Distribution Models of Timber Species for Forest Conservation and Restoration in the Andean-Amazonian Landscape, North of Peru

 ,
,  ,
,  ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Methodological Design
2.3. Selection and Collection of Occurrences of the Most Important Forest Species in the Amazonas Region
2.4. Basic and Thematic Cartography Conditioning
2.5. Bioclimatic, Soil, and Physiographic Variables
2.6. Extraction of Values of Climatic, Soil, and Physiographic Variables
2.7. Variable Correlation and Clustering
2.8. Model Execution
2.9. Identification of Potential Areas for Conservation and Restoration
3. Results
3.1. Biographical Distribution of Timber Species
3.2. Model Performance
3.3. Potential Areas of Restoration and Coexistence of Species
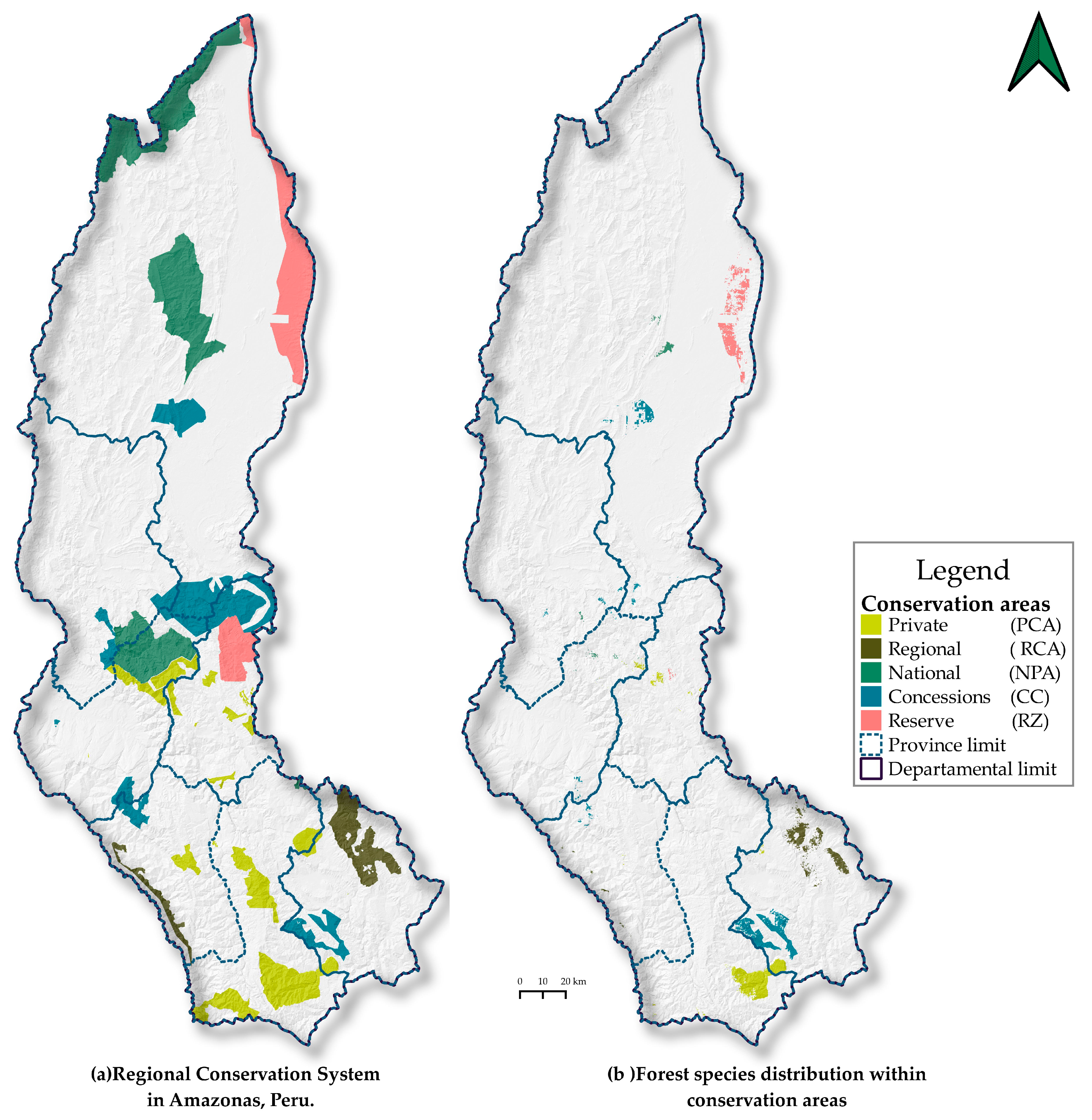
3.4. Potential Biogeographic Distribution in Conservation Areas
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Flores, Y. Crecimiento y Productividad de Plantaciones Forestales en la Amazonía Peruana; Corporación Grafica Andina SAC: Lima, Peru, 2010. [Google Scholar]
- MINAM. Perú Reino de Bosques; Editorial Etiqueta Negra/Pool Editores SAC: Lima, Peru, 2014. [Google Scholar]
- Andersen, H.E.; Reutebuch, S.E.; McGaughey, R.J.; d’Oliveira, M.V.N.; Keller, M. Monitoring selective logging in western amazonia with repeat lidar flights. Remote Sens. Environ. 2014, 151, 157–165. [Google Scholar] [CrossRef] [Green Version]
- Killick, E. Extractive Relations: Natural Resource Use, Indigenous Peoples and Environmental Protection in Peru. Bull. Lat. Am. Res. 2020, 39, 290–304. [Google Scholar] [CrossRef]
- Dourojeanni, M. Aprovechamiento del barbecho forestal en áreas de agricultura migratoria en la Amazonía peruana. Rev. For. Perú 1987, 14, 1–33. [Google Scholar]
- Adhikari, D.; Barik, S.K.; Upadhaya, K. Habitat distribution modelling for reintroduction of Ilex khasiana Purk., a critically endangered tree species of northeastern India. Ecol. Eng. 2012, 40, 37–43. [Google Scholar] [CrossRef] [Green Version]
- Cánovas, F.; Magliozzi, C.; Mestre, F.; Palazón, J.A.; González-Wangüemert, M. ENiRG: R-GRASS interface for efficiently characterizing the ecological niche of species and predicting habitat suitability. Ecography 2016, 39, 593–598. [Google Scholar] [CrossRef]
- Tripathi, P.; Dev Behera, M.; Roy, P.S. Optimized grid representation of plant species richness in India-Utility of an existing national database in integrated ecological analysis. PLoS ONE 2017, 12, e0173774. [Google Scholar] [CrossRef] [Green Version]
- Guisan, A.; Thuiller, W. Predicting species distribution: Offering more than simple habitat models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef]
- Peterson, A.T.; Soberón, J.; Pearson, R.G.; Anderson, R.P.; Martínez-Meyer, E.; Nakamura, M.; Araújo, M.B. Ecological Niches and Geographic Distributions (MPB-49); Princeton University Press: Princeton, NJ, USA, 2011. [Google Scholar]
- Fagundes, C.K.; Vogt, R.C.; De Marco Júnior, P. Testing the efficiency of protected areas in the Amazon for conserving freshwater turtles. Divers. Distrib. 2016, 22, 123–135. [Google Scholar] [CrossRef]
- Antúnez, P.; Suárez-Mota, M.E.; Valenzuela-Encinas, C.; Ruiz-Aquino, F. The potential distribution of tree species in three periods of time under a climate change scenario. Forests 2018, 9, 628. [Google Scholar] [CrossRef] [Green Version]
- Guisan, A.; Zimmermann, N.E. Predictive Habitat Distribution Models in Ecology. Ecol. Model. 2000, 135, 147–186. [Google Scholar] [CrossRef]
- Phillips, S.B.; Aneja, V.P.; Kang, D.; Arya, S.P. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Bai, Y.; Wei, X.; Li, X. Distributional dynamics of a vulnerable species in response to past and future climate change: A window for conservation prospects. PeerJ 2018, 2018, 1–25. [Google Scholar] [CrossRef]
- Beltramino, A.A.; Vogler, R.E.; Gutiérrez Gregoric, D.E.; Rumi, A. Impact of climate change on the distribution of a giant land snail from South America: Predicting future trends for setting conservation priorities on native malacofauna. Clim. Chang. 2015, 131, 621–633. [Google Scholar] [CrossRef]
- Naveda-Rodríguez, A.; Vargas, F.H.; Kohn, S.; Zapata-Ríos, G. Andean Condor (Vultur gryphus) in Ecuador: Geographic Distribution, Population Size and Extinction Risk. PLoS ONE 2016, 11, e0151827. [Google Scholar] [CrossRef] [PubMed]
- Alfonso-Corrado, C.; Naranjo-Luna, F.; Clark-Tapia, R.; Campos, J.E.; Rojas-Soto, O.R.; Luna-Krauletz, M.D.; Bodenhorn, B.; Gorgonio-Ramírez, M.; Pacheco-Cruz, N. Effects of environmental changes on the occurrence of Oreomunnea mexicana (Juglandaceae) in a biodiversity hotspot cloud forest. Forests 2017, 8, 261. [Google Scholar] [CrossRef]
- Qin, A.; Liu, B.; Guo, Q.; Bussmann, R.W.; Ma, F.; Jian, Z.; Xu, G.; Pei, S. Maxent modeling for predicting impacts of climate change on the potential distribution of Thuja sutchuenensis Franch., an extremely endangered conifer from southwestern China. Glob. Ecol. Conserv. 2017, 10, 139–146. [Google Scholar] [CrossRef]
- Abdelaal, M.; Fois, M.; Fenu, G.; Bacchetta, G. Using MaxEnt modeling to predict the potential distribution of the endemic plant Rosa arabica Crép. in Egypt. Ecol. Inform. 2019, 50, 68–75. [Google Scholar] [CrossRef]
- Otieno, B.A.; Nahrung, H.F.; Steinbauer, M.J. Where did you come from? Where did you go? Investigating the origin of invasive Leptocybe species using distribution modelling. Forests 2019, 10, 115. [Google Scholar] [CrossRef] [Green Version]
- Kariyawasam, C.S.; Kumar, L.; Ratnayake, S.S. Invasive plant species establishment and range dynamics in Sri Lanka under climate change. Entropy 2019, 21, 571. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Zhang, H.; Yue, J.; Xie, T.; Xu, Y.; Tian, Y. Predicting shifts in the suitable climatic distribution of walnut (Juglans regia L.) in China: Maximum entropy model paves the way to forest management. Forests 2018, 9, 103. [Google Scholar]
- Frey, G.P.; West, T.A.P.; Hickler, T.; Rausch, L.; Gibbs, H.K.; Börner, J. Simulated impacts of soy and infrastructure expansion in the Brazilian Amazon: A maximum entropy approach. Forests 2018, 9, 600. [Google Scholar] [CrossRef] [Green Version]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Xu, D.; Agee, E.; Wang, J.; Ivanov, V.Y. Estimation of Evapotranspiration of Amazon Rainforest Using the Maximum Entropy Production Method. Geophys. Res. Lett. 2019, 46, 1402–1412. [Google Scholar] [CrossRef]
- Fonseca, M.G.; Aragão, L.E.O.C.; Lima, A.; Shimabukuro, Y.E.; Arai, E.; Anderson, L.O. Modelling fire probability in the Brazilian Amazon using the maximum entropy method. Int. J. Wildland Fire 2016, 25, 955–969. [Google Scholar] [CrossRef]
- Gilani, H.; Arif Goheer, M.; Ahmad, H.; Hussain, K. Under predicted climate change: Distribution and ecological niche modelling of six native tree species in Gilgit-Baltistan, Pakistan. Ecol. Indic. 2020, 111, 106049. [Google Scholar] [CrossRef]
- Doland Nichols, J.; Rosemeyer, M.E.; Lynn Carpenter, F.; Kettler, J. Intercropping legume trees with native timber trees rapidly restores cover to eroded tropical pasture without fertilization. For. Ecol. Manag. 2001, 152, 195–209. [Google Scholar] [CrossRef]
- Carpenter, F.L.; Nichols, J.D.; Pratt, R.T.; Young, K.C. Methods of facilitating reforestation of tropical degraded land with the native timber tree, Terminalia amazonia. For. Ecol. Manag. 2004, 202, 281–291. [Google Scholar] [CrossRef]
- Walentowski, H.; Heinrichs, S.; Hohnwald, S.; Wiegand, A.; Heinen, H.; Thren, M.; Gamarra Torres, O.A.; Sabogal, A.B.; Zerbe, S. Vegetation succession on degraded sites in the Pomacochas Basin (Amazonas, N Peru)-Ecological options for forest restoration. Sustainability 2018, 10, 609. [Google Scholar] [CrossRef] [Green Version]
- Lamb, D.; Erskine, P.D.; Parrotta, J.A. Restoration of Degraded Tropical Forest Landscapes. Science 2005, 310, 1628–1632. [Google Scholar] [CrossRef] [Green Version]
- Chazdon, R.L. Towards more effective integration of tropical forest restoration and conservation. Biotropica 2019, 51, 463–472. [Google Scholar] [CrossRef]
- Cole, R.J.; Holl, K.D.; Zahawi, R.A. Seed rain under tree islands planted to restore degraded lands in a tropical agricultural landscape. Ecol. Appl. 2010, 20, 1255–1269. [Google Scholar] [CrossRef] [PubMed]
- Ymber, F.B. Especies Forestales Nativas Para Recuperacion de Areas Degradadas en la Region Ucayali; Imprenta Davidson: Pucallpa, Peru, 2014. [Google Scholar]
- Schmidt, I.B.; de Urzedo, D.I.; Piña-Rodrigues, F.C.M.; Vieira, D.L.M.; de Rezende, G.M.; Sampaio, A.B.; Junqueira, R.G.P. Community-based native seed production for restoration in Brazil—The role of science and policy. Plant Biol. 2019, 21, 389–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romijn, E.; Coppus, R.; De Sy, V.; Herold, M.; Roman-Cuesta, R.M.; Verchot, L. Land restoration in Latin America and the Caribbean: An overview of recent, ongoing and planned restoration initiatives and their potential for climate change mitigation. Forests 2019, 10, 510. [Google Scholar] [CrossRef] [Green Version]
- Blignaut, J.; Aronson, J.; de Groot, R. Restoration of natural capital: A key strategy on the path to sustainability. Ecol. Eng. 2014, 65, 54–61. [Google Scholar] [CrossRef]
- Jacobs, D.F.; Oliet, J.A.; Aronson, J.; Bolte, A.; Bullock, J.M.; Donoso, P.J.; Landhäusser, S.M.; Madsen, P.; Peng, S.; Rey-Benayas, J.M.; et al. Restoring forests: What constitutes success in the twenty-first century? New For. 2015, 46, 601–614. [Google Scholar] [CrossRef]
- MINAM. El Perú de los Bosques; MINAM: Lima, Peru, 2011. [Google Scholar]
- MINAM. Mapa Nacional de Cobertura Vegetal: Memoria Descriptiva; MINAM: Lima, Peru, 2015; p. 105. [Google Scholar]
- GRA; IIAP. Zonificación Ecológica y Económica (ZEE) del Departamento de Amazonas; IIAP: Iquitos, Peru, 2010. [Google Scholar]
- OSINFOR. Modelamieto Espacial de Nichos Ecológicos Para la Evaluación de Presencia de Especies Forestales Maderables en la Amazonía Peruana; Punto & Grafía S.A.C.: Lima, Peru, 2013. [Google Scholar]
- Hengl, T.; De Jesus, J.M.; Heuvelink, G.B.M.; Gonzalez, M.R.; Kilibarda, M.; Blagotić, A.; Shangguan, W.; Wright, M.N.; Geng, X.; Bauer-Marschallinger, B.; et al. SoilGrids250m: Global gridded soil information based on machine learning. PLoS ONE 2017, 12, e0169748. [Google Scholar] [CrossRef] [Green Version]
- Rojas, N.B.; Cotrina, D.A.; Castillo, E.B.; Oliva, M.; Salas, R. Current and Future Distribution of Five Timber Forest Species in Amazonas, Northeast Peru: Contributions towards a Restoration Strategy. Diversity 2020, 12, 305. [Google Scholar] [CrossRef]
- OSINFOR. Sistema de Información Gerencial del OSINFOR. Reportes Estadísticos: Principales Especies Forestales Maderables Aprobadas. Available online: https://observatorio.osinfor.gob.pe/Estadisticas/Home/Reportes/1 (accessed on 15 April 2020).
- Pliscoff, P.; Fuentes, T. Modelación de la distribución de especies y ecosistemas en el tiempo y en el espacio: Una revisión de las nuevas herramientas y enfoques disponibles. Rev. Geogr. Norte Gd. 2011, 48, 61–79. [Google Scholar] [CrossRef] [Green Version]
- Scheldeman, X.; van Zonneveld, M. Training Manual on Spatial Analysis of Plant Diversity and Distribution; Bioversity International: Rome, Italy, 2010. [Google Scholar]
- OSINFOR. Modelamiento de la Distribución Potencial de 18 Especies Forestales en el Departamento de Loreto; OSINFOR: Lima, Peru, 2016. [Google Scholar]
- Stevens, G.C. The latitudinal gradient in geographic range: How so many species coexist in the tropics. Am. Nat. 1989, 133, 240–256. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- De Marco, P.; Nobrega, C.C. Evaluating collinearity effects on species distribution models: An approach based on virtual species simulation. PLoS ONE 2018, 13, e0202403. [Google Scholar] [CrossRef] [PubMed]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquez, J.R.G.; Gruber, B.; Lafourcade, B.; Leitao, P.J.; et al. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 027–046. [Google Scholar] [CrossRef]
- Zhang, K.; Zhang, Y.; Zhou, C.; Meng, J.; Sun, J.; Zhou, T.; Tao, J. Impact of Climate Factors on Future Distributions of Paeonia Ostii across China Estimated by MaxEnt; Elsevier: Amsterdam, The Netherlands, 2019. [Google Scholar]
- Zhang, K.; Zhang, Y.; Tao, J. Predicting the potential distribution of Paeonia veitchii (Paeoniaceae) in China by incorporating climate change into a maxent model. Forests 2019, 10, 190. [Google Scholar] [CrossRef] [Green Version]
- Heikkinen, R.K.; Luoto, M.; Araújo, M.B.; Virkkala, R.; Thuiller, W.; Sykes, M.T. Methods and uncertainties in bioclimatic envelope modelling under climate change. Prog. Phys. Geogr. 2006, 30, 751–777. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.Q.; Kushwaha, S.P.S.; Saran, S.; Xu, J.; Roy, P.S. Maxent modeling for predicting the potential distribution of medicinal plant, Justicia adhatoda L. in Lesser Himalayan foothills. Ecol. Eng. 2013, 51, 83–87. [Google Scholar] [CrossRef]
- Patel, V.R.; Mehta, R.G. Impact of Outlier Removal and Normalization Approach in Modified k-Means Clustering Algorithm. IJCSI Int. J. Comput. Sci. Issues 2011, 8, 331–336. [Google Scholar]
- Jaynes, E.T. Information Theory and Statistical Mechanics. Phys. Rev. 1957, 106, 620–630. [Google Scholar] [CrossRef]
- Merow, C.; Smith, M.J.; Silander, J.A. A practical guide to MaxEnt for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Manel, S.; Williams, C.; Ormerod, S.J. Evaluating presence—Absence models in ecology: The need to account for prevalence. J. Appl. Ecol. 2001, 38, 921–931. [Google Scholar] [CrossRef]
- Hanley, J.A.; McNeil, B.J. The meaning and use of the area under a receiver operating characteristic (ROC) curve. Radiology 1982, 143, 29–36. [Google Scholar] [CrossRef] [Green Version]
- Araujo, M.; Pearson, R.; Thuiller, W.; Erhard, M. Validation of species-climate impact models under climate change. Glob. Chang. Biol. 2005, 11, 1504–1513. [Google Scholar] [CrossRef] [Green Version]
- Jiménez-Valverde, A. Insights into the area under the receiver operating characteristic curve (AUC) as a discrimination measure in species distribution modelling. Glob. Ecol. Biogeogr. 2012, 21, 498–507. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Zhang, L.; Jing, Z.; Li, Z.; Liu, Y.; Fang, S. Predictive modeling of suitable habitats for Cinnamomum Camphora (L.) presl using maxent model under climate change in China. Int. J. Environ. Res. Public Health 2019, 16, 3185. [Google Scholar] [CrossRef] [Green Version]
- MINAM. Estudio para la Identificación de Áreas Degradadas y Propuesta de Monitoreo; MINAM: Lima, Peru, 2017. [Google Scholar]
- Sears, R.R.; Pinedo-Vasquez, M. Forest Policy Reform and the Organization of Logging in Peruvian Amazonia. Dev. Chang. 2011, 42, 609–631. [Google Scholar] [CrossRef]
- Sears, R.R.; Cronkleton, P.; Polo Villanueva, F.; Miranda Ruiz, M.; Pérez-Ojeda del Arco, M. Farm-forestry in the Peruvian Amazon and the feasibility of its regulation through forest policy reform. For. Policy Econ. 2018, 87, 49–58. [Google Scholar] [CrossRef]
- Arce, R. Aproximaciones a la filosofía forestal de la sustentabilidad del manejo forestal en el Perú. Rev. For. Perú 2019, 34, 113. [Google Scholar] [CrossRef]
- Reynel, C.; Pennington, T.; Pennington, R.; Flores, C.; Daza, A. Árboles Útiles de la Amazonía Peruana y sus Usos; ICRAF: Lima, Peru, 2003. [Google Scholar]
- Reynel, C.; Pennington, T.D.; Pennington, R.T. Árboles del Perú; Imprenta Bellido: Lima, Peru, 2016. [Google Scholar]
- OSINFOR. Fichas de Identificación de Especies Forestales Maderables de la Concesión Forestal Industrial Maderera Zapote (IMAZA)”, Como Producto del Curso Taller: “V Curso Taller Fortalecimiento de Capacidades a Supervisores Forestales en Aplicación de Criterios Técnicos, Reconocimiento de Especies Maderables y uso de Tecnologías en las Supervisiones; Ficha Técnica; MLB Impresiones: Lima, Peru, 2018; pp. 1–58. [Google Scholar]
- Bandopadhyay, S. Does elevation impact local level climate change? An analysis based on fifteen years of daily diurnal data and time series forecasts. Pac. Sci. Rev. A Nat. Sci. Eng. 2016, 18, 241–253. [Google Scholar] [CrossRef]
- Godsoe, W.; Franklin, J.; Blanchet, F.G. Effects of biotic interactions on modeled species’ distribution can be masked by environmental gradients. Ecol. Evol. 2017, 7, 654–664. [Google Scholar] [CrossRef] [Green Version]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudík, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A.; et al. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef] [Green Version]
- Roces-Díaz, J.V.; Jiménez-Alfaro, B.; Álvarez-Álvarez, P.; Álvarez-García, M.A. Environmental niche and distribution of six deciduous tree species in the spanish atlantic region. IForest 2014, 8, 214–221. [Google Scholar] [CrossRef] [Green Version]
- Boiffin, J.; Badeau, V.; Bréda, N. Species distribution models may misdirect assisted migration: Insights from the introduction of Douglas-fir to Europe: Insights. Ecol. Appl. 2017, 27, 446–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laurente, M. Modeling the Effects of Climate Change on the Distribution of Cedrela odorata L. “Cedro” in the Peruvian Amazon. Biologist 2015, 13, 213–224. [Google Scholar]
- OSINFOR. Fichas de Identificación de Especies Forestales Maderables y Silvicultura Tropical”, Como Producto del IV Curso—Taller: “Fortalecimiento de las Capacidades en la Identificación de Especies Forestales Maderables y Silvicultura Tropical”. Dirigido a Supervisores Forestales y de Fauna Silvestre; Ficha Técnica; MLB Impresiones: Lima, Peru, 2017; pp. 1–51. [Google Scholar]
- Pennington, T.; Muellner, A.N.; Wise, R. A Monograph of Cedrela (Meliaceae); Dh Books: Milborne Port, UK, 2010. [Google Scholar]
- OSINFOR. Fichas de Identificación de Especies Forestales Maderables de la Selva Central”, Como Producto del III Curso Taller: “Identificación de Especies Forestales Maderables; Ficha Técnica; Vistay Publicidad E.I.R.L.: Lima, Peru, 2015; pp. 1–69. [Google Scholar]
- OSINFOR. Ficha de Identificación de Especies Forestales Maderables de la Provincia de Tahuamanu, Departamento de Madre de Dios; Ficha Técnica; OSINFOR: Lima, Peru, 2019; pp. 1–59. [Google Scholar]
- Pennington, T.D.; Reynel, C.; Daza, A. Illustrated Guide to the Trees of Peru; David Hunt: Totnes, UK, 2004. [Google Scholar]
- Vásquez, R. Flórula de las Reservas Biológicas de Iquitos, Perú: Allpahuayo-Mishana, Explornapo Camp, Explorama Lodge; Missouri Botanical Garden: Iquitos, Peru, 1997. [Google Scholar]
- Reynel, C.; Pennington, T.D. El Género Inga en el Perú: Morfología, Distribución y Usos; Royal Botanic Gardens: Lima, Peru, 1997. [Google Scholar]
- Abolmaali, S.M.R.; Tarkesh, M.; Bashari, H. MaxEnt modeling for predicting suitable habitats and identifying the effects of climate change on a threatened species, Daphne mucronata, in central Iran. Ecol. Inform. 2018, 43, 116–123. [Google Scholar] [CrossRef]
- Hipólito, J.; Hasui, É.; Viana, B.F. Solving problems involving the distribution of a species of unknown distribution via ecological niche modeling. Nat. Conserv. 2015, 13, 15–23. [Google Scholar] [CrossRef] [Green Version]
- Pramanik, M.; Paudel, U.; Mondal, B.; Chakraborti, S.; Deb, P. Predicting climate change impacts on the distribution of the threatened Garcinia indica in the Western Ghats, India. Clim. Risk Manag. 2018, 19, 94–105. [Google Scholar] [CrossRef]
- Bañuelos-Revilla, J.E.; Palacio-Núñez, J.; Martínez-Montoya, J.F.; Olmos-Oropeza, G.; Flores-Cano, J.A. Potential distribution and abundance of candelilla (Euphorbia antisyphilitica) in northern zacatecas, mexico. Madera Bosques 2019, 25, e2511657. [Google Scholar]
- Velazco, S.J.E.; Galvão, F.; Villalobos, F.; De Marco, P. Using worldwide edaphic data to model plant species niches: An assessment at a continental extent. PLoS ONE 2017, 12, e0186025. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, L.O.; Young, K.R. Biological diversity of Peru: Determining priority areas for conservation. Ambio 2000, 29, 329–337. [Google Scholar] [CrossRef]
- Fajardo, J.; Lessmann, J.; Bonaccorso, E.; Devenish, C.; Muñoz, J. Combined use of systematic conservation planning, species distribution modelling, and connectivity analysis reveals severe conservation gaps in a megadiverse country (Peru). PLoS ONE 2014, 9, e0114367. [Google Scholar] [CrossRef] [Green Version]
- Mark, J.; Rivers, M.C. Cedrela Odorata, Spanish Cedar The IUCN Red List of Threatened Species; IUCN: Gland, Switzerland, 2017. [Google Scholar]
- Durán, A.P.; Casalegno, S.; Marquet, P.A.; Gaston, K.J. Representation of ecosystem services by terrestrial protected areas: Chile as a case study. PLoS ONE 2013, 8, e0082643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Possingham, H.P.; Wilson, K.A.; Andelman, S.J.; Vynne, C.H.; Vynme, C.H. Protected areas: Goals, limitations, and design. In Principles of Conservation Biology; Sinauer Associates: Sunderland, MA, USA, 2006; pp. 509–551. [Google Scholar]
- Dourojeanni, M.J. Protected areas and scientific research in Peru. Rev. For. Perú 2018, 33, 91–101. [Google Scholar]
- Romo, M.; Leo, M.; Epiquién, M. SICRE-AMAZONAS; APECO: Chachapoyas, Peru, 2009. [Google Scholar]
- Román, F.; Mamani, A.; Cruz, A.D.; Sandoval, C.; Cuesta, F. Orientaciones para la Restauración de Ecosistemas Forestales y Otros Ecosistemas de Vegetación Silvestre; SERFOR: Lima, Peru, 2018. [Google Scholar]
- Laestadius, L.; Maginnis, S.; Minnemeyer, S.; Potapov, P.; Saint-Laurent, C.; Sizer, N. Mapa de oportunidades de restauración del paisaje forestal. Unasylva 2011, 62, 47–48. [Google Scholar]
- Burnett, K.M.; Ticktin, T.; Bremer, L.L.; Quazi, S.A.; Geslani, C.; Wada, C.A.; Kurashima, N.; Mandle, L.; Pascua, P.; Depraetere, T.; et al. Restoring to the future: Environmental, cultural, and management trade-offs in historical versus hybrid restoration of a highly modified ecosystem. Conserv. Lett. 2019, 12, e12606. [Google Scholar] [CrossRef] [Green Version]
- Griscom, B.W.; Adams, J.; Ellis, P.W.; Houghton, R.A.; Lomax, G.; Miteva, D.A.; Schlesinger, W.H.; Shoch, D.; Siikamäki, J.V.; Smith, P.; et al. Natural climate solutions. Proc. Natl. Acad. Sci. USA 2017, 114, 11645–11650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schnabel, F.; Schwarz, J.A.; Dănescu, A.; Fichtner, A.; Nock, C.A.; Bauhus, J.; Potvin, C. Drivers of productivity and its temporal stability in a tropical tree diversity experiment. Glob. Chang. Biol. 2019, 25, 4257–4272. [Google Scholar] [CrossRef]
- Gamfeldt, L.; Snäll, T.; Bagchi, R.; Jonsson, M.; Gustafsson, L.; Kjellander, P.; Ruiz-Jaen, M.C.; Fröberg, M.; Stendahl, J.; Philipson, C.D.; et al. Higher levels of multiple ecosystem services are found in forests with more tree species. Nat. Commun. 2013, 4, 1340. [Google Scholar] [CrossRef]
- Van Der Plas, F.; Manning, P.; Soliveres, S.; Allan, E.; Scherer-Lorenzen, M.; Verheyen, K.; Wirth, C.; Zavala, M.A.; Ampoorter, E.; Baeten, L.; et al. Biotic homogenization can decrease landscape-scale forest multifunctionality. Proc. Natl. Acad. Sci. USA 2016, 113, 3557–3562. [Google Scholar] [CrossRef] [Green Version]
- IITO. ITTO Guidelines for the Restoration, Management and Rehabilitation of Degraded and Secondary Tropical Forests; Internaitonal Tropical Timber Organization: Yokohama, Japan, 2002. [Google Scholar]
- Rodríguez, J.C.; Sabogal, C. Restoring degraded forest land with native tree species: The experience of “Bosques Amazónicos” in Ucayali, Peru. Forests 2019, 10, 851. [Google Scholar] [CrossRef] [Green Version]
- Nalvarte, W.; Sabogal, C.; Galvan, O.; Marmillod, D.; Angulo, W.; Cordova, N.; Colan, V. Silvicultura en la Amazonia Peruana: Diagnostico de Experiencias en la Region Ucanyali y la Provincia de Puerto Inca; CIFOR: Ucayali, Peru, 2004. [Google Scholar]
- Pecchi, M.; Marchi, M.; Burton, V.; Giannetti, F.; Moriondo, M.; Bernetti, I.; Bindi, M.; Chirici, G. Species distribution modelling to support forest management. A literature review. Ecol. Model. 2019, 411, 108817. [Google Scholar] [CrossRef]
- Dos Santos, V.A.H.F.; Ferreira, M.J. Initial establishment of commercial tree species under enrichment planting in a Central Amazon secondary forest: Effects of silvicultural treatments. For. Ecol. Manag. 2020, 460, 117822. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nº | Species | Local Name | Family | Conservation Status 1 | Records Number | Vol. m3 (r) |
|---|---|---|---|---|---|---|
| 1 | Cedrelinga cateniformis | Tornillo | Fabaceae | LC | 2193 | 20,786.22 |
| 2 | Otoba parvifolia | Sempo | Myristicaceae | NL | 414 | 1264.81 |
| 3 | Ceiba pentandra | Lupuna | Malvaceae | LC | 333 | 8144.38 |
| 4 | Inga sp. | Guabilla | Fabaceae | NL | 311 | 1125.54 |
| 5 | Apuleia leiocarpa | Anacaspi | Fabaceae | NL | 241 | 1608.01 |
| 6 | Cedrela montana | Cedro | Meliaceae | EN | 215 | 2952.86 |
| 7 | Cariniana decandra | Papelillo | Lecythidaceae | NL | 210 | 2120.07 |
| 8 | Calycophyllum spruceanum | Capirona | Rubiaceae | NL | 201 | 877.16 |
| 9 | Cedrela odorata | Cedro amargo | Meliaceae | VU | 184 | 579.57 |
| 10 | Hura crepitans | Catahua | Euphorbiaceae | NL | 152 | 1569.96 |
| Total | 4454 | 37,686.17 | ||||
| Category | Variable | Description | Units | Species 1 |
|---|---|---|---|---|
| Bioclimatic | Bio01 | Annual Mean Temperature | °C | |
| Bio02 | Mean Diurnal Range (monthly mean (max temp–min temp)) | °C | d; g | |
| Bio03 | Isothermality ((Bio02/Bio07) × 100) | % | a; b; c; f; g; h; i; j | |
| Bio04 | Temperature Seasonality (standard deviation × 100) | °C | d; e; h; j | |
| Bio05 | Max Temperature of Warmest Month | °C | ||
| Bio06 | Min Temperature of Coldest Month | °C | d | |
| Bio07 | Annual Temperature Range (Bio05-Bio06) | °C | ||
| Bio08 | Mean Temperature of Wettest Quarter | °C | b; h | |
| Bio09 | Mean Temperature of Driest Quarter | °C | c; f; i | |
| Bio10 | Mean Temperature of Warmest Quarter | °C | a | |
| Bio11 | Mean Temperature of Coldest Quarter | °C | ||
| Bio12 | Annual Precipitation | mm | ||
| Bio13 | Precipitation of Wettest Month | mm | i | |
| Bio14 | Precipitation of Driest Month | mm | ||
| Bio15 | Precipitation Seasonality (coefficient of variation) | mm | d; e; f; h | |
| Bio16 | Precipitation of Wettest Quarter | mm | a; b; c; d; g | |
| Bio17 | Precipitation of Driest Quarter | mm | i | |
| Bio18 | Precipitation of Warmest Quarter | mm | a; b; d; e; f; g; i; j | |
| Bio19 | Precipitation of Coldest Quarter | mm | d | |
| Rad | Solar Radiation | kJ m−2 day−1 | e; f; g; h; j | |
| Topographic | Elevation | Elevation | m | a; b; c; d; e; f; g; h; i; j |
| Slope | Slope | ° | a; e; i; j | |
| Aspect | Cardinal Slope Direction | ° | a; b; c; e; f; g; i; j | |
| Soil | pH | pH x 10 to 0.30 m | KCl | a; c; d; e; f; g; h; i |
| CIC | Cation Exchange Capacity (at pH 7) 0.30 m | cmolc kg−1 | a; b; c; f; g; i; j | |
| CO | Soil Organic Carbon Stock (fine-grained soil fraction) to 0.15 m | g kg−1 | b; e; f; h; j |
| Species | Variable 1 (%) | Variable 2 (%) | Variable 3 (%) | Total Contribution |
|---|---|---|---|---|
| A. leiocarpa | Elevation (70.1) | Bio 16 (12.5) | Bio 10 (5.4) | 88% |
| C. spruceanum | Elevation (60.5) | Bio08 (24.2) | Aspect (5.3) | 90% |
| C. decandra | Elevation (56.6) | Bio 09 (25.1) | Bio 16 (12.1) | 93.8% |
| C. montana | Bio 19 (78.4) | pH (15.4) | Bio 02 (5.1) | 98.9% |
| C. odorata | Bio08 (45.7) | Elevation (24.7) | Rad (15.5) | 85.9% |
| C.cateniformis | Elevation (47.6) | Bio 18 (21.1) | Bio 09 (9.6) | 78.3% |
| C. pentandra | Elevation (61.5) | Bio 01 (11.6) | Rad (10.9) | 84% |
| H. crepitans | Bio 04 (38.8) | Elevation (23.5) | Bio 08 (14.1) | 76.4% |
| Inga sp. | Elevation (22.7) | Cic (22.4) | Bio 17 (19.9) | 65% |
| O. parvifolia | Elevation (48.3) | Bio 18 (19.4) | Bio 04 (9.3) | 77% |
| Species | A. leiocarpa | C. spruceanum | C. decandra | C. montana | C. odorata |
|---|---|---|---|---|---|
| AUC | 0.954 | 0.985 | 0.958 | 0.868 | 0.985 |
| Species | C. cateniformis | C. pentandra | H. crepitans | Inga sp. | O. parvifolia |
| AUC | 0.914 | 0.952 | 0.977 | 0.965 | 0.948 |
| Species | Potential Distribution | |||||||
|---|---|---|---|---|---|---|---|---|
| High | Moderate | Low | Total | |||||
| km2 | % Amazonas | km2 | % Amazonas | km2 | % Amazonas | km2 | % Amazonas | |
| A. leiocarpa | 761.55 | 1.81 | 2449.89 | 5.83 | 2822.01 | 6.71 | 6033.45 | 14.35 |
| C. spruceanum | 167.52 | 0.40 | 192.64 | 0.46 | 617.64 | 1.47 | 977.8 | 2.33 |
| C. decandra | 761.67 | 1.81 | 1885.82 | 4.48 | 2903.42 | 6.90 | 550.9 | 13.20 |
| C. montana | 2625.42 | 6.24 | 5832.68 | 13.87 | 8649.03 | 20.57 | 17107.1 | 40.68 |
| C. odorata | 210.72 | 0.50 | 423.74 | 1.01 | 893.48 | 2.12 | 1527.93 | 3.63 |
| C.cateniformis | 1194.75 | 2.84 | 2666.85 | 6.34 | 4757.6 | 11.31 | 8619.21 | 20.50 |
| C. pentandra | 584.39 | 1.39 | 1966.59 | 4.68 | 3128.21 | 7.44 | 5679.18 | 13.51 |
| H. crepitans | 411.88 | 0.98 | 890.42 | 2.12 | 1260.59 | 3.00 | 2562.89 | 6.09 |
| Inga sp. | 461.2 | 1.10 | 866.69 | 2.06 | 1542.38 | 3.67 | 1280.27 | 6.83 |
| O. parvifolia | 1000.82 | 2.38 | 2525.53 | 6.01 | 4203.92 | 10.00 | 7730.28 | 18.38 |
| Coexistence | 3.73 | 0.008 | 324.64 | 0.77 | 5971.42 | 14.20 | 6299.79 | 14.97 |
| Species | Potential for Restoration | |||||||
|---|---|---|---|---|---|---|---|---|
| High | Medium | Low | Total | |||||
| km2 | % Amazonas | km2 | % Amazonas | km2 | % Amazonas | km2 | % Amazonas | |
| A. leiocarpa | 305.07 | 0.73 | 931.18 | 2.21 | 902.75 | 2.15 | 2139.00 | 5.09 |
| C. spruceanum | 70.07 | 0.17 | 88.99 | 0.21 | 269.69 | 0.64 | 428.75 | 1.02 |
| C. decandra | 403.36 | 0.96 | 965.52 | 2.30 | 481.79 | 1.15 | 1850.67 | 4.40 |
| C. montana | 1221.79 | 2.91 | 2553.71 | 6.07 | 2349.50 | 5.59 | 6125.00 | 14.57 |
| C. odorata | 3.99 | 0.01 | 125.65 | 0.30 | 236.61 | 0.56 | 366.25 | 0.87 |
| C. cateniformis | 449.20 | 1.07 | 917.02 | 2.18 | 1620.20 | 3.85 | 2986.42 | 7.10 |
| C. pentandra | 297.86 | 0.71 | 880.93 | 2.09 | 986.43 | 2.35 | 2165.22 | 5.15 |
| H. crepitans | 213.54 | 0.51 | 452.09 | 1.08 | 601.97 | 1.43 | 1267.60 | 3.01 |
| Inga sp. | 254.84 | 0.61 | 419.91 | 1.00 | 629.70 | 1.50 | 1304.45 | 3.10 |
| O. parvifolia | 491.96 | 1.17 | 1015.02 | 2.41 | 1291.14 | 3.07 | 2798.12 | 6.65 |
| Coexistence | 1.35 | 0.003 | 137.63 | 0.33 | 2619.48 | 6.23 | 2758.45 | 6.56 |
| Species | High Potential Distribution in the Regional Conservation System | ||||||
|---|---|---|---|---|---|---|---|
| Conservation Category | Total | ||||||
| PCA | RCA | PNA | CC | RZ | km2 | % Amazonas | |
| A. leiocarpa | 4.16 | 23.30 | 4.61 | 32.07 | 0.08 | ||
| C. spruceanum | 3.92 | 3.92 | 0.01 | ||||
| C. decandra | 1.17 | 11.74 | 16.54 | 124.82 | 154.26 | 0.37 | |
| C. montana | 220.89 | 140.14 | 8.37 | 185.43 | 3.16 | 557.98 | 1.33 |
| C. odorata | 3.94 | 1.76 | 5.70 | 0.01 | |||
| C.cateniformis | 11.19 | 42.68 | 24.29 | 78.16 | 0.19 | ||
| C. pentandra | 1.39 | 16.29 | 17.68 | 0.04 | |||
| H. crepitans | 2.39 | 9.61 | 12.00 | 0.03 | |||
| Inga sp. | 0.30 | 0.16 | 0.46 | 0.00 | |||
| O. parvifolia | 4.68 | 7.48 | 32.33 | 44.49 | 0.11 | ||
| Total | 222.06 | 140.14 | 46.7 | 288.34 | 209.34 | 906.72 | 2.16 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cotrina Sánchez, D.A.; Barboza Castillo, E.; Rojas Briceño, N.B.; Oliva, M.; Torres Guzman, C.; Amasifuen Guerra, C.A.; Bandopadhyay, S. Distribution Models of Timber Species for Forest Conservation and Restoration in the Andean-Amazonian Landscape, North of Peru. Sustainability 2020, 12, 7945. https://doi.org/10.3390/su12197945
Cotrina Sánchez DA, Barboza Castillo E, Rojas Briceño NB, Oliva M, Torres Guzman C, Amasifuen Guerra CA, Bandopadhyay S. Distribution Models of Timber Species for Forest Conservation and Restoration in the Andean-Amazonian Landscape, North of Peru. Sustainability. 2020; 12(19):7945. https://doi.org/10.3390/su12197945
Chicago/Turabian StyleCotrina Sánchez, Dany A., Elgar Barboza Castillo, Nilton B. Rojas Briceño, Manuel Oliva, Cristóbal Torres Guzman, Carlos A. Amasifuen Guerra, and Subhajit Bandopadhyay. 2020. "Distribution Models of Timber Species for Forest Conservation and Restoration in the Andean-Amazonian Landscape, North of Peru" Sustainability 12, no. 19: 7945. https://doi.org/10.3390/su12197945