Oocyte Selection for In Vitro Embryo Production in Bovine Species: Noninvasive Approaches for New Challenges of Oocyte Competence

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Morphological and Visual Markers for the Selection of the Best Oocytes
2.1. Ovarian Morphology
2.2. Follicle Size
2.3. Morphology of the Cumulus–Oocyte Complexes
2.4. Lipid Content
2.5. Cumulus Expansion and Oocyte Size
2.6. First Polar Body Assessment
2.7. Polarized Light Microscopy
2.7.1. Evaluation of the Meiotic Spindle and Zona Pellucida Birefringence
2.7.2. Assessment of the Zona Pellucida Birefringence
2.8. Brilliant Cresyl Blue (BCB) Staining
3. Non-invasive Molecular Approaches
3.1. Cell Death (Apoptosis) in Cumulus Cells
3.2. Transcriptomic and Proteomic of Cumulus Cells
3.3. Follicular Fluid Analysis
4. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Conti, M.; Franciosi, F. Acquisition of oocyte competence to develop as an embryo: Integrated nuclear and cytoplasmic events. Hum. Reprod. Update 2018, 24, 245–266. [Google Scholar] [CrossRef] [PubMed]
- Ruvolo, G.; Fattouh, R.R.; Bosco, L.; Brucculeri, A.M.; Cittadini, E. New molecular markers for the evaluation of gamete quality. J. Assist Reprod. Genet. 2013, 30, 207–212. [Google Scholar] [CrossRef] [Green Version]
- Van Wagtendonk-de Leeuw, A.M. Ovum pick up and in vitro production in the bovine after use in several generations: A 2005 status. Theriogenology 2006, 65, 914–925. [Google Scholar] [CrossRef] [PubMed]
- Rizos, D.; Clemente, M.; Bermejo-Alvarez, P.; de La Fuente, J.; Lonergan, P.; Gutierrez-Adan, A. Consequences of in vitro culture conditions on embryo development and quality. Reprod. Domest. Anim. 2008, 43 (Suppl. 4), 44–50. [Google Scholar] [CrossRef] [PubMed]
- Telfer, E.E.; Sakaguchi, K.; Clarkson, Y.L.; McLaughlin, M. In vitro growth of immature bovine follicles and oocytes. Reprod. Fertil. Dev. 2019, 32, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Lonergan, P.; Fair, T. Maturation of Oocytes in Vitro. Annu. Rev. Anim. Biosci. 2016, 4, 255–268. [Google Scholar] [CrossRef]
- Blondin, P.; Sirard, M.A. Oocyte and follicular morphology as determining characteristics for developmental competence in bovine oocytes. Mol. Reprod. Dev. 1995, 41, 54–62. [Google Scholar] [CrossRef]
- Krisher, R.L. The effect of oocyte quality on development. J. Anim. Sci. 2004, 82 (E-Suppl), E14–E23. [Google Scholar] [CrossRef]
- Rizos, D.; Ward, F.; Duffy, P.; Boland, M.P.; Lonergan, P. Consequences of bovine oocyte maturation, fertilization or early embryo development in vitro versus in vivo: Implications for blastocyst yield and blastocyst quality. Mol. Reprod. Dev. 2002, 61, 234–248. [Google Scholar] [CrossRef]
- Bogliotti, Y.S.; Wu, J.; Vilarino, M.; Okamura, D.; Soto, D.A.; Zhong, C.; Sakurai, M.; Sampaio, R.V.; Suzuki, K.; Izpisua Belmonte, J.C.; et al. Efficient derivation of stable primed pluripotent embryonic stem cells from bovine blastocysts. Proc. Natl. Acad. Sci. USA 2018, 115, 2090–2095. [Google Scholar] [CrossRef] [Green Version]
- Navarro, M.; Soto, D.A.; Pinzon, C.A.; Wu, J.; Ross, P.J. Livestock pluripotency is finally captured in vitro. Reprod. Fertil. Dev. 2019, 32, 11–39. [Google Scholar] [CrossRef] [PubMed]
- Goszczynski, D.E.; Cheng, H.; Demyda-Peyras, S.; Medrano, J.F.; Wu, J.; Ross, P.J. In vitro breeding: Application of embryonic stem cells to animal productiondagger. Biol. Reprod. 2019, 100, 885–895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebner, T.; Moser, M.; Sommergruber, M.; Tews, G. Selection based on morphological assessment of oocytes and embryos at different stages of preimplantation development: A review. Hum. Reprod. Update 2003, 9, 251–262. [Google Scholar] [CrossRef] [PubMed]
- Koyama, K.; Kang, S.S.; Huang, W.; Yanagawa, Y.; Takahashi, Y.; Nagano, M. Estimation of the optimal timing of fertilization for embryo development of in vitro-matured bovine oocytes based on the times of nuclear maturation and sperm penetration. J. Vet. Med. Sci. 2014, 76, 653–659. [Google Scholar] [CrossRef] [Green Version]
- Coticchio, G.; Sereni, E.; Serrao, L.; Mazzone, S.; Iadarola, I.; Borini, A. What criteria for the definition of oocyte quality? Ann. N. Y. Acad. Sci. 2004, 1034, 132–144. [Google Scholar] [CrossRef]
- Assidi, M.; Montag, M.; Van der Ven, K.; Sirard, M.A. Biomarkers of human oocyte developmental competence expressed in cumulus cells before ICSI: A preliminary study. J. Assist. Reprod. Genet. 2011, 28, 173–188. [Google Scholar] [CrossRef] [Green Version]
- Goovaerts, I.G.; Leroy, J.L.; Jorssen, E.P.; Bols, P.E. Noninvasive bovine oocyte quality assessment: Possibilities of a single oocyte culture. Theriogenology 2010, 74, 1509–1520. [Google Scholar] [CrossRef]
- Varisanga, M.D.; Sumantri, C.; Murakami, M.; Fahrudin, M.; Suzuki, T. Morphological classification of the ovaries in relation to the subsequent oocyte quality for IVF-produced bovine embryos. Theriogenology 1998, 50, 1015–1023. [Google Scholar] [CrossRef]
- Hagemann, L.J.; Beaumont, S.E.; Berg, M.; Donnison, M.J.; Ledgard, A.; Peterson, A.J.; Schurmann, A.; Tervit, H.R. Development during single IVP of bovine oocytes from dissected follicles: Interactive effects of estrous cycle stage, follicle size and atresia. Mol. Reprod. Dev. 1999, 53, 451–458. [Google Scholar] [CrossRef]
- Hagemann, L.J. Influence of the dominant follicle on oocytes from subordinate follicles. Theriogenology 1999, 51, 449–459. [Google Scholar] [CrossRef]
- Manjunatha, B.M.; Gupta, P.S.; Ravindra, J.P.; Devaraj, M.; Ramesh, H.S.; Nandi, S. In vitro developmental competence of buffalo oocytes collected at various stages of the estrous cycle. Theriogenology 2007, 68, 882–888. [Google Scholar] [CrossRef] [PubMed]
- Pirestani, A.; Hosseini, S.M.; Hajian, M.; Forouzanfar, M.; Moulavi, F.; Abedi, P.; Gourabi, H.; Shahverdi, A.; Taqi Dizaj, A.V.; Nasr Esfahani, M.H. Effect of ovarian cyclic status on in vitro embryo production in cattle. Int. J. Fertil. Steril. 2011, 4, 172–175. [Google Scholar] [PubMed]
- Penitente-Filho, J.M.; Jimenez, C.R.; Zolini, A.M.; Carrascal, E.; Azevedo, J.L.; Silveira, C.O.; Oliveira, F.A.; Torres, C.A. Influence of corpus luteum and ovarian volume on the number and quality of bovine oocytes. Anim. Sci. J. 2015, 86, 148–152. [Google Scholar] [CrossRef] [PubMed]
- Hajarian, H.; Shahsavari, M.H.; Karami-shabankareh, H.; Dashtizad, M. The presence of corpus luteum may have a negative impact on in vitro developmental competency of bovine oocytes. Reprod. Biol. 2016, 16, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Karami Shabankareh, H.; Shahsavari, M.H.; Hajarian, H.; Moghaddam, G. In vitro developmental competence of bovine oocytes: Effect of corpus luteum and follicle size. Iran J. Reprod. Med. 2015, 13, 615–622. [Google Scholar]
- Chohan, K.R.; Hunter, A.G. Effect of reproductive status on in vitro developmental competence of bovine oocytes. J. Vet. Sci. 2003, 4, 67–72. [Google Scholar] [CrossRef]
- Gandolfi, F.; Luciano, A.M.; Modina, S.; Ponzini, A.; Pocar, P.; Armstrong, D.T.; Lauria, A. The in vitro developmental competence of bovine oocytes can be related to the morphology of the ovary. Theriogenology 1997, 48, 1153–1160. [Google Scholar] [CrossRef]
- Smith, L.C.; Olivera-Angel, M.; Groome, N.P.; Bhatia, B.; Price, C.A. Oocyte quality in small antral follicles in the presence or absence of a large dominant follicle in cattle. J. Reprod. Fertil. 1996, 106, 193–199. [Google Scholar] [CrossRef] [Green Version]
- Arlotto, T.; Schwartz, J.L.; First, N.L.; Leibfried-Rutledge, M.L. Aspects of follicle and oocyte stage that affect in vitro maturation and development of bovine oocytes. Theriogenology 1996, 45, 943–956. [Google Scholar] [CrossRef]
- De Wit, A.A.; Wurth, Y.A.; Kruip, T.A. Effect of ovarian phase and follicle quality on morphology and developmental capacity of the bovine cumulus-oocyte complex. J. Anim. Sci. 2000, 78, 1277–1283. [Google Scholar] [CrossRef]
- Chian, R.C.; Chung, J.T.; Downey, B.R.; Tan, S.L. Maturational and developmental competence of immature oocytes retrieved from bovine ovaries at different phases of folliculogenesis. Reprod. Biomed. Online 2002, 4, 127–132. [Google Scholar] [CrossRef]
- Lonergan, P.; Monaghan, P.; Rizos, D.; Boland, M.P.; Gordon, I. Effect of follicle size on bovine oocyte quality and developmental competence following maturation, fertilization, and culture in vitro. Mol. Reprod. Dev. 1994, 37, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Iwata, H.; Hashimoto, S.; Ohota, M.; Kimura, K.; Shibano, K.; Miyake, M. Effects of follicle size and electrolytes and glucose in maturation medium on nuclear maturation and developmental competence of bovine oocytes. Reproduction 2004, 127, 159–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lequarre, A.S.; Vigneron, C.; Ribaucour, F.; Holm, P.; Donnay, I.; Dalbies-Tran, R.; Callesen, H.; Mermillod, P. Influence of antral follicle size on oocyte characteristics and embryo development in the bovine. Theriogenology 2005, 63, 841–859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caixeta, E.S.; Ripamonte, P.; Franco, M.M.; Junior, J.B.; Dode, M.A. Effect of follicle size on mRNA expression in cumulus cells and oocytes of Bos indicus: An approach to identify marker genes for developmental competence. Reprod. Fertil. Dev. 2009, 21, 655–664. [Google Scholar] [CrossRef]
- Nivet, A.L.; Bunel, A.; Labrecque, R.; Belanger, J.; Vigneault, C.; Blondin, P.; Sirard, M.A. FSH withdrawal improves developmental competence of oocytes in the bovine model. Reproduction 2012, 143, 165–171. [Google Scholar] [CrossRef] [Green Version]
- De Bem, T.; Adona, P.R.; Bressan, F.F.; Mesquita, L.G.; Chiaratti, M.R.; Meirelles, F.V.; Leal, C. The influence of morphology, follicle size and Bcl-2 and Bax transcripts on the developmental competence of bovine oocytes. Reprod. Domest. Anim. 2014, 49, 576–583. [Google Scholar] [CrossRef]
- Kauffold, J.; Amer, H.A.; Bergfeld, U.; Weber, W.; Sobiraj, A. The in vitro developmental competence of oocytes from juvenile calves is related to follicular diameter. J. Reprod. Dev. 2005, 51, 325–332. [Google Scholar] [CrossRef] [Green Version]
- Hendriksen, P.J.; Vos, P.L.; Steenweg, W.N.; Bevers, M.M.; Dieleman, S.J. Bovine follicular development and its effect on the in vitro competence of oocytes. Theriogenology 2000, 53, 11–20. [Google Scholar] [CrossRef]
- Annes, K.; Muller, D.B.; Vilela, J.A.P.; Valente, R.S.; Caetano, D.P.; Cibin, F.W.S.; Milazzotto, M.P.; Mesquita, F.S.; Belaz, K.R.A.; Eberlin, M.N.; et al. Influence of follicle size on bovine oocyte lipid composition, follicular metabolic and stress markers, embryo development and blastocyst lipid content. Reprod. Fertil. Dev. 2019, 31, 462–472. [Google Scholar] [CrossRef]
- Alves, G.P.; Cordeiro, F.B.; Bruna de Lima, C.; Annes, K.; Cristina Dos Santos, E.; Ispada, J.; Fontes, P.K.; Nogueira, M.F.G.; Nichi, M.; Milazzotto, M.P. Follicular environment as a predictive tool for embryo development and kinetics in cattle. Reprod. Fertil. Dev. 2019, 31, 451–461. [Google Scholar] [CrossRef] [PubMed]
- Labrecque, R.; Fournier, E.; Sirard, M.A. Transcriptome analysis of bovine oocytes from distinct follicle sizes: Insights from correlation network analysis. Mol. Reprod. Dev. 2016, 83, 558–569. [Google Scholar] [CrossRef] [PubMed]
- Moussa, M.; Shu, J.; Zhang, X.H.; Zeng, F. Maternal control of oocyte quality in cattle “a review”. Anim. Reprod. Sci. 2015, 155, 11–27. [Google Scholar] [CrossRef] [PubMed]
- Tello, M.F.; Lorenzo, M.S.; Luchetti, C.G.; Maruri, A.; Cruzans, P.R.; Alvarez, G.M.; Gambarotta, M.C.; Salamone, D.F.; Cetica, P.D.; Lombardo, D.M. Apoptosis in porcine cumulus-oocyte complexes: Relationship with their morphology and the developmental competence. Mol. Reprod. Dev. 2020, 87, 274–283. [Google Scholar] [CrossRef] [PubMed]
- Leibfried, L.; First, N.L. Characterization of bovine follicular oocytes and their ability to mature in vitro. J. Anim. Sci. 1979, 48, 76–86. [Google Scholar] [CrossRef] [PubMed]
- Hazeleger, N.L.; Hill, D.J.; Stubbing, R.B.; Walton, J.S. Relationship of morphology and follicular fluid environment of bovine oocytes to their developmental potential in vitro. Theriogenology 1995, 43, 509–522. [Google Scholar] [CrossRef]
- Madison, V.; Avery, B.; Greve, T. Selection of immature bovine oocytes for developmental potential in vitro. Anim. Reprod. Sci. 1992, 27, 1–11. [Google Scholar] [CrossRef]
- Boni, R.; Cuomo, A.; Tosti, E. Developmental potential in bovine oocytes is related to cumulus-oocyte complex grade, calcium current activity, and calcium stores. Biol. Reprod. 2002, 66, 836–842. [Google Scholar] [CrossRef] [Green Version]
- Bilodeau-Goeseels, S.; Panich, P. Effects of oocyte quality on development and transcriptional activity in early bovine embryos. Anim. Reprod. Sci. 2002, 71, 143–155. [Google Scholar] [CrossRef]
- Emanuelli, I.P.; Costa, C.B.; Rafagnin Marinho, L.S.; Seneda, M.M.; Meirelles, F.V. Cumulus-oocyte interactions and programmed cell death in bovine embryos produced in vitro. Theriogenology 2019, 126, 81–87. [Google Scholar] [CrossRef]
- Yuan, Y.Q.; Van Soom, A.; Leroy, J.L.; Dewulf, J.; Van Zeveren, A.; de Kruif, A.; Peelman, L.J. Apoptosis in cumulus cells, but not in oocytes, may influence bovine embryonic developmental competence. Theriogenology 2005, 63, 2147–2163. [Google Scholar] [CrossRef]
- De Loos, F.; van Vliet, C.; van Maurik, P.; Kruip, T.A. Morphology of immature bovine oocytes. Gamete Res. 1989, 24, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Hawk, H.; Wall, R. Improved yields of bovine blastocysts from in vitro-produced oocytes. I. Selection of oocytes and zygotes. Theriogenology 1994, 41, 1571–1583. [Google Scholar] [CrossRef]
- Dunning, K.R.; Russell, D.L.; Robker, R.L. Lipids and oocyte developmental competence: The role of fatty acids and beta-oxidation. Reproduction 2014, 148, R15–R27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Genicot, G.; Leroy, J.L.; Soom, A.V.; Donnay, I. The use of a fluorescent dye, Nile red, to evaluate the lipid content of single mammalian oocytes. Theriogenology 2005, 63, 1181–1194. [Google Scholar] [CrossRef]
- Van Blerkom, J.; Bell, H.; Weipz, D. Cellular and developmental biological aspects of bovine meiotic maturation, fertilization, and preimplantation embryogenesis in vitro. J. Electron. Microsc. Tech. 1990, 16, 298–323. [Google Scholar] [CrossRef]
- McKeegan, P.J.; Sturmey, R.G. The role of fatty acids in oocyte and early embryo development. Reprod. Fertil. Dev. 2011, 24, 59–67. [Google Scholar] [CrossRef]
- Sturmey, R.G.; Reis, A.; Leese, H.J.; McEvoy, T.G. Role of fatty acids in energy provision during oocyte maturation and early embryo development. Reprod. Domest. Anim. 2009, 44 (Suppl. 3), 50–58. [Google Scholar] [CrossRef]
- Kim, J.Y.; Kinoshita, M.; Ohnishi, M.; Fukui, Y. Lipid and fatty acid analysis of fresh and frozen-thawed immature and in vitro matured bovine oocytes. Reproduction 2001, 122, 131–138. [Google Scholar] [CrossRef]
- Pavani, K.C.; Rocha, A.; Oliveira, E.; da Silva, F.M.; Sousa, M. Novel ultrastructural findings in bovine oocytes matured in vitro. Theriogenology 2020, 143, 88–97. [Google Scholar] [CrossRef]
- Dadarwal, D.; Honparkhe, M.; Dias, F.C.; Alce, T.; Lessard, C.; Singh, J. Effect of superstimulation protocols on nuclear maturation and distribution of lipid droplets in bovine oocytes. Reprod. Fertil. Dev. 2015, 27, 1137–1146. [Google Scholar] [CrossRef] [PubMed]
- Auclair, S.; Uzbekov, R.; Elis, S.; Sanchez, L.; Kireev, I.; Lardic, L.; Dalbies-Tran, R.; Uzbekova, S. Absence of cumulus cells during in vitro maturation affects lipid metabolism in bovine oocytes. Am. J. Physiol. Endocrinol. Metab. 2013, 304, E599–E613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fair, T.; Hulshof, S.C.; Hyttel, P.; Greve, T.; Boland, M. Oocyte ultrastructure in bovine primordial to early tertiary follicles. Anat. Embryol. (Berl) 1997, 195, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Prates, E.G.; Nunes, J.T.; Pereira, R.M. A role of lipid metabolism during cumulus-oocyte complex maturation: Impact of lipid modulators to improve embryo production. Mediators Inflamm. 2014, 2014, 692067. [Google Scholar] [CrossRef] [PubMed]
- Leroy, J.L.; Genicot, G.; Donnay, I.; Van Soom, A. Evaluation of the lipid content in bovine oocytes and embryos with nile red: A practical approach. Reprod. Domest. Anim. 2005, 40, 76–78. [Google Scholar] [CrossRef]
- Salamone, D.F.; Canel, N.G.; Rodriguez, M.B. Intracytoplasmic sperm injection in domestic and wild mammals. Reproduction 2017, 154, F111–F124. [Google Scholar] [CrossRef]
- Nagano, M.; Katagiri, S.; Takahashi, Y. Relationship between bovine oocyte morphology and in vitro developmental potential. Zygote 2006, 14, 53–61. [Google Scholar] [CrossRef]
- Jeong, W.J.; Cho, S.J.; Lee, H.S.; Deb, G.K.; Lee, Y.S.; Kwon, T.H.; Kong, I.K. Effect of cytoplasmic lipid content on in vitro developmental efficiency of bovine IVP embryos. Theriogenology 2009, 72, 584–589. [Google Scholar] [CrossRef]
- Nagano, M. Acquisition of developmental competence and in vitro growth culture of bovine oocytes. J. Reprod. Dev. 2019, 65, 195–201. [Google Scholar] [CrossRef] [Green Version]
- Prates, E.G.; Marques, C.C.; Baptista, M.C.; Vasques, M.I.; Carolino, N.; Horta, A.E.; Charneca, R.; Nunes, J.T.; Pereira, R.M. Fat area and lipid droplet morphology of porcine oocytes during in vitro maturation with trans-10, cis-12 conjugated linoleic acid and forskolin. Animal 2013, 7, 602–609. [Google Scholar] [CrossRef]
- Jasensky, J.; Boughton, A.P.; Khmaladze, A.; Ding, J.; Zhang, C.; Swain, J.E.; Smith, G.W.; Chen, Z.; Smith, G.D. Live-cell quantification and comparison of mammalian oocyte cytosolic lipid content between species, during development, and in relation to body composition using nonlinear vibrational microscopy. Analyst 2016, 141, 4694–4706. [Google Scholar] [CrossRef] [PubMed]
- Machado, M.F.; Caixeta, E.S.; Sudiman, J.; Gilchrist, R.B.; Thompson, J.G.; Lima, P.F.; Price, C.A.; Buratini, J. Fibroblast growth factor 17 and bone morphogenetic protein 15 enhance cumulus expansion and improve quality of in vitro-produced embryos in cattle. Theriogenology 2015, 84, 390–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, K.; Hansen, P.J.; Ealy, A.D. Fibroblast growth factor 10 enhances bovine oocyte maturation and developmental competence in vitro. Reproduction 2010, 140, 815–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, K.; Yamashita, S.; Hoshi, H. Influence of epidermal growth factor and transforming growth factor-alpha on in vitro maturation of cumulus cell-enclosed bovine oocytes in a defined medium. J. Reprod. Fertil. 1994, 100, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Allworth, A.E.; Albertini, D.F. Meiotic maturation in cultured bovine oocytes is accompanied by remodeling of the cumulus cell cytoskeleton. Dev. Biol. 1993, 158, 101–112. [Google Scholar] [CrossRef]
- Furnus, C.C.; de Matos, D.G.; Moses, D.F. Cumulus expansion during in vitro maturation of bovine oocytes: Relationship with intracellular glutathione level and its role on subsequent embryo development. Mol. Reprod. Dev. 1998, 51, 76–83. [Google Scholar] [CrossRef]
- Choi, Y.H.; Carnevale, E.M.; Seidel, G.E., Jr.; Squire, E.L. Effects of gonadotropins on bovine oocytes matured in TCM-199. Theriogenology 2001, 56, 661–670. [Google Scholar] [CrossRef]
- Anchordoquy, J.P.; Anchordoquy, J.M.; Sirini, M.A.; Testa, J.A.; Peral-Garcia, P.; Furnus, C.C. The importance of manganese in the cytoplasmic maturation of cattle oocytes: Blastocyst production improvement regardless of cumulus cells presence during in vitro maturation. Zygote 2016, 24, 139–148. [Google Scholar] [CrossRef]
- Dovolou, E.; Messinis, I.E.; Periquesta, E.; Dafopoulos, K.; Gutierrez-Adan, A.; Amiridis, G.S. Ghrelin accelerates in vitro maturation of bovine oocytes. Reprod. Domest. Anim. 2014, 49, 665–672. [Google Scholar] [CrossRef]
- Rosa, D.E.; Anchordoquy, J.M.; Anchordoquy, J.P.; Sirini, M.A.; Testa, J.A.; Mattioli, G.A.; Furnus, C.C. Analyses of apoptosis and DNA damage in bovine cumulus cells after in vitro maturation with different copper concentrations: Consequences on early embryo development. Zygote 2016, 24, 869–879. [Google Scholar] [CrossRef]
- Marei, W.F.; Ghafari, F.; Fouladi-Nashta, A.A. Role of hyaluronic acid in maturation and further early embryo development of bovine oocytes. Theriogenology 2012, 78, 670–677. [Google Scholar] [CrossRef] [PubMed]
- Fukui, Y. Effect of follicle cells on the acrosome reaction, fertilization, and developmental competence of bovine oocytes matured in vitro. Mol. Reprod. Dev. 1990, 26, 40–46. [Google Scholar] [CrossRef]
- Fair, T.; Hyttel, P.; Greve, T. Bovine oocyte diameter in relation to maturational competence and transcriptional activity. Mol. Reprod. Dev. 1995, 42, 437–442. [Google Scholar] [CrossRef]
- Fair, T.; Hyttel, P.; Greve, T.; Boland, M. Nucleus structure and transcriptional activity in relation to oocyte diameter in cattle. Mol. Reprod. Dev. 1996, 43, 503–512. [Google Scholar] [CrossRef]
- Anguita, B.; Jimenez-Macedo, A.R.; Izquierdo, D.; Mogas, T.; Paramio, M.T. Effect of oocyte diameter on meiotic competence, embryo development, p34 (cdc2) expression and MPF activity in prepubertal goat oocytes. Theriogenology 2007, 67, 526–536. [Google Scholar] [CrossRef] [PubMed]
- Otoi, T.; Yamamoto, K.; Koyama, N.; Tachikawa, S.; Suzuki, T. Bovine oocyte diameter in relation to developmental competence. Theriogenology 1997, 48, 769–774. [Google Scholar] [CrossRef]
- Huang, W.; Nagano, M.; Kang, S.S.; Yanagawa, Y.; Takahashi, Y. Effects of in vitro growth culture duration and prematuration culture on maturational and developmental competences of bovine oocytes derived from early antral follicles. Theriogenology 2013, 80, 793–799. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Kanno, C.; Huang, W.; Kang, S.S.; Yanagawa, Y.; Nagano, M. Effect of bone morphogenetic protein-4 on in vitro growth, steroidogenesis and subsequent developmental competence of the oocyte-granulosa cell complex derived from bovine early antral follicles. Reprod. Biol. Endocrinol. 2016, 14, 3. [Google Scholar] [CrossRef] [Green Version]
- Cavalera, F.; Zanoni, M.; Merico, V.; Sacchi, L.; Bellazzi, R.; Garagna, S.; Zuccotti, M. Chromatin organization and timing of polar body I extrusion identify developmentally competent mouse oocytes. Int. J. Dev. Biol. 2019, 63, 245–251. [Google Scholar] [CrossRef]
- Holubcova, Z.; Kyjovska, D.; Martonova, M.; Paralova, D.; Klenkova, T.; Otevrel, P.; Stepanova, R.; Kloudova, S.; Hampl, A. Egg maturity assessment prior to ICSI prevents premature fertilization of late-maturing oocytes. J. Assist. Reprod. Genet. 2019, 36, 445–452. [Google Scholar] [CrossRef] [Green Version]
- Nandi, S.; Ravindranatha, B.M.; Gupta, P.S.; Sarma, P.V. Timing of sequential changes in cumulus cells and first polar body extrusion during in vitro maturation of buffalo oocytes. Theriogenology 2002, 57, 1151–1159. [Google Scholar] [CrossRef]
- Van der Westerlaken, L.A.; van der Schans, A.; Eyestone, W.H.; de Boer, H.A. Kinetics of first polar body extrusion and the effect of time of stripping of the cumulus and time of insemination on developmental competence of bovine oocytes. Theriogenology 1994, 42, 361–370. [Google Scholar] [CrossRef]
- Park, Y.S.; Kim, S.S.; Kim, J.M.; Park, H.D.; Byun, M.D. The effects of duration of in vitro maturation of bovine oocytes on subsequent development, quality and transfer of embryos. Theriogenology 2005, 64, 123–134. [Google Scholar] [CrossRef] [PubMed]
- Ward, F.; Enright, B.; Rizos, D.; Boland, M.; Lonergan, P. Optimization of in vitro bovine embryo production: Effect of duration of maturation, length of gamete co-incubation, sperm concentration and sire. Theriogenology 2002, 57, 2105–2117. [Google Scholar] [CrossRef]
- Dominko, T.; First, N.L. Timing of meiotic progression in bovine oocytes and its effect on early embryo development. Mol. Reprod. Dev. 1997, 47, 456–467. [Google Scholar] [CrossRef]
- Hu, J.; Jin, C.; Zheng, H.; Liu, Q.; Zhu, W.; Zeng, Z.; Wu, J.; Wang, Y.; Li, J.; Zhang, X.; et al. First polar body morphology affects potential development of porcine parthenogenetic embryo in vitro. Zygote 2015, 23, 615–621. [Google Scholar] [CrossRef]
- Ebner, T.; Moser, M.; Sommergruber, M.; Yaman, C.; Pfleger, U.; Tews, G. First polar body morphology and blastocyst formation rate in ICSI patients. Hum. Reprod. 2002, 17, 2415–2418. [Google Scholar] [CrossRef] [Green Version]
- Zhou, W.; Fu, L.; Sha, W.; Chu, D.; Li, Y. Relationship of polar bodies morphology to embryo quality and pregnancy outcome. Zygote 2016, 24, 401–407. [Google Scholar] [CrossRef]
- Ebner, T.; Yaman, C.; Moser, M.; Sommergruber, M.; Feichtinger, O.; Tews, G. Prognostic value of first polar body morphology on fertilization rate and embryo quality in intracytoplasmic sperm injection. Hum. Reprod. 2000, 15, 427–430. [Google Scholar] [CrossRef]
- Rose, B.I.; Laky, D. Polar body fragmentation in IVM oocytes is associated with impaired fertilization and embryo development. J. Assist. Reprod. Genet. 2013, 30, 679–682. [Google Scholar] [CrossRef] [Green Version]
- Halvaei, I.; Khalili, M.A.; Soleimani, M.; Razi, M.H. Evaluating the Role of First Polar Body Morphology on Rates of Fertilization and Embryo Development in ICSI Cycles. Int. J. Fertil. Steril. 2011, 5, 110–115. [Google Scholar] [PubMed]
- De Santis, L.; Cino, I.; Rabellotti, E.; Calzi, F.; Persico, P.; Borini, A.; Coticchio, G. Polar body morphology and spindle imaging as predictors of oocyte quality. Reprod. Biomed. Online 2005, 11, 36–42. [Google Scholar] [CrossRef]
- Ciotti, P.M.; Notarangelo, L.; Morselli-Labate, A.M.; Felletti, V.; Porcu, E.; Venturoli, S. First polar body morphology before ICSI is not related to embryo quality or pregnancy rate. Hum. Reprod. 2004, 19, 2334–2339. [Google Scholar] [CrossRef] [PubMed]
- Verlinsky, Y.; Lerner, S.; Illkevitch, N.; Kuznetsov, V.; Kuznetsov, I.; Cieslak, J.; Kuliev, A. Is there any predictive value of first polar body morphology for embryo genotype or developmental potential? Reprod. Biomed. Online 2003, 7, 336–341. [Google Scholar] [CrossRef]
- Caamano, J.N.; Munoz, M.; Diez, C.; Gomez, E. Polarized light microscopy in mammalian oocytes. Reprod. Domest. Anim. 2010, 45 (Suppl. 2), 49–56. [Google Scholar] [CrossRef] [PubMed]
- Montag, M.; Koster, M.; van der Ven, K.; van der Ven, H. Gamete competence assessment by polarizing optics in assisted reproduction. Hum. Reprod. Update 2011, 17, 654–666. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Yan, J.; Liu, Z.C.; Yan, L.Y.; Li, M.; Zhou, Q.; Qiao, J. Optimal timing of oocyte maturation and its relationship with the spindle assembly and developmental competence of in vitro matured human oocytes. Fertil. Steril. 2011, 96, 73–78. [Google Scholar] [CrossRef]
- Kilani, S.; Cooke, S.; Tilia, L.; Chapman, M. Does meiotic spindle normality predict improved blastocyst development, implantation and live birth rates? Fertil. Steril. 2011, 96, 389–393. [Google Scholar] [CrossRef]
- Tomari, H.; Honjo, K.; Kunitake, K.; Aramaki, N.; Kuhara, S.; Hidaka, N.; Nishimura, K.; Nagata, Y.; Horiuchi, T. Meiotic spindle size is a strong indicator of human oocyte quality. Reprod. Med. Biol. 2018, 17, 268–274. [Google Scholar] [CrossRef]
- Rienzi, L.; Ubaldi, F.; Martinez, F.; Iacobelli, M.; Minasi, M.G.; Ferrero, S.; Tesarik, J.; Greco, E. Relationship between meiotic spindle location with regard to the polar body position and oocyte developmental potential after ICSI. Hum. Reprod. 2003, 18, 1289–1293. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Oldenbourg, R.; Trimarchi, J.R.; Keefe, D.L. A reliable, noninvasive technique for spindle imaging and enucleation of mammalian oocytes. Nat. Biotechnol. 2000, 18, 223–225. [Google Scholar] [CrossRef] [PubMed]
- Lu, F.; Shi, D.; Wei, J.; Yang, S.; Wei, Y. Development of embryos reconstructed by interspecies nuclear transfer of adult fibroblasts between buffalo (Bubalus bubalis) and cattle (Bos indicus). Theriogenology 2005, 64, 1309–1319. [Google Scholar] [CrossRef] [PubMed]
- Caamano, J.N.; Maside, C.; Gil, M.A.; Munoz, M.; Cuello, C.; Diez, C.; Sanchez-Osorio, J.R.; Martin, D.; Gomis, J.; Vazquez, J.M.; et al. Use of polarized light microscopy in porcine reproductive technologies. Theriogenology 2011, 76, 669–677. [Google Scholar] [CrossRef] [PubMed]
- Rienzi, L.; Martinez, F.; Ubaldi, F.; Minasi, M.G.; Iacobelli, M.; Tesarik, J.; Greco, E. Polscope analysis of meiotic spindle changes in living metaphase II human oocytes during the freezing and thawing procedures. Hum. Reprod. 2004, 19, 655–659. [Google Scholar] [CrossRef] [Green Version]
- Caamano, J.N.; Diez, C.; Trigal, B.; Munoz, M.; Morato, R.; Martin, D.; Carrocera, S.; Mogas, T.; Gomez, E. Assessment of meiotic spindle configuration and post-warming bovine oocyte viability using polarized light microscopy. Reprod. Domest Anim. 2013, 48, 470–476. [Google Scholar] [CrossRef]
- Montag, M.; Schimming, T.; Koster, M.; Zhou, C.; Dorn, C.; Rosing, B.; van der Ven, H.; Ven der Ven, K. Oocyte zona birefringence intensity is associated with embryonic implantation potential in ICSI cycles. Reprod. Biomed. Online 2008, 16, 239–244. [Google Scholar] [CrossRef]
- Madaschi, C.; de Souza Bonetti, T.C.; de Almeida Ferreira Braga, D.P.; Pasqualotto, F.F.; Iaconelli, A., Jr.; Borges, E., Jr. Spindle imaging: A marker for embryo development and implantation. Fertil. Steril. 2008, 90, 194–198. [Google Scholar] [CrossRef]
- De Almeida Ferreira Braga, D.P.; de Cassia Savio Figueira, R.; Queiroz, P.; Madaschi, C.; Iaconelli, A., Jr.; Borges, E., Jr. Zona pellucida birefringence in in vivo and in vitro matured oocytes. Fertil. Steril. 2010, 94, 2050–2053. [Google Scholar] [CrossRef]
- Petersen, C.G.; Vagnini, L.D.; Mauri, A.L.; Massaro, F.C.; Silva, L.F.; Cavagna, M.; Baruffi, R.L.; Oliveira, J.B.; Franco, J.G., Jr. Evaluation of zona pellucida birefringence intensity during in vitro maturation of oocytes from stimulated cycles. Reprod. Biol. Endocrinol. 2011, 9, 53. [Google Scholar] [CrossRef] [Green Version]
- Ashourzadeh, S.; Khalili, M.A.; Omidi, M.; Mahani, S.N.; Kalantar, S.M.; Aflatoonian, A.; Habibzadeh, V. Noninvasive assays of in vitro matured human oocytes showed insignificant correlation with fertilization and embryo development. Arch. Gynecol. Obstet. 2015, 292, 459–463. [Google Scholar] [CrossRef]
- Held, E.; Mertens, E.M.; Mohammadi-Sangcheshmeh, A.; Salilew-Wondim, D.; Besenfelder, U.; Havlicek, V.; Herrler, A.; Tesfaye, D.; Schellander, K.; Holker, M. Zona pellucida birefringence correlates with developmental capacity of bovine oocytes classified by maturational environment, COC morphology and G6PDH activity. Reprod. Fertil. Dev. 2012, 24, 568–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koester, M.; Mohammadi-Sangcheshmeh, A.; Montag, M.; Rings, F.; Schimming, T.; Tesfaye, D.; Schellander, K.; Hoelker, M. Evaluation of bovine zona pellucida characteristics in polarized light as a prognostic marker for embryonic developmental potential. Reproduction 2011, 141, 779–787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertero, A.; Ritrovato, F.; Evangelista, F.; Stabile, V.; Fortina, R.; Ricci, A.; Revelli, A.; Vincenti, L.; Nervo, T. Evaluation of equine oocyte developmental competence using polarized light microscopy. Reproduction 2017, 153, 775–784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohammadi-Sangcheshmeh, A.; Held, E.; Rings, F.; Ghanem, N.; Salilew-Wondim, D.; Tesfaye, D.; Sieme, H.; Schellander, K.; Hoelker, M. Developmental competence of equine oocytes: Impacts of zona pellucida birefringence and maternally derived transcript expression. Reprod. Fertil. Dev. 2014, 26, 441–452. [Google Scholar] [CrossRef]
- Pujol, M.; Lopez-Bejar, M.; Paramio, M.T. Developmental competence of heifer oocytes selected using the brilliant cresyl blue (BCB) test. Theriogenology 2004, 61, 735–744. [Google Scholar] [CrossRef]
- Alm, H.; Torner, H.; Lohrke, B.; Viergutz, T.; Ghoneim, I.M.; Kanitz, W. Bovine blastocyst development rate in vitro is influenced by selection of oocytes by brillant cresyl blue staining before IVM as indicator for glucose-6-phosphate dehydrogenase activity. Theriogenology 2005, 63, 2194–2205. [Google Scholar] [CrossRef]
- Bhojwani, S.; Alm, H.; Torner, H.; Kanitz, W.; Poehland, R. Selection of developmentally competent oocytes through brilliant cresyl blue stain enhances blastocyst development rate after bovine nuclear transfer. Theriogenology 2007, 67, 341–345. [Google Scholar] [CrossRef]
- Opiela, J.; Katska-Ksiazkiewicz, L. The utility of Brilliant Cresyl Blue (BCB) staining of mammalian oocytes used for in vitro embryo production (IVP). Reprod. Biol. 2013, 13, 177–183. [Google Scholar] [CrossRef]
- Silva, D.S.; Rodriguez, P.; Galuppo, A.; Arruda, N.S.; Rodrigues, J.L. Selection of bovine oocytes by brilliant cresyl blue staining: Effect on meiosis progression, organelle distribution and embryo development. Zygote 2013, 21, 250–255. [Google Scholar] [CrossRef]
- Fakruzzaman, M.; Bang, J.I.; Lee, K.L.; Kim, S.S.; Ha, A.N.; Ghanem, N.; Han, C.H.; Cho, K.W.; White, K.L.; Kong, I.K. Mitochondrial content and gene expression profiles in oocyte-derived embryos of cattle selected on the basis of brilliant cresyl blue staining. Anim. Reprod. Sci. 2013, 142, 19–27. [Google Scholar] [CrossRef]
- Castaneda, C.A.; Kaye, P.; Pantaleon, M.; Phillips, N.; Norman, S.; Fry, R.; D’Occhio, M.J. Lipid content, active mitochondria and brilliant cresyl blue staining in bovine oocytes. Theriogenology 2013, 79, 417–422. [Google Scholar] [CrossRef] [PubMed]
- Salviano, M.B.; Collares, F.J.; Becker, B.S.; Rodrigues, B.A.; Rodrigues, J.L. Bovine non-competent oocytes (BCB-) negatively impact the capacity of competent (BCB+) oocytes to undergo in vitro maturation, fertilisation and embryonic development. Zygote 2016, 24, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Karami Shabankareh, H.; Azimi, G.; Torki, M. Developmental competence of bovine oocytes selected based on follicle size and using the brilliant cresyl blue (BCB) test. Iran. J. Reprod. Med. 2014, 12, 771–778. [Google Scholar] [PubMed]
- Mirshamsi, S.M.; Karamishabankareh, H.; Ahmadi-Hamedani, M.; Soltani, L.; Hajarian, H.; Abdolmohammadi, A.R. Combination of oocyte and zygote selection by brilliant cresyl blue (BCB) test enhanced prediction of developmental potential to the blastocyst in cattle. Anim. Reprod. Sci. 2013, 136, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Anguita, B.; Vandaele, L.; Mateusen, B.; Maes, D.; Van Soom, A. Developmental competence of bovine oocytes is not related to apoptosis incidence in oocytes, cumulus cells and blastocysts. Theriogenology 2007, 67, 537–549. [Google Scholar] [CrossRef]
- Ikeda, S.; Imai, H.; Yamada, M. Apoptosis in cumulus cells during in vitro maturation of bovine cumulus-enclosed oocytes. Reproduction 2003, 125, 369–376. [Google Scholar] [CrossRef]
- Janowski, D.; Salilew-Wondim, D.; Torner, H.; Tesfaye, D.; Ghanem, N.; Tomek, W.; El-Sayed, A.; Schellander, K.; Holker, M. Incidence of apoptosis and transcript abundance in bovine follicular cells is associated with the quality of the enclosed oocyte. Theriogenology 2012, 78, 656–669. [Google Scholar] [CrossRef]
- Warzych, E.; Pers-Kamczyc, E.; Krzywak, A.; Dudzinska, S.; Lechniak, D. Apoptotic index within cumulus cells is a questionable marker of meiotic competence of bovine oocytes matured in vitro. Reprod. Biol. 2013, 13, 82–87. [Google Scholar] [CrossRef]
- Lee, K.S.; Joo, B.S.; Na, Y.J.; Yoon, M.S.; Choi, O.H.; Kim, W.W. Cumulus cells apoptosis as an indicator to predict the quality of oocytes and the outcome of IVF-ET. J. Assist. Reprod. Genet. 2001, 18, 490–498. [Google Scholar] [CrossRef]
- Fan, Y.; Chang, Y.; Wei, L.; Chen, J.; Li, J.; Goldsmith, S.; Silber, S.; Liang, X. Apoptosis of mural granulosa cells is increased in women with diminished ovarian reserve. J. Assist. Reprod. Genet. 2019, 36, 1225–1235. [Google Scholar] [CrossRef] [Green Version]
- Clavero, A.; Castilla, J.A.; Nunez, A.I.; Garcia-Pena, M.L.; Maldonado, V.; Fontes, J.; Mendoza, N.; Martinez, L. Apoptosis in human granulosa cells after induction of ovulation in women participating in an intracytoplasmic sperm injection program. Eur. J. Obstet. Gynecol. Reprod. Biol. 2003, 110, 181–185. [Google Scholar] [CrossRef]
- Bunel, A.; Jorssen, E.P.; Merckx, E.; Leroy, J.L.; Bols, P.E.; Sirard, M.A. Individual bovine in vitro embryo production and cumulus cell transcriptomic analysis to distinguish cumulus-oocyte complexes with high or low developmental potential. Theriogenology 2015, 83, 228–237. [Google Scholar] [CrossRef] [PubMed]
- Donnison, M.; Pfeffer, P.L. Isolation of genes associated with developmentally competent bovine oocytes and quantitation of their levels during development. Biol. Reprod. 2004, 71, 1813–1821. [Google Scholar] [CrossRef]
- Bettegowda, A.; Patel, O.V.; Lee, K.B.; Park, K.E.; Salem, M.; Yao, J.; Ireland, J.J.; Smith, G.W. Identification of novel bovine cumulus cell molecular markers predictive of oocyte competence: Functional and diagnostic implications. Biol. Reprod. 2008, 79, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Mourot, M.; Dufort, I.; Gravel, C.; Algriany, O.; Dieleman, S.; Sirard, M.A. The influence of follicle size, FSH-enriched maturation medium, and early cleavage on bovine oocyte maternal mRNA levels. Mol. Reprod. Dev. 2006, 73, 1367–1379. [Google Scholar] [CrossRef] [PubMed]
- Ashry, M.; Lee, K.; Mondal, M.; Datta, T.K.; Folger, J.K.; Rajput, S.K.; Zhang, K.; Hemeida, N.A.; Smith, G.W. Expression of TGFbeta superfamily components and other markers of oocyte quality in oocytes selected by brilliant cresyl blue staining: Relevance to early embryonic development. Mol. Reprod. Dev. 2015, 82, 251–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nivet, A.L.; Vigneault, C.; Blondin, P.; Sirard, M.A. Changes in granulosa cells’ gene expression associated with increased oocyte competence in bovine. Reproduction 2013, 145, 555–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melo, E.O.; Cordeiro, D.M.; Pellegrino, R.; Wei, Z.; Daye, Z.J.; Nishimura, R.C.; Dode, M.A. Identification of molecular markers for oocyte competence in bovine cumulus cells. Anim. Genet. 2017, 48, 19–29. [Google Scholar] [CrossRef] [Green Version]
- Assidi, M.; Dufort, I.; Ali, A.; Hamel, M.; Algriany, O.; Dielemann, S.; Sirard, M.A. Identification of potential markers of oocyte competence expressed in bovine cumulus cells matured with follicle-stimulating hormone and/or phorbol myristate acetate in vitro. Biol. Reprod. 2008, 79, 209–222. [Google Scholar] [CrossRef] [Green Version]
- Kussano, N.R.; Leme, L.O.; Guimaraes, A.L.; Franco, M.M.; Dode, M.A. Molecular markers for oocyte competence in bovine cumulus cells. Theriogenology 2016, 85, 1167–1176. [Google Scholar] [CrossRef]
- Khurchabilig, A.; Sato, A.; Ashibe, S.; Hara, A.; Fukumori, R.; Nagao, Y. Expression levels of FSHR, IGF1R, CYP11al and HSD3beta in cumulus cells can predict in vitro developmental competence of bovine oocytes. Zygote 2020, 1–7. [Google Scholar] [CrossRef]
- O’Shea, L.C.; Daly, E.; Hensey, C.; Fair, T. ATRX is a novel progesterone-regulated protein and biomarker of low developmental potential in mammalian oocytes. Reproduction 2017, 153, 671–682. [Google Scholar] [CrossRef] [PubMed]
- Xiong, X.R.; Lan, D.L.; Li, J.; Yin, S.; Xiong, Y.; Zi, X.D. Identification of differential abundances of mRNA transcript in cumulus cells and CCND1 associated with yak oocyte developmental competence. Anim. Reprod. Sci. 2019, 208, 106135. [Google Scholar] [CrossRef] [PubMed]
- Bunel, A.; Nivet, A.L.; Blondin, P.; Vigneault, C.; Richard, F.J.; Sirard, M.A. Cumulus cell gene expression associated with pre-ovulatory acquisition of developmental competence in bovine oocytes. Reprod. Fertil. Dev. 2014, 26, 855–865. [Google Scholar] [CrossRef] [PubMed]
- Dieci, C.; Lodde, V.; Labreque, R.; Dufort, I.; Tessaro, I.; Sirard, M.A.; Luciano, A.M. Differences in cumulus cell gene expression indicate the benefit of a pre-maturation step to improve in-vitro bovine embryo production. Mol. Hum. Reprod. 2016, 22, 882–897. [Google Scholar] [CrossRef] [PubMed]
- Walter, J.; Monthoux, C.; Fortes, C.; Grossmann, J.; Roschitzki, B.; Meili, T.; Riond, B.; Hofmann-Lehmann, R.; Naegeli, H.; Bleul, U. The bovine cumulus proteome is influenced by maturation condition and maturational competence of the oocyte. Sci. Rep. 2020, 10, 9880. [Google Scholar] [CrossRef]
- Peddinti, D.; Memili, E.; Burgess, S.C. Proteomics-based systems biology modeling of bovine germinal vesicle stage oocyte and cumulus cell interaction. PLoS ONE 2010, 5, e11240. [Google Scholar] [CrossRef] [Green Version]
- Memili, E.; Peddinti, D.; Shack, L.A.; Nanduri, B.; McCarthy, F.; Sagirkaya, H.; Burgess, S.C. Bovine germinal vesicle oocyte and cumulus cell proteomics. Reproduction 2007, 133, 1107–1120. [Google Scholar] [CrossRef] [Green Version]
- Sutton, M.L.; Gilchrist, R.B.; Thompson, J.G. Effects of in-vivo and in-vitro environments on the metabolism of the cumulus-oocyte complex and its influence on oocyte developmental capacity. Hum. Reprod. Update 2003, 9, 35–48. [Google Scholar] [CrossRef]
- Revelli, A.; Delle Piane, L.; Casano, S.; Molinari, E.; Massobrio, M.; Rinaudo, P. Follicular fluid content and oocyte quality: From single biochemical markers to metabolomics. Reprod. Biol. Endocrinol. 2009, 7, 40. [Google Scholar] [CrossRef] [Green Version]
- Wrenzycki, C.; Stinshoff, H. Maturation environment and impact on subsequent developmental competence of bovine oocytes. Reprod. Domest. Anim. 2013, 48 (Suppl. 1), 38–43. [Google Scholar] [CrossRef] [PubMed]
- Matoba, S.; Bender, K.; Fahey, A.G.; Mamo, S.; Brennan, L.; Lonergan, P.; Fair, T. Predictive value of bovine follicular components as markers of oocyte developmental potential. Reprod. Fertil. Dev. 2014, 26, 337–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bender, K.; Walsh, S.; Evans, A.C.; Fair, T.; Brennan, L. Metabolite concentrations in follicular fluid may explain differences in fertility between heifers and lactating cows. Reproduction 2010, 139, 1047–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zachut, M.; Sood, P.; Levin, Y.; Moallem, U. Proteomic analysis of preovulatory follicular fluid reveals differentially abundant proteins in less fertile dairy cows. J. Proteomics 2016, 139, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Alves, B.G.; Alves, K.A.; Lucio, A.C.; Martins, M.C.; Silva, T.H.; Alves, B.G.; Braga, L.S.; Silva, T.V.; Viu, M.A.; Beletti, M.E.; et al. Ovarian activity and oocyte quality associated with the biochemical profile of serum and follicular fluid from Girolando dairy cows postpartum. Anim. Reprod. Sci. 2014, 146, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, C.; Fujito, A.; Kazuka, M.; Sugiyama, R.; Ito, H.; Isaka, K. Anti-Mullerian hormone substance from follicular fluid is positively associated with success in oocyte fertilization during in vitro fertilization. Fertil. Steril. 2008, 89, 586–591. [Google Scholar] [CrossRef]
- Kim, J.H.; Lee, J.R.; Chang, H.J.; Jee, B.C.; Suh, C.S.; Kim, S.H. Anti-Mullerian hormone levels in the follicular fluid of the preovulatory follicle: A predictor for oocyte fertilization and quality of embryo. J. Korean Med. Sci. 2014, 29, 1266–1270. [Google Scholar] [CrossRef] [Green Version]
- Tramisak Milakovic, T.; Panic Horvat, L.; Cavlovic, K.; Smiljan Severinski, N.; Vlasic, H.; Vlastelic, I.; Ljiljak, D.; Radojcic Badovinac, A. Follicular fluid anti-Mullerian hormone: A predictive marker of fertilization capacity of MII oocytes. Arch. Gynecol. Obstet. 2015, 291, 681–687. [Google Scholar] [CrossRef]
- Revelli, A.; Canosa, S.; Bergandi, L.; Skorokhod, O.A.; Biasoni, V.; Carosso, A.; Bertagna, A.; Maule, M.; Aldieri, E.; D’Eufemia, M.D.; et al. Oocyte polarized light microscopy, assay of specific follicular fluid metabolites, and gene expression in cumulus cells as different approaches to predict fertilization efficiency after ICSI. Reprod. Biol. Endocrinol. 2017, 15, 47. [Google Scholar] [CrossRef]
- Ciepiela, P.; Duleba, A.J.; Kario, A.; Chelstowski, K.; Branecka-Wozniak, D.; Kurzawa, R. Oocyte matched follicular fluid anti-Mullerian hormone is an excellent predictor of live birth after fresh single embryo transfer. Hum. Reprod. 2019, 34, 2244–2253. [Google Scholar] [CrossRef]
- Rico, C.; Fabre, S.; Medigue, C.; di Clemente, N.; Clement, F.; Bontoux, M.; Touze, J.L.; Dupont, M.; Briant, E.; Remy, B.; et al. Anti-mullerian hormone is an endocrine marker of ovarian gonadotropin-responsive follicles and can help to predict superovulatory responses in the cow. Biol. Reprod. 2009, 80, 50–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rico, C.; Drouilhet, L.; Salvetti, P.; Dalbies-Tran, R.; Jarrier, P.; Touze, J.L.; Pillet, E.; Ponsart, C.; Fabre, S.; Monniaux, D. Determination of anti-Mullerian hormone concentrations in blood as a tool to select Holstein donor cows for embryo production: From the laboratory to the farm. Reprod. Fertil. Dev. 2012, 24, 932–944. [Google Scholar] [CrossRef]
- Monniaux, D.; Barbey, S.; Rico, C.; Fabre, S.; Gallard, Y.; Larroque, H. Anti-Mullerian hormone: A predictive marker of embryo production in cattle? Reprod. Fertil. Dev. 2010, 22, 1083–1091. [Google Scholar] [CrossRef] [PubMed]
- Gebremedhn, S.; Salilew-Wondim, D.; Ahmad, I.; Sahadevan, S.; Hossain, M.M.; Hoelker, M.; Rings, F.; Neuhoff, C.; Tholen, E.; Looft, C.; et al. MicroRNA Expression Profile in Bovine Granulosa Cells of Preovulatory Dominant and Subordinate Follicles during the Late Follicular Phase of the Estrous Cycle. PLoS ONE 2015, 10, e0125912. [Google Scholar] [CrossRef] [PubMed]
- Sohel, M.M.; Hoelker, M.; Noferesti, S.S.; Salilew-Wondim, D.; Tholen, E.; Looft, C.; Rings, F.; Uddin, M.J.; Spencer, T.E.; Schellander, K.; et al. Exosomal and Non-Exosomal Transport of Extra-Cellular microRNAs in Follicular Fluid: Implications for Bovine Oocyte Developmental Competence. PLoS ONE 2013, 8, e78505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasquariello, R.; Manzoni, E.F.M.; Fiandanese, N.; Viglino, A.; Pocar, P.; Brevini, T.A.L.; Williams, J.L.; Gandolfi, F. Implications of miRNA expression pattern in bovine oocytes and follicular fluids for developmental competence. Theriogenology 2020, 145, 77–85. [Google Scholar] [CrossRef] [PubMed]



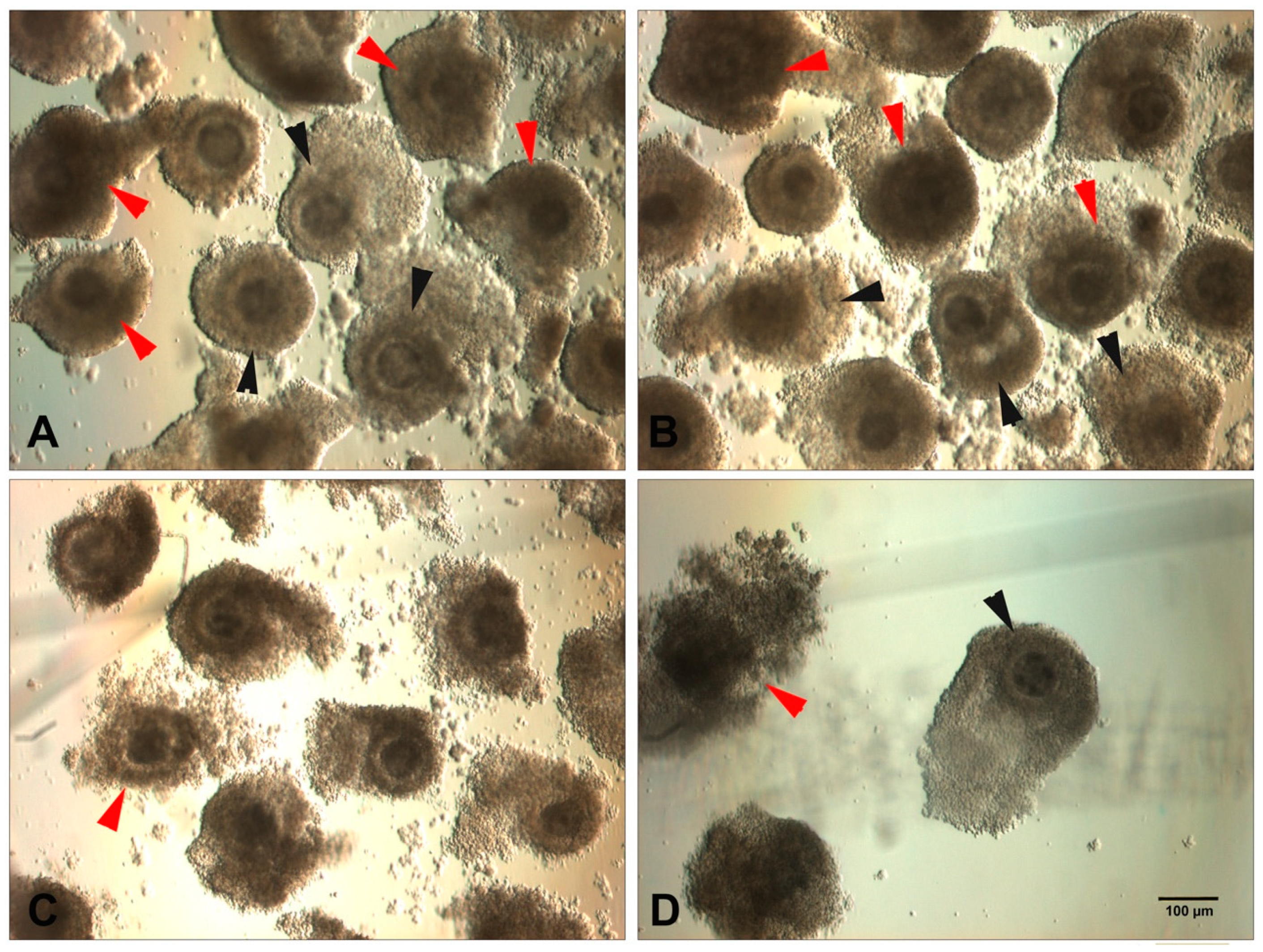{kind=link}
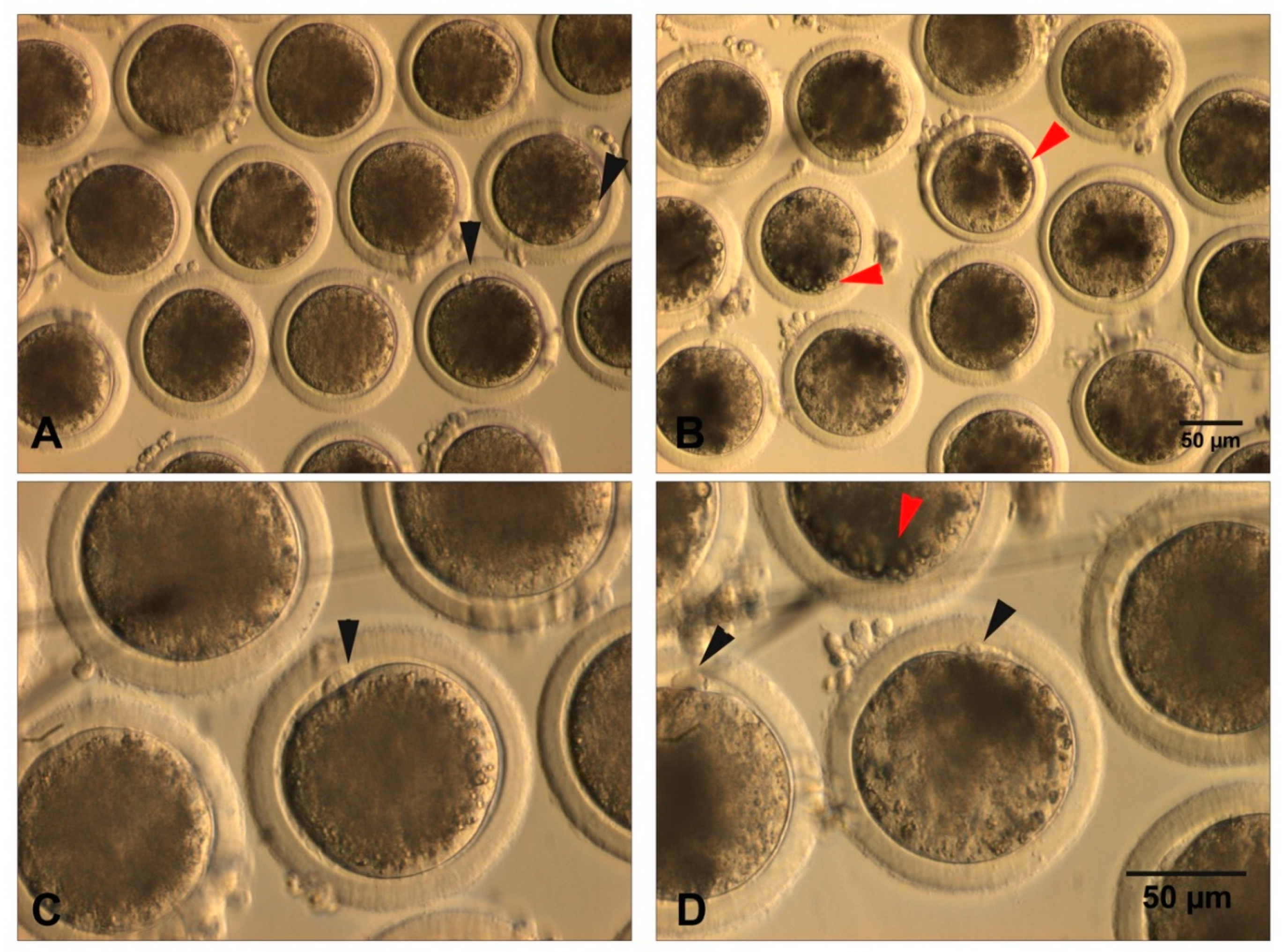{kind=link}
| Reference | Criteria | Recommendation |
|---|---|---|
| [28,30,31] | Ovarian morphology | Presence of cycle-related structures |
| [7,33,34,35] | Follicle size | >5 mm |
| [30,49,50] | Morphology of the cumulus–oocyte complexes (COCs) | COCs with at least five layers of cumulus cells (CC), compact and/or slightly expanded cumulus, with or without dark spots in the oocyte and cumulus |
| [53,67,68] | Lipid content | Dark ooplasm indicates high competence, light-colored indicates lacking lipids and poor competence, and black ooplasm indicates aging |
| [77,78,79,80] | Cumulus expansion and oocyte size | Not associated to oocyte quality; important role in fertilization |
| [29,83,86,135] | Oocyte size | Diameters >115 and <130 microns |
| [96,97,98,99] | First polar body (PB1) morphology | PB1 of a homogeneous, round shape with a smooth or intact surface |
| [112,114,115] | Meiotic spindle and zona pellucida birefringence | Useful tool for micromanipulation procedures (intracytoplasmic sperm injection (ICSI) or somatic cell nuclear transfer (SCNT)) and for assessing post-warming integrity of meiotic spindle of vitrified bovine oocytes |
| [121,122] | Zona pellucida birefringence (ZPB) | Lower ZPB is related to high quality oocytes and improved blastocyst development |
| [115,128,129,130,134] | Brilliant cresyl blue staining | BCB+ oocytes have higher developmental competence than BCB− oocytes |
| Reference | Oocyte Stage | Criterion of Developmental Competence | Technique Used | Genes and/or Pathways Associated with High Competence | Genes and/or Pathways Associated with Low Competence |
|---|---|---|---|---|---|
| Transcriptomic | |||||
| [16] | GV | GC collected 2 h before and 6 h after LH surge | qPCR and microarray analysis | TNFAIP6, HAS2, HSD3B2, PLOD2, CHSY1 (differentiation, cell growth, protein translation, apoptosis-related, lipid and glucose metabolism, ECM formation) | ENO1, DNAJB6, GJA1, SYNPO, ZNF330, MYO1D (protein synthesis cellular movement, cell signaling, molecular transport, nucleic acid metabolism) |
| [35] | GV | follicle size (1.0–3.0, 3.1–6.0, 6.1–8.0, and ≥8.1 mm) | qPCR | FSHR (follicle stimulant hormone receptor), GH (cell growth), and EGF (cell growth and differentiation) | N.A |
| [142] | GV | Cell arrest and oocyte fate | qPCR and microarray analysis | GATM (post-translational modification, amino-acid metabolism, and free-radical scavenging) | AGPAT9 (lipid metabolism), CLIC3 (chloride ion concentration control, cell volume regulation, and apoptosis), KRT8 (cellular assembly and organization, apoptosis) |
| [143] | GV | Follicle size (>5 mm vs. <2mm) | qPCR and SSH | Oct4, Msx1 (transcription factors), Znf198 (TFGb and activin signaling), NDFIP1(posttranslational modification), CCNA2 (cell cycle), SLB (stabilization and translation of mRNAs encoding histones) | N.A |
| [144] | GV | Adult vs. prepuberal donors | qPCR and microarray analysis | N.A | CTSB, CTSK, CTSS, and CTSZ (cathepsin family of lysosomal cysteine proteinases) |
| [145] | GV | OPU 6 h post LH vs. slaughterhouse oocytes after 6 h IVM | qPCR and microarray analysis | PTTG1, CDC5L, CKS1B, CCNB2 (cell cycle), PSMB2, PRDX1 (cell metabolism), RGS16 (cell signaling), SKIIP (gene expression), and chromatin support H2A | BMP15, GDF9, CCNB1, and STK6 (follicle–oocyte interaction and cell cycle) |
| [146] | GV | Brilliant cresyl blue staining | qPCR | N.A | CTSB, CTSK, CTSS, and CTSZ (cathepsin family of lysosomal cysteine proteinases) |
| [147] | GV | GC collected after FSH withdrawal | qPCR and microarray analysis | SMAD7, STAT1 (transcription), PRL and GH (cell growth, proliferation), BMPR1B, IGF2, RELN, and TFPI2 (follicle growth), NRP1 (angiogenesis), GFPT2, TF, and VNN1 (oxidative stress response) | KCNJ8 and ANKRD1 (apoptosis and inflammation) |
| [148] | GV | Follicle size (>8 mm vs. <3mm) | qPCR and microarray analysis | FGF11 (cell growth, and differentiation), IGFBP4 and SPRY1 (cell cycle, DNA repair) | ARHGAP22, COL18A1, and GPC4 (cell cycle, signaling) |
| [149] | GV | IVM plus FSH or phorbol myristate acetate (PMA) treatment | qPCR and microarray analysis | HAS2, INHBA, EGFR, GREM1, CD44, TNFAIP6, PTGS2, HSP90B1, SERPINE2, PTX3 (differentiation, cell growth, protein translation, apoptosis, lipid and glucose metabolism, ECM formation) | N.A |
| [150] | GV | Follicle size and oocyte fate | qPCR | GPC4 (regulation of growth factors, adhesion, signaling, proliferation, and differentiation) | N.A |
| [151] | GV | COCs morphology and oocyte fate | qPCR | N.A | FSHR, IGF1R, CYP11al, and HSD3β (cell growth, cell differentiation, steroidogenesis) |
| [153] | GV | Maturation outcome and oocyte fate | RNA-seq | CCND1, BMP15, GDF9, H19, KLF4, GPC1, SYCP3, and CTSB (cell cycle, meiosis, cell signaling, metabolism, and apoptosis) | N.A |
| [154] | GV | FSH withdrawals; follicles from 5 mm aspirated by OPU | qPCR and microarray analysis | CYP11A1 (steroidogenesis), NSDHL (cholesterol synthesis), GATM (creatine biosynthesis), MAN1A1 (functional gap junction-mediated communication), VNN1 (oxidative stress response), NRP1 (angiogenesis), TGFB1 (cell growth and differentiation) | N.A |
| [155] | GV | Chromatin compaction, follicle size, and BCB staining | qPCR and microarray analysis | GATM (posttranslational modification, amino-acid metabolism, and free-radical scavenging), MAN1A1 (functional gap junction-mediated COC communication), ZIP8 (zinc transporter) | ANGPT2 (cell death, apoptosis) |
| Proteomic | |||||
| [156] | MII | Matured in vivo vs. IVM | MALDI TOF | KEGG pathways of the complement and coagulation cascade, ECM–receptor interactions, steroid biosynthesis, glucose and carbohydrate metabolism | N.A |
| [157] | GV | COC morphology and follicle size (>2 mm to 8 mm) | 2-DLCMS | Integrin signaling, actin cytoskeleton signaling, ephrin receptor signaling, PI3K signaling, MAPK signaling | N.A |
| [158] | GV | COC morphology and follicle size (>2 mm to 8 mm) | 2-DLCMS | 4395 proteins were expressed in the CCs; 858 proteins were common to both CCs and oocytes | N.A |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aguila, L.; Treulen, F.; Therrien, J.; Felmer, R.; Valdivia, M.; Smith, L.C. Oocyte Selection for In Vitro Embryo Production in Bovine Species: Noninvasive Approaches for New Challenges of Oocyte Competence. Animals 2020, 10, 2196. https://doi.org/10.3390/ani10122196
Aguila L, Treulen F, Therrien J, Felmer R, Valdivia M, Smith LC. Oocyte Selection for In Vitro Embryo Production in Bovine Species: Noninvasive Approaches for New Challenges of Oocyte Competence. Animals. 2020; 10(12):2196. https://doi.org/10.3390/ani10122196
Chicago/Turabian StyleAguila, Luis, Favian Treulen, Jacinthe Therrien, Ricardo Felmer, Martha Valdivia, and Lawrence C Smith. 2020. "Oocyte Selection for In Vitro Embryo Production in Bovine Species: Noninvasive Approaches for New Challenges of Oocyte Competence" Animals 10, no. 12: 2196. https://doi.org/10.3390/ani10122196