
Bioremediation Potential of Native Bacillus sp. Strains as a Sustainable Strategy for Cadmium Accumulation of Theobroma cacao in Amazonas Region
 , ,
, ,  ,
,  and
and
Abstract
:1. Introduction
2. Methodology
2.1. Soil Samples Collection
2.2. Bacterial Isolation
2.3. Cadmium Tolerance Phenotypic Assay and Morphological Characterization
2.4. BOX Fingerprinting and 16S rRNA Phylogenetic Identification
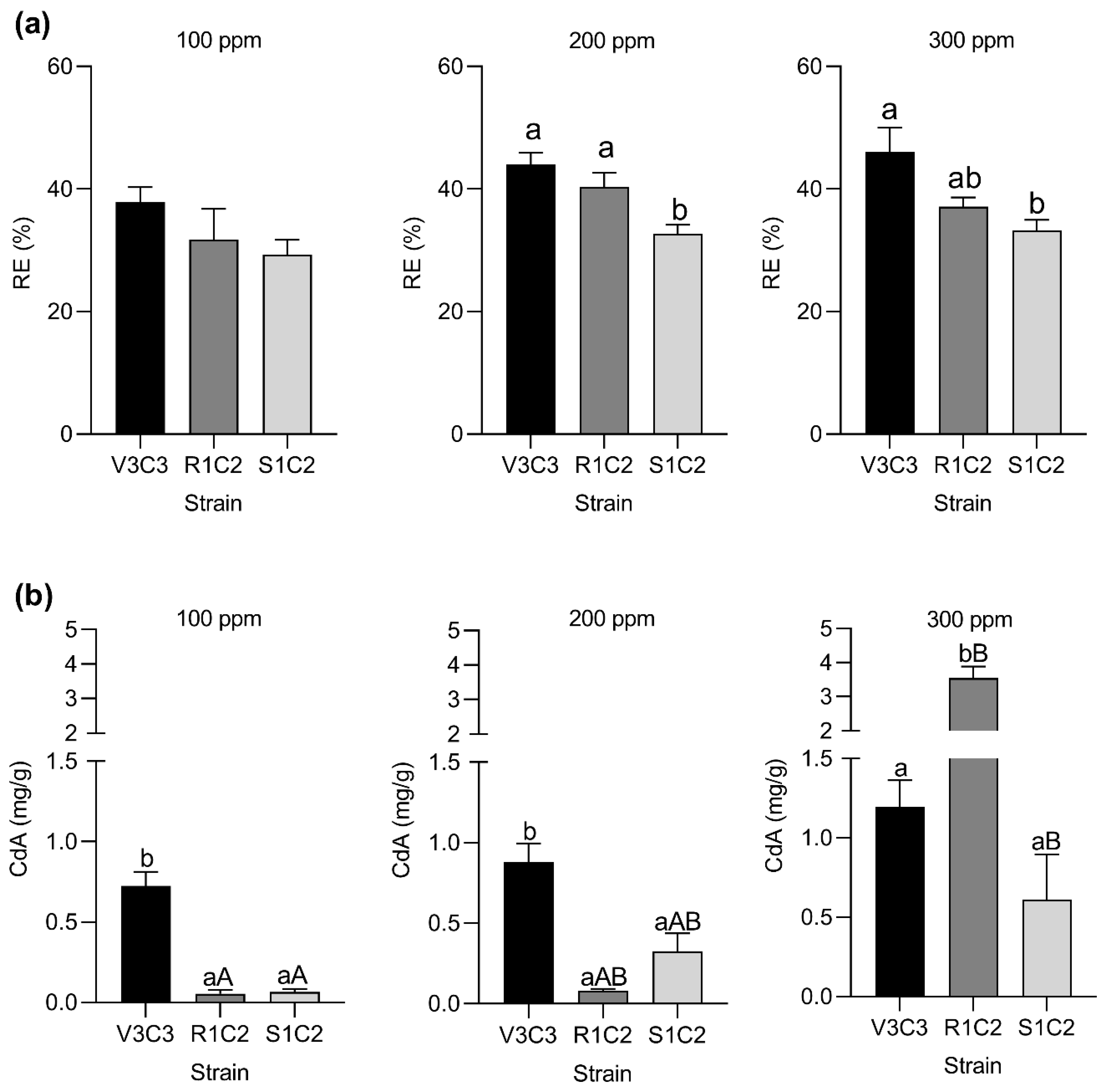
2.5. In Vitro Removal Cadmium Efficiency
2.6. Intracellular Cadmium Bioaccumulation Capacity
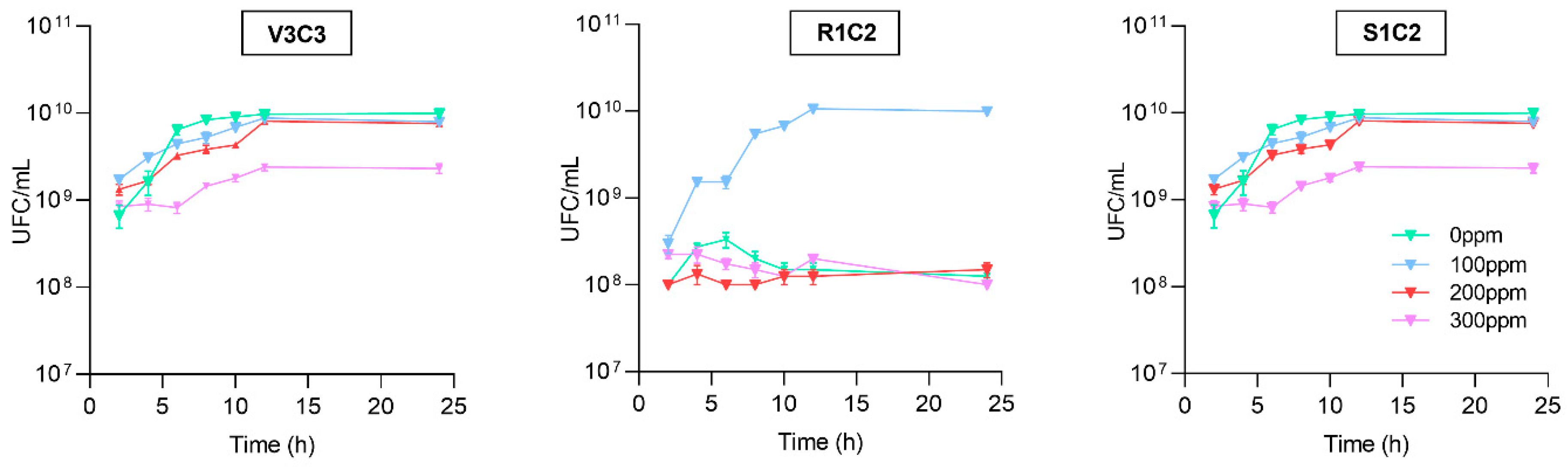
2.7. Bacterial Growth Kinetic
2.8. Plant Growth Promoting Traits Characterization
2.9. Growth Plant Promotion Capacity under Cadmium Conditions
2.10. Data Analysis
3. Results
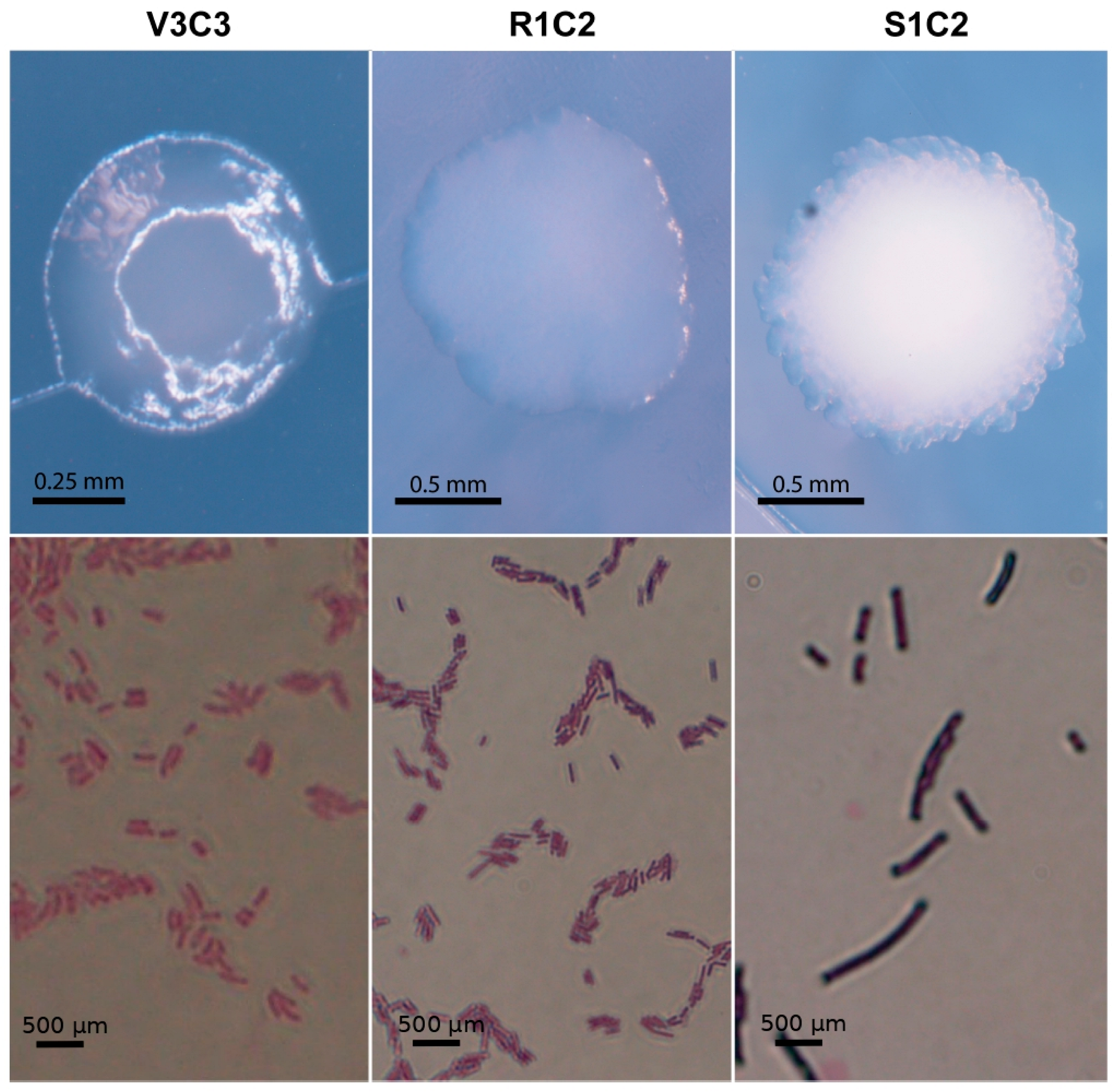
3.1. Cadmium Tolerant Bacteria
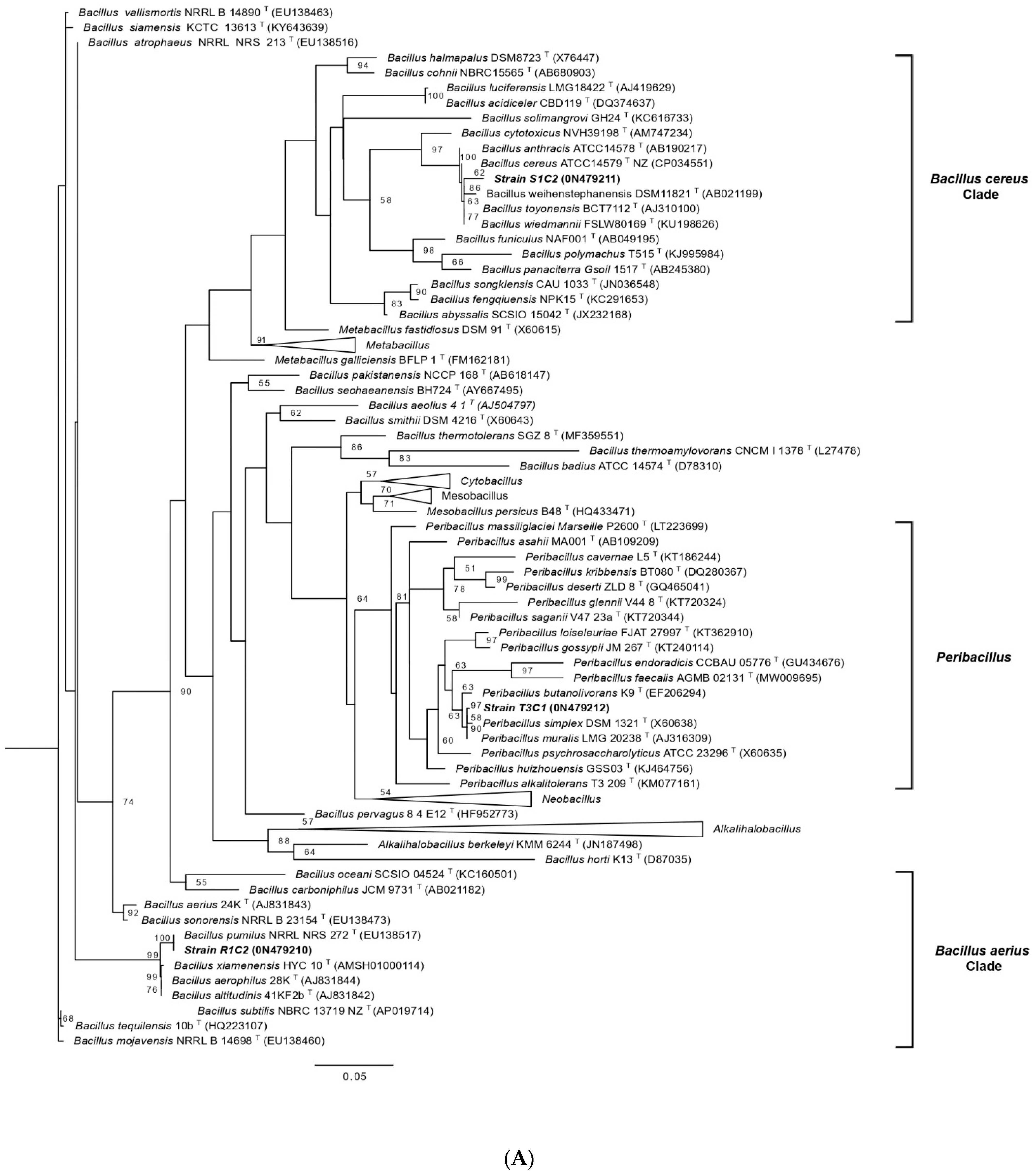
3.2. Bacterial Identification
3.3. Bacterial Behavior under Cadmium Toxicity and Complementary PGPR Traits
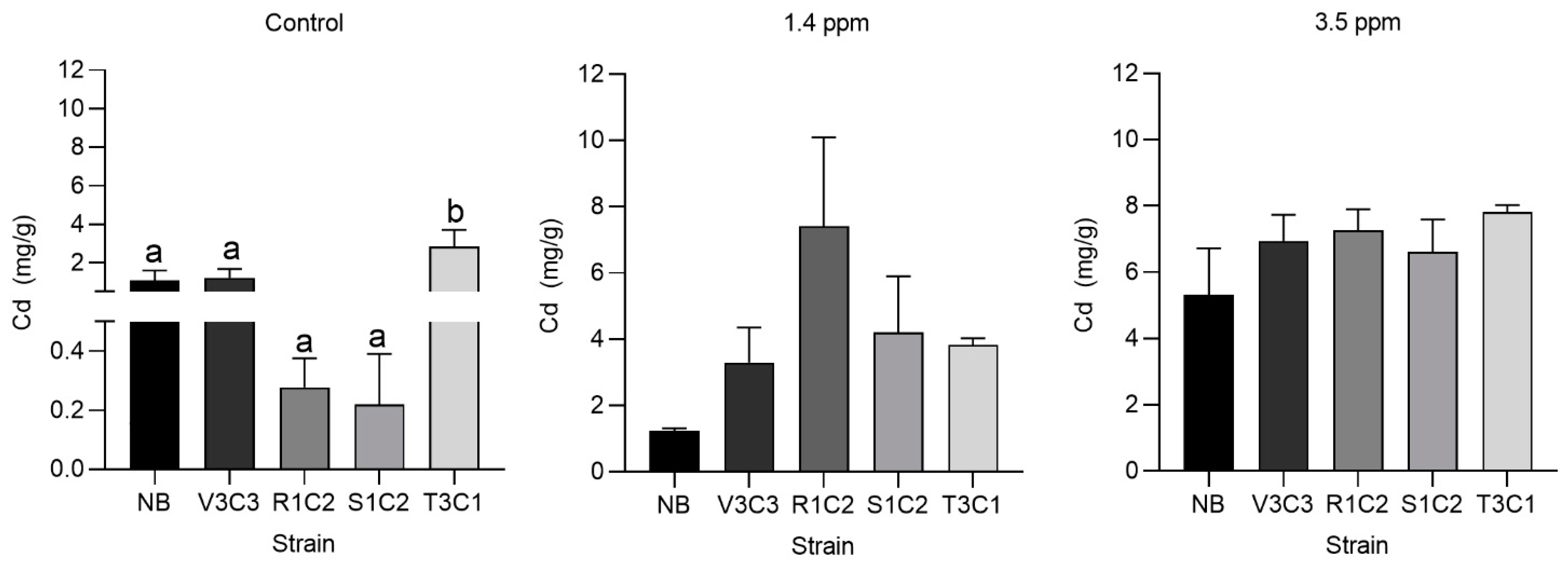
3.4. Effect of Selected Strains Inoculation on the Growing Variables and Cadmium Accumulation of Cacao
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sarkar, A.; Ravindran, G.; Krishnamurthy, V. A Brief Review on the Effect of Cadmium Toxicity: From Cellular to Organ Level. Int. J. Biol.-Technol. Res. 2013, 3, 17–36. [Google Scholar]
- Huang, Y.; He, C.; Shen, C.; Guo, J.; Mubeen, S.; Yuan, J.; Yang, Z. Toxicity of Cadmium and Its Health Risks from Leafy Vegetable Consumption. Food Funct. 2017, 8, 1373–1401. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Plant, J.A.; Voulvoulis, N.; Oates, C.J.; Ihlenfeld, C. Cadmium Levels in Europe: Implications for Human Health. Environ. Geochem. Health 2010, 32, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Sharma, H.; Rawal, N.; Mathew, B.B. The Characteristics, Toxicity and Effects of Cadmium. Int. J. Nanotechnol. Nanosci. 2015, 3, 1–9. [Google Scholar]
- Rahimzadeh, M.R.; Kazemi, S.; Moghadamnia, A.A. Cadmium Toxicity and Treatment: An Update. Casp. J. Intern. Med. 2017, 8, 135–145. [Google Scholar] [CrossRef]
- European Food Safety Authority Commission Regulation (EU) N° 488/2014. 2011. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX:32014R0488 (accessed on 15 July 2022).
- Mercosur Reglamento Técnico Mercosur Sobre Límites Máximos de Contaminantes Inorgánicos En Alimentos (Derogación De Las Res. GMC N° 102/94 y N° 36/96). 2011. Available online: http://www.puntofocal.gov.ar/doc/r_gmc_12-11.pdf (accessed on 25 April 2022).
- USDA. Foreign Agricultural Service China Releases the Standard for Maximum Levels of Contaminants in Foods; Beijing. 2018. Available online: https://www.fsis.usda.gov/sites/default/files/media_file/2021-02/GB-2762-2017.pdf (accessed on 25 April 2022).
- Chaney, R.L. Cadmium and Zinc in Trace Elements in Soils; Hooda, P.S., Ed.; Willey Online Library: Hoboken, NJ, USA, 2010. [Google Scholar] [CrossRef]
- ICCO International Cocoa Organization. 2018. Available online: https://www.icco.org/wp-content/uploads/FFP-5-2-Rev.1-A-Working-Definition-of-Fine-or-Flavour-Cocoa-English.pdf (accessed on 25 April 2022).
- Gomez-juaristi, M.; Torres, L.G.; Bravo, L.; Vaquero, M.P.; Bastida, S.; Sánchez-Muniz, F.J. Beneficial Effects of Chocolate on Cardiovascular Health. Nutr. Hosp. 2011, 26, 289–292. [Google Scholar] [CrossRef] [PubMed]
- Martín, M.Á.; Ramos, S. Health Beneficial Effects of Cocoa Phenolic Compounds: A Mini-Review. Curr. Opin. Food Sci. 2017, 14, 20–25. [Google Scholar] [CrossRef] [Green Version]
- Anyimah-ackah, E.; Ofosu, I.W.; Lutterodt, H.E.; Darko, G. Exposures and Risks of Arsenic, Cadmium, Lead, and Mercury in Cocoa Beans and Cocoa-Based Foods: A Systematic Review. Food Qual. Saf. 2018, 3, 1–8. [Google Scholar] [CrossRef]
- Yanus, R.L.; Sela, H.; Borojovich, E.J.C.; Zakon, Y.; Saphier, M.; Nikolski, A.; Gutflai, E.; Lorber, A.; Karpas, Z. Trace Elements in Cocoa Solids and Chocolate: An ICPMS Study. Talanta 2014, 119, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Alloway, B.J. Heavy Metals in Soils Trace: Metals and Metalloids in Soils and Their Bioavailability; Alloway, B.J., Ed.; Springer: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Vanderschueren, R.; Argüello, D.; Blommaert, H.; Montalvo, D.; Barraza, F.; Maurice, L.; Schreck, E.; Schulin, R.; Lewis, C.; Luis, J.; et al. Mitigating the Level of Cadmium in Cacao Products: Reviewing the Transfer of Cadmium from Soil to Chocolate Bar. Sci. Total Environ. 2021, 781, 146779. [Google Scholar] [CrossRef]
- Bertoldi, D.; Barbero, A.; Camin, F.; Caligiani, A.; Larcher, R. Multielemental Fi Ngerprinting and Geographic Traceability of Theobroma cacao Beans and Cocoa Products. Food Control 2016, 65, 46–53. [Google Scholar] [CrossRef]
- Abt, E.; Sam, J.F.; Gray, P.; Robin, L.P. Cadmium and Lead in Cocoa Powder and Chocolate Products in the U.S. Market. Food Addit. Contam. Part B 2018, 11, 92–102. [Google Scholar] [CrossRef]
- Fechner, C.; Greiner, M.; Heseker, H.; Lindtner, O. Dietary Exposure Assessment of Aluminium and Cadmium from Cocoa in Relation to Cocoa Origin. PLoS ONE 2019, 14, e0217990. [Google Scholar] [CrossRef] [PubMed]
- Kruszewski, B.; Obiedziński, M.W.; Kowalska, J. Nickel, Cadmium and Lead Levels in Raw Cocoa and Processed Chocolate Mass Materials from Three Different Manufacturers. J. Food Compos. Anal. 2018, 66, 127–135. [Google Scholar] [CrossRef]
- Mounicou, S.; Szpunar, J.; Andrey, D.; Blake, C.; Lobinski, R. Concentrations and Bioavailability of Cadmium and Lead in Cocoa Powder and Related Products Concentrations. Food Addit. Contam. 2003, 20, 343–352. [Google Scholar] [CrossRef]
- MIDAGRI “Ministerio de Desarrollo Agrario y Riego”. Producción Nacional de Cacao En Grano Creció En La Última Década a Un Promedio de 12.6% Al Año. 2020. Available online: https://www.gob.pe/institucion/midagri/noticias/305143-produccion-nacional-de-cacao-en-grano-crecio-en-la-ultima-decada-a-un-promedio-de-12-6-al-año (accessed on 29 April 2022).
- Arévalo-gardini, E.; Arévalo-hernández, C.O.; Baligar, V.C.; He, Z.L. Heavy Metal Accumulation in Leaves and Beans of Cacao (Theobroma Cacao L.) in Major Cacao Growing Regions in Peru. Sci. Total Environ. 2017, 605–606, 792–800. [Google Scholar] [CrossRef] [PubMed]
- Rosales-Huamani, J.A.; Breña-Ore, J.L.; Sespedes-Varkarsel, S.; De La Cuba, L.H.; Centeno-Rojas, L.; Otiniano-Zavala, A.; Andrade-Choque, J.; Valverde-Espinoza, S.; Castillo-Sequera, J.L. Study to Determine Levels of Cadmium in Cocoa Crops Applied to Inland Areas of Peru: The Case of the Campo Verde-Honoria Tournavista Corridor. Agronomy 2020, 10, 1576. [Google Scholar] [CrossRef]
- Oliva, M.; Rubio, K.; Epquin, M.; Marlo, G.; Leiva, S. Cadmium Uptake in Native Cacao Trees in Agricultural Lands of Bagua, Peru. Agronomy 2020, 10, 1551. [Google Scholar] [CrossRef]
- Gupta, G.; Parihar, S.S.; Ahirwar, N.K.; Snehi, S.K.; Singh, V. Plant Growth Promoting Rhizobacteria (PGPR): Current and Future Prospects for Development of Sustainable Agriculture. Microb. Biochem. Technol. 2015, 7, 96–102. [Google Scholar] [CrossRef]
- Nanda, M.; Kumar, V.; Sharma, D.K. Multimetal Tolerance Mechanisms in Bacteria: The Resistance Strategies Acquired by Bacteria That Can Be Exploited to ‘Clean-up’ Heavy Metal Contaminants from Water. Aquat. Toxicol. 2019, 212, 1–10. [Google Scholar] [CrossRef]
- Bravo, D.; Pardo-Díaz, S.; Benavides-Erazo, J.; Rengifo-Estrada, G.; Braissant, O.; Leon-Moreno, C. Cadmium and Cadmium-tolerant Soil Bacteria in Cacao Crops from Northeastern Colombia. J. Appl. Microbiol. 2018, 124, 1175–1194. [Google Scholar] [CrossRef]
- Bravo, D.; Braissant, O. Cadmium-Tolerant Bacteria: Current Trends and Applications in Agriculture. Lett. Appl. Microbiol. 2022, 74, 311–333. [Google Scholar] [CrossRef] [PubMed]
- Aryal, M. A Comprehensive Study on the Bacterial Biosorption of Heavy Metals: Materials, Performances, Mechanisms, and Mathematical Modellings. Rev. Chem. Eng. 2021, 37, 715–754. [Google Scholar] [CrossRef]
- Ghosh, S.; Bhattacharya, J.; Nitnavare, R.; Webster, T.J. Heavy Metal Removal by Bacillus for Sustainable Agriculture; Springer: Berlin/Heidelberg, Germany, 2022. [Google Scholar]
- Li, X.; Li, D.; Yan, Z.; Ao, Y. Biosorption and Bioaccumulation Characteristics of Cadmium by Plant Growth-Promoting Rhizobacteria. RSC Adv. 2018, 8, 30902–30911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbas, S.Z.; Rafatullah, M.; Hossain, K.; Ismail, N.; Tajarudin, H.A.; Abdul Khalil, H.P.S. A Review on Mechanism and Future Perspectives of Cadmium-Resistant Bacteria. Int. J. Environ. Sci. Technol. 2018, 15, 243–262. [Google Scholar] [CrossRef]
- Siripornadulsil, S.; Siripornadulsil, W. Cadmium-Tolerant Bacteria Reduce the Uptake of Cadmium in Rice: Potential for Microbial Bioremediation. Ecotoxicol. Environ. Saf. 2013, 94, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Feria-Cáceres, P.F.; Penagos-Velez, L.; Moreno-Herrera, C.X. Tolerance and Cadmium (Cd) Immobilization by Native Bacteria Isolated in Cocoa Soils with Increased Metal Content. Microbiol. Res. 2022, 13, 556–573. [Google Scholar] [CrossRef]
- Quiroga-Mateus, R.; López-Zuleta, S.; Chávez, E.; Bravo, D. Cadmium-Tolerant Bacteria in Cacao Farms from Antioquia, Colombia: Isolation, Characterization and Potential Use to Mitigate Cadmium Contamination. Processes 2022, 10, 1457. [Google Scholar] [CrossRef]
- Segura, A.; Ramos, J.L. Plant—Bacteria Interactions in the Removal of Pollutants. Curr. Opin. Biotechnol. 2013, 24, 467–473. [Google Scholar] [CrossRef] [PubMed]
- Kuiper, I.; Bloemberg, G.V.; Lugtenberg, B.J.J. Selection of a Plant-Bacterium Pair as a Novel Tool for Rhizostimulation of Polycyclic Aromatic Hydrocarbon-Degrading Bacteria. Mol. Plant-Microbe Interact. 2001, 14, 1197–1205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ministerio del Ambiente del Peú Resolución Ministerial N° 307_2012_MINAM. 2012. Available online: https://www.minam.gob.pe/wp-content/uploads/2013/09/rm_307-1.pdf (accessed on 29 April 2022).
- Bashir, A.H. Diversity and Morphology of Bacterial Community Characterized in Topsoil Samples from the Gaza Strip, Palestine. Res. J. Microbiol. 2012, 7, 309–318. [Google Scholar] [CrossRef]
- Dos Santos, H.R.M.; Argolo, C.S.; Argôlo-Filho, R.C.; Loguercio, L.L. A 16S RDNA PCR-Based Theoretical to Actual Delta Approach on Culturable Mock Communities Revealed Severe Losses of Diversity Information. BMC Microbiol. 2019, 19, 74. [Google Scholar] [CrossRef] [PubMed]
- Sannazzaro, A.I.; Bergottini, V.M.; Paz, R.C.; Castagno, L.N.; Menéndez, A.B.; Ruiz, O.A.; Pieckenstain, F.L.; Estrella, M.J. Comparative Symbiotic Performance of Native Rhizobia of the Flooding Pampa and Strains Currently Used for Inoculating Lotus tenuis in This Region. Antonie Van Leeuwenhoek 2011, 99, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Cumpa-Velásquez, L.M.; Moriconi, J.I.; Dip, D.P.; Castagno, L.N.; Puig, M.L.; Maiale, S.J.; Santa-María, G.E.; Sannazzaro, A.I.; Estrella, M.J. Prospecting Phosphate Solubilizing Bacteria in Alkaline-Sodic Environments Reveals Intra-Specific Variability in Pantoea eucalypti Affecting Nutrient Acquisition and Rhizobial Nodulation in Lotus tenuis. Appl. Soil Ecol. 2021, 168, 104125. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information (NCBI). Available online: https://www.ncbi.nlm.nih.gov (accessed on 20 May 2022).
- The Ribosomal Database Project (RDP). Available online: http://rdp.cme.msu.edu/ (accessed on 20 May 2022).
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Shi, Z.; Zhang, Z.; Yuan, M.; Wang, S.; Yang, M.; Yao, Q.; Ba, W.; Zhao, J.; Xie, B. Characterization of a High Cadmium Accumulating Soil Bacterium, Cupriavidus Sp. WS2. Chemosphere 2020, 247, 125834. [Google Scholar] [CrossRef]
- Pandey, N.; Bhatt, R. Arsenic Resistance and Accumulation by Two Bacteria Isolated from a Natural Arsenic Contaminated Site. J. Basic Microbiol. 2015, 55, 1275–1286. [Google Scholar] [CrossRef]
- Sharma, P.; Kumawat, K.C.; Kaur, S.; Kaur, N. Assessment of Zinc Solubilization by Endophytic Bacteria in Legume Rhizosphere. Indian J. Appl. Res. 2014, 4, 439–441. [Google Scholar] [CrossRef]
- Di Rienzo, J.A.; Balzarini, M.G.; Tablada, M.; Gonzalez, L.A.; Casanoves, F. InfoStat: Manual Del Usuario, Primera ed.; Editorial Brujas: Córdoba, Argentina, 2008. [Google Scholar]
- Patel, S.; Gupta, R.S. A Phylogenomic and Comparative Genomic Framework for Resolving the Polyphyly of the Genus Bacillus: Proposal for Six New Genera of Bacillus Species, Peribacillus Gen. Nov., Cytobacillus Gen. Nov., Mesobacillus Gen. Nov., Neobacillus Gen. Nov. Int. J. Syst. Evol. Microbiol. 2020, 70, 406–438. [Google Scholar] [CrossRef]
- Chaturvedi, A.D.; Pal, D.; Penta, S.; Kumar, A. Ecotoxic Heavy Metals Transformation by Bacteria and Fungi in Aquatic Ecosystem. World J. Microbiol. Biotechnol. 2015, 31, 1595–1603. [Google Scholar] [CrossRef]
- Al-Gheethi, A.A.S.; Lalung, J.; Noman, E.A.; Bala, J.D.; Norli, I. Removal of Heavy Metals and Antibiotics from Treated Sewage Effluent by Bacteria. Clean Technol. Environ. Policy 2015, 17, 2101–2123. [Google Scholar] [CrossRef]
- Priyadarshanee, M.; Das, S. Biosorption and Removal of Toxic Heavy Metals by Metal Tolerating Bacteria for Bioremediation of Metal Contamination: A Comprehensive Review. J. Environ. Chem. Eng. 2020, 9, 104686. [Google Scholar] [CrossRef]
- Rajkumar, M.; Ae, N.; Freitas, H. Chemosphere Endophytic Bacteria and Their Potential to Enhance Heavy Metal Phytoextraction. Chemosphere 2009, 77, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Cáceres, P.F.F.; Vélez, L.P.; Junca, H.; Moreno-Herrera, C.X. Theobroma cacao L. Agricultural Soils with Natural Low and High Cadmium (Cd) in Santander (Colombia), Contain a Persistent Shared Bacterial Composition Shaped by Multiple Soil Variables and Bacterial Isolates Highly Resistant to Cd Concentrations. Curr. Res. Microb. Sci. 2021, 2, 100086. [Google Scholar] [CrossRef] [PubMed]
- Cordoba-novoa, H.A.; Torres-rojas, E.; Sciences, A. Assessment of Native Cadmium-Resistant Bacteria in Cacao. BioRxiv 2021. [Google Scholar] [CrossRef]
- Chen, Y.P.; Tsai, C.F.; Rekha, P.D.; Ghate, S.D.; Huang, H.Y.; Hsu, Y.H.; Liaw, L.L.; Young, C. Agricultural Management Practices Influence the Soil Enzyme Activity and Bacterial Community Structure in Tea Plantations. Bot. Stud. 2021, 62, 8. [Google Scholar] [CrossRef]
- Li, W.-H.; Liu, Q.-Z.; Chen, P. Effect of Long-Term Continuous Cropping of Strawberry on Soil Bacterial Community Structure and Diversity. J. Integr. Agric. 2018, 17, 2570–2582. [Google Scholar] [CrossRef]
- Chu, D. Effects of Heavy Metals on Soil Microbial Community. Earth Environ. Sci. 2018, 113, 012009. [Google Scholar] [CrossRef]
- Gomes, N.C.M.; Landi, L.; Smalla, K.; Nannipieri, P.; Brookes, P.C.; Renella, G. Effects of Cd- and Zn-Enriched Sewage Sludge on Soil Bacterial and Fungal Communities. Ecotoxicol. Environ. Saf. 2010, 73, 1255–1263. [Google Scholar] [CrossRef]
- Marques, A.P.G.C.; Moreira, H.; Franco, A.R.; Rangel, A.O.S.S.; Castro, P.M.L. Inoculating Helianthus annuus (Sunflower) Grown in Zinc and Cadmium Contaminated Soils with Plant Growth Promoting Bacteria—Effects on Phytoremediation Strategies. Chemosphere 2013, 92, 74–83. [Google Scholar] [CrossRef]
- Xu, M.; Hao, X.; Xiong, Z.; Liao, H.; Wang, L.; Zhang, T.; Luo, X.; Chen, W.; Huang, Q. Soil Amendments Change Bacterial Functional Genes More than Taxonomic Structure in a Cadmium-Contaminated Soil. Soil Biol. Biochem. 2021, 154, 108126. [Google Scholar] [CrossRef]
- Lata, S.; Mishra, T.; Kaur, S. Cadmium Bioremediation Potential of Bacillus sp. and Cupriavidus sp. J. Pure Appl. Microbiol. 2021, 15, 1665–1680. [Google Scholar] [CrossRef]
- Sheng, X.; He, L.; Wang, Q.; Ye, H.; Jiang, C. Effects of Inoculation of Biosurfactant-Producing Bacillus Sp. J119 on Plant Growth and Cadmium Uptake in a Cadmium-Amended Soil. J. Hazard. Mater. 2008, 155, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Koua, S.H.; Coulibaly, D.; Waze, N.; Alloue, M.; Fernique, B. Bacillus subtilis Strains Isolated from Cocoa Trees (Theobroma cacao L.) Rhizosphere for Their Use as Potential Plant Growth Promoting Rhizobacteria in Côte d’ Ivoire. Curr. Microbiol. 2020, 77, 2258–2264. [Google Scholar] [CrossRef]
- Leite, H.A.C.; Silva, A.B.; Gomes, F.P.; Gramacho, K.P.; Faria, J.C.; de Souza, J.T.; Loguercio, L.L. Bacillus subtilis and Enterobacter cloacae Endophytes from Healthy Theobroma Cacao L. Trees Can Systemically Colonize Seedlings and Promote Growth. Appl. Microb. CELL Physiol. Bacillus 2013, 97, 2639–2651. [Google Scholar] [CrossRef]
- Melnick, R.L.; Suárez, C.; Bailey, B.A.; Backman, P.A. Isolation of Endophytic Endospore-Forming Bacteria from Theobroma Cacao as Potential Biological Control Agents of Cacao Diseases. Biol. Control 2011, 57, 236–245. [Google Scholar] [CrossRef]
- Thomas, L.; Gupta, A.; Gopal, M.; George, P.; Thomas, G.V. Efficacy of Rhizospheric Bacillus spp. for Growth Promotion in Theobroma cacao L. Seedlings. J. Plant. Crop. 2011, 39, 19–25. [Google Scholar]
- Valentine, N.B.; Bolton, H.; Kingsley, M.T.; Drake, G.R.; Balkwilf, D.L.; Plymale, A.E. Biosorption of Cadmium, Cobalt, Nickel, and Strontium by a Bacillus Simplex Strain Isolated from the Vadose Zone. J. Ind. Microbiol. 1996, 16, 189–196. [Google Scholar] [CrossRef]
- Vijayaraghavan, K.; Yun, Y. Bacterial Biosorbents and Biosorption. Biotechnol. Adv. 2008, 26, 266–291. [Google Scholar] [CrossRef]
- Alotaibi, B.S.; Khan, M.; Shamim, S. Unraveling the Underlying Heavy Metal Detoxification Mechanisms of Bacillus Species. Microorganisms 2021, 9, 1628. [Google Scholar] [CrossRef]
- Njoku, K.L.; Akinyede, O.R.; Obidi, O.F. Microbial Remediation of Heavy Metals Contaminated Media by Bacillus megaterium and Rhizopus stolonifer. Sci. Afr. 2020, 10, e00545. [Google Scholar] [CrossRef]
- Ding, J.; Chen, W.; Zhang, Z.; Qin, F.; Jiang, J.; He, A.; Sheng, G.D. Enhanced Removal of Cadmium from Wastewater with Coupled Biochar and Bacillus subtilis. Water Sci. Technol. 2021, 83, 2075–2086. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Dang, Z.; Guo, C.; Lu, G.; Gu, R.R.; Liu, H.; Zhang, H. Biosorption of Cd (II) by Live and Dead Cells of Bacillus cereus RC-1 Isolated from Cadmium-Contaminated Soil. Colloids Surf. B Biointerfaces 2013, 107, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.L.; Michels, P.C.; Dawson, S.C.; Kitisakkul, S.; Baross, J.A.; Keasling, J.D.; Clark, D.S. Cadmium Removal by a New Strain of Pseudomonas aeruginosa in Aerobic Culture. Appl. Environ. Microbiol. 1997, 63, 4075–4078. [Google Scholar] [CrossRef] [Green Version]
- Pramanik, K.; Mandal, S.; Banerjee, S.; Ghosh, A.; Kanti, T.; Chandra, N. Chemosphere Unraveling the Heavy Metal Resistance and Biocontrol Potential of Pseudomonas sp. K32 Strain Facilitating Rice Seedling Growth under Cd Stress. Chemosphere 2021, 274, 129819. [Google Scholar] [CrossRef]
- Hrynkiewicz, K.; Kowalkowski, T. Strain-Specific Bioaccumulation and Intracellular Distribution of Cd2+ in Bacteria Isolated from the Rhizosphere, Ectomycorrhizae, and Fruitbodies of Ectomycorrhizal Fungi. Environ. Sci. Pollut. Res. 2015, 22, 3055–3067. [Google Scholar] [CrossRef] [Green Version]
- Li, W.-W.; Yu, H.-Q. Insight into the Roles of Microbial Extracellular Polymer Substances in Metal Biosorption. Bioresour. Technol. 2014, 160, 15–23. [Google Scholar] [CrossRef]
- Lian, Z.; Yang, Z.; Song, W.; Sun, M.; Gan, Y.; Bai, X. Effects of Different Exogenous Cadmium Compounds on the Chemical Composition and Adsorption Properties of Two Gram-Negative Bacterial EPS. Sci. Total Environ. 2022, 806, 150511. [Google Scholar] [CrossRef]
- Xie, Y.; He, N.; Wei, M.; Wen, T.; Wang, X.; Liu, H.; Zhong, S.; Xu, H. Cadmium Biosorption and Mechanism Investigation Using a Novel Bacillus subtilis KC6 Isolated from Pyrite Mine. J. Clean. Prod. 2021, 312, 127749. [Google Scholar] [CrossRef]
- Huang, F.; Guo, C.; Lu, G.; Yi, X.; Zhu, L.; Dang, Z. Bioaccumulation Characterization of Cadmium by Growing Bacillus cereus RC-1 and Its Mechanism. Chemosphere 2014, 109, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Wang, Z.; Cai, Y.; Chen, S.; Tian, J.; Cai, K.-Z. Ecotoxicology and Environmental Safety Heavy Metal Bioaccumulation and Cation Release by Growing Bacillus cereus RC-1 under Culture Conditions. Ecotoxicol. Environ. Saf. 2018, 157, 216–226. [Google Scholar] [CrossRef] [PubMed]
- Hui, C.; Guo, Y.; Liu, L.; Yi, J. Recent Advances in Bacterial Biosensing and Bioremediation of Cadmium Pollution: A Mini-Review. World J. Microbiol. Biotechnol. 2022, 38, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Blindauer, C.A. Bacterial Metallothioneins: Past, Present, and Questions for the Future. J. Biol. Inorg. Chem. 2011, 16, 1011–1024. [Google Scholar] [CrossRef] [PubMed]
- Habjanič, J.; Mathew, A.; Eberl, L.; Freisinger, E. Deciphering the Enigmatic Function of Pseudomonas Metallothioneins. Front. Microbiol. 2020, 11, 1709. [Google Scholar] [CrossRef] [PubMed]
- Higham, D.P.; Sadler, P.J.; Scawen, M.D. Cadmium-Resistant Pseudomonas putida Synthesizes Novel Cadmium Proteins. Science 1984, 225, 1043–1046. [Google Scholar] [CrossRef]
- Huckle, J.W.; Morby, A.P.; Turner, J.S.; Robinson, N.J. Isolation of a Prokaryotic Metallothionein Locus and Analysis of Transcriptional Control by Trace Metal Ions. Mol. Microbiol. 1993, 7, 177–187. [Google Scholar] [CrossRef]
- Chavez, E.; He, Z.L.; Stoffella, P.J.; Mylavarapu, R.; Li, Y.; Baligar, V.C. Evaluation of Soil Amendments as a Remediation Alternative for Cadmium-Contaminated Soils under Cacao Plantations. Environ. Sci. Pollut. Res. 2016, 23, 17571–17580. [Google Scholar] [CrossRef]
- Ramtahal, G.; Umaharan, P.; Hanuman, A.; Davis, C.; Ali, L. The Effectiveness of Soil Amendments, Biochar and Lime, in Mitigating Cadmium bioaccumulation in Theobroma cacao L. Sci. Total Environ. 2019, 693, 133563. [Google Scholar] [CrossRef]
- Ramtahal, G.; Umaharan, P.; Davis, C.; Roberts, C.; Hanuman, A.; Ali, L. Mitigation of Cadmium Uptake in Theobroma cacao L.: Efficacy of Soil Application Methods of Hydrated Limeand Biochar. Plant Soil 2022, 477, 281–296. [Google Scholar] [CrossRef]
- Moncada, U.A.P.; Gómez, M.R.; Ordoñez, D.P.S.; Rolón, A.M.P.; Ortiz, W.A.W.; Ramírez, L.; Estrada, G.A.R. Arbuscular Mycorrhizal Fungi (AMF) as a Strategy to Reduce the Absorption of Cadmium in Cocoa (Theobroma cacao) Plants. Terra Latinoam. 2019, 37, 121–130. [Google Scholar] [CrossRef]
- Shahzad, A.; Qin, M.; Elahie, M.; Naeem, M.; Bashir, T.; Yasmin, H.; Younas, M.; Areeb, A.; Irfan, M.; Billah, M.; et al. Bacillus pumilus Induced Tolerance of Maize (Zea mays L.) against Cadmium (Cd) Stress. Sci. Rep. 2021, 11, 17196. [Google Scholar] [CrossRef] [PubMed]
- Jan, M.; Shah, G.; Masood, S.; Shinwari, K.I.; Hameed, R.; Rha, E.S.; Jamil, M. Bacillus cereus enhanced Phytoremediation Ability of Rice Seedlings under Cadmium Toxicity. BioMed Res. Int. 2019, 2019, 8134651. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Liu, Z.; Huang, Y.; Zhang, Y.; Wang, X.; Hu, Z. Cadmium-Resistant Rhizobacterium Bacillus cereus M4 Promotes the Growth and Reduces Cadmium Accumulation in Rice (Oryza sativa L.). Environ. Toxicol. Pharmacol. 2019, 72, 103265. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Zhu, L.; Ma, Z.; Wang, J. Bacillus Amyloliquefaciens SAY09 Increases Cadmium Resistance in Plants by Activation of Auxin-Mediated. Genes 2017, 8, 173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahile, A.A.; Khan, M.A.; Hamayun, M.; Imran, M.; Kang, S.; Lee, I. Novel Bacillus cereus strain, ALT1, Enhance Growth and Strengthens the Antioxidant System of Soybean under Cadmium Stress. Agronomy 2021, 11, 404. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Sample Site | Phenotype | Gram (+) | Gram (−) | |||
|---|---|---|---|---|---|---|---|
| Shape | Border | Colour | Elevation | ||||
| O2C1 | Aramango-O | C | E | nP | c | ● | |
| F1C3 | Aramango-F | C | E | nP | Fl | ● | |
| H1C2 | La peca-H | C | E | p | R | ● | |
| H2C1 | La peca-H | C | E | nP | c | ● | |
| H3C1 | La peca-H | C | E | nP | c | ● | |
| H4C1 | La peca-H | C | E | nP | c | ● | |
| H4C2 | La peca-H | P | E | nP | c | ● | |
| H5C1 | La peca-H | C | E | nP | c | ● | |
| H5C2 | La peca-H | C | E | nP | c | ● | |
| J1C2 | La peca-J | C | E | p | R | ● | |
| J2C1 | La peca-J | P | E | nP | R | ● | |
| J2C2 | La peca-J | C | E | p | c | ● | |
| J3C1 | La peca-J | C | E | p | c | ● | |
| J4C1 | La peca-J | I | C | nP | Fl | ● | |
| J4C2 | La peca-J | C | E | nP | R | ● | |
| J5C1 | La peca-J | C | E | nP | c | ● | |
| T2C1 | La peca-T | C | E | nP | Fl | ● | |
| T2C2 | La peca-T | I | C | nP | c | ● | |
| T3C1 | La peca-T | P | E | nP | R | ● | |
| T3C2 | La peca-T | C | E | p | c | ● | |
| T4C1 | La peca-T | C | E | p | c | ● | |
| T4C2 | La peca-T | I | C | nP | Fl | ● | |
| T5C1 | La peca-T | C | E | nP | R | ● | |
| T5C2 | La peca-T | C | E | nP | c | ● | |
| V3C3 | Copallin-V | C | E | nP | Fl | ● | |
| S1C2 | Copallin-S | I | C | nP | R | ● | |
| S5C1 | Copallin-S | C | E | p | c | ● | |
| Z5C1 | Copallin-Z | C | E | nP | c | ● | |
| C1C1 | Copallin-C | C | E | p | c | ● | |
| C1C2 | Copallin-C | C | E | p | c | ● | |
| C2C1 | Copallin-C | P | C | nP | R | ● | |
| C3C1 | Copallin-C | C | E | nP | R | ● | |
| C5C1 | Copallin-C | C | E | nP | c | ● | |
| M1C1 | Copallin-M | I | C | nP | Fl | ● | |
| M1C2 | Copallin-M | C | E | p | R | ● | |
| M2C1 | Copallin-M | P | E | nP | R | ● | |
| M3C1 | Copallin-M | P | E | nP | R | ● | |
| M4C1 | Copallin-M | C | E | p | c | ● | |
| M4C2 | Copallin-M | I | L | nP | R | ● | |
| M5C2 | Copallin-M | C | E | p | c | ● | |
| E1C2 | Copallin-E | C | E | p | c | ● | |
| E2C2 | Copallin-E | C | E | p | c | ● | |
| E3C1 | Copallin-E | C | E | p | R | ● | |
| R1C1 | Copallin-R | C | E | nP | R | ● | |
| R1C2 | Copallin-R | I | L | nP | c | ● | |
| R2C1 | Copallin-R | C | E | nP | c | ● | |
| R2C2 | Copallin-R | C | E | p | c | ● | |
| R3C1 | Copallin-R | C | E | nP | R | ● | |
| R3C2 | Copallin-R | P | E | p | R | ● | |
| R4C1 | Copallin-R | C | E | nP | c | ● | |
| R5C2 | Copallin-R | F | F | nP | Fl | ● | |
| R5C3 | Copallin-R | P | E | p | R | ● | |
| Variable | Strain | ||||
|---|---|---|---|---|---|
| NB | V3C3 | R1C2 | S1C2 | T3C1 | |
| Control | |||||
| FWstem | 5.7 ± 1.61 | 5.64 ± 1.61 | 7.13 ± 1.8 | 7.3 ± 2.08 | 7.97 ± 2.08 |
| Fwroot | 1.96 ± 0.51 | 2.52 ± 0.51 | 2 ± 0.57 | 2.57 ± 0.66 | 1.77 ± 0.66 |
| DWstem | 1.96 ± 0.47 | 2.07 ± 0.47 | 2.18 ± 0.52 | 2.2 ± 0.6 | 2.51 ± 0.6 |
| Dwroot | 0.52 ± 0.16 | 0.65 ± 0.16 | 0.49 ± 0.18 | 0.61 ± 0.21 | 0.74 ± 0.21 |
| 1.4 ppm | |||||
| FWstem | 6.8 ± 2.83 | 10.88 ± 2.0 | 5.77 ± 2.31 | 13.12 ± 1.79 | 9.6 ± 2 |
| Fwroot | 2.05 ± 0.65 | 2.78 ± 0.46 | 1.7 ± 0.53 | 2.52 ± 0.41 | 2.23 ± 0.46 |
| DWstem | 2.15 ± 0.89 | 3.2 ± 0.63 | 1.71 ± 0.72 | 4.1 ± 0.56 | 3.23 ± 0.63 |
| Dwroot | 0.25 ± 0.17 | 0.78 ± 0.12 | 0.42 ± 0.14 | 0.7 ± 0.11 | 0.65 ± 0.12 |
| 3.5 ppm | |||||
| FWstem | 13.55 ± 1.54 b | 4.87 ± 1.54 a | 9.58 ± 1.34 ab | 13.55 ± 1.34 b | 11.9 ± 1.89 ab |
| Fwroot | 2.68 ± 0.54 | 1.95 ± 0.54 | 1.83 ± 0.54 | 2.72 ± 0.48 | 2.07 ± 0.62 |
| DWstem | 3.52 ± 0.67 | 1.75 ± 0.78 | 2.9 ± 0.67 | 3.83 ± 0.6 | 2.67 ± 0.78 |
| Dwroot | 0.86 ± 0.17 | 0.53 ± 0.17 | 0.52 ± 0.17 | 0.76 ± 0.15 | 0.61 ± 0.19 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arce-Inga, M.; González-Pérez, A.R.; Hernandez-Diaz, E.; Chuquibala-Checan, B.; Chavez-Jalk, A.; Llanos-Gomez, K.J.; Leiva-Espinoza, S.T.; Oliva-Cruz, S.M.; Cumpa-Velasquez, L.M. Bioremediation Potential of Native Bacillus sp. Strains as a Sustainable Strategy for Cadmium Accumulation of Theobroma cacao in Amazonas Region. Microorganisms 2022, 10, 2108. https://doi.org/10.3390/microorganisms10112108
Arce-Inga M, González-Pérez AR, Hernandez-Diaz E, Chuquibala-Checan B, Chavez-Jalk A, Llanos-Gomez KJ, Leiva-Espinoza ST, Oliva-Cruz SM, Cumpa-Velasquez LM. Bioremediation Potential of Native Bacillus sp. Strains as a Sustainable Strategy for Cadmium Accumulation of Theobroma cacao in Amazonas Region. Microorganisms. 2022; 10(11):2108. https://doi.org/10.3390/microorganisms10112108
Chicago/Turabian StyleArce-Inga, Marielita, Alex Ricardo González-Pérez, Elgar Hernandez-Diaz, Beimer Chuquibala-Checan, Antony Chavez-Jalk, Kelvin James Llanos-Gomez, Santos Triunfo Leiva-Espinoza, Segundo Manuel Oliva-Cruz, and Liz Marjory Cumpa-Velasquez. 2022. "Bioremediation Potential of Native Bacillus sp. Strains as a Sustainable Strategy for Cadmium Accumulation of Theobroma cacao in Amazonas Region" Microorganisms 10, no. 11: 2108. https://doi.org/10.3390/microorganisms10112108