
Cytoplasmic Male Sterility Incidence in Potato Breeding Populations with Late Blight Resistance and Identification of Breeding Lines with a Potential Fertility Restorer Mechanism

 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. Cytoplasmic Type Composition of Late Blight Resistant Populations
2.2. Effects of Cytoplasmic Type on Reproductive Traits
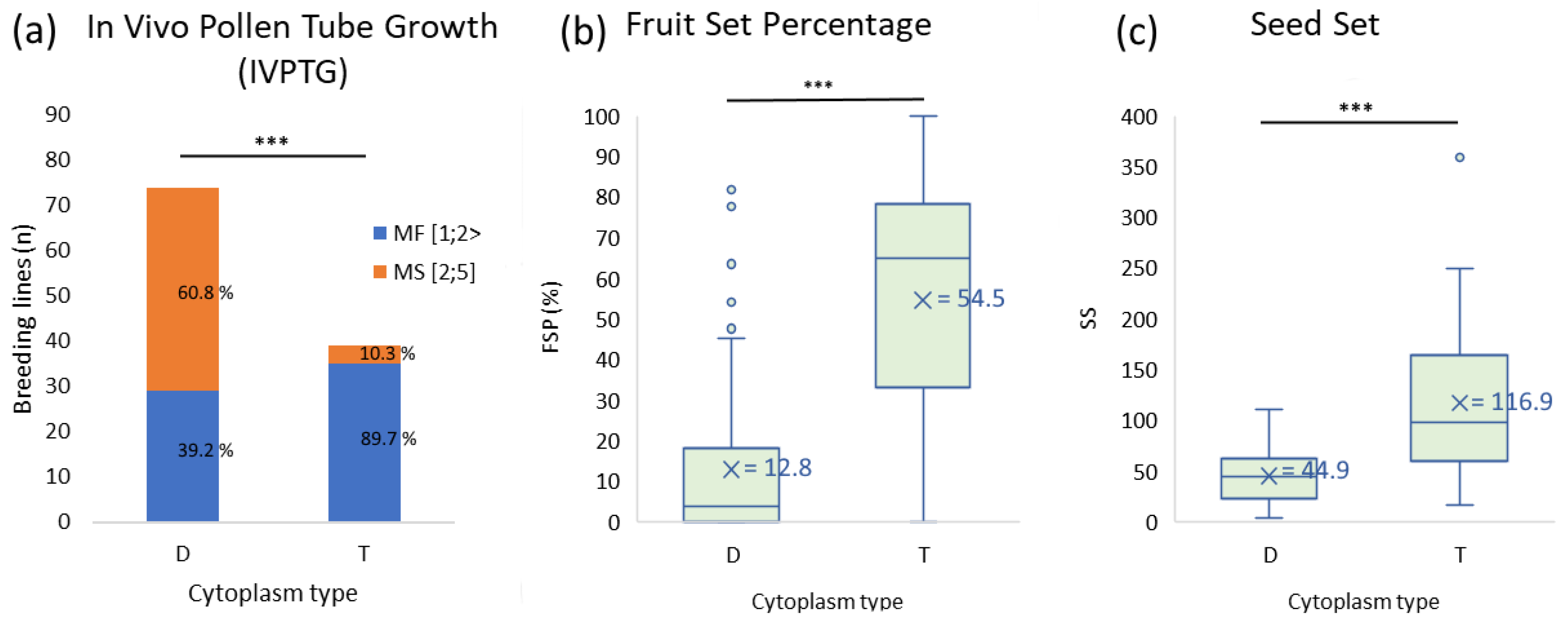
2.3. Effects of Cytoplasmic Type on Pollen Tube Germination and Fruit and Seed Production in Test Crosses
2.4. Correlations among Traits
2.5. Identification of Potential Fertility Restorer Breeding Lines
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Reproductive Trait Evaluation
4.3. Floral Abnormalities Evaluation
4.4. Test Cross Evaluation
4.5. Statistical Analysis and Software
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bradshaw, J.E. Breeding Diploid F1 Hybrid Potatoes for Propagation from Botanical Seed (TPS): Comparisons with Theory and Other Crops. Plants 2022, 11, 1121. [Google Scholar] [CrossRef] [PubMed]
- Busse, J.S.; Jansky, S.H.; Agha, H.I.; Carley, C.A.S.; Shannon, L.M.; Bethke, P.C. A High Throughput Method for Generating Dihaploids from Tetraploid Potato. Am. J. Potato Res. 2021, 98, 304–314. [Google Scholar] [CrossRef]
- Ordoñez, B.; Santayana, M.; Aponte, M.; Henry, I.M.; Comai, L.; Eyzaguirre, R.; Lindqvist-Kreuze, H.; Bonierbale, M. PL-4 (CIP596131.4): An Improved Potato Haploid Inducer. Am. J. Potato Res. 2021, 98, 255–262. [Google Scholar] [CrossRef]
- Eggers, E.-J.; van der Burgt, A.; van Heusden, S.A.W.; de Vries, M.E.; Visser, R.G.F.; Bachem, C.W.B.; Lindhout, P. Neofunctionalisation of the Sli gene leads to self-compatibility and facilitates precision breeding in potato. Nat. Commun. 2021, 12, 4141. [Google Scholar] [CrossRef]
- Hosaka, K.; Sanetomo, R. Creation of a highly homozygous diploid potato using the S locus inhibitor (Sli) gene. Euphytica 2020, 216, 169. [Google Scholar] [CrossRef]
- Jansky, S.H.; Charkowski, A.O.; Douches, D.S.; Gusmini, G.; Richael, C.; Bethke, P.C.; Spooner, D.M.; Novy, R.G.; De Jong, H.; De Jong, W.S.; et al. Reinventing Potato as a Diploid Inbred Line-Based Crop. Crop Sci. 2016, 56, 1412–1422. [Google Scholar] [CrossRef] [Green Version]
- Kardile, H.B.; Yilma, S.; Sathuvalli, V. Molecular Approaches to Overcome Self-Incompatibility in Diploid Potatoes. Plants 2022, 11, 1328. [Google Scholar] [CrossRef]
- Almekinders, C.J.M.; Chujoy, E.; Thiele, G. The Use of True Potato Seed as Pro-poor Technology: The Efforts of an International Agricultural Research Institute to Innovating Potato Production. Potato Res. 2009, 52, 275–293. [Google Scholar] [CrossRef] [Green Version]
- Anisimova, I.N.; Gavrilenko, T.A. Cytoplasmic Male Sterility and Prospects for Its Utilization in Potato Breeding, Genetic Studies and Hybrid Seed Production. Russ. J. Genet. Appl. Res. 2017, 7, 721–735. [Google Scholar] [CrossRef]
- Grun, P.; Ochoa, C.; Capage, D. Evolution of Cytoplasmic Factors in Tetraploid Cultivated Potatoes (Solanaceae). Am. J. Bot. 1977, 64, 412–420. [Google Scholar] [CrossRef]
- Frei, U.; Peiretti, E.G.; Wenzel, G. Significance of Cytoplasmic DNA in Plant Breeding. In Plant Breeding Reviews; John Wiley & Sons, Inc.: Oxford, UK, 2003; pp. 175–210. [Google Scholar]
- Bohra, A.; Jha, U.C.; Adhimoolam, P.; Bisht, D.; Singh, N.P. Cytoplasmic male sterility (CMS) in hybrid breeding in field crops. Plant Cell Rep. 2016, 35, 967–993. [Google Scholar] [CrossRef] [PubMed]
- Iwanaga, M.; Ortiz, R.; Cipar, M.S.; Peloquin, S.J. A restorer gene for genetic-cytoplasmic male sterility in cultivated potatoes. Am. Potato J. 1991, 68, 19–28. [Google Scholar] [CrossRef]
- Mihovilovich, E.; Sanetomo, R.; Hosaka, K.; Ordoñez, B.; Aponte, M.; Bonierbale, M. Cytoplasmic diversity in potato breeding: Case study from the International Potato Center. Mol. Breed. 2015, 35, 137. [Google Scholar] [CrossRef]
- Sanetomo, R.; Nashiki, A. Identification of the tetrad-sterility-causing Solanum stoloniferum Schltdl. & Bouché cytoplasm in interspecific hybrids with S. tuberosum L. Genet. Resour. Crop Evol. 2021, 68, 3383–3397. [Google Scholar] [CrossRef]
- Dionne, L.A. Cytoplasmic sterility in derivatives of Solanum demissum. Am. Potato J. 1961, 38, 117–120. [Google Scholar] [CrossRef]
- Gavrilenko, A.; Klimenko, N.S.; Alpatieva, N.V.; Kostina, L.I.; Lebedeva, V.A.; Evdokimova, Z.Z.; Apalikova, O.V.; Novikova, L.Y.; Antonova, O.Y. Cytoplasmic genetic diversity of potato varieties bred in Russia and FSU countries. Vavilov J. Genet. Breed. 2019, 23, 753–764. [Google Scholar] [CrossRef] [Green Version]
- Hosaka, K.; Sanetomo, R. Development of a rapid identification method for potato cytoplasm and its use for evaluating Japanese collections. Theor. Appl. Genet. 2012, 125, 1237–1251. [Google Scholar] [CrossRef]
- Bentolila, S.; Alfonso, A.A.; Hanson, M.R. A pentatricopeptide repeat-containing gene restores fertility to cytoplasmic male-sterile plants. Proc. Natl. Acad. Sci. USA 2002, 99, 10887–10892. [Google Scholar] [CrossRef] [Green Version]
- Gaborieau, L.; Brown, G.G.; Mireau, H. The propensity of pentatricopeptide repeat genes to evolve into restorers of cytoplasmic male sterility. Front. Plant Sci. 2016, 7, 1816. [Google Scholar] [CrossRef] [Green Version]
- Anisimova, I.N.; Alpatieva, N.V.; Karabitsina, Y.I.; Gavrilenko, T.A. Nucleotide Sequence Polymorphism in the RFL-PPR Genes of Potato. J. Genet. 2019, 98, 87. [Google Scholar] [CrossRef]
- Sanetomo, R.; Akai, K.; Nashiki, A. Discovery of a novel mitochondrial DNA molecule associated with tetrad pollen sterility in potato. BMC Plant Biol. 2022, 22, 302. [Google Scholar] [CrossRef] [PubMed]
- Hosaka, K. Who is the mother of the potato?—Restriction endonuclease analysis of chloroplast DNA of cultivated potatoes. Theor. Appl. Genet. 1986, 72, 606–618. [Google Scholar] [CrossRef] [PubMed]
- Lössl, A.; Götz, M.; Braun, A.; Wenzel, G. Molecular markers for cytoplasm in potato: Male sterility and contribution of different plastid-mitochondrial configurations to starch production. Euphytica 2000, 116, 221–230. [Google Scholar] [CrossRef]
- Provan, J.; Powell, W.; Dewar, H.; Bryan, G.; Machray, G.C.; Waugh, R. An Extreme Cytoplasmic Bottleneck in the Modern European Cultivated Potato (Solanum Tuberosum) Is Not Reflected in Decreased Levels of Nuclear Diversity. Proc. R. Soc. B Biol. Sci. 1999, 266, 633–639. [Google Scholar] [CrossRef] [Green Version]
- Landeo, J. Late Blight Strategy at CIP. In Proceedings of the International Potato Center. 1989. Fungal Diseases of Potatoes. Report of the Planning Conference on Fungal Diseases of the Potato Held at CIP, Lima, Peru, 1989; pp. 57–73. [Google Scholar]
- Vanishree, G.; Patil, V.U.; Kaur, R.P.; Bhardwaj, V.; Chakrabarti, S.K.; Kumar, M. Cytoplasmic Types of Indian Potato Cultivars and Their Effect on Important Agronomic Traits. Agric. Res. 2021, 11, 390–397. [Google Scholar] [CrossRef]
- Mori, K.; Mukojima, N.; Nakao, T.; Tamiya, S.; Sakamoto, Y.; Sohbaru, N.; Hayashi, K.; Watanuki, H.; Nara, K.; Yamazaki, K.; et al. Germplasm Release: Saikai 35, a Male and Female Fertile Breeding Line Carrying Solanum Phureja-Derived Cytoplasm and Potato Cyst Nematode Resistance (H1) and Potato Virus Y Resistance (Ry chc) Genes. Am. J. Potato Res. 2012, 89, 63–72. [Google Scholar] [CrossRef]
- Hoopes, R.W.; Plaisted, R.L.; Cubillos, A.G. Yield and fertility of reciprocal-cross Tuberosum-Andigena hybrids. Am. Potato J. 1980, 57, 275–284. [Google Scholar] [CrossRef]
- Vilaró, F.L.; Plaisted, R.L.; Hoopes, R.W. Comparison of cytoplasmic male sterilities in progenies of Tuberosum × Andigena and Tuberosum × Neo-Tuberosum crosses. Am. Potato J. 1989, 66, 13–24. [Google Scholar] [CrossRef]
- Wang, R.; Cai, X.; Hu, S.; Li, Y.; Fan, Y.; Tan, S.; Liu, Q.; Zhou, W. Comparative Analysis of the Mitochondrial Genomes of Nicotiana tabacum: Hints Toward the Key Factors Closely Related to the Cytoplasmic Male Sterility Mechanism. Front. Genet. 2020, 11, 257. [Google Scholar] [CrossRef] [Green Version]
- Jindal, S.K.; Dhaliwal, M.S.; Meena, O.P. Molecular advancements in male sterility systems of Capsicum: A review. Plant Breed. 2019, 139, 42–64. [Google Scholar] [CrossRef]
- Yoshimi, M.; Kitamura, Y.; Isshiki, S.; Saito, T.; Yasumoto, K.; Terachi, T.; Yamagishi, H. Variations in the structure and transcription of the mitochondrial atp and cox genes in wild Solanum species that induce male sterility in eggplant (S. melongena). Theor. Appl. Genet. 2013, 126, 1851–1859. [Google Scholar] [CrossRef] [PubMed]
- Dafni, A.; Firmage, D. Pollen viability and longevity: Practical, ecological and evolutionary implications. In Pollen and Pollination; Springer: Berlin/Heidelberg, Germany, 2000; pp. 113–132. [Google Scholar] [CrossRef]
- Trognitz, B.R. Comparison of different pollen viability assays to evaluate pollen fertility of potato dihaploids. Euphytica 1991, 56, 143–148. [Google Scholar] [CrossRef]
- Khan, M.R.; Isshiki, S. Cytoplasmic Male Sterility in Eggplant. Hortic. J. 2016, 85, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; An, D.; Cao, Y.; Yu, H.; Zhu, Y.; Mei, Y.; Zhang, B.; Wang, L. Development and application of KASP markers associated with Restorer-of-fertility gene in Capsicum annuum L. Physiol. Mol. Biol. Plants 2021, 27, 2757–2765. [Google Scholar] [CrossRef] [PubMed]
- Alsahlany, M.; Enciso-Rodriguez, F.; Lopez-Cruz, M.; Coombs, J.; Douches, D.S. Developing self-compatible diploid potato germplasm through recurrent selection. Euphytica 2021, 217, 47. [Google Scholar] [CrossRef]
- Lindhout, P.; Meijer, D.; Schotte, T.; Hutten, R.C.B.; Visser, R.G.F.; van Eck, H.J. Towards F1 Hybrid Seed Potato Breeding. Potato Res. 2011, 54, 301–312. [Google Scholar] [CrossRef] [Green Version]
- Jansky, S.H.; Peloquin, S.J. Advantages of Wild Diploid Solanum Species Over Cultivated Diploid Relatives in Potato Breeding Programs. Genet. Resour. Crop Evol. 2006, 53, 669–674. [Google Scholar] [CrossRef]
- Phumichai, C.; Ikeguchi-Samitsu, Y.; Fujimatsu, M.; Kitanishi, S.; Kobayashi, A.; Mori, M.; Hosaka, K. Expression of S-locus inhibitor gene (Sli) in various diploid potatoes. Euphytica 2006, 148, 227–234. [Google Scholar] [CrossRef] [Green Version]
- Gastelo, M.; Burgos, G.; Bastos, C.; Perez, J.M.; Otiniano, R.; Cabrera, H.; Ccanto, R.; de Scurrah, M.M.; Felde, T.Z. Identification of Elite Potato Clones with Resistance to Late Blight Through Participatory Varietal Selection in Peru. Potato Res. 2021, 64, 611–634. [Google Scholar] [CrossRef]
- Landeo, J.A.; Gastelo, M.; Beltran, G.; Diaz, L. Quantifying Genetic Variance for Horizontal Resistance to Late Blight in Potato Breeding Population B3C1. CIP Program Rep. 1999, 2000, 63–68. [Google Scholar]
- Gastelo, M.; Diaz, L.; Landeo, J.A.; Bonierbale, M. New elite potato clones with heat tolerance, late blight and virus resistance to address climate change. In Potato and Sweetpotato in Africa: Transforming the Value Chains for Food and Nutrition Security; CABI: Oxfordshire, UK, 2015; pp. 143–152. [Google Scholar]
- Gastelo, M.; Kleinwechter, U.; Bonierbale, M. Gobal Potato Research for a Changing World—Technical Report; International Potato Center: Lima, Peru, 2014. [Google Scholar]
- Santayana, M.; Osorio, F.; Mendes, T.; Lindqvist-Kreuze, H. Guide for Evaluation of Potato Essential Reproductive Characteristics in Greenhouse Conditions; International Potato Center: Lima, Peru, 2021. [Google Scholar]
- Ordoñez, B.; Orrillo, M.; Bonierbale, M. Technical Manual Potato Reproductive and Cytological Biology; International Potato Center: Lima, Peru, 2017. [Google Scholar]
- Huaman, Z.; Williams, J.T.; Salhuana, W.; Vincent, L. Descriptors for the Cultivated Potato; International Board for Plant Genetic Resources: Rome, Italy, 1977; p. 50. [Google Scholar]
- Camadro, E.L.; Peloquin, S.J. Cross-incompatibility between two sympatric polyploid Solanum species. Theor. Appl. Genet. 1981, 60, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Rife, T.W.; Poland, J.A. Field Book: An Open-Source Application for Field Data Collection on Android. Crop Sci. 2014, 54, 1624–1627. [Google Scholar] [CrossRef]
- Mutterer, J.; Zinck, E. Quick-and-clean article figures with FigureJ. J. Microsc. 2013, 252, 89–91. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2022. [Google Scholar]
- Butler, D.G. Asreml: Fits the Linear Mixed Model. R Package Version 4.1.0.130. Available online: http://www.vsni.co.uk (accessed on 25 October 2022).
- Brien, C. Asremlplus: Augments ‘ASReml-R’ in Fitting Mixed Models and Packages Generally in Exploring Prediction Differences. R Package Version 4.3.36. Available online: https://CRAN.R-project.org/package=asremlPlus (accessed on 25 October 2022).




{kind=link}
{kind=link}
{kind=link}
| Populations | ||||
|---|---|---|---|---|
| Oldest Maternal Ancestor | Ancestor Generations 1 | B3C3 | LBHT | LBHT x LTVR |
| Proportion of D-type breeding lines (N = 83) | 96.8% | 39.5% | 16.2% | |
| AC 25953 2 | 8 | 2 | -- | -- |
| 54-Q-2 | 8 | 16 | -- | -- |
| INDIA-832 | 6 | 10 | 3 | -- |
| GRETA | 6 | -- | 2 | 2 |
| 66-563-13C | 6 | 2 | -- | -- |
| NEVADA | 6 | 9 | 11 | 3 |
| CIP676084 | 5 | 13 | 1 | 1 |
| INDIA-1058 B 3 | 5 | 4 | -- | -- |
| TXY.4 | 2 | 4 | -- | -- |
| Proportion of T-type breeding lines (N = 58) | 3.2% | 58.1% | 83.8% | |
| AC 25953 2 | 8 | -- | 2 | 2 |
| CIP800122 | 7 | -- | 23 | 29 |
| CIP750627 | 5 | 2 | -- | -- |
| Proportion of W/γ-type breeding lines (N = 1) | 0% | 2.3% | 0% | |
| PG-232 | 2 | -- | 1 | -- |
| D and T Cytoplasm | D Cytoplasm | T Cytoplasm | ||||
|---|---|---|---|---|---|---|
| Fruit Set (%) | Seed Set | Fruit Set (%) | Seed Set | Fruit Set (%) | Seed Set | |
| Anther-lobe overlap | 0.29 *** | 0.23 ** | 0.10 | 0.08 | 0.18 | 0.04 |
| Pollen viability | −0.17 *** | 0.05 | −0.10 | −0.10 | 0.18 | 0.35 *** |
| In vivo pollen tube growth | −0.53 *** | −0.46 *** | −0.28 *** | −0.16 | −0.48 *** | −0.41 *** |
| Breeding Lines | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| D-Type | T-Type | Total MF | Total MS | Candidate | |||||
| Parent Type | CIP Number | CyT | MF | MS | MF | MS | |||
| Male | CIP392633.64 | D | 3 | 0 | 7 | 0 | 10 | 0 | Rf+ |
| CIP396272.43 | D | 0 | 0 | 6 | 0 | 6 | 0 | Rf+ | |
| CIP304372.7 | T | 0 | 1 | 4 | 3 | 4 | 4 | Rf+ | |
| CIP392639.2 | D | 2 | 2 | 0 | 0 | 2 | 2 | Rf+ | |
| CIP395017.229 | D | 2 | 2 | 0 | 0 | 2 | 2 | Rf+ | |
| CIP396012.288 | D | 5 | 3 | 0 | 0 | 5 | 3 | Rf+ | |
| CIP396038.107 | D | 0 | 6 | 0 | 0 | 0 | 6 | Rf− | |
| CIP396041.102 | T | 0 | 4 | 0 | 0 | 0 | 4 | Rf− | |
| Female | CIP395117.3 | D | -- | -- | -- | -- | 0 | 2 | Rf− |
| CIP396034.103 | D | -- | -- | -- | -- | 0 | 3 | Rf− | |
| CIP396046.105 | D | -- | -- | -- | -- | 0 | 3 | Rf− | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santayana, M.; Aponte, M.; Kante, M.; Eyzaguirre, R.; Gastelo, M.; Lindqvist-Kreuze, H. Cytoplasmic Male Sterility Incidence in Potato Breeding Populations with Late Blight Resistance and Identification of Breeding Lines with a Potential Fertility Restorer Mechanism. Plants 2022, 11, 3093. https://doi.org/10.3390/plants11223093
Santayana M, Aponte M, Kante M, Eyzaguirre R, Gastelo M, Lindqvist-Kreuze H. Cytoplasmic Male Sterility Incidence in Potato Breeding Populations with Late Blight Resistance and Identification of Breeding Lines with a Potential Fertility Restorer Mechanism. Plants. 2022; 11(22):3093. https://doi.org/10.3390/plants11223093
Chicago/Turabian StyleSantayana, Monica, Mariela Aponte, Moctar Kante, Raúl Eyzaguirre, Manuel Gastelo, and Hannele Lindqvist-Kreuze. 2022. "Cytoplasmic Male Sterility Incidence in Potato Breeding Populations with Late Blight Resistance and Identification of Breeding Lines with a Potential Fertility Restorer Mechanism" Plants 11, no. 22: 3093. https://doi.org/10.3390/plants11223093