
A Peculiar Distribution of the Emerging Nematode Angiostrongylus cantonensis in the Canary Islands (Spain): Recent Introduction or Isolation Effect?



 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, Q.P.; Lai, D.H.; Zhu, X.Q.; Chen, X.G.; Lun, Z.R. Human angiostrongyliasis. Lancet Infect. Dis. 2008, 8, 621–630. [Google Scholar] [CrossRef]
- Chen, H.T. Un nouveau nematode pulmonaire, Pulmonema cantonensisn.g., n. sp. des rats de Canton. Ann. Parasitol. Hum. Comp. 1935, 13, 312–317. [Google Scholar] [CrossRef] [Green Version]
- Foronda, P.; López-Gonzalez, M.; Miquel, J.; Torres, J.; Segovia, M.; Abreu-Acosta, N.; Casanova, J.C.; Valladares, B.; Mas-Coma, S.; Bargues, M.D.; et al. Finding of Parastrongylus cantonensis (Chen, 1935) in Rattus rattus in Tenerife, Canary Islands (Spain). Acta Trop. 2010, 114, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Paredes-Esquivel, C.; Sola, J.; Delgado-Serra, S.; Puig Riera, M.; Negre, N.; Miranda, M.A.; Jurado-Rivera, J.A. Angiostrongylus cantonensis in North African hedgehogs as vertebrate hosts, Mallorca, Spain, October 2018. EuroSurveillance 2019, 15, 1900489. [Google Scholar] [CrossRef] [Green Version]
- Mugisha, L.; Bwangamoi, O.; Cranfeld, M.R. Angiostrongylus cantonensis and other parasites infections of rodents of Budongo Forest Reserve, Uganda. Am. J. Agric. Biol. Sci. 2012, 7, 91–95. [Google Scholar]
- York, E.M.; Creecy, J.P.; Lord, W.D.; Caire, W. Geographic range expansion for rat lungworm in North America. Emerg. Infect. Dis. 2015, 21, 1234–1236. [Google Scholar] [CrossRef]
- Stockdale Walden, H.D.; Slapcinsky, J.D.; Roff, S.; Mendieta Calle, J.; Diaz Goodwin, Z.; Stern, J.; Corlett, R.; Conway, J.; McIntosh, A. Geographic distribution of Angiostrongylus cantonensis in wild rats (Rattus rattus) and terrestrial snails in Florida, USA. PLoS ONE 2017, 12, e0177910. [Google Scholar] [CrossRef] [PubMed]
- Eamsobhana, P.; Lim, P.E.; Zhang, H.; Gan, X.; Yong, H.S. Molecular differentiation and phylogenetic relationships of three Angiostrongylus species and Angiostrongylus cantonensis geographical isolates based on a 66-kDa protein gene of A. cantonensis (Nematoda: Angiostrongylidae). Exp. Parasitol. 2010, 126, 564–569. [Google Scholar] [CrossRef]
- Alicata, J.E. Life cycle and biology. In Angiostrongyliasis in the Pacific and Southeast Asia; Alicata, J.E., Jindrak, K., Eds.; Springfield: Thomas, IL, USA, 1970; pp. 17–27. [Google Scholar]
- Lunn, J.A.; Lee, R.; Smaller, J.; MacKay, B.M.; King, T.; Hunt, G.B.; Martin, P.; Krockenberg, M.B.; Spielman, D.; Malik, R. Twenty-two cases of canine neural angiostronglyosis in eastern Australia (2002–2005) and a review of the literature. Parasites Vectors 2012, 5, 70. [Google Scholar] [CrossRef] [Green Version]
- Kliks, M.M.; Palumbo, N.E. Eosinophilic meningitis beyond the Pacific Basin: The global dispersal of a peridomestic zoonosis caused by Angiostrongylus cantonensis, the nematode lungworm of rats. Soc. Sci. Med. 1992, 34, 199–212. [Google Scholar] [CrossRef]
- Wang, Q.P.; Wu, Z.D.; Wei, J.; Owen, R.L.; Lun, Z.R. Human Angiostrongylus cantonensis: An update. Eur. J. Clin. Microbiol. 2012, 31, 389–395. [Google Scholar] [CrossRef]
- Cross, J.H.; Chen, E.R. Food-borne parasitic zoonoses. In Angiostrongyliasis; Murrell, K.D., Fried, B., Eds.; Springer: New York, NY, USA, 2007; pp. 263–290. [Google Scholar]
- Ma, G.; Dennis, M.; Rose, K.; Spratt, D.; Spielman, D. Tawny frogmouths and brushtail possums as sentinels for Angiostrongylus cantonensis, the rat lungworm. Vet. Parasitol. 2013, 192, 158–165. [Google Scholar] [CrossRef]
- Qvarnstrom, Y.; Sullivan, J.J.; Bishop, H.S.; Hollingsworth, R.; da Silva, A. PCR based detection of Angiostrongylus cantonensis in tissue and mucus secretions from molluscan hosts. Appl. Environ. Microbiol. 2007, 73, 1415–1419. [Google Scholar] [CrossRef] [Green Version]
- Beaver, P.C.; Rosen, L. Memorandum on the first report of Angiostrongylus in man, by Nomura and Lin, 1945. Am. J. Trop. Med. Hyg. 1964, 13, 589–590. [Google Scholar] [CrossRef]
- Murphy, G.S.; Johnson, S. Clinical aspects of eosinophilic meningitis and meningoencephalitis caused by Angiostrongylus cantonensis, the rat lungworm. Hawaii J. Public Health Med. 2013, 72, 35–40. [Google Scholar]
- Cowie, R.H. Angiostrongylus cantonensis: Agent of a sometimes fatal globally emerging infectious disease (rat lungworm disease). ACS Chem. Neurosci. 2017, 8, 2102–2104. [Google Scholar] [CrossRef] [Green Version]
- Prasidthrathsint, K.; Lewis, J.; Couturier, M.R. The Briefcase: Angiostrongylus cantonensis eosinophilic meningitis in a returned traveler. J. Clin. Microbiol. 2017, 55, 2880–2883. [Google Scholar] [CrossRef] [Green Version]
- Valente, R.; Robles, M.; Navone, T.G.; Diaz, I.J. Angiostrongylus spp. in the Americas: Geographical and chronological distribution of definitive hosts versus disease reports. Mem. Inst. Oswaldo Cruz 2018, 113, 143–152. [Google Scholar] [CrossRef]
- Alicata, J.E. Biology and distribution of the rat lungworm, Angiostrongylus cantonensis, and its relationship to eosinophilic meningoencephalitis and other neurological disorders of man and animals. Adv. Parasitol. 1965, 3, 223–248. [Google Scholar]
- Tokiwa, T.; Hashimoto, T.; Yabe, T.; Komatsu, N.; Akao, N.; Ohta, N. First report of Angiostrongylus cantonensis (Nematoda: Angiostrongylidae) infections in invasive rodents from five islands of the Ogasawara Archipelago, Japan. PLoS ONE 2013, 8, e70729. [Google Scholar] [CrossRef]
- Wallace, G.D.; Rosen, L. Studies on eosinophilic meningitis. I. Observations on the geographic distribution of Angiostrongylus cantonensis in the Pacific Area and its prevalence in wild rats. Am. J. Epidemiol. 1965, 81, 52–62. [Google Scholar] [CrossRef]
- Hochberg, N.S.; Park, S.Y.; Blackburn, B.G.; Sejvar, J.J.; Gaynor, K.; Chung, H.; Leniek, K.; Herwaldt, B.L.; Effler, P.V. Distribution of eosinophilic meningitis cases attributable to Angiostrongylus cantonensis. Hawaii Emerg. Infect. Dis. 2007, 13, 1675–1680. [Google Scholar]
- York, E.M.; Butler, C.J.; Lord, W.D. Global decline in suitable habitat for Angiostrongylus (=Parastrongylus) cantonensis: The role of climate change. PLoS ONE 2014, 9, e103831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- State Meteorological Agency of Spain (Ed.) Climate Atlas of the Archipelagos of the Canary Islands, Madeira and the Azores. Ministry of Agriculture, Food and Environment, Spain. p. 80. 2012. Available online: http://www.aemet.es/documentos/es/conocermas/publicaciones/2Atlasclimatologico/AtlasClimaMacaronesiaBaja.pdf.</doc (accessed on 3 March 2021).
- Fernández-Palacios, J.M.; Santos, A.D.L. Ecología de las Islas Canarias; Soc. La Cosmológica: Santa Cruz de Tenerife, Spain, 1996. [Google Scholar]
- Fernández-Palacios, J.M.; Whittaker, R.J. The Canaries: An important biogeographical meeting place. J. Biogeogr. 2008, 35, 379–387. [Google Scholar] [CrossRef]
- Kinsella, J.M. Angiostrongylus schmidti sp.n. (Nematoda: Metastrongyloidea) from the rice rat, Oryzomys palustris, in Florida, with a key to the species of Angiostrongylus kamensky, 1905. J. Parasitol. 1971, 57, 494–497. [Google Scholar] [CrossRef] [PubMed]
- Drozdz, J.; Gorecka, T.; Binh, N.H. The occurrence of nematodes of the subfamily Angiostrongylinae in Viet-Nam and the question of geographical origin of Parastrongylus cantonensis (Chen, 1935). Acta Parasitol. Pol. 1975, 23, 115–126. [Google Scholar]
- Ubelaker, J.E. Systematics of species referred to the genus Angiostrongylus. J. Parasitol. 1986, 72, 237–244. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. A language and environment for statistical computing. In R Foundation for Statistical Computing; R Core Team: Vienna, Austria, 2019. [Google Scholar]
- Martin, A.; Hernández, M.A.; Lorenzo, J.A.; Nogales, M.; González, C. Las Palomas Endémicas de Canarias; Viceconsejería de Medio Ambiente del Gobierno de Canarias y SEO/BirdLife: Santa Cruz de Tenerife, Spain, 2000; p. 191. [Google Scholar]
- Zamora, I.I.; Esquivel, J.L.M.; Pérez, N.Z.; Hernández, M.A. Lista de Especies Silvestres de Canarias: Hongos, Plantas y Animales Terrestres; Consejería de Medio Ambiente y Ordenación Territorial del Gobierno de Canarias: San Cristóbal de La Laguna, Spain, 2004. [Google Scholar]
- Martin-Alonso, A.; Abreu-Yanes, E.; Feliu, C.; Mas-Coma, S.; Bargues, M.D.; Valladares, B.; Foronda, P. Intermediate Hosts of Angiostrongylus cantonensis in Tenerife, Spain. PLoS ONE 2015, 10, e0120686. [Google Scholar] [CrossRef] [Green Version]
- Jefferies, R.; Shaw, S.E.; Viney, M.E.; Morgan, E.R. Angiostrongylus vasorum from South America and Europe represent distinct lineages. Parasitology 2009, 136, 107–115. [Google Scholar] [CrossRef]
- Monte, T.C.C.; Simões, R.O.; Oliveira, A.P.; Novaes, C.F.; Thiengo, S.C.; Silva, A.J.; Estrela, P.C.; Maldonado, A., Jr. Phylogenetic relationship of the Brazilian isolates of the rat lungworm Angiostrongylus cantonensis (Nematoda: Metastrongylidae) employing mitochondrial COI gene sequence data. Parasit Vectors 2012, 5, 248. [Google Scholar] [CrossRef] [Green Version]
- Červená, B.; Modrý, D.; Fecková, B.; Hrazdilová, K.; Foronda, P.; Alonso, A.M.; Lee, R.; Walker, J.; Niebuhr, C.N.; Malik, R.; et al. Low diversity of Angiostrongylus cantonensis complete mitochondrial DNA sequences from Australia, Hawaii, French Polynesia and the Canary Islands revealed using whole genome next-generation sequencing. Parasit Vectors 2019, 12, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, S.; Zhou, X.N.; Zhang, Y.; Liu, H.X.; Zhu, D.; Yin, W.G.; Steinmann, P.; Wang, X.H.; Jia, T.W. The effect of temperature on the development of Angiostrongylus cantonensis (Chen 1935) in Pomacea canaliculata (Lamarck 1822). Parasitol. Res. 2006, 99, 583–587. [Google Scholar] [CrossRef] [PubMed]
- Puertos del Estado. Autoridad Portuaria Santa Cruz de Tenerife. Ministerios de Transporte, Movilidad y Agenda Urbana. 2019. Available online: https://www.puertosdetenerife.org (accessed on 10 March 2021).
- Puertos del Estado. Autoridad Portuaria de Las Palmas. Ministerios de Transporte, Movilidad y Agenda Urbana. 2019. Available online: http://www.palmasport.es/es/ (accessed on 10 March 2021).
- Lafferty, K.D. The ecology of climate change and infectious diseases. Ecology 2009, 90, 888–900. [Google Scholar] [CrossRef] [PubMed]
- Kutz, S.J.; Hoberg, E.P.; Polley, L.; Jenkins, E.J. Global warming is changing the dynamics of Arctic host-parasite systems. Proc. R. Soc. Lond. Biol. 2005, 272, 2571–2576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salb, A.L.; Barkema, H.W.; Elkin, V.T.; Thompson, R.C.; Whiteside, D.P.; Black, S.R.; Dubey, J.P.; Kutz, S.J. Dogs as sources and sentinels of parasites in humans and wildlife, Northern Canada. Emerg. Infect. Dis. 2008, 14, 60–63. [Google Scholar] [CrossRef]
- Kutz, S.J.; Jenkins, E.J.; Veitch, A.M.; Ducrocq, J.; Polley, L.; Elkin, B.; Lair, S. The Arctic as a model for anticipating, preventing, and mitigating climate change impacts on host parasite interactions. Vet. Parasitol. 2009, 163, 217–228. [Google Scholar] [CrossRef]
- Yang, T.B.; Wu, Z.D.; Lun, Z.R. The apple snail Pomacea canaliculata, a novel vector of the rat lungworm, Angiostrongylus cantonensis: Its introduction, spread, and control in China. Hawaii J. Med. Public Health. 2013, 72, 23. [Google Scholar]
- Cowie, R.H. Pathways for transmission of angiostrongyliasis and the risk of disease associated with them. Hawaii J. Med. Public Health 2013, 72, 70–74. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
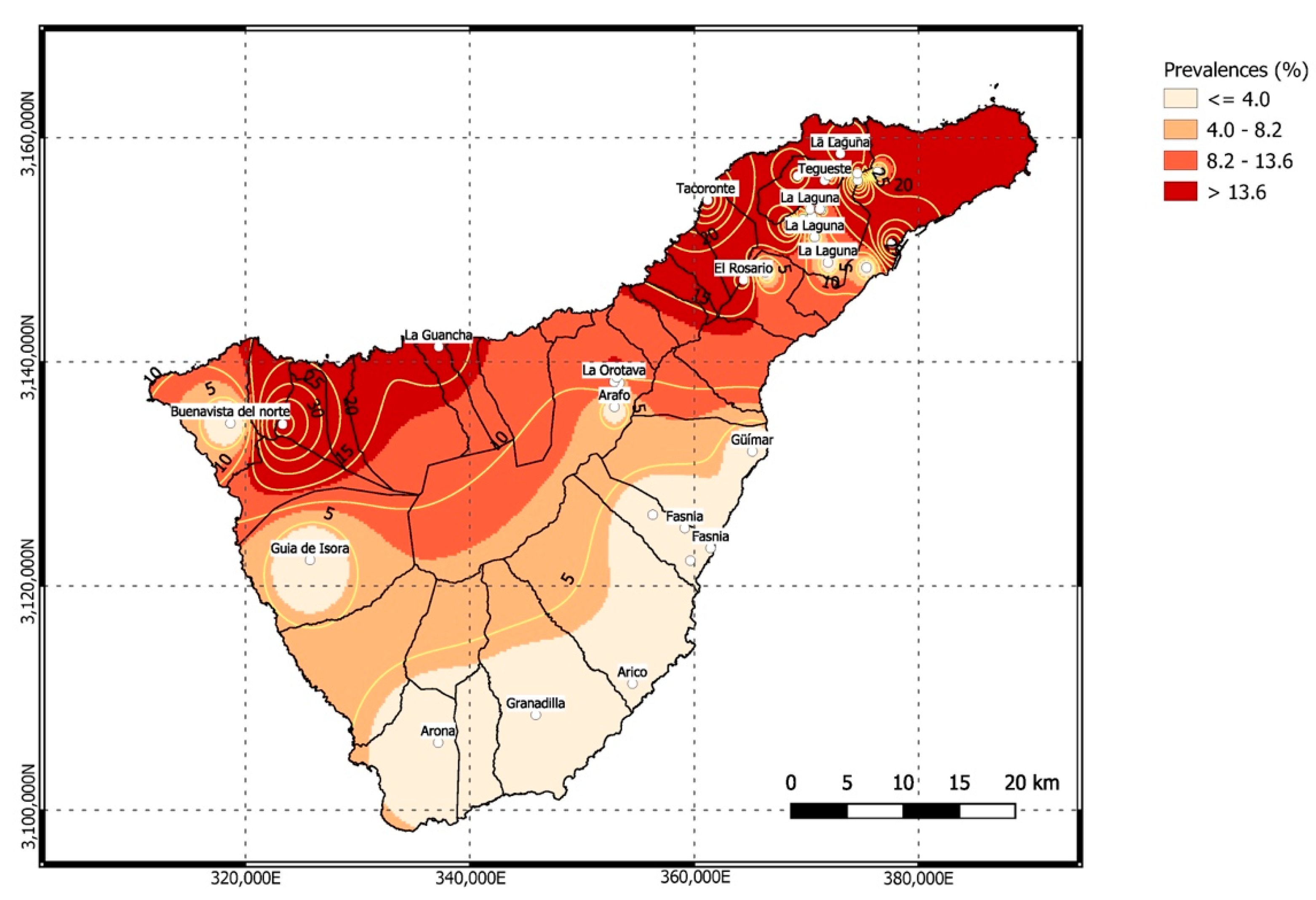
| Locations | Rattus rattus | Rattus norvegicus | Mus musculus domesticus | Total |
|---|---|---|---|---|
| P (%) (+/n) | P (%) (+/n) | P (%) (+/n) | P (%) (+/n) | |
| North | 22.79 (31/136) | 9.09 (1/11) | 0 (0/107) | 12.59 (32/254) |
| Buenavista del Norte | 0 (0/5) | 0 (0/1) | 0 (0/2) | 0 (0/8) |
| El Rosario | 21.05 (4/19) | 0 (0/0) | 0 (0/4) | 17.39 (4/23) |
| El Sauzal | 100 (4/4) | 0 (0/9) | 0 (0/9) | 18.18 (4/22) |
| El Tanque | 37.5 (6/16) | 0 (0/0) | 0 (0/1) | 35.29 (6/17) |
| La Guancha | 16.6 (1/6) | 0 (0/0) | 0 (0/5) | 9.09 (1/11) |
| La Orotava | 20 (7/35) | 0 (0/0) | 0 (0/54) | 7.86 (7/89) |
| Tegueste | 17.64 (9/51) | 100 (1/1) | 0 (0/32) | 11.90 (10/84) |
| South | 0 (0/28) | 0 (0/8) | 0 (0/13) | 0 (0/49) |
| Arico | 0 (0/0) | 0 (0/6) | 0 (0/0) | 0 (0/6) |
| Arona | 0 (0/2) | 0 (0/0) | 0 (0/2) | 0 (0/4) |
| Fasnia | 0 (0/19) | 0 (0/2) | 0 (0/1) | 0 (0/22) |
| Granadilla | 0 (0/1) | 0 (0/0) | 0 (0/3) | 0 (0/4) |
| Guía de Isora | 0 (0/6) | 0 (0/0) | 0 (0/0) | 0 (0/6) |
| Güímar | 0 (0/0) | 0 (0/0) | 0 (0/3) | 0 (0/3) |
| San Miguel de Abona | 0 (0/0) | 0 (0/0) | 0 (0/4) | 0 (0/4) |
| Santiago del Teide | 0 (0/0) | 0 (0/0) | 0 (0/1) | 0 (0/1) |
| Main cities | 24.04 (25/104) | 0 (0/0) | 0 (0/51) | 16.13 (25/155) |
| San Cristóbal de La Laguna | 22.82 (21/92) | 0 (0/0) | 0 (0/36) | 16.4 (21/128) |
| Santa Cruz de Tenerife | 33.33 (4/12) | 0 (0/0) | 0 (0/15) | 14.81(4/27) |
| Total | 21.20 (60/283) | 7.14 (1/14) | (0/181) | 12.76 (61/478) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martín-Carrillo, N.; Feliu, C.; Abreu-Acosta, N.; Izquierdo-Rodriguez, E.; Dorta-Guerra, R.; Miquel, J.; Abreu-Yanes, E.; Martin-Alonso, A.; García-Livia, K.; Quispe-Ricalde, M.A.; et al. A Peculiar Distribution of the Emerging Nematode Angiostrongylus cantonensis in the Canary Islands (Spain): Recent Introduction or Isolation Effect? Animals 2021, 11, 1267. https://doi.org/10.3390/ani11051267
Martín-Carrillo N, Feliu C, Abreu-Acosta N, Izquierdo-Rodriguez E, Dorta-Guerra R, Miquel J, Abreu-Yanes E, Martin-Alonso A, García-Livia K, Quispe-Ricalde MA, et al. A Peculiar Distribution of the Emerging Nematode Angiostrongylus cantonensis in the Canary Islands (Spain): Recent Introduction or Isolation Effect? Animals. 2021; 11(5):1267. https://doi.org/10.3390/ani11051267
Chicago/Turabian StyleMartín-Carrillo, Natalia, Carlos Feliu, Néstor Abreu-Acosta, Elena Izquierdo-Rodriguez, Roberto Dorta-Guerra, Jordi Miquel, Estefanía Abreu-Yanes, Aarón Martin-Alonso, Katherine García-Livia, María Antonieta Quispe-Ricalde, and et al. 2021. "A Peculiar Distribution of the Emerging Nematode Angiostrongylus cantonensis in the Canary Islands (Spain): Recent Introduction or Isolation Effect?" Animals 11, no. 5: 1267. https://doi.org/10.3390/ani11051267