
The Efficacy of a Trivalent Inactivated Salmonella Vaccine Combined with the Live S. Gallinarum 9R Vaccine in Young Layers after Experimental Infections with S. Enteritidis, S. Typhimurium, and S. Infantis

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Salmonella Strains
2.2. Chickens
2.3. Feed and Water
2.4. Vaccines and Vaccination
2.5. Preparation of the Inoculum and Avianization
2.6. Bacteriology
2.7. Pre Trials
2.8. Experimental Infections with Salmonella
2.9. Cloacal Swabbing and Samplings
2.10. Serology
2.11. Statistics
2.12. Animal Welfare
3. Results
3.1. Cloacal Swabs
3.2. Salmonella Recovery from Livers, Spleens, and Caecum Contents
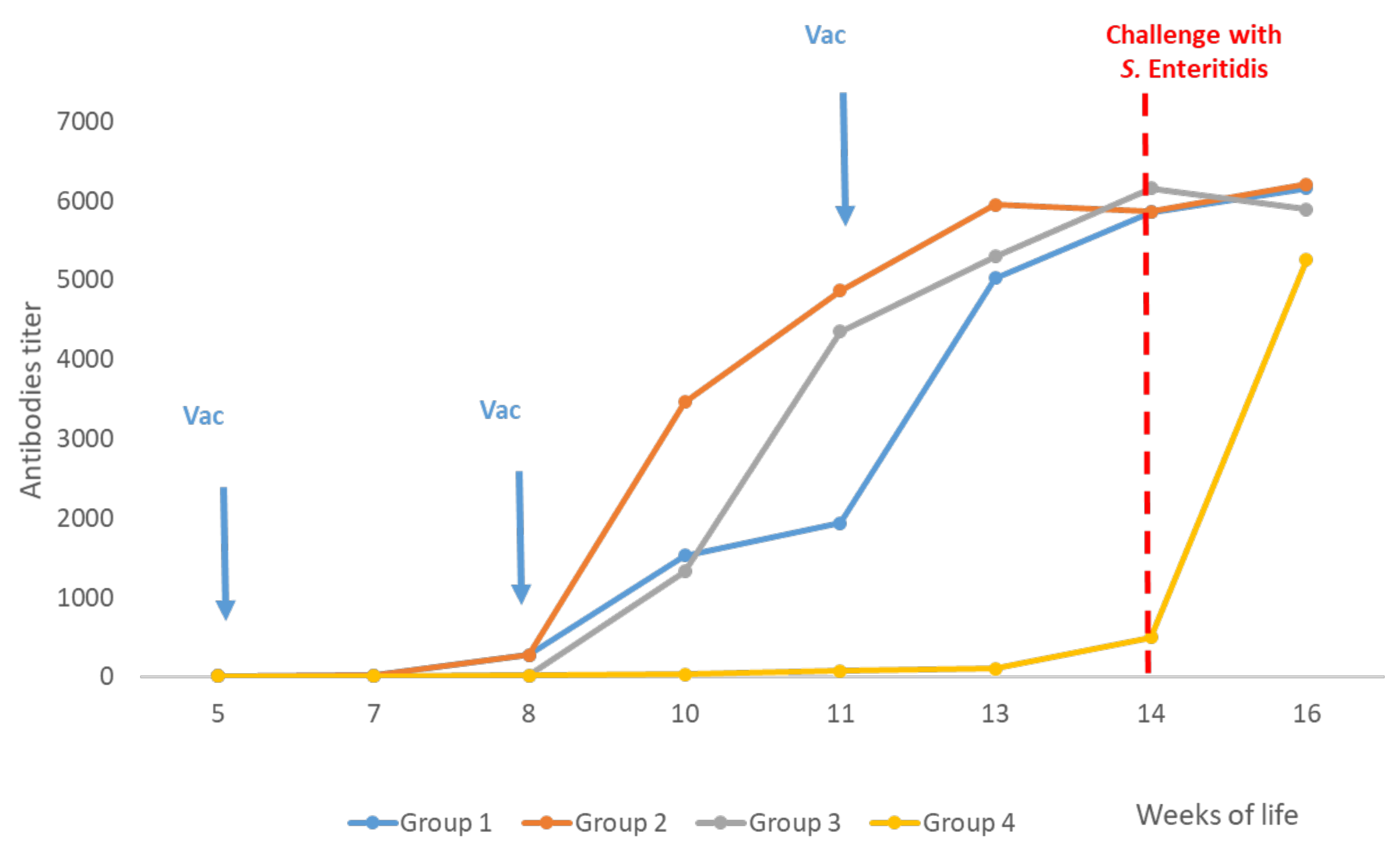
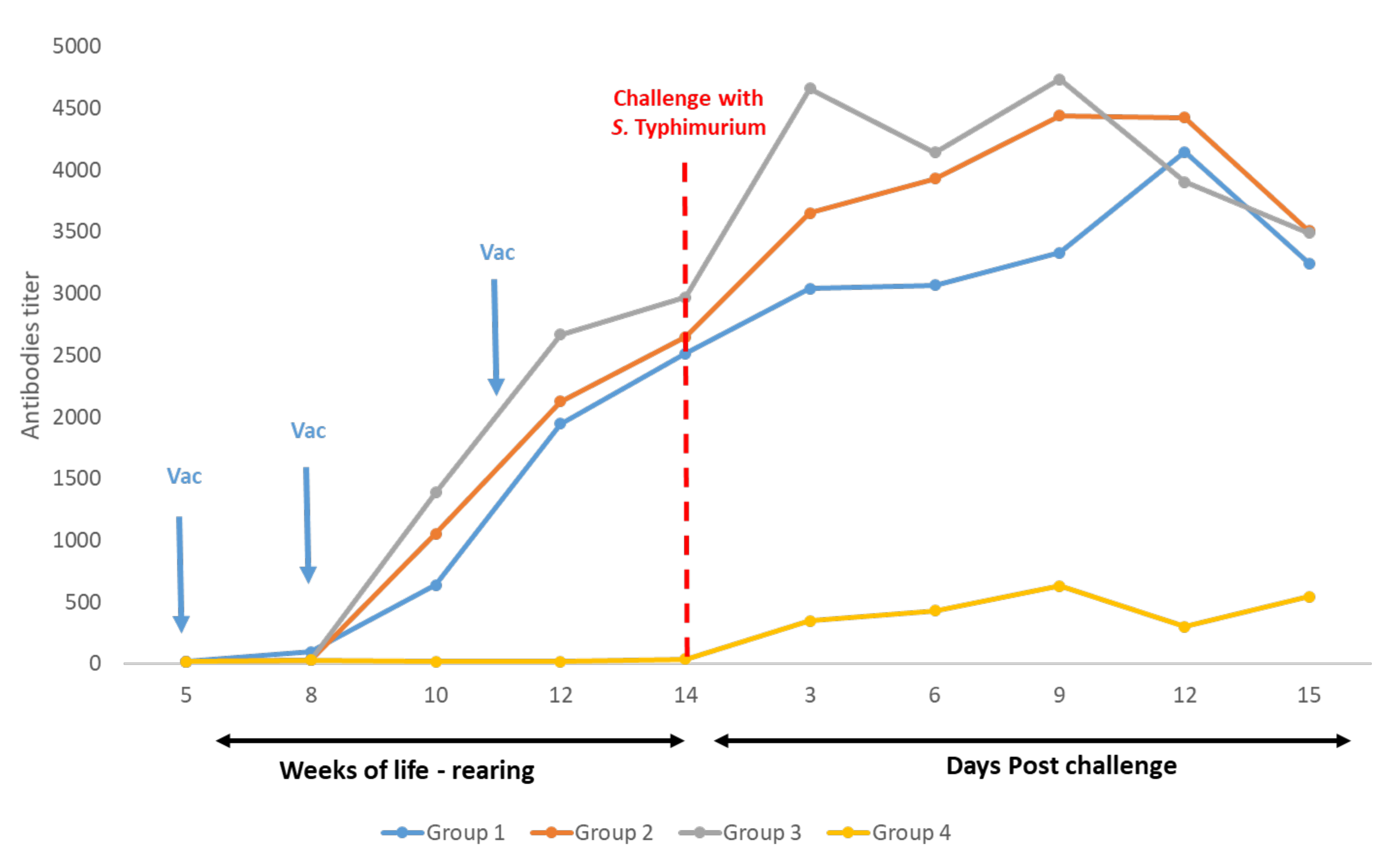
3.3. Serology
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mahmoud, B.S.M. Salmonella—A Dangerous Foodborne Pathogen; Barakat, S.M.M., Ed.; InTech: Rijeka, Croatia, 2012; ISBN 978-953-307-782-6. [Google Scholar]
- Zou, Q.; Li, R.; Liu, G.; Liu, S. Genotyping of Salmonella with Lineage-Specific Genes: Correlation with Serotyping. Int. J. Infect. Dis. 2016, 49, 134–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frenzen, P.D.; Riggs, T.L.; Buzby, J.C.; Breuer, T.; Roberts, T.; Voetsch, D.; Reddy, S.; FoodNet Working Group. Salmonella Cost Estimate Updated Using FoodNet Data. FoodReviews 1999, 22, 10–15. [Google Scholar] [CrossRef] [Green Version]
- Batz, M.; Hoffmann, S.; Morris, J.G. Disease-Outcome Trees, EQ-5D Scores and Estimated Annual Losses of Quality-Adjusted Life Years (QALYs) for 14 Foodborne Pathogens in the United States. Foodborne Pathog. Dis. 2014, 11, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Gast, R.K. Salmonella Infections. In Diseases of Poultry; Saif, Y., Fadly, A., Glisson, J., McDougald, L., Nolan, L., Swayne, D., Eds.; Blackwell Publishing: Athens, GA, USA, 2008; pp. 619–674. [Google Scholar]
- EFSA. The European Union One Health 2020 Zoonoses Report. EFSA J. 2021, 19, e06971. [Google Scholar] [CrossRef]
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.A.; Roy, S.L.; Jones, J.L.; Griffin, P.M. Foodborne Illness Acquired in the United States-Major Pathogens. Emerg. Infect. Dis. 2011, 17, 7–15. [Google Scholar] [CrossRef]
- Gast, R.K. Microbiology of Shell Egg Production in the United States. In Producing Safe Eggs: Microbial Ecology of Salmonella; Ricke, C., Gast, R.K., Eds.; Academic Press: London, UK, 2017; pp. 25–44. ISBN 9780128025826. [Google Scholar]
- EC Regulation (EC) No 2160/2003 of the European Parliament and of the Council. Off. J. Eur. Union 2003, 2003, 1–15.
- Bueno, D.J.; Soria, M.A.; Soria, M.C.; Procura, F.; Rodriguez, F.I.; Godano, E.I. Egg Production Systems and Salmonella in South America. In Producing Safe Eggs—Microbial Ecology of Salmonella; Ricke, S.C., Gast, R.K., Eds.; Academic Press—Nikki Levy: London, UK, 2017; pp. 87–110. ISBN 978-0-12-802582-6. [Google Scholar]
- Gomez-Aldapa, C.A.; del Torres-Vitela, R.M.; Villaruel-Lopez, A.; Castro-Rosas, J. The Role of Food in Salmonella Infections. In Salmonella—A Dangerous Foodborne Pathogen; Mahmoud, B.S.M., Ed.; InTech: Rijeka, Croatia, 2012; pp. 17–31. [Google Scholar]
- Cox, J.; Woolcock, J.; Sartor, A. The Significance of Salmonella, Particularly S. Infantis, to the Australian Egg Industry; Cox, J.M., Woolcock, J.B., Sartor, A.L., Eds.; RIRDC Web Publication: Sydney, Australia, 2002. [Google Scholar]
- Zamudio, M.L.; Meza, A.; Bailón, H.; Martinez-Urtaza, J.; Campos, J. Experiencias En La Vigilancia Epidemiológica de Agentes Patógenos Transmitidos Por Alimentos a Través de Electroforesis En Campo Pulsado (PFGE) En El Perú. Rev. Peru. Med. Exp. Salud Publica 2011, 28, 128–135. [Google Scholar] [CrossRef] [Green Version]
- Walderama, W.; Quevedo, M.; Pastor, J.; Mantilla, Y.; Ortiz, M. Estudio de Prevalencia de Serotipos de Salmonella En Granjas Avícolas Tecnificadas En El Perú—Parte 1. MAP 2014, 5, 50–58. [Google Scholar]
- Revolledo, L.; Ferreira, A.J.P.P. Current Perspectives in Avian Salmonellosis: Vaccines and Immune Mechanisms of Protection. J. Appl. Poult. Res. 2012, 21, 418–431. [Google Scholar] [CrossRef]
- Holt, P.S.; Mitchell, B.W.; Gast, R.K. Airborne Horizontal Transmission of Salmonella Enteritidis in Molted Laying Chickens. Avian Dis. 1998, 42, 45–52. [Google Scholar] [CrossRef]
- Holt, P.S. Horizontal Transmission of Salmonella Enteritidis in Molted and Unmolted Laying Chickens. Avian Dis. 1995, 39, 239–249. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, M.; Takagi, M.; Takahashi, T.; Suzuki, S.; Sato, S.; Takehara, K. The Effect of the Flow of Air on Horizontal Transmission of Salmonella Enteritidis in Chickens. Avian Dis. 1997, 41, 354–360. [Google Scholar] [CrossRef]
- Palermo-Neto, J.; Quinteiro-Filho, W.M.; Ribeiro, A.; Ferraz-de-Paula, V.; Pinheiro, M.L.; Baskeville, E.; Akamine, A.T.; Astolfi-Ferreira, C.S.; Ferreira, A.J.P.; Gomes, A.V. Overcrowding Stress Decreases Macrophage Activity and Increases Salmonella Enteritidis Invasion in Broiler Chickens. Avian Pathol. 2014, 40, e30–e31. [Google Scholar] [CrossRef]
- Davies, R.H.; Breslin, M. Persistence of Salmonella Enteritidis Phage Type 4 in the Environment and Arthropod Vectors on an Empty Free-Range Chicken Farm. Environ. Microbiol. 2003, 5, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Zhang-Barber, L.; Turner, A.K.; Barrow, P.A. Vaccination for Control of Salmonella in Poultry. Vaccine 1999, 17, 2538–2545. [Google Scholar] [CrossRef]
- Djordjevic, S.P.; Cain, A.K.; Evershed, N.J.; Falconer, L.; Levings, R.S.; Lightfoot, D.; Hall, R.M. Emergence and Evolution of Multiply Antibiotic-Resistant Salmonella Enterica Serovar Paratyphi B D-Tartrate-Utilizing Strains Containing SGI1. Antimicrob. Agents Chemother. 2009, 53, 2319–2326. [Google Scholar] [CrossRef] [Green Version]
- Glynn, M.K.A.; Bopp, C.; Dewitt, W.; Dabney, P.; Mokhtar, M.; Angullo, F. Emergence of Multidrug-Resistant Salmonella Enterica Serotype Typhimurium DT104 Infections in the United States. N. Engl. J. Med. 1998, 338, 1333–1338. [Google Scholar] [CrossRef] [Green Version]
- Wegener, H.C. Antibiotics in Animal Feed and Their Role in Resistance Development. Curr. Opin. Microbiol. 2003, 6, 439–445. [Google Scholar] [CrossRef]
- Temelli, S.; Kahya, S.; Eyigor, A.; Carli, K.T. Incidence of Salmonella Enteritidis in Chicken Layer Flocks in Turkey: Results by Real-Time Polymerase Chain Reaction and International Organization for Standardization Culture Methods. Poult. Sci. 2010, 89, 1406–1410. [Google Scholar] [CrossRef]
- Gast, R.K. Pre-Harvest Measures to Control Salmonella in Laying Hens. In Improving the Safety and Quality of Eggs and Egg Products; Woodhead Publishing: Sawston, UK, 2011; Volume 2, pp. 120–145. ISBN 9780857090720. [Google Scholar]
- Cerquetti, M.C.; Gherardi, M.M. Orally Administered Attenuated Salmonella Enteritidis Reduces Chicken Cecal Carriage of Virulent Salmonella Challenge Organisms. Vet. Microbiol. 2000, 76, 185–192. [Google Scholar] [CrossRef]
- Van Immerseel, F.; Methner, U.; Rychlik, I.; Nagy, B.; Velge, P.; Martin, G.; Foster, N.; Ducatelle, R.; Barrow, P.A.; Ducatelle, R. Vaccination and Early Protection against Non-Host-Specific Salmonella Serotypes in Poultry: Exploitation of Innate Immunity and Microbial Activity. Epidemiol. Infect. 2005, 133, 959–978. [Google Scholar] [CrossRef] [PubMed]
- Gast, R.K. Serotype-Specific and Serotype-Independent Strategies for Preharvest Control of Food-Borne Salmonella in Poultry. Avian Dis. 2007, 51, 817–828. [Google Scholar] [CrossRef] [PubMed]
- De Cort, W.; Ducatelle, R.; Van Immerseel, F. Preharvest Measures to Improve the Safety of Eggs. In Producing Safe Eggs: Microbial Ecology of Salmonella; Ricke, S.C., Gast, R.K., Eds.; Elsevier: London, UK, 2016; pp. 259–280. ISBN 9780128025826. [Google Scholar]
- Desin, T.S.; Köster, W.; Potter, A.A. Salmonella Vaccines in Poultry: Past, Present and Future. Expert Rev. Vaccines 2013, 12, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Muotiala, A.; Hovi, M.; Mäkelä, P.H. Protective Immunity in Mouse Salmonellosis: Comparison of Smooth and Rough Live and Killed Vaccines. Microb. Pathog. 1989, 6, 51–60. [Google Scholar] [CrossRef]
- Babu, U.; Dalloul, R.A.; Okamura, M.; Lillehoj, H.S.; Xie, H.; Raybourne, R.B.; Gaines, D.; Heckert, R.A. Salmonella Enteritidis Clearance and Immune Responses in Chickens Following Salmonella Vaccination and Challenge. Vet. Immunol. Immunopathol. 2004, 101, 251–257. [Google Scholar] [CrossRef]
- Smith, H.W.H. The Use of Live Vaccines in Experimental Salmonella Gallinarum Infection in Chickens with Observations on Their Interference Effect. J. Hyg. 1956, 54, 419–432. [Google Scholar] [CrossRef] [Green Version]
- Feberwee, A.; de Vries, T.S.; Hartman, E.G.; de Wit, J.J.; Elbers, A.R.; de Jong, W.A. Vaccination against Salmonella Enteritidis in Dutch Commercial Layer Flocks with a Vaccine Based on a Live Salmonella Gallinarum 9R Strain: Evaluation of Efficacy, Safety, and Performance of Serologic Salmonella Tests. Avian Dis. 2001, 45, 83–91. [Google Scholar] [CrossRef]
- Feberwee, A.; De Vries, T.S.; Elbers, A.R.W.; De Jong, W.A. Results of a Salmonella Enteritidis Vaccination Field Trial in Broiler-Breeder Flocks in The Netherlands. Avian Dis. 2000, 44, 249–255. [Google Scholar] [CrossRef]
- Gast, R.K. Paratyphoid Infections. In Diseases of Poultry; Swayne, D.E., Glisson, J.R., McDougald, L.R., Nolan, L.K., Suarez, D.L., Nair, V., Eds.; John Wiley & Sons Inc.: Hoboken, NJ, USA, 2013; pp. 693–706. ISBN 978-0-470-95899-5. [Google Scholar]
- Filho, R.A.C.P.; de Paiva, J.B.; Argüello, Y.M.S.; da Silva, M.D.; Gardin, Y.; Resende, F.; Junior, A.B.; Sesti, L. Efficacy of Several Vaccination Programmes in Commercial Layer and Broiler Breeder Hens against Experimental Challenge with Salmonella Enterica Serovar Enteritidis. Avian Pathol. 2009, 38, 367–375. [Google Scholar] [CrossRef] [Green Version]
- Deguchi, K.; Yokoyama, E.; Honda, T.; Mizuno, K. Efficacy of a Novel Trivalent Inactivated Vaccine Against the Shedding of Salmonella in a Chicken Challenge Model. Avian Dis. 2009, 53, 281–286. [Google Scholar] [CrossRef]
- Aehle, S.; Curtiss, R. Current and Future Perspectives on Development of Salmonella Vaccine Technologies. In Producing Safe Eggs: Microbial Ecology of Salmonella; Rickie, S.C., Gast, R.K., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 281–299. ISBN 9780128025826. [Google Scholar]
- Pascual Anderson, M.D.R.; Calderón, Y.; Pascual, V. Investigación y Recuento de Enterobacteriaceae Lactosapositivas (Coliformes). In Microbiología Alimentaria: Metodología Analítica para Alimentos y Bebidas; Díaz de Santos, S.A., Ed.; Universidad de Alcalá: Madrid, Spain, 1992; pp. 17–20. [Google Scholar]
- Huberman, Y.D.; Velilla, A.V.; Terzolo, H.R. Evaluation of Different Live Salmonella Enteritidis Vaccine Schedules Administered during Layer Hen Rearing to Reduce Excretion, Organ Colonization, and Egg Contamination. Poult. Sci. 2019, 98, 2422–2431. [Google Scholar] [CrossRef]
- Huberman, Y.D.; Terzolo, H.R. Fowl Typhoid: Assessment of a Disinfectant Oral Dose to Reduce Horizontal Spread and Mortality. Avian Dis. 2008, 52, 320–323. [Google Scholar] [CrossRef]
- Audisio, M.C.; Terzolo, H.R. Virulence Analysis of a Salmonella Gallinarum Strain by Oral Inoculation of 20-Day-Old Chickens. Avian Dis. 2002, 46, 186–191. [Google Scholar] [CrossRef]
- Hoysak, D.J.; Weatherhead, P.J. Sampling Blood from Birds: A Technique and an Assessment of Its Effect. Condor 1991, 93, 746–752. [Google Scholar] [CrossRef] [Green Version]
- Williams, J.E.; Whittemore, A.D. Serological Diagnosis of Pullorum Disease with the Microagglutination System. Appl. Microbiol. 1971, 21, 394–399. [Google Scholar] [CrossRef] [PubMed]
- El-Enbaawy, M.I.; Ahmed, Z.A.M.; Sadek, M.A.; Ibrahim, H.M. Protective Efficacy of Salmonella Local Strains Representing Groups B, C, D and E in a Prepared Polyvalent Formalin Inactivated Oil Adjuvant Vaccine in Layers. Int. J. Microbiol. Res. 2013, 4, 288–295. [Google Scholar] [CrossRef]
- Brown, S.L.; Klein, G.C.; McKinney, F.T.; Jones, W.L. Safranin O-Stained Antigen Microagglutination Test for Detection of Brucella Antibodies. J. Clin. Microbiol. 1981, 13, 398–400. [Google Scholar] [CrossRef] [Green Version]
- National Research Council. Guide for the Care and Use of Laboratory Animals, 8th ed.; The National Academies Press: Washington, DC, USA, 2011; Volume 46, ISBN 97803-09154000. [Google Scholar]
- AVMA. Guidelines for the Euthanasia of Animals: 2013 Edition; 2013.0.1; AVMA: Schaumburg, IL, USA, 2013; ISBN 978-1-882691-21-0. [Google Scholar]
- Revolledo, L. Vaccines and Vaccination against Fowl Typhoid and Pullorum Disease: An Overview and Approaches in Developing Countries. J. Appl. Poult. Res. 2018, 3, 1–13. [Google Scholar] [CrossRef]
- Wigley, P.; Barrow, P. Salmonella in Preharvest Chickens: Current Understanding and Approaches to Control. In Producing Safe Eggs. Microbial Ecology of Salmonella; Rickie, S.C., Gast, R.K., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 139–159. ISBN 9780128025826. [Google Scholar]
- Barrow, P.A.; Lovell, M.A. Experimental Infection of Egg-Laying Hens with Salmonella Enteritidis Phage Type 4. Avian Pathol. 1991, 20, 335–348. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.J.; Mo, I.P.; Kang, M.S. Safety and Efficacy of Salmonella Gallinarum 9R Vaccine in Young Laying Chickens. Avian Pathol. 2005, 34, 362–366. [Google Scholar] [CrossRef]
- OIE. Fowl Typhoid and Pullorum Disease. In OIE Terrestrial Manual; OIE: Paris, France, 2018; pp. 1–17. [Google Scholar]
- Toyota-Hanatani, Y.; Ekawa, T.; Ohta, H.; Igimi, S.; Hara-Kudo, Y.; Sasai, K.; Baba, E. Public Health Assessment of Salmonella Enterica Serovar Enteritidis Inactivated-Vaccine Treatment in Layer Flocks. Appl. Environ. Microbiol. 2009, 75, 1005–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, E.; Snoeyenbos, G.; Weinack, O.; Smyser, C. Studies on the Use of 9R Strain of Salmonella Gallinarum as a Vaccine in Chickens. Avian Dis. 1981, 25, 38–52. [Google Scholar] [CrossRef] [PubMed]
- Dewaele, I.; Van Meirhaeghe, H.; Rasschaert, G.; Vanrobaeys, M.; De Graef, E.; Herman, L.; Ducatelle, R.; Heyndrickx, M.; De Reu, K. Persistent Salmonella Enteritidis Environmental Contamination on Layer Farms in the Context of an Implemented National Control Program with Obligatory Vaccination. Poult. Sci. 2012, 91, 282–291. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Group | Age of Vaccination (Weeks) | n | ||||
|---|---|---|---|---|---|---|
| Trial 1 | Trial 2 | Trial 3 | ||||
| Challenge Strain | ||||||
| 5 | 8 | 11 | S. Enteritidis | S. Typhimurium | S. Infantis | |
| 1 | 9R | - | Inactivated | 32 | 30 | 30 |
| 2 | 9R | Inactivated | Inactivated | 32 | 30 | 30 |
| 3 | - | Inactivated | Inactivated | 32 | 28 | 30 |
| 4 | - | - | - | 32 | 30 | 30 |
| Inoculation Dose (CFU) * | 1 × 107 | 4 × 107 | 6 × 107 | |||
| Group | Number of Positive Cloacal Swabs Samples | ||
|---|---|---|---|
| Trial 1 | Trial 2 | Trial 3 | |
| S. Enteritidis | S. Typhimurium | S. Infantis | |
| 1 | 124/191 a | 7/90 a | 5/90 a |
| 2 | 63/191 b | 7/90 a | 3/90 a |
| 3 | 106/191 c | 1/81 b | 5/90 a |
| 4 | 140/191 d | 17/90 c | 23/90 b |
| Number of Trial/Salmonella Strain | Group | n | Number of Positive Samples | Number of Positive Birds | |||
|---|---|---|---|---|---|---|---|
| Liver | Spleen | Caecum Contents | Total | ||||
| Trial 1 Salmonella Enteritidis | 1 | 25 | 8 a | 13 a | 20 ab | 41 a | 21 a |
| 2 | 25 | 7 a | 8 a | 15 a | 30 b | 18 a | |
| 3 | 25 | 18 b | 22 b | 22 b | 62 c | 23 ab | |
| 4 | 25 | 17 b | 24 b | 23 b | 64 c | 25 b | |
| Trial 2 Salmonella Typhimurium | 1 | 30 | 1 a | 1 a | 5 a | 7 a | 6 a |
| 2 | 30 | 2 a | 0 a | 4 a | 6 a | 6 a | |
| 3 | 28 | 4 a | 7 b | 6 a | 17 b | 11 a | |
| 4 | 30 | 10 b | 20 c | 13 b | 43 c | 23 b | |
| Trial 3 Salmonella Infantis | 1 | 30 | 3 a | 10 a | 13 a | 26 a | 18 a |
| 2 | 30 | 1 a | 8 a | 7 a | 16 a | 13 b | |
| 3 | 30 | 6 ab | 7 a | 6 a | 19 a | 13 b | |
| 4 | 30 | 13 b | 26 b | 23 b | 62 b | 29 c | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huberman, Y.D.; Caballero-García, M.; Rojas, R.; Ascanio, S.; Olmos, L.H.; Malena, R.; Lomónaco, J.; Nievas, P.; Chero, P.; Lévano-Gracía, J.; et al. The Efficacy of a Trivalent Inactivated Salmonella Vaccine Combined with the Live S. Gallinarum 9R Vaccine in Young Layers after Experimental Infections with S. Enteritidis, S. Typhimurium, and S. Infantis. Vaccines 2022, 10, 1113. https://doi.org/10.3390/vaccines10071113
Huberman YD, Caballero-García M, Rojas R, Ascanio S, Olmos LH, Malena R, Lomónaco J, Nievas P, Chero P, Lévano-Gracía J, et al. The Efficacy of a Trivalent Inactivated Salmonella Vaccine Combined with the Live S. Gallinarum 9R Vaccine in Young Layers after Experimental Infections with S. Enteritidis, S. Typhimurium, and S. Infantis. Vaccines. 2022; 10(7):1113. https://doi.org/10.3390/vaccines10071113
Chicago/Turabian StyleHuberman, Yosef Daniel, Melanie Caballero-García, Rober Rojas, Silvia Ascanio, Leandro Hipólito Olmos, Rosana Malena, Jorgelina Lomónaco, Paula Nievas, Paula Chero, Julio Lévano-Gracía, and et al. 2022. "The Efficacy of a Trivalent Inactivated Salmonella Vaccine Combined with the Live S. Gallinarum 9R Vaccine in Young Layers after Experimental Infections with S. Enteritidis, S. Typhimurium, and S. Infantis" Vaccines 10, no. 7: 1113. https://doi.org/10.3390/vaccines10071113