
Growth, Physiological, Nutrient-Uptake-Efficiency and Shade-Tolerance Responses of Cacao Genotypes under Different Shades

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Conditions
2.1.1. Cacao Genotypes
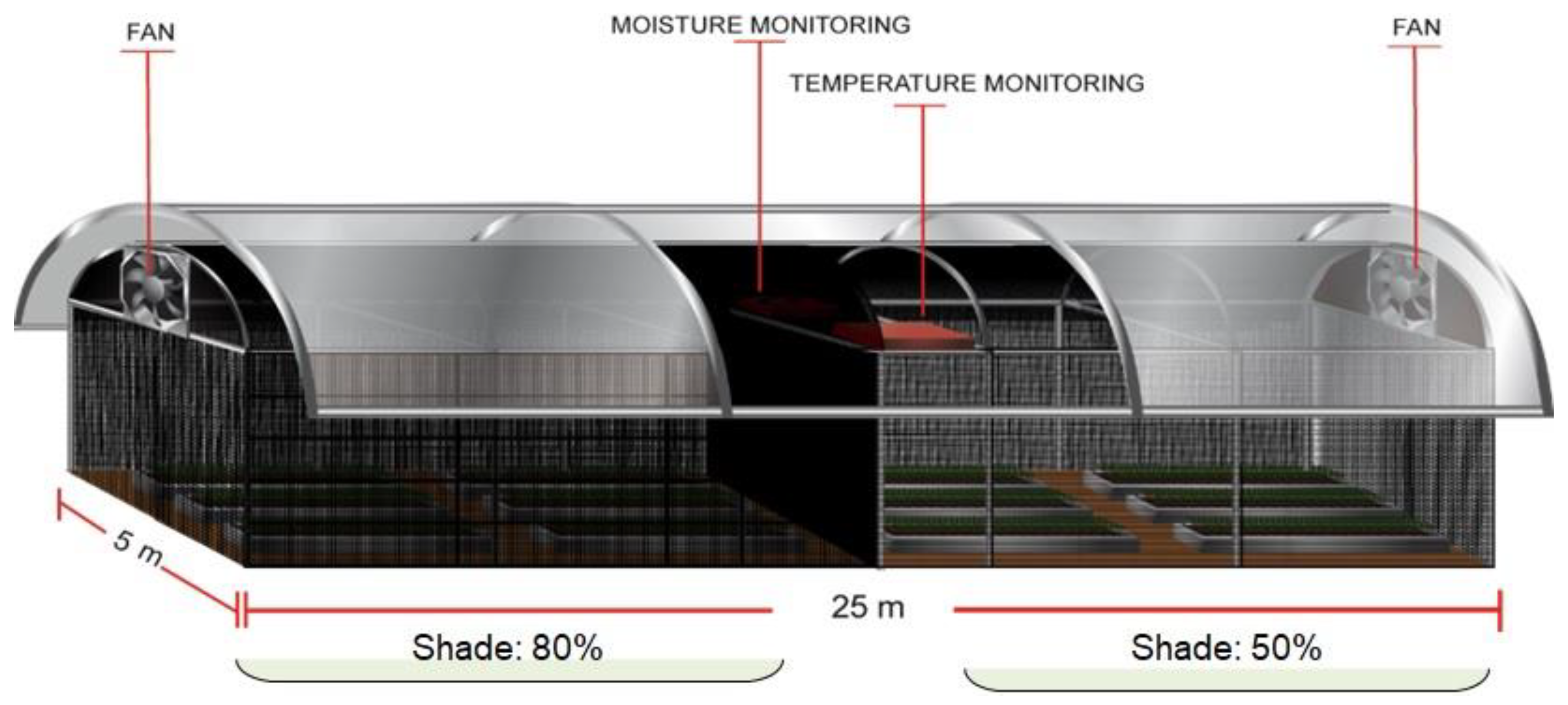
2.1.2. Shade (PPFD) Levels
2.1.3. Plant Growth Conditions
2.2. Biometric Parameters
2.3. Physiological Parameters
2.4. Nutrient Uptake Parameters
2.4.1. Concentration of Nutrients
2.4.2. Nutrient Uptake (U)
- SDW (shoot dry weight) = in g plant−1
- Element concentration = in g kg−1 (for macronutrients) or mg kg−1 (for micronutrients)
- U (or content) = g plant−1 (macronutrients) or mg plant−1 (micronutrients)
2.4.3. Nutrient Uptake Efficiency (NUE)
- element concentration = g kg−1 for macronutrients and mg kg−1 for micronutrients
- NUE = in g shoot g−1 of any macronutrient or in g shoot mg−1 of any micronutrient
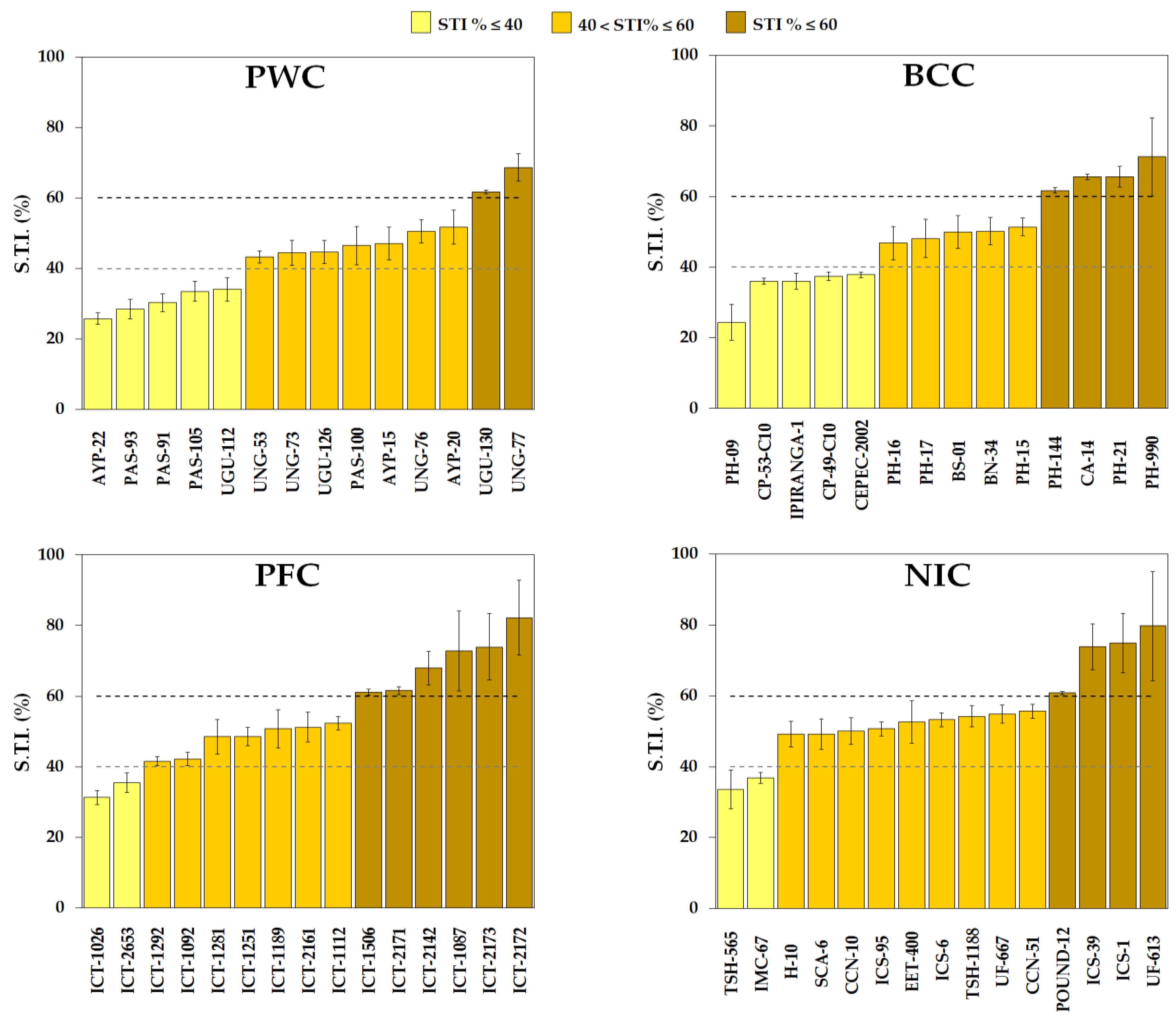
2.5. Shade Tolerance Index (STI)
- total (shoot + root) dry biomass (g plant−1) at 80% shade (PPFD 400 μmol m−2 s−1); this represents low light or heavy shade;
- total (shoot + root) dry biomass (g plant−1) at 50% shade (PPFD 1000 μmol m−2 s−1); this represents high light or low shade.
- Genotypes were classified into 3 groups: sensitive to shade (STI% ≤ 40), medium shade-tolerant (40 < STI% ≤ 60), and tolerant to shade (STI% > 60).
2.6. Statistical Analysis
3. Results and Discussion
3.1. Growth Parameters Influenced by Shade Levels
3.2. Physiological Parameters Influenced by Shade
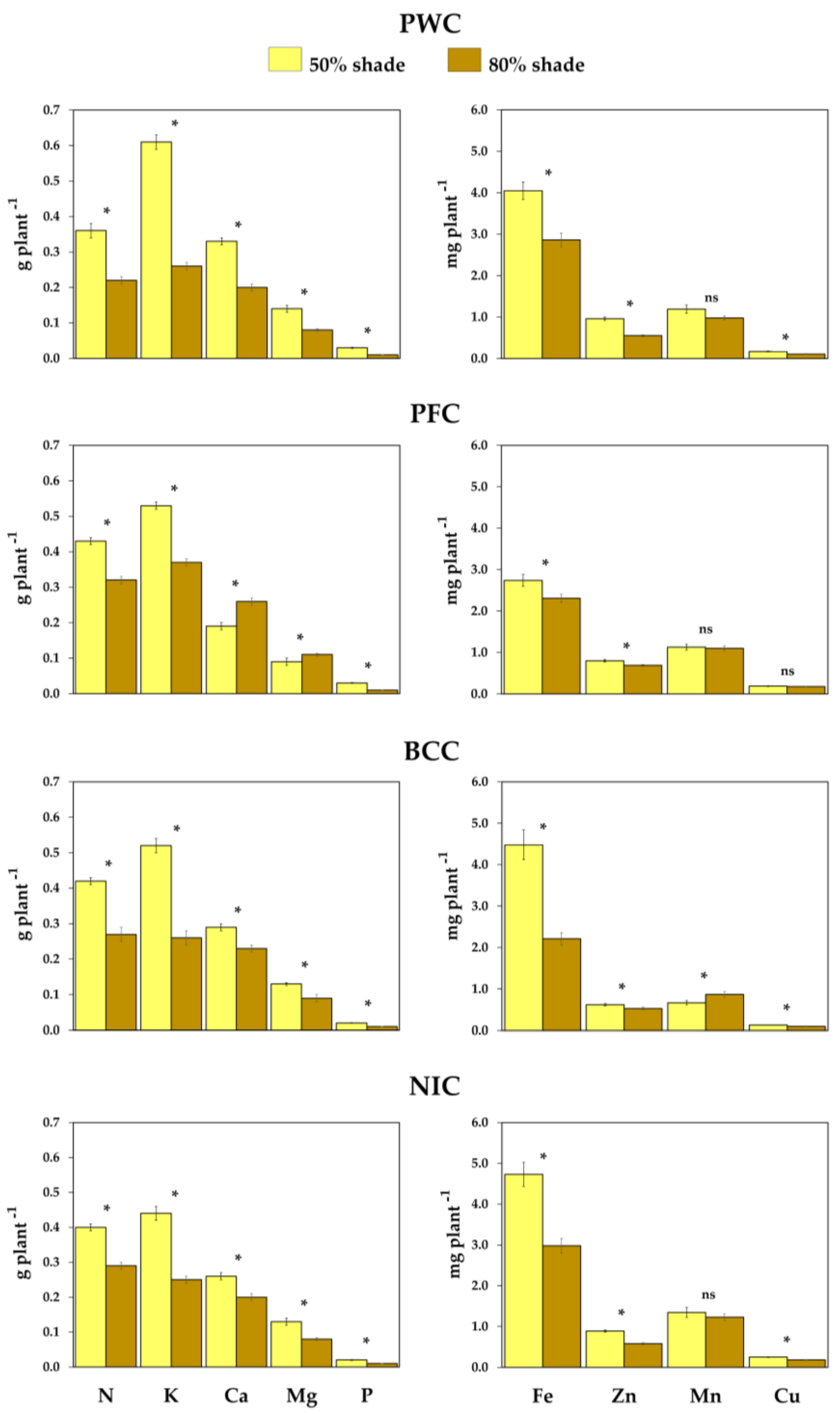
3.3. Nutritional Status Influenced by Shade
3.4. Cacao Genotypes Tolerant to Shade
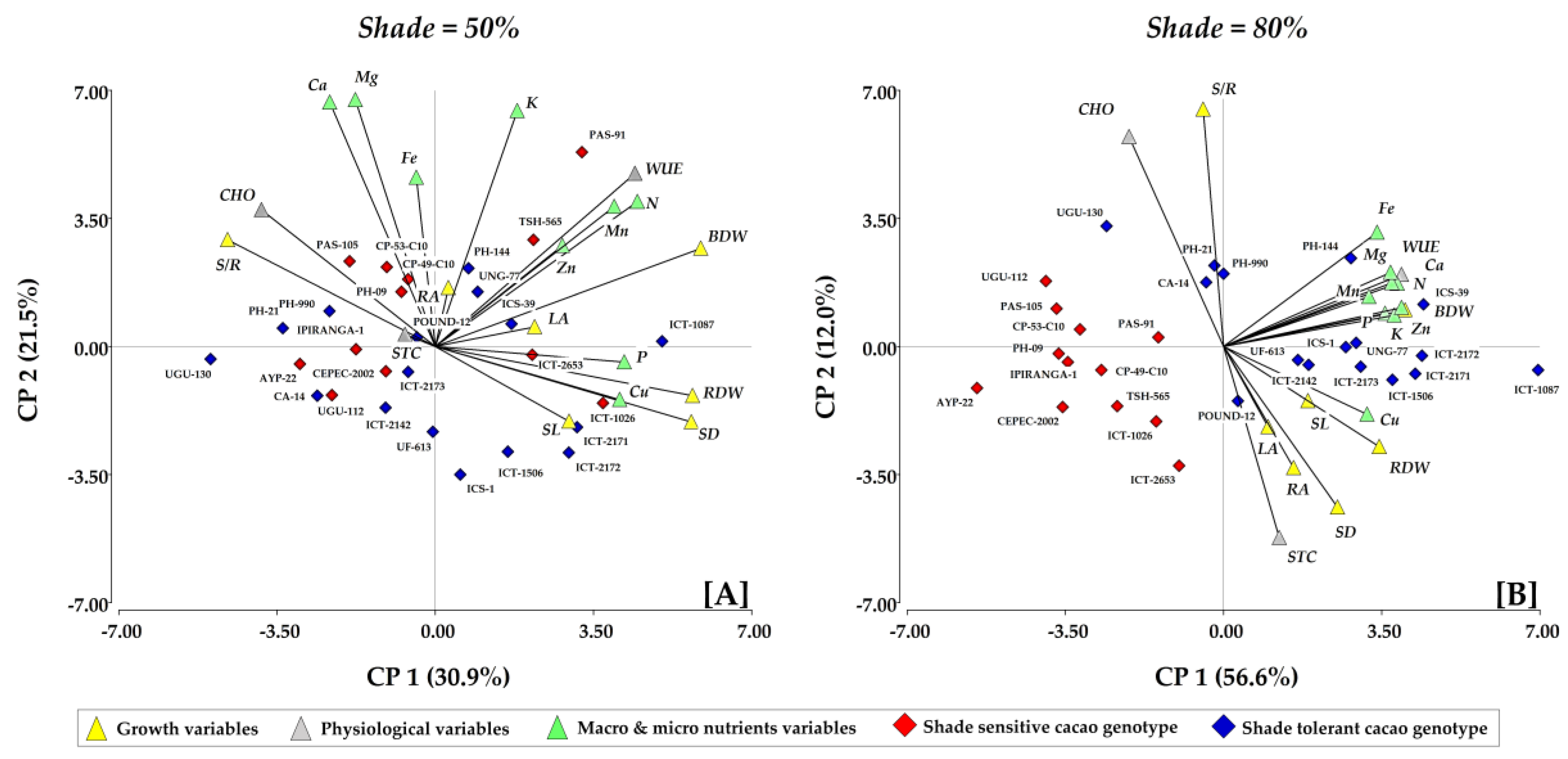
3.5. Interaction between Growth, Physiological Parameters, and Nutrient Uptake
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Feng, J.; Zhao, K.; He, D.; Fang, S.; Lee, T.; Chu, C.; He, F. Comparing shade tolerance measures of woody forest species. PeerJ 2018, 6, e5736. [Google Scholar] [CrossRef] [PubMed]
- Lienard, J.; Florescu, I.; Strigul, N. An appraisal of the classic forest succession paradigm with the shade tolerance index. PLoS ONE 2015, 10, e0117138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valladares, F.; Laanisto, L.; Niinemets, Ü.; Zavala, M.A. Shedding light on shade: Ecological perspectives of understorey plant life. Plant Ecol. Divers. 2016, 9, 237–251. [Google Scholar] [CrossRef] [Green Version]
- Martínez-García, J.F.; Galstyan, A.; Salla-Martret, M.; Cifuentes-Esquivel, N.; Gallemí, M.; Bou-Torrent, J. Regulatory components of shade avoidance syndrome. In Advances in Botanical Research; Elsevier: Amsterdam, The Netherlands, 2010; pp. 65–116. [Google Scholar] [CrossRef]
- Semchenko, M.; Lepik, M.; Götzenberger, L.; Zobel, K. Positive effect of shade on plant growth: Amelioration of stress or active regulation of growth rate?: Effects of shade on plant growth. J. Ecol. 2012, 100, 459–466. [Google Scholar] [CrossRef]
- Lahive, F.; Hadley, P.; Daymond, A.J. The physiological responses of cacao to the environment and the implications for climate change resilience. A review. Agron. Sustain. Dev. 2019, 39, 5. [Google Scholar] [CrossRef] [Green Version]
- Almeida, A.-A.F.; de Valle, R.R. Ecophysiology of the cacao tree. Braz. J. Plant Physiol. 2007, 19, 425–448. [Google Scholar] [CrossRef] [Green Version]
- Rice, R.A.; Greenberg, R. Cacao Cultivation and the Conservation of Biological Diversity. AMBIO J. Hum. Environ. 2000, 29, 167–173. [Google Scholar] [CrossRef]
- Acheampong, K.; Hadley, P.; Daymond, A.J. Photosynthetic activity and early growth of four cacao genotypes as influenced by different shade regimes under West African dry and wet season conditions. Exp. Agric. 2013, 49, 31–42. [Google Scholar] [CrossRef] [Green Version]
- Da Silva Branco, M.C.; De Almeida, A.-A.F.; Dalmolin, Â.C.; Ahnert, D.; Baligar, V.C. Influence of low light intensity and soil flooding on cacao physiology. Sci. Hortic. 2017, 217, 243–257. [Google Scholar] [CrossRef]
- Galyuon, I.K.A.; McDavid, C.R.; Lopez, F.B.; Spence, J.A. The effect of irradiance level on cocoa (Theobroma cacao L): I. Growth and leaf adaptations. Trop. Agric. 1996, 73, 23–28. [Google Scholar]
- Galyuon, I.K.A.; McDavid, C.R.; Lopez, F.B.; Spence, J.A. The effect of irradiance level on cocoa (Theobroma cacao L.): II. Gas exchange and chlorophyll fluorescence. Trop. Agric. 1996, 73, 29–33. [Google Scholar]
- Jagoret, P.; Ngnogue, H.T.; Malézieux, E.; Michel, I. Trajectories of cocoa agroforests and their drivers over time: Lessons from the Cameroonian experience. Eur. J. Agron. 2018, 101, 183–192. [Google Scholar] [CrossRef]
- Nijmeijer, A.; Lauri, P.-E.; Harmand, J.-M.; Freschet, G.T.; Essobo Nieboukaho, J.-D.; Fogang, P.K.; Enock, S.; Saj, S. Long-term dynamics of cocoa agroforestry systems established on lands previously occupied by savannah or forests. Agric. Ecosyst. Environ. 2019, 275, 100–111. [Google Scholar] [CrossRef]
- Saj, S.; Durot, C.; Mvondo Sakouma, K.; Tayo Gamo, K.; Avana-Tientcheu, M.-L. Contribution of associated trees to long-term species conservation, carbon storage, and sustainability: A functional analysis of tree communities in cacao plantations of Central Cameroon. Int. J. Agric. Sustain. 2017, 15, 282–302. [Google Scholar] [CrossRef]
- Tscharntke, T.; Clough, Y.; Bhagwat, S.A.; Buchori, D.; Faust, H.; Hertel, D.; Hölscher, D.; Juhrbandt, J.; Kessler, M.; Perfecto, I.; et al. Multifunctional shade-tree management in tropical agroforestry landscapes—A review: Multifunctional shade-tree management. J. Appl. Ecol. 2011, 48, 619–629. [Google Scholar] [CrossRef] [Green Version]
- Asante, W.A.; Acheampong, E.; Kyereh, E.; Kyereh, B. Farmers’ perspectives on climate change manifestations in smallholder cocoa farms and shifts in cropping systems in the forest-savannah transitional zone of Ghana. Land Use Policy 2017, 66, 374–381. [Google Scholar] [CrossRef]
- Niether, W.; Armengot, L.; Andres, C.; Schneider, M.; Gerold, G. Shade trees and tree pruning alter through fall and microclimate in cocoa (Theobroma cacao L.) production systems. Ann. For. Sci. 2018, 75, 38. [Google Scholar] [CrossRef] [Green Version]
- Abou Rajab, Y.; Leuschner, C.; Barus, H.; Tjoa, A.; Hertel, D. Cacao cultivation under diverse shade tree cover allows high carbon storage and sequestration without yield losses. PLoS ONE 2016, 11, e0149949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andres, C.; Blaser, W.J.; Dzahini-Obiatey, H.K.; Ameyaw, G.A.; Domfeh, O.K.; Awiagah, M.A.; Gattinger, A.; Schneider, M.; Offei, S.K.; Six, J. Agroforestry systems can mitigate the severity of cocoa swollen shoot virus disease. Agric. Ecosyst. Environ. 2018, 252, 83–92. [Google Scholar] [CrossRef]
- Bos, M.M.; Steffan-Dewenter, I.; Tscharntke, T. Shade tree management affects fruit abortion, insect pests and pathogens of cacao. Agric. Ecosyst. Environ. 2007, 120, 201–205. [Google Scholar] [CrossRef]
- Bunn, C.; Läderach, P.; Quaye, A.; Muilerman, S.; Noponen, M.R.A.; Lundy, M. Recommendation domains to scale out climate change adaptation in cocoa production in Ghana. Clim. Serv. 2019, 100123. [Google Scholar] [CrossRef]
- Hosseini Bai, S.; Trueman, S.J.; Nevenimo, T.; Hannet, G.; Bapiwai, P.; Poienou, M.; Wallace, H.M. Effects of shade-tree species and spacing on soil and leaf nutrient concentrations in cocoa plantations at 8 years after establishment. Agric. Ecosyst. Environ. 2017, 246, 134–143. [Google Scholar] [CrossRef]
- Sambuichi, R.H.R.; Vidal, D.B.; Piasentin, F.B.; Jardim, J.G.; Viana, T.G.; Menezes, A.A.; Mello, D.L.N.; Ahnert, D.; Baligar, V.C. Cabruca agroforests in southern Bahia, Brazil: Tree component, management practices and tree species conservation. Biodivers Conserv. 2012, 21, 1055–1077. [Google Scholar] [CrossRef]
- Vanhove, W.; Vanhoudt, N.; Van Damme, P. Effect of shade tree planting and soil management on rehabilitation success of a 22-year-old graded cocoa (Theobroma cacao L.) plantation. Agric. Ecosyst. Environ. 2016, 219, 14–25. [Google Scholar] [CrossRef]
- Vliet, J.A.; van Giller, K.E. Mineral nutrition of cocoa: A review. In Advances in Agronomy; Donald, L.S., Ed.; Academic Press: Cambridge, MA, USA, 2017; Volume 141, pp. 185–270. [Google Scholar]
- Abdulai, I.; Jassogne, L.; Graefe, S.; Asare, R.; Van Asten, P.; Läderach, P.; Vaast, P. Characterization of cocoa production, income diversification and shade tree management along a climate gradient in Ghana. PLoS ONE 2018, 13, e0195777. [Google Scholar] [CrossRef]
- Asare, R.; Markussen, B.; Asare, R.A.; Anim-Kwapong, G.; Ræbild, A. On-farm cocoa yields increase with canopy cover of shade trees in two agro-ecological zones in Ghana. Clim. Dev. 2019, 11, 435–445. [Google Scholar] [CrossRef] [Green Version]
- Blaser, W.J.; Oppong, J.; Hart, S.P.; Landolt, J.; Yeboah, E.; Six, J. Climate-smart sustainable agriculture in low-to-intermediate shade agroforests. Nat. Sustain. 2018, 1, 234–239. [Google Scholar] [CrossRef]
- Blaser, W.J.; Oppong, J.; Yeboah, E.; Six, J. Shade trees have limited benefits for soil fertility in cocoa agroforests. Agric. Ecosyst. Environ. 2017, 243, 83–91. [Google Scholar] [CrossRef] [Green Version]
- Jezeer, R.E.; Verweij, P.A.; Santos, M.J.; Boot, R.G.A. Shaded Coffee and Cocoa—Double Dividend for Biodiversity and Small-scale Farmers. Ecol. Econ. 2017, 140, 136–145. [Google Scholar] [CrossRef] [Green Version]
- Almeida, A.-A.F.; Gattward, J.N. Respostas do cacaueiro as variacoes da intensidad de luz. In Em Capitulo 2 de Cacao: Cultivo, Pesquisa e Inovação; de Souza Junior, J.O., Ed.; Editora da Universidade Estadual de Santa Cruz: Ilhéus, Brazil, 2018; pp. 35–58. [Google Scholar]
- Suárez Salazar, J.C.; Melgarejo, L.M.; Casanoves, F.; Di Rienzo, J.A.; DaMatta, F.M.; Armas, C. Photosynthesis limitations in cacao leaves under different agroforestry systems in the Colombian Amazon. PLoS ONE 2018, 13, e0206149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, G.A.R.; Lass, R.A. Cocoa, 4th ed.; Blackwell Science: Oxford, UK, 2001; 619p. [Google Scholar]
- Chazdon, R.L. Sunflecks and their importance to forest understorey plants. In Advances in Ecological Research; Elsevier: Amsterdam, The Netherlands, 1988; pp. 1–63. [Google Scholar] [CrossRef]
- De Castro, F. Light spectral composition in a tropical forest: Measurements and model. Tree Physiol. 2000, 20, 49–56. [Google Scholar] [CrossRef]
- Goldstein, G.; Santiago, L.S.; Campanello, P.I.; Avalos, G.; Zhang, Y.-J.; Villagra, M. Facing shortage or excessive light: How tropical and subtropical trees adjust their photosynthetic behavior and life history traits to a dynamic forest environment. In Tropical Tree Physiology; Goldstein, G., Santiago, L.S., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 319–336. [Google Scholar] [CrossRef]
- Liang, N.; Tang, Y.; Okuda, T. Is elevation of carbon dioxide concentration beneficial to seedling photosynthesis in the understory of tropical rain forests? Tree Physiol. 2001, 21, 1047–1055. [Google Scholar] [CrossRef] [Green Version]
- Pallardy, S.G. Photosynthesis. In Physiology of Woody Plants; Elsevier: Amsterdam, The Netherlands, 2008; pp. 107–167. [Google Scholar] [CrossRef]
- Pearcy, R.W. Photosynthetic gas exchange responses of Australian tropical forest trees in canopy, gap and understory micro-environments. Funct. Ecol. 1987, 1, 169. [Google Scholar] [CrossRef]
- Tang, Y.; Kachi, N.; Furukawa, A.; Awang, M.B. Heterogeneity of light availability and its effects on simulated carbon gain of tree leaves in a small gap and the understory in a tropical rain forest. Biotropica 1999, 31, 268–278. [Google Scholar] [CrossRef]
- Miyaji, K.-I.; Da Silva, W.S.; Alvim, P.D.T. Longevity of leaves of a tropical tree, Theobroma cacao, grown under shading, in relation to position within the canopy and time of emergence. New Phytol. 1997, 135, 445–454. [Google Scholar] [CrossRef] [Green Version]
- Baligar, V.C.; Bunce, J.A.; Machado, R.C.R.; Elson, M.K. Carbon dioxide and photosynthetic photon flux density effects on growth and mineral uptake of cacao. J. Food Agric. Environ. 2005, 3, 142–147. [Google Scholar]
- Baligar, V.C.; Bunce, J.A.; Machado, R.C.R.; Elson, M.K. Photosynthetic photon flux density, carbon dioxide concentration, and vapor pressure deficit effects on photosynthesis in cacao seedlings. Photosynthetica 2008, 46, 216–221. [Google Scholar] [CrossRef]
- Bastid, P.; Jimmy, I. Gas transfer measurements on young cocoa trees in field and modeling of photosynthetic activity. In Proceedings of the 14th International Cocoa Research Conference, Accra, Ghana, 13–18 October 2003; pp. 195–203. [Google Scholar]
- Hutcheon, W.F. Photosynthesis of Cocoa: Photosynthesis in Relation to the Light and Plant Nutrient Status. 1973–1974; Cocoa Research Institute of Ghana: Accra, Ghana, 1976. [Google Scholar]
- Raja Harun, R.M.; Hardwick, K. The effects of prolonged exposure to different light intensities on the photosynthesis of cocoa leaves. In Proceedings of the 10th International Cocoa Research Conference, Santo Domingo, Dominican Republic, 17–23 May 1988; pp. 205–209. [Google Scholar]
- Daymond, A.J.; Hadley, P.; Machado, R.C.R.; Ng, E. Canopy characteristics of contrasting clones of cacao (Theobroma cacao). Exp. Agric. 2002, 38, 359–367. [Google Scholar] [CrossRef]
- Motamayor, J.C.; Lachenaud, P.; da Silva e Mota, J.W.; Loor, R.; Kuhn, D.N.; Brown, J.S.; Schnell, R.J. Geographic and genetic population differentiation of the Amazonian chocolate tree (Theobroma cacao L.). PLoS ONE 2008, 3, e3311. [Google Scholar] [CrossRef] [Green Version]
- Pereira de Araújo, R.; Furtado de Almeida, A.-A.; Pinto Barroso, J.; Aparecida de Oliveira, R.; Pinto Gomes, F.; Ahnert, D.; Baligar, V. Molecular and morphophysiological responses cocoa leaves with different concentrations of anthocyanin to variations in light levels. Sci. Hortic. 2017, 224, 188–197. [Google Scholar] [CrossRef] [Green Version]
- Silva, F.C. Manual de análises químicas de solos, plantas e fertilizantes. In 2ª edição Revista e Ampliada; Embrapa Informação Tecnológica: Brasília, Brazil, 2009; 627p. [Google Scholar]
- Lamari, L. ASSESS 2.0. Image Analysis Sofware for Plant Disease Quantification. American Phytopathological Society; APS Press: St. Paul, MN, USA, 2008; 125p. [Google Scholar]
- United States Environmental Protection Agency (US EPA). Method 3050B: Acid Digestion of Sediments, Sludges, and Soils, Revision 2; United States Environmental Protection Agency (US EPA): Washington, DC, USA, 1996.
- Baligar, V.C.; Fageria, N.K.; He, Z.L. Nutrient use efficiency in plants. Commun. Soil Sci. Plant Anal. 2001, 32, 921–950. [Google Scholar] [CrossRef]
- McDonald, G.; Bovill, W.; Huang, C.; Lightfoot, D. Nutrient use efficiency. In Genomics and Breeding for Climate-Resilient Crops; Kole, C., Ed.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 333–393. [Google Scholar] [CrossRef]
- Di Rienzo, J.A.; Casanoves, F.; Balzarini, M.G.; Gonzalez, L.; Tablada, M.; Robledo, C.W. InfoStat Versión 2020; Centro de Transferencia InfoStat, FCA, Universidad Nacional de Córdoba: Córdoba, Argentina, 2020; Available online: http://www.infostat.com.ar (accessed on 25 March 2021).
- Poorter, H.; Niklas, K.J.; Reich, P.B.; Oleksyn, J.; Poot, P.; Mommer, L. Biomass allocation to leaves, stems and roots: Meta-analyses of interspecific variation and environmental control: Tansley review. New Phytologist. 2012, 193, 30–50. [Google Scholar] [CrossRef]
- Wartenberg, A.C.; Blaser, W.J.; Roshetko, J.M.; Van Noordwijk, M.; Six, J. Soil fertility and Theobroma cacao growth and productivity under commonly intercropped shade-tree species in Sulawesi, Indonesia. Plant Soil 2020, 453, 87–104. [Google Scholar] [CrossRef]
- Daymond, A.J.; Hadley, P. The effects of temperature and light integral on early vegetative growth and chlorophyll fluorescence of four contrasting genotypes of cacao (Theobroma cacao). Ann. Appl. Biol. 2004, 145, 257–262. [Google Scholar] [CrossRef]
- Daymond, A.J.; Tricker, P.J.; Hadley, P. Genotypic variation in photosynthesis in cacao is correlated with stomatal conductance and leaf nitrogen. Biol. Plant. 2011, 55, 99–104. [Google Scholar] [CrossRef]
- Baligar, V.C.; Elson, M.K.; Almeida, A.A.F.; de Araujo, Q.R.; Ahnert, D.; He, Z. The impact of carbon dioxide concentrations and low to adequate photosynthetic photon flux density on growth, physiology and nutrient use efficiency of juvenile cacao genotypes. Agronomy 2021, 11, 397. [Google Scholar] [CrossRef]
- Niether, W.; Schneidewind, U.; Fuchs, M.; Schneider, M.; Armengot, L. Below- and aboveground production in cocoa monocultures and agroforestry systems. Sci. Total Environ. 2019, 657, 558–567. [Google Scholar] [CrossRef]
- Almeida, A.-A.F.; Valle, R.R. Cacao: Ecophysiology of growth and production. In Ecophysiology of Tropical Tree Crops; DaMatta, F., Ed.; Nova Science Publishers: New York, NY, USA, 2010; p. 389. [Google Scholar]
- Acheampong, K.; Hadley, P.; Daymond, A.; Adu-Yeboah, P. The influence of shade and organic fertilizer treatments on the physiology and establishment of Theobroma cacao clones. Am. J. Exp. Agric. 2015, 6, 347–360. [Google Scholar] [CrossRef] [Green Version]
- Agudelo-Castañeda, G.A.; Cadena-Torres, J.; Almanza-Merchán, P.J.; Pinzón-Sandoval, E.H. Desempeño fisiológico de nueve genotipos de cacao (Theobroma cacao L.) bajo la sombra de tres especies forestales en Santander, Colombia. Rev. Colomb. Cienc. Hortic. 2018, 12, 223–232. [Google Scholar] [CrossRef] [Green Version]
- Jaimez, R.E.; Amores Puyutaxi, F.; Vasco, A.; Loor, R.G.; Tarqui, O.; Quijano, G.; Jimenez, J.C.; Tezara, W. Photosynthetic response to low and high light of cacao growing without shade in an area of low evaporative demand. Acta Biológica Colomb. 2018, 23, 95–103. [Google Scholar] [CrossRef]
- López-Marín, J.; Gálvez, A.; González, A.; Egea-Gilabert, C.; Fernández, J.A. Effect of shade on yield, quality and photosynthesis-related parameters of sweet pepper plants. Acta Hortic. 2012, 956, 545–552. [Google Scholar] [CrossRef]
- Yang, M.; Liu, M.; Lu, J.; Yang, H. Effects of shading on the growth and leaf photosynthetic characteristics of three forages in an apple orchard on the Loess Plateau of eastern Gansu, China. PeerJ 2019, 7, e7594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baliza, D.P.; Cunha, R.L.; Guimarães, R.J.; Barbosa, J.P.R.A.D.; Ávila, F.W.; Passos, A.M.A. Physiological characteristics and development of coffee plants under different shading levels. Rev. Bras. Ciênc. Agrár. Recife. 2012, 7, 7–43. [Google Scholar] [CrossRef] [Green Version]
- Cabala Rosand, P.; Santana, M.B.M.; de Santana, C.J.L. Cacao. In Detecting Mineral Nutrient Deficiencies in Tropical and Temperate Crops; Plucknett, D., Sprague, H.B., Eds.; CRC Press: Boca Raton, FL, USA, 1989. [Google Scholar] [CrossRef]
- Murray, D.B. Leaf analysis applied to cocoa. In Cocoa Growers’ Bulletin; Cadbury Brothers Ltd., Bournville: Birmingham, UK, 1967; Volume 9, pp. 25–35. [Google Scholar]
- Ahenkorah, Y.; Halm, B.; Appiah, M.; Akrofi, G.; Yirenkyi, J. Twenty Years’ Results from a Shade and Fertilizer Trial on Amazon Cocoa (Theobroma cacao) in Ghana. Exp. Agric. 1987, 23, 31–39. [Google Scholar] [CrossRef]
- Loue, A. Etude des Carences et des Deficincies Minerales sur le Cacaoyer (Study of Deficiencies and Mineral Deficiencies of Cacao); Institut Français du Café et du Cacao: Paris, France; Bulletin: Santa Rosa, CA, USA, 1961; Volume 1, p. 53. [Google Scholar]
- Snoeck, J. Cacao. In L’analyse Végétale dans le Contrôle de L’alimentation des Plantes Temperées et Tropicales; Martin-Prevel, P., Gagnard, J., Gautier, P., Eds.; Technique et doc. (Plant Analysis of Nutrient Requirements in Temperate and Tropical Plants); Lavoisier Publishing Inc.: New York, NY, USA, 1987; pp. 432–439. [Google Scholar]
- Bhargava, B.S.; Raghupati, H.B. Analysis of plants. In Methods of Analysis of Soils, Water and Fertilizers; Tondon, H.S., Ed.; FDCO: New Delhi, India, 1993; pp. 49–82. [Google Scholar]
- Isaac, M.E.; Timmer, V.R.; Quashie-Sam, S.J. Shade tree effects in an 8-year-old cocoa agroforestry system: Biomass and nutrient diagnosis of Theobroma cacao by vector analysis. Nutr. Cycl. Agroecosyst. 2007, 78, 155–165. [Google Scholar] [CrossRef]
- Bongers, F.J.; Evers, J.B.; Anten, N.P.R.; Pierik, R. From shade avoidance responses to plant performance at vegetation level: Using virtual plant modeling as a tool. New Phytol. 2014, 204, 268–272. [Google Scholar] [CrossRef] [Green Version]
- Portsmuth, A.; Niinemets, Ü. Structural and physiological plasticity in response to light and nutrients in five temperate deciduous woody species of contrasting shade tolerance. Funct. Ecol. 2007, 21. [Google Scholar] [CrossRef]
- Markesteijn, L.; Poorter, L. Seedling root morphology and biomass allocation of 62 tropical tree species in relation to drought- and shade-tolerance. J. Ecol. 2009, 97, 311–325. [Google Scholar] [CrossRef]
- Macfie, S.M.; Taylor, G.J. The effects of excess manganese on photosynthetic rate and concentration of chlorophyll in Triticum aestivum grown in solution culture. Physiol. Plant 1992, 85, 467–475. [Google Scholar] [CrossRef]
- Moroni, J.S.; Briggs, K.G.; Taylor, G.J. Chlorophyll content and leaf elongation rate in wheat seedlings as a measure of manganese tolerance. Plant Soil 1991, 136, 1–9. [Google Scholar] [CrossRef]
- Singh, P.; Misra, A.; Srivastava, N.K. Influence of Mn deficiency on growth, chlorophyll content, physiology, and essential monoterpene oil(s) in genotypes of spearmint (Mentha spicata L.). Photosynthetica 2001, 39, 473–476. [Google Scholar] [CrossRef]
- Bazihizina, N.; Colzi, I.; Giorni, E.; Mancuso, S.; Gonnelli, C. Photosynthesizing on metal excess: Copper differently induced changes in various photosynthetic parameters in copper tolerant and sensitive Silene paradoxa L. populations. Plant Sci. 2015, 232, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Mendoza, D.; Gil, F.E.; Escoboza-Garcia, F.; Santamaria, J.M.; Zapata-Perez, O. Copper stress on photosynthesis of black mangle (Avicennia germinans). An. Acad. Bras. Ciênc. 2013, 85, 665–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moustakas, M.; Ouzounidou, G.; Symeonidis, L.; Karataglis, S. Field study of the effects of excess copper on wheat photosynthesis and productivity. Soil Sci. Plant Nutr. 1997, 43, 531–539. [Google Scholar] [CrossRef] [Green Version]
- Pätsikkä, E.; Kairavuo, M.; Šeršen, F.; Aro, E.-M.; Tyystjärvi, E. Excess copper predisposes photosystem II to photoinhibition in vivo by outcompeting iron and causing decrease in leaf chlorophyll. Plant Physiol. 2002, 129, 1359–1367. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Wild Cacao Genotypes from River Basins of Peru Amazon (PWC) | Peruvian Farmers’ Cacao Genotypes (PFC) | Brazilian Genotypes (BCC) | National and International Genotypes (NIC) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| N° | Genotype | Origin | N° | Genotype | Origin | N° | Genotype | Origin | N° | Genotype | Origin |
| 1 | AYP-15 | Aypena | 1 | ICT-1026 | Mariscal Cáceres—Juanjui | 1 | BN-34 | Fazenda Boa Nova | 1 | CCN-10 | Ecuador |
| 2 | AYP-20 | Aypena | 2 | ICT-1087 | Mariscal Cáceres—Juanjui | 2 | BS-01 | Fazenda Bom Sossego | 2 | CCN-51 | Ecuador |
| 3 | AYP-22 | Aypena | 3 | ICT-1092 | Mariscal Cáceres—Juanjui | 3 | CA-14 | Fazenda Canta Galo | 3 | EET-400 | Ecuador |
| 4 | PAS-91 | Pastaza | 4 | ICT-1112 | Mariscal Cáceres—Juanjui | 4 | CEPEC-2002 | Centro de pesquisa do cacau | 4 | H-10 | Peru |
| 5 | PAS-93 | Pastaza | 5 | ICT-1189 | Mariscal Cáceres—Juanjui | 5 | CP-49-C10 | Centro de pesquisa do cacau | 5 | ICS-1 | Trinidad and Tobago |
| 6 | PAS-100 | Pastaza | 6 | ICT-1251 | Mariscal Cáceres—Juanjui | 6 | CP-53-C10 | Centro de pesquisa do cacau | 6 | ICS-6 | Trinidad and Tobago |
| 7 | PAS-105 | Pastaza | 7 | ICT-1281 | Mariscal Cáceres—Juanjui | 7 | IPIRANGA-1 | Cidade de Ipiranga | 7 | ICS-39 | Trinidad and Tobago |
| 8 | UGU-112 | Ungumayo | 8 | ICT-1292 | Mariscal Cáceres—Juanjui | 8 | PH-09 | Fazenda Porto Hibrido | 8 | ICS-95 | Trinidad and Tobago |
| 9 | UGU-126 | Ungumayo | 9 | ICT-1506 | Mariscal Cáceres—Juanjui | 9 | PH-15 | Fazenda Porto Hibrido | 9 | IMC-67 | Peru |
| 10 | UGU-130 | Ungumayo | 10 | ICT-2142 | Tocache | 10 | PH-16 | Fazenda Porto Hibrido | 10 | POUND-12 | Peru |
| 11 | UNG-53 | Ungurahui | 11 | ICT-2161 | Tocache | 11 | PH-17 | Fazenda Porto Hibrido | 11 | SCA-6 | Peru |
| 12 | UNG-73 | Ungurahui | 12 | ICT-2171 | Tocache | 12 | PH-21 | Fazenda Porto Hibrido | 12 | TSH-565 | Trinidad and Tobago |
| 13 | UNG-76 | Ungurahui | 13 | ICT-2172 | Tocache | 13 | PH-144 | Fazenda Porto Hibrido | 13 | TSH-1188 | Trinidad and Tobago |
| 14 | UNG-77 | Ungurahui | 14 | ICT-2173 | Tocache | 14 | PH-990 | Fazenda Porto Hibrido | 14 | UF-613 | Costa Rica |
| 15 | ICT-2653 | Tocache | 15 | UF-667 | Costa Rica | ||||||
| Growth Parameter * | Unit per Plant | PWC | PFC | BCC | NIC | Total | Source of Variation [df] | ||
|---|---|---|---|---|---|---|---|---|---|
| Shade (S) [1] | Genotype (G) [57] | G*S [57] | |||||||
| pv | |||||||||
| SL 50% | cm | 33.62 | 39.56 | 34.56 | 39.05 | 36.79 | 0.0004 | <0.0001 | 0.0072 |
| SL 80% | cm | 44.60 | 51.90 | 42.46 | 45.30 | 46.15 | |||
| SL Δ | cm | −10.98 | −12.34 | −7.90 | −6.25 | −9.36 | |||
| SD 50% | mm | 10.17 | 12.14 | 9.80 | 12.04 | 11.07 | 0.0001 | <0.0001 | 0.0003 |
| SD 80% | mm | 7.88 | 9.27 | 8.06 | 9.03 | 8.58 | |||
| SD Δ | mm | 2.29 | 2.87 | 1.74 | 3.01 | 2.49 | |||
| LA 50% | cm2 | 780.59 | 898.51 | 834.11 | 952.53 | 868.47 | 0.0001 | <0.0001 | 0.6657 |
| LA 80% | cm2 | 1023.99 | 1145.91 | 1037.56 | 1188.25 | 1110.27 | |||
| LA Δ | cm2 | −243.40 | −247.40 | −203.45 | −235.72 | −241.80 | |||
| RA 50% | cm2 | 241.50 | 300.99 | 317.97 | 327.91 | 297.69 | <0.0001 | <0.0001 | 0.0008 |
| RA 80% | cm2 | 295.97 | 388.69 | 337.21 | 418.28 | 361.54 | |||
| RA Δ | cm2 | −54.47 | −87.70 | −19.24 | −90.37 | −63.85 | |||
| RDW 50% | g | 6.94 | 10.61 | 7.21 | 9.02 | 8.50 | <0.0001 | <0.0001 | <0.0001 |
| RDW 80% | g | 2.38 | 3.93 | 2.92 | 3.60 | 3.23 | |||
| RDW Δ | g | 4.56 | 6.68 | 4.29 | 5.42 | 5.27 | |||
| BDW 50% | g | 30.81 | 35.58 | 30.05 | 32.08 | 32.19 | <0.0001 | <0.0001 | <0.0001 |
| BDW 80% | g | 13.83 | 18.76 | 14.43 | 17.16 | 16.11 | |||
| BDW Δ | g | 16.98 | 16.82 | 15.62 | 14.92 | 16.08 | |||
| S/R 50% | 3.92 | 2.44 | 3.45 | 2.64 | 3.09 | 0.0001 | <0.0001 | 0.0006 | |
| S/R 80% | 5.53 | 4.01 | 4.23 | 3.99 | 4.42 | ||||
| S/R Δ | −1.61 | −1.57 | −0.78 | −1.35 | −1.33 | ||||
| Physiological Parameter * | Unit | PWC | PFC | BCC | NIC | Total | Source of Variation [df] | ||
|---|---|---|---|---|---|---|---|---|---|
| Shade (S) [1] | Genotype (G) [57] | G*S [57] | |||||||
| pv | |||||||||
| CHL 50% | SPAD | 33.74 | 29.17 | 31.71 | 28.66 | 30.75 | <0.0001 | <0.0001 | <0.0001 |
| CHL 80% | SPAD | 33.55 | 30.44 | 33.46 | 31.19 | 32.12 | |||
| CHL Δ | SPAD | 0.19 | −1.27 | −1.75 | −2.53 | −1.37 | |||
| gs 50% | mmol m−2 s−1 | 188.40 | 184.01 | 226.86 | 183.78 | 195.35 | 0.0001 | 0.0002 | 0.0007 |
| gs 80% | mmol m−2 s−1 | 70.56 | 169.41 | 115.04 | 139.80 | 124.77 | |||
| gs Δ | mmol m−2 s−1 | 117.84 | 14.60 | 111.82 | 43.98 | 70.58 | |||
| WUE 50% | g L−1 | 1.33 | 1.39 | 1.27 | 1.28 | 1.31 | 0.0003 | <0.0001 | <0.0001 |
| WUE 80% | g L−1 | 0.98 | 1.27 | 0.98 | 1.16 | 1.09 | |||
| WUE Δ | g L−1 | 0.35 | 0.12 | 0.29 | 0.12 | 0.22 | |||
| Nutrient * | Unit per Plant | PWC | PFC | BCC | NIC | Total | Source of Variation [df] | ||
|---|---|---|---|---|---|---|---|---|---|
| Shade (S) [1] | Genotype (G) [57] | G*S [57] | |||||||
| N 50% | g kg−1 | 15.09 | 17.45 | 18.26 | 17.20 | 17.01 | <0.0001 | <0.0001 | 0.002 |
| N 80% | g kg−1 | 19.02 | 21.78 | 22.53 | 21.45 | 21.21 | |||
| N Δ | g kg−1 | −3.93 | −4.33 | −4.27 | −4.25 | −4.20 | |||
| K 50% | g kg−1 | 25.64 | 21.17 | 23.00 | 18.79 | 22.08 | 0.1034 | <0.0001 | <0.0001 |
| K 80% | g kg−1 | 21.82 | 24.66 | 21.52 | 18.53 | 21.63 | |||
| K Δ | g kg−1 | 3.82 | −3.49 | 1.48 | 0.26 | 0.45 | |||
| Ca 50% | g kg−1 | 13.81 | 7.81 | 12.66 | 11.01 | 11.26 | <0.0001 | <0.0001 | <0.0001 |
| Ca 80% | g kg−1 | 17.47 | 17.67 | 19.08 | 15.14 | 17.31 | |||
| Ca Δ | g kg−1 | −3.66 | −9.86 | −6.42 | −4.13 | −6.05 | |||
| Mg 50% | g kg−1 | 6.02 | 3.60 | 5.69 | 5.78 | 5.25 | <0.0001 | <0.0001 | <0.0001 |
| Mg 80% | g kg−1 | 6.40 | 7.20 | 7.52 | 5.69 | 6.69 | |||
| Mg Δ | g kg−1 | −0.38 | −3.60 | −1.83 | 0.09 | −1.44 | |||
| P 50% | g kg−1 | 1.18 | 1.14 | 0.93 | 0.87 | 1.03 | 0.0002 | <0.0001 | <0.0001 |
| P 80% | g kg−1 | 0.74 | 0.80 | 0.54 | 0.83 | 0.73 | |||
| P Δ | g kg−1 | 0.44 | 0.34 | 0.39 | 0.04 | 0.30 | |||
| Fe 50% | mg kg−1 | 173.09 | 110.65 | 194.57 | 202.88 | 169.83 | 0.0017 | <0.0001 | <0.0001 |
| Fe 80% | mg kg−1 | 240.11 | 156.13 | 181.33 | 218.20 | 198.54 | |||
| Fe Δ | mg kg−1 | −67.02 | −45.48 | 13.24 | −15.32 | −28.71 | |||
| Zn 50% | mg kg−1 | 40.39 | 32.26 | 27.74 | 39.20 | 34.93 | <0.0001 | <0.0001 | <0.0001 |
| Zn 80% | mg kg−1 | 47.91 | 46.62 | 44.50 | 43.18 | 45.53 | |||
| Zn Δ | mg kg−1 | −7.52 | −14.36 | −16.76 | −3.98 | −10.60 | |||
| Mn 50% | mg kg−1 | 50.60 | 44.73 | 29.28 | 57.25 | 45.66 | 0.0006 | 0.0006 | <0.0001 |
| Mn 80% | mg kg−1 | 84.15 | 72.87 | 76.98 | 88.24 | 80.56 | |||
| Mn Δ | mg kg−1 | −33.55 | −28.14 | −47.70 | −30.99 | −34.90 | |||
| Cu 50% | mg kg−1 | 7.04 | 7.49 | 5.71 | 10.85 | 7.82 | <0.0001 | <0.0001 | 0.0017 |
| Cu 80% | mg kg−1 | 9.43 | 12.42 | 8.33 | 14.15 | 11.16 | |||
| Cu Δ | mg kg−1 | −2.39 | −4.93 | −2.62 | −3.30 | −3.34 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arévalo-Gardini, E.; Farfán, A.; Barraza, F.; Arévalo-Hernández, C.O.; Zúñiga-Cernades, L.B.; Alegre, J.; Baligar, V.C. Growth, Physiological, Nutrient-Uptake-Efficiency and Shade-Tolerance Responses of Cacao Genotypes under Different Shades. Agronomy 2021, 11, 1536. https://doi.org/10.3390/agronomy11081536
Arévalo-Gardini E, Farfán A, Barraza F, Arévalo-Hernández CO, Zúñiga-Cernades LB, Alegre J, Baligar VC. Growth, Physiological, Nutrient-Uptake-Efficiency and Shade-Tolerance Responses of Cacao Genotypes under Different Shades. Agronomy. 2021; 11(8):1536. https://doi.org/10.3390/agronomy11081536
Chicago/Turabian StyleArévalo-Gardini, Enrique, Abel Farfán, Fiorella Barraza, César O. Arévalo-Hernández, Luis B. Zúñiga-Cernades, Julio Alegre, and Virupax C. Baligar. 2021. "Growth, Physiological, Nutrient-Uptake-Efficiency and Shade-Tolerance Responses of Cacao Genotypes under Different Shades" Agronomy 11, no. 8: 1536. https://doi.org/10.3390/agronomy11081536