Repeated Episodes of Heroin Cause Enduring Alterations of Circadian Activity in Protracted Abstinence

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
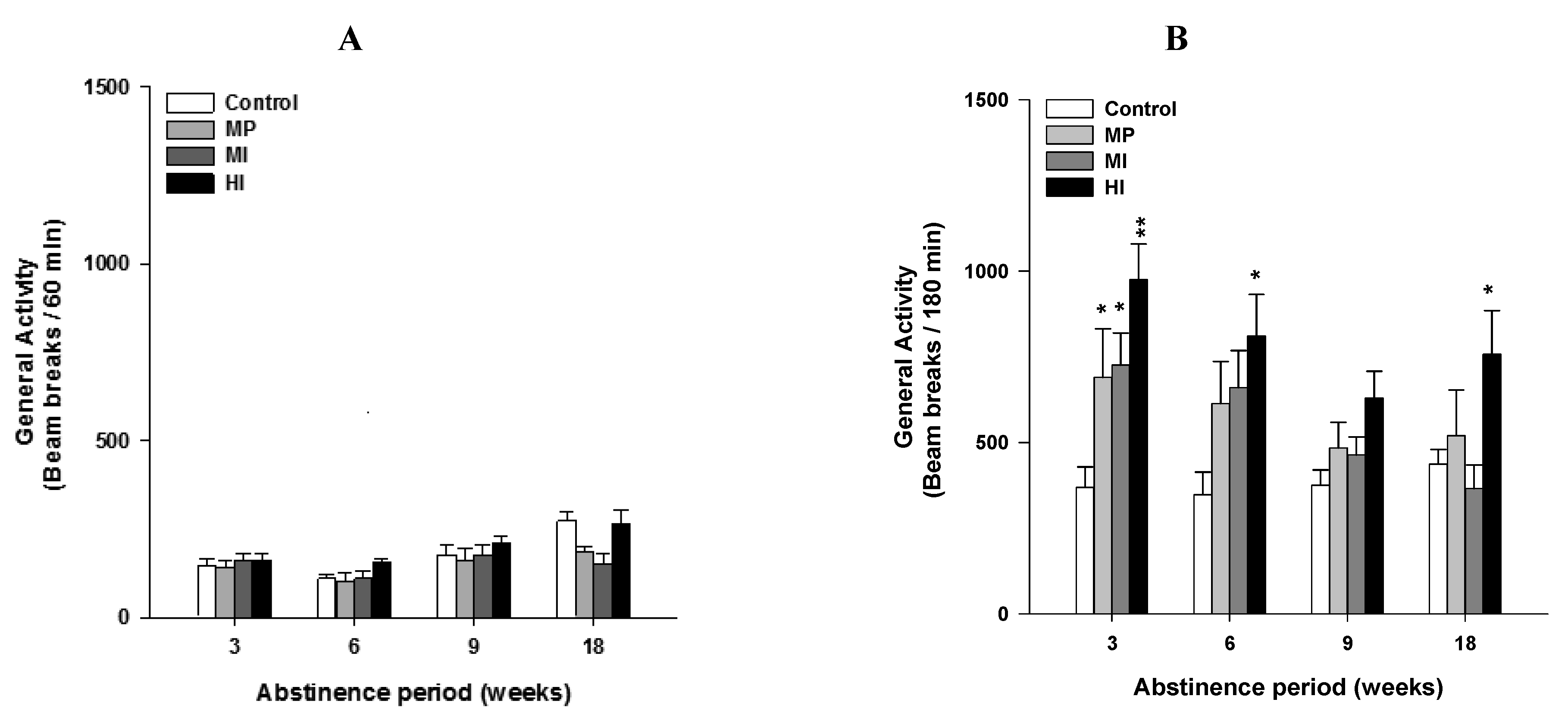
2.1. Experiment 1: Long Lasting Sensitivity to Heroin Challenge Depends on Previous History of Opiate Dependence (Figure 1)

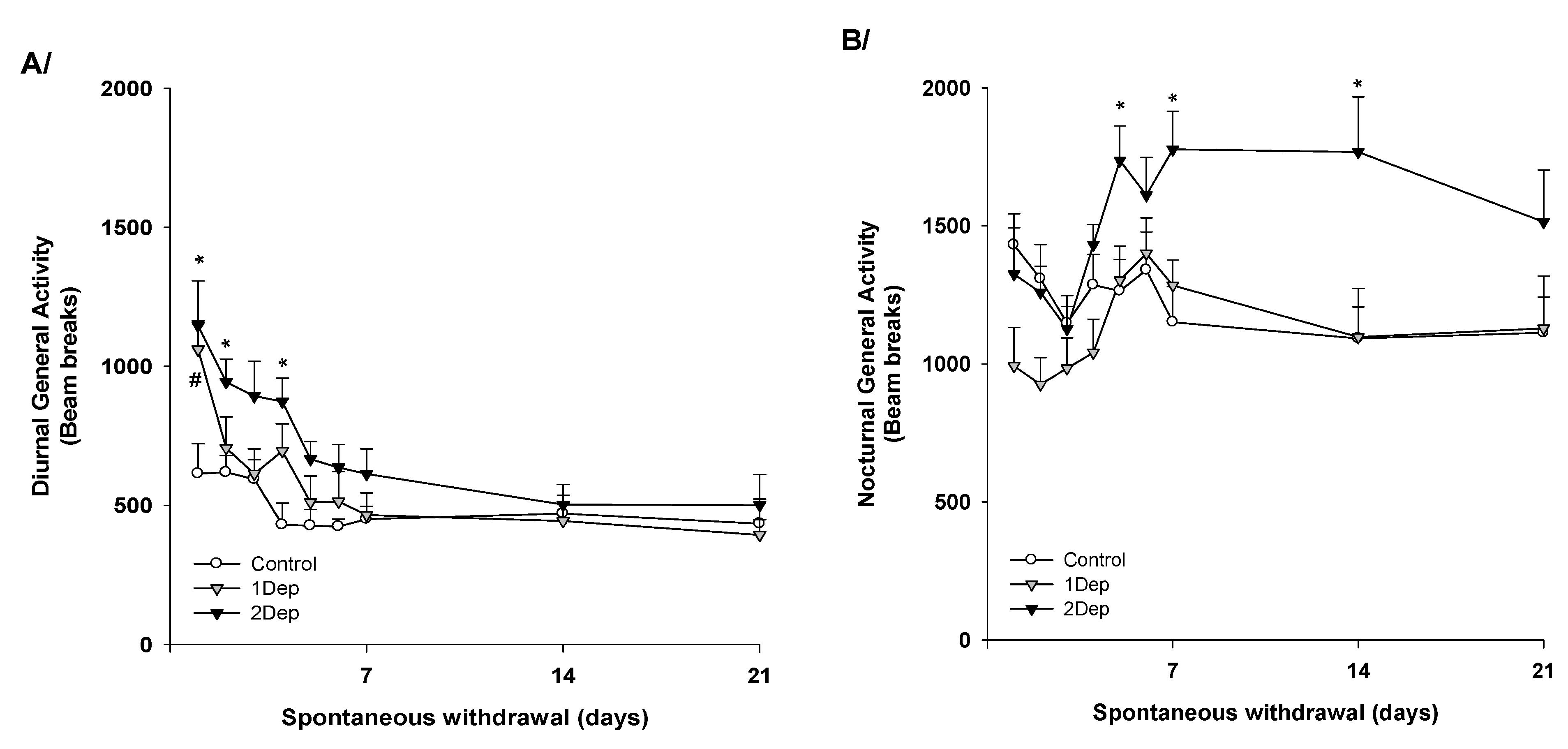
2.2. Experiment 2: Repeated Episodes of Heroin Dependence: Consequences on the Circadian Activity upon Abrupt Cessation (Figure 2)

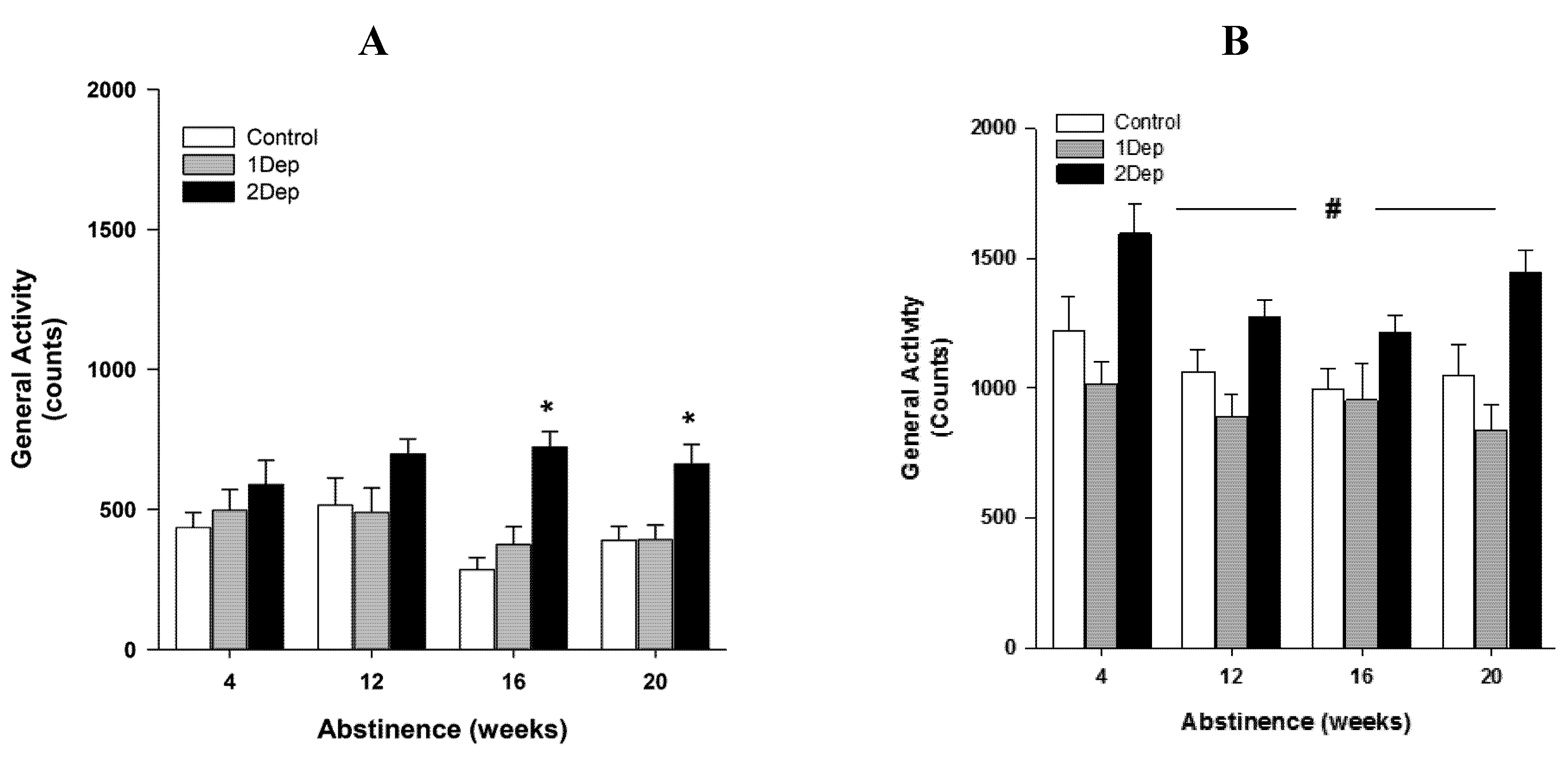
2.3. Experiment 3: Effect of Intermittent Exposures to Heroin: A Time-Course Analysis of the Circadian Activity throughout Extended Abstinence (Figure 3)

3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Opiate Dependence Induction
4.2.1. Morphine Pellets (MP)
4.2.2. Morphine Injections (MI)
4.2.3. Heroin Injections (HI)
4.3. Activity Recording
4.4. Experimental Design
4.4.1. Experiment 1: Long Lasting Sensitivity to Heroin Challenge Depends on the Previous History of Opiate Dependence
4.4.2. Experiment 2: Repeated Episodes of Heroin Dependence: Consequences on the Circadian Activity upon Abrupt Cessation
4.4.3. Experiment 3: Effects of Intermittent Exposures to Heroin: A Time-Course Analysis of the Circadian Activity throughout Extended Abstinence
4.5. Statistical Analyses
5. Conclusion
Acknowledgments
References
- Deroche-Gamonet, V.; Belin, D.; Piazza, P.V. Evidence for addiction-like behavior in the rat. Science 2004, 305, 1014–1017. [Google Scholar] [CrossRef]
- Koob, G.F.; Le Moal, M. Drug addiction, dysregulation of reward, and allostasis. Neuropsychopharmacology 2001, 24, 97–129. [Google Scholar] [CrossRef]
- Vanderschuren, L.J.; Everitt, B.J. Drug seeking becomes compulsive after prolonged cocaine self-administration. Science 2004, 305, 1017–1019. [Google Scholar] [CrossRef]
- World Health Organization, International Statistical Classification of Diseases and Related Problems; World Health Organization: Geneva, Switzerland, 1992.
- O’Brien, C.P. Anticraving medications for relapse prevention: A possible new class of psychoactive medications. Am. J. Psychiatry 2005, 162, 1423–1431. [Google Scholar] [CrossRef]
- Shaham, Y.; Shalev, U.; Lu, L.; de Wit, H.; Stewart, J. The reinstatement model of drug relapse: History, methodology and major findings. Psychopharmacology (Berl.) 2003, 168, 3–20. [Google Scholar] [CrossRef]
- Gossop, M.; Green, L.; Phillips, G.; Bradley, B. Lapse, relapse and survival among opiate addicts after treatment. A prospective follow-up study. Br. J. Psychiatry 1989, 154, 348–353. [Google Scholar] [CrossRef]
- Li, S.X.; Shi, J.; Epstein, D.H.; Wang, X.; Zhang, X.L.; Bao, Y.P.; Zhang, D.; Zhang, X.Y.; Kosten, T.R.; Lu, L. Circadian alteration in neurobiology during 30 days of abstinence in heroin users. Biol. Psychiatry 2009, 65, 905–912. [Google Scholar] [CrossRef]
- Martin, W.R.; Jasinski, D.R. Physiological parameters of morphine dependence in man—Tolerance, early abstinence, protracted abstinence. J. Psychiatr. Res. 1969, 7, 9–17. [Google Scholar] [CrossRef]
- Khazan, N.; Colasanti, B. Decline in the mean integrated electroencephalogram voltage during morphine abstinence in the rat. J. Pharmacol. Exp. Ther. 1971, 177, 491–499. [Google Scholar]
- Caillé, S.; Espejo, E.F.; Koob, G.F.; Stinus, L. Dorsal and median raphe serotonergic system lesion does not alter the opiate withdrawal syndrome. Pharmacol. Biochem. Behav. 2002, 72, 979–986. [Google Scholar] [CrossRef]
- Caillé, S.; Espejo, E.F.; Reneric, J.P.; Cador, M.; Koob, G.F.; Stinus, L. Total neurochemical lesion of noradrenergic neurons of the locus coeruleus does not alter either naloxone-precipitated or spontaneous opiate withdrawal nor does it influence ability of clonidine to reverse opiate withdrawal. J. Pharmacol. Exp. Ther. 1999, 290, 881–892. [Google Scholar]
- Robert, C.; Stinus, L.; Limoge, A. Sleep impairments in the rats implanted with morphine pellets. Neuropsychobiology 1999, 40, 214–217. [Google Scholar] [CrossRef]
- Koob, G.F.; Stinus, L.; Le Moal, M.; Bloom, F.E. Opponent process theory of motivation: Neurobiological evidence from studies of opiate dependence. Neurosci. Biobehav. Rev. 1989, 13, 135–149. [Google Scholar] [CrossRef]
- O’Brien, C.P.; Jaffe, J.H. Addictive States; Raven Press: New York, NY, USA, 1992. [Google Scholar]
- Solomon, R.L.; Corbit, R. An opponent-process theory of motivation: Temporal dynamics of effect. Psychol. Rev. 1974, 81, 119–145. [Google Scholar] [CrossRef]
- Wikler, A.; Pescor, F.T. Classical conditioning of a morphine abstinence phenomenon, reinforcement of opioid-drinking behavior and “Relapse”in morphine-addicted rats. Psychopharmacologia 1967, 10, 255–284. [Google Scholar] [CrossRef]
- Kenny, P.J.; Chen, S.A.; Kitamura, O.; Markou, A.; Koob, G.F. Conditioned withdrawal drives heroin consumption and decreases reward sensitivity. J. Neurosci. 2006, 26, 5894–5900. [Google Scholar] [CrossRef]
- Harris, G.C.; Aston-Jones, G. Enhanced morphine preference following prolonged abstinence: Association with increased Fos expression in the extended amygdala. Neuropsychopharmacology 2003, 28, 292–299. [Google Scholar] [CrossRef]
- Shalev, U.; Morales, M.; Hope, B.; Yap, J.; Shaham, Y. Time-dependent changes in extinction behavior and stress-induced reinstatement of drug seeking following withdrawal from heroin in rats. Psychopharmacology (Berl.) 2001, 156, 98–107. [Google Scholar] [CrossRef]
- Vanderschuren, L.J.; Tjon, G.H.; Nestby, P.; Mulder, A.H.; Schoffelmeer, A.N.; de Vries, T.J. Morphine-induced long-term sensitization to the locomotor effects of morphine and amphetamine depends on the temporal pattern of the pretreatment regimen. Psychopharmacology (Berl.) 1997, 131, 115–122. [Google Scholar] [CrossRef]
- Shaham, Y.; Rajabi, H.; Stewart, J. Relapse to heroin-seeking in rats under opioid maintenance: The effect of stress, heroin priming and withdrawal. J. Neurosci. 1996, 16, 1957–1963. [Google Scholar]
- Hellemans, K.G.; Dickinson, A.; Everitt, B.J. Motivational control of heroin seeking by conditioned stimuli associated with withdrawal and heroin taking by rats. Behav. Neurosci. 2006, 120, 103–114. [Google Scholar] [CrossRef]
- Lu, L.; Chen, H.; Su, W.; Ge, X.; Yue, W.; Su, F.; Ma, L. Role of withdrawal in reinstatement of morphine-conditioned place preference. Psychopharmacology (Berl.) 2005, 181, 90–100. [Google Scholar] [CrossRef]
- Aston-Jones, G.; Harris, G.C. Brain substrates for increased drug seeking during protracted withdrawal. Neuropharmacology 2004, 47, 167–179. [Google Scholar] [CrossRef]
- Stinus, L.; Caille, S.; Koob, G.F. Opiate withdrawal induced place aversion lasts for up to 16 weeks. Psychopharmacology (Berl.) 2000, 149, 115–120. [Google Scholar] [CrossRef]
- Schulteis, G.; Heyser, C.J.; Koob, G.F. Opiate withdrawal signs precipitated by naloxone following a single exposure to morphine: Potentiation with repeated morphine treatment. Psychopharmacology (Berl.) 1997, 129, 56–65. [Google Scholar] [CrossRef]
- Azorlosa, J.L.; Stitzer, M.L.; Greenwald, M.K. Opioid physical dependence development: Effects of single versus repeated pretreatments and of subjects opioid exposure history. Psychopharmacology (Berl.) 1994, 114, 71–80. [Google Scholar] [CrossRef]
- Cohen, A.; Koob, G.F.; George, O. Robust escalation of nicotine intake with extended access to nicotine self-administration and intermittent periods of abstinence. Neuropsychopharmacology 2012, 37, 2153–2160. [Google Scholar] [CrossRef]
- Laorden, M.L.; Fuertes, G.; Gonzalez-Cuello, A.; Milanes, M.V. Changes in catecholaminergic pathways innervating paraventricular nucleus and pituitary-adrenal axis response during morphine dependence: Implication of alpha(1)- and alpha(2)-adrenoceptors. J. Pharmacol. Exp. Ther. 2000, 293, 578–584. [Google Scholar]
- Milanes, M.V.; Laorden, M.L.; Chapleur-Chateau, M.; Burlet, A. Alterations in corticotropin-releasing factor and vasopressin content in rat brain during morphine withdrawal: Correlation with hypothalamic noradrenergic activity and pituitary-adrenal response. J. Pharmacol. Exp. Ther. 1998, 285, 700–706. [Google Scholar]
- Houshyar, H.; Galigniana, M.D.; Pratt, W.B.; Woods, J.H. Differential responsivity of the hypothalamic-pituitary-adrenal axis to glucocorticoid negative-feedback and corticotropin releasing hormone in rats undergoing morphine withdrawal: Possible mechanisms involved in facilitated and attenuated stress responses. J. Neuroendocrinol. 2001, 13, 875–886. [Google Scholar]
- Houshyar, H.; Gomez, F.; Manalo, S.; Bhargava, A.; Dallman, M.F. Intermittent morphine administration induces dependence and is a chronic stressor in rats. Neuropsychopharmacology 2003, 28, 1960–1972. [Google Scholar] [CrossRef]
- Harrigan, S.E.; Downs, D.A. Self-administration of heroin, acetylmethadol, morphine, and methadone in rhesus monkeys. Life Sci. 1978, 22, 619–623. [Google Scholar] [CrossRef]
- Hubner, C.B.; Kornetsky, C. Heroin, 6-acetylmorphine and morphine effects on threshold for rewarding and aversive brain stimulation. J. Pharmacol. Exp. Ther. 1992, 260, 562–567. [Google Scholar]
- Van Ree, J.M.; Slangen, J.L.; de Wied, D. Intravenous self-administration of drugs in rats. J. Pharmacol. Exp. Ther. 1978, 204, 547–557. [Google Scholar]
- Ungless, M.A.; Whistler, J.L.; Malenka, R.C.; Bonci, A. Single cocaine exposure in vivo induces long-term potentiation in dopamine neurons. Nature 2001, 411, 583–587. [Google Scholar] [CrossRef]
- Ciccocioppo, R.; Martin-Fardon, R.; Weiss, F. Stimuli associated with a single cocaine experience elicit long-lasting cocaine-seeking. Nat. Neurosci. 2004, 7, 495–496. [Google Scholar] [CrossRef]
- McLellan, A.T.; Lewis, D.C.; O’Brien, C.P.; Kleber, H.D. Drug dependence, a chronic medical illness: Implications for treatment, insurance, and outcomes evaluation. JAMA 2000, 284, 1689–1695. [Google Scholar] [CrossRef]
- Chen, S.A.; O’Dell, L.E.; Hoefer, M.E.; Greenwell, T.N.; Zorrilla, E.P.; Koob, G.F. Unlimited access to heroin self-administration: Independent motivational markers of opiate dependence. Neuropsychopharmacology 2006, 31, 2692–2707. [Google Scholar] [CrossRef]
- Ren, Z.Y.; Zhang, X.L.; Liu, Y.; Zhao, L.Y.; Shi, J.; Bao, Y.; Zhang, X.Y.; Kosten, T.R.; Lu, L. Diurnal variation in cue-induced responses among protracted abstinent heroin users. Pharmacol. Biochem. Behav. 2009, 91, 468–472. [Google Scholar] [CrossRef]
- Boutrel, B.; Cannella, N.; de Lecea, L. The role of hypocretin in driving arousal and goal-oriented behaviors. Brain Res. 2010, 1314, 103–111. [Google Scholar] [CrossRef]
- Mahler, S.V.; Smith, R.J.; Moorman, D.E.; Sartor, G.C.; Aston-Jones, G. Multiple roles for orexin/hypocretin in addiction. Prog. Brain Res. 2012, 198, 79–121. [Google Scholar] [CrossRef]
- Gawin, F.H.; Kleber, H.D. Abstinence symptomatology and psychiatric diagnosis in cocaine abusers. Clinical observations. Arch. Gen. Psychiatry 1986, 43, 107–113. [Google Scholar] [CrossRef]
- Grimm, J.W.; Hope, B.T.; Wise, R.A.; Shaham, Y. Neuroadaptation: Incubation of cocaine craving after withdrawal. Nature 2001, 412, 141–142. [Google Scholar] [CrossRef]
- Gold, L.H.; Stinus, L.; Inturrisi, C.E.; Koob, G.F. Prolonged tolerance, dependence and abstinence following subcutaneous morphine pellet implantation in the rat. Eur. J. Pharmacol. 1994, 253, 45–51. [Google Scholar] [CrossRef]
- Robbins, T.W.; Ersche, K.D.; Everitt, B.J. Drug addiction and the memory systems of the brain. Ann. N. Y. Acad. Sci. 2008, 1141, 1–21. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Stinus, L.; Cador, M.; Caille, S. Repeated Episodes of Heroin Cause Enduring Alterations of Circadian Activity in Protracted Abstinence. Brain Sci. 2012, 2, 421-433. https://doi.org/10.3390/brainsci2030421
Stinus L, Cador M, Caille S. Repeated Episodes of Heroin Cause Enduring Alterations of Circadian Activity in Protracted Abstinence. Brain Sciences. 2012; 2(3):421-433. https://doi.org/10.3390/brainsci2030421
Chicago/Turabian StyleStinus, Luis, Martine Cador, and Stephanie Caille. 2012. "Repeated Episodes of Heroin Cause Enduring Alterations of Circadian Activity in Protracted Abstinence" Brain Sciences 2, no. 3: 421-433. https://doi.org/10.3390/brainsci2030421