef]
- Li, Y.; Huang, H.; Liu, B.; Zhang, Y.; Pan, X.; Yu, X.Y.; Shen, Z.; Song, Y.H. Inflammasomes as therapeutic targets in human diseases. Signal Transduct. Target Ther. 2021, 6, 247. [Google Scholar] [CrossRef]
- Wannamaker, W.; Davies, R.; Namchuk, M.; Pollard, J.; Ford, P.; Ku, G.; Decker, C.; Charifson, P.; Weber, P.; Germann, U.A.; et al. [(S)-1-((S)-2-{[1-(4-Amino-3-chloro-phenyl)-methanoyl]-amino}-3,3-dimethyl-butanoyl)-pyrrolidine-2-carboxylic acid ((2R,3S)-2-ethoxy-5-oxo-tetrahydro-furan-3-yl)-amide (VX-765), an Orally Available Selective Interleukin (IL)-Converting Enzyme/Caspase-1 Inhibitor, Exhibits Potent Anti-Inflammatory Activities by Inhibiting the Release of IL-1β and IL. J. Pharmacol. Exp. Therap. 2007, 321, 509–516. [Google Scholar] [CrossRef]
- Cao, D.Y.; Zhang, Z.H.; Li, R.Z.; Shi, X.K.; Xi, R.Y.; Zhang, G.L.; Li, F.; Wang, F. A small molecule inhibitor of caspase-1 inhibits NLRP3 inflammasome activation and pyroptosis to alleviate gouty inflammation. Immunol. Lett. 2022, 244, 28–39. [Google Scholar] [CrossRef] [PubMed]
- González-Cofrade, L.; Green, J.P.; Cuadrado, I.; Amesty, Á.; Oramas-Royo, S.; Brough, D.; Estévez-Braun, A.; Hortelano, S.; de Las Heras, B. Phenolic and quinone methide nor-triterpenes as selective NLRP3 inflammasome inhibitors. Bioorg. Chem. 2023, 132, 106362. [Google Scholar] [CrossRef] [PubMed]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| P1 | P2 | P3 | P4 | P5 | P6 | P7 | P8 | P9 | P10 | |
|---|---|---|---|---|---|---|---|---|---|---|
| H-Tingenone | Cys285His237 −7.2 | Cys285 His237 −7.2 | Arg341 Cys285 −8.9 | Arg341 His237 −13.7 | Trp340 π OUT | Trp340 π OUT | Trp340 OUT | Arg341 His237 | Trp340 π OUT | Trp340 Cys285 (unfavored) |
| Tingenone | Arg341 His237 −11.8 | His237 −11.8 | His237 Cys285 −11.8 | His237 −11.8 | Arg341 * −13.0 | His237 −11.8 | Arg341 His342 (π-π) * −13.0 | His237 Cys285 −11.8 | Arg341 His342 (π-π) * −13.0 | His237 Cys285 |
| Pristimerin | Arg341 Cys285 OUT | Asp288 −9.3 | Asp288 −9.3 | Arg341 Cys285 (π-S) * −9.4 | His342 (π-π) * −5.8 | none −5.8 | Arg341 Cys285 (π-S) * −9.4 | His342 (π-π) * −5.8 | none | Arg341 Cys285 (π-S) * −9.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caruso, F.; Rossi, M.; Eberhardt, E.; Berinato, M.; Sakib, R.; Surco-Laos, F.; Chavez, H. Maytenus octogona Superoxide Scavenging and Anti-Inflammatory Caspase-1 Inhibition Study Using Cyclic Voltammetry and Computational Docking Techniques. Int. J. Mol. Sci. 2023, 24, 10750. https://doi.org/10.3390/ijms241310750
Caruso F, Rossi M, Eberhardt E, Berinato M, Sakib R, Surco-Laos F, Chavez H. Maytenus octogona Superoxide Scavenging and Anti-Inflammatory Caspase-1 Inhibition Study Using Cyclic Voltammetry and Computational Docking Techniques. International Journal of Molecular Sciences. 2023; 24(13):10750. https://doi.org/10.3390/ijms241310750
Chicago/Turabian StyleCaruso, Francesco, Miriam Rossi, Eric Eberhardt, Molly Berinato, Raiyan Sakib, Felipe Surco-Laos, and Haydee Chavez. 2023. "Maytenus octogona Superoxide Scavenging and Anti-Inflammatory Caspase-1 Inhibition Study Using Cyclic Voltammetry and Computational Docking Techniques" International Journal of Molecular Sciences 24, no. 13: 10750. https://doi.org/10.3390/ijms241310750