Circulating miR-16-5p, miR-92a-3p, and miR-451a in Plasma from Lung Cancer Patients: Potential Application in Early Detection and a Regulatory Role in Tumorigenesis Pathways

 ,
, 
 ,
, 
Abstract
:1. Introduction
2. Results
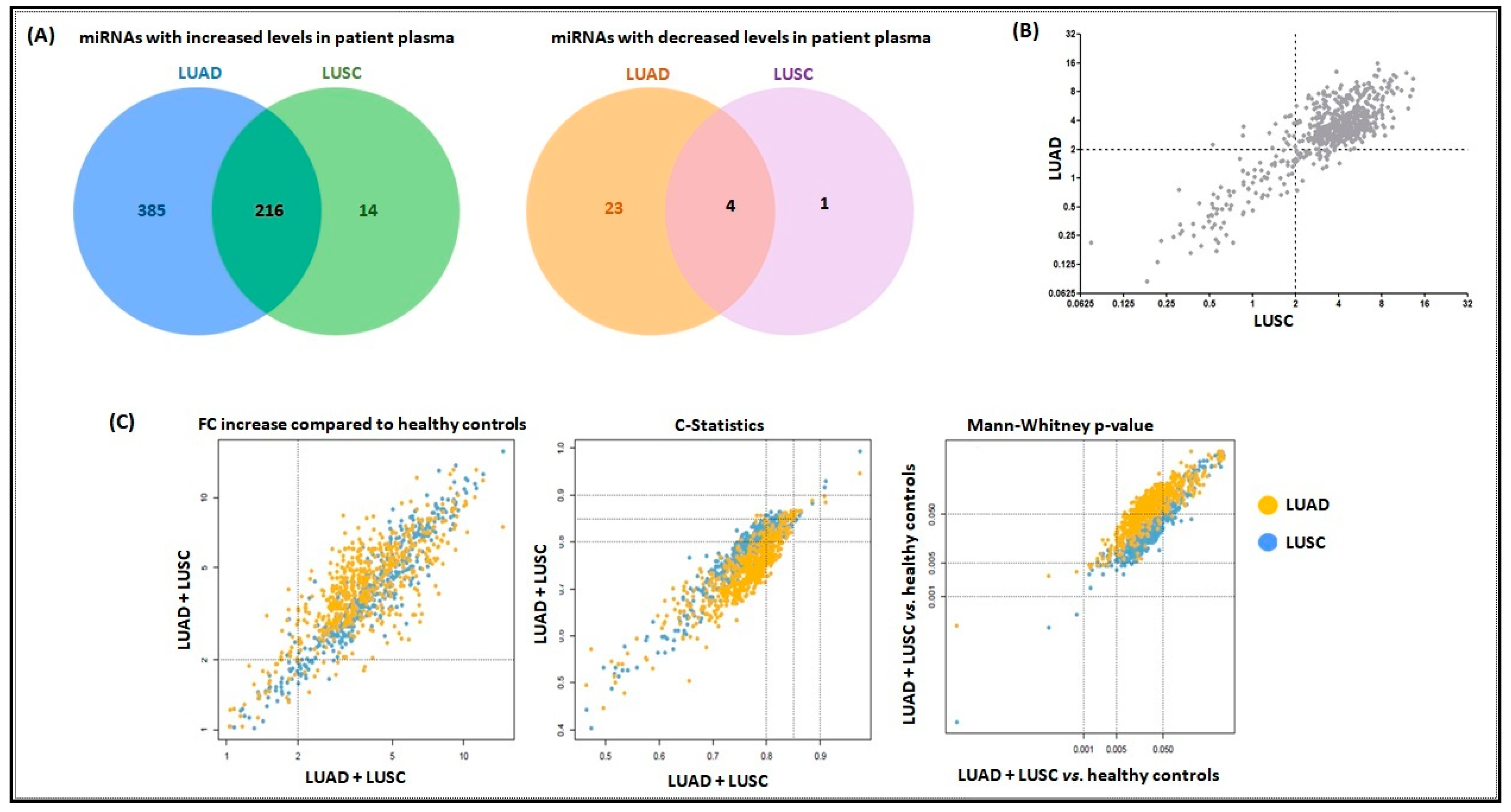
2.1. Higher Levels of Plasma miRNAs are Found in LUAD and LUSC Cases Versus Controls
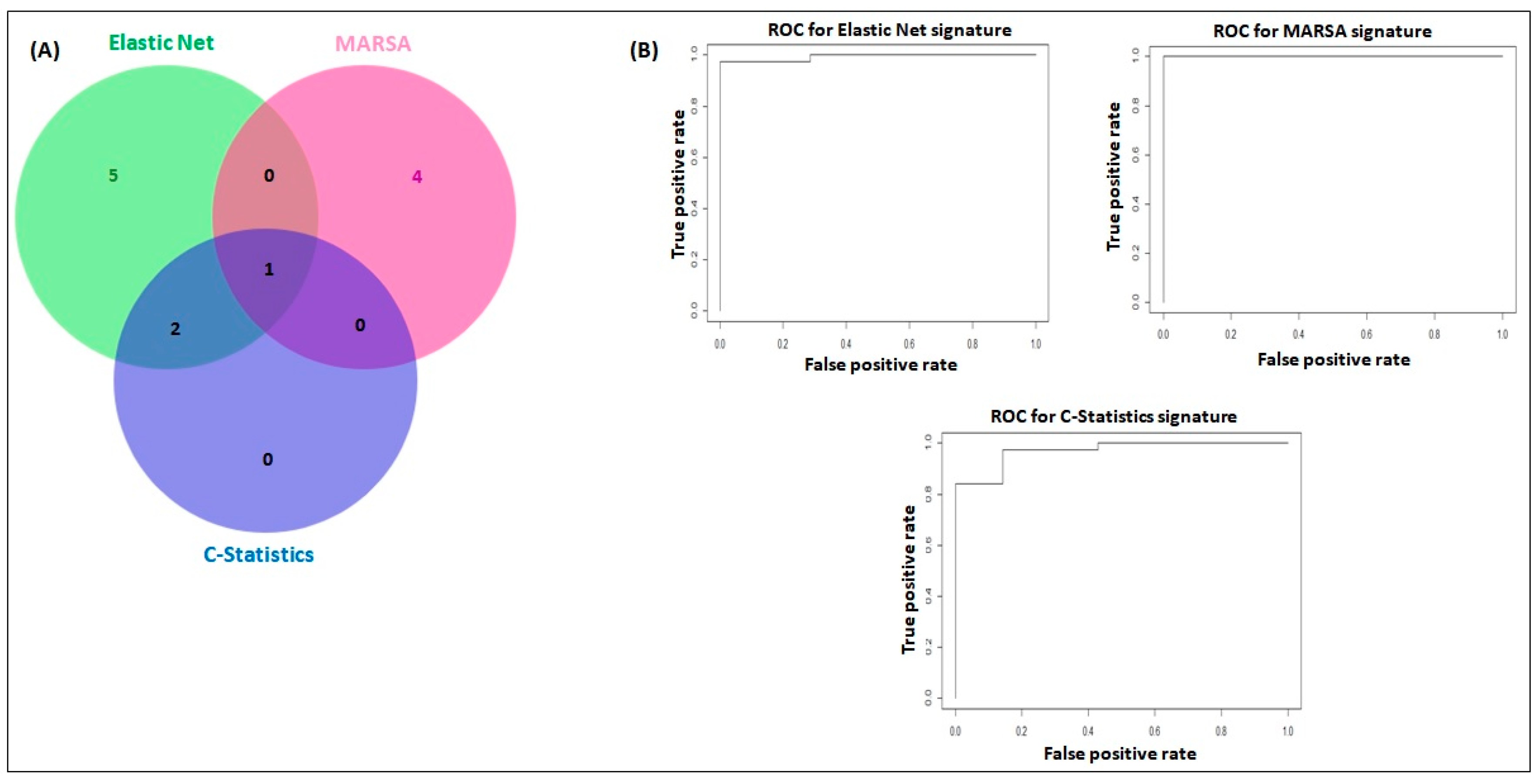
2.2. miRNA Signatures Have Potential Application in Early Detection of LUAD and LUSC
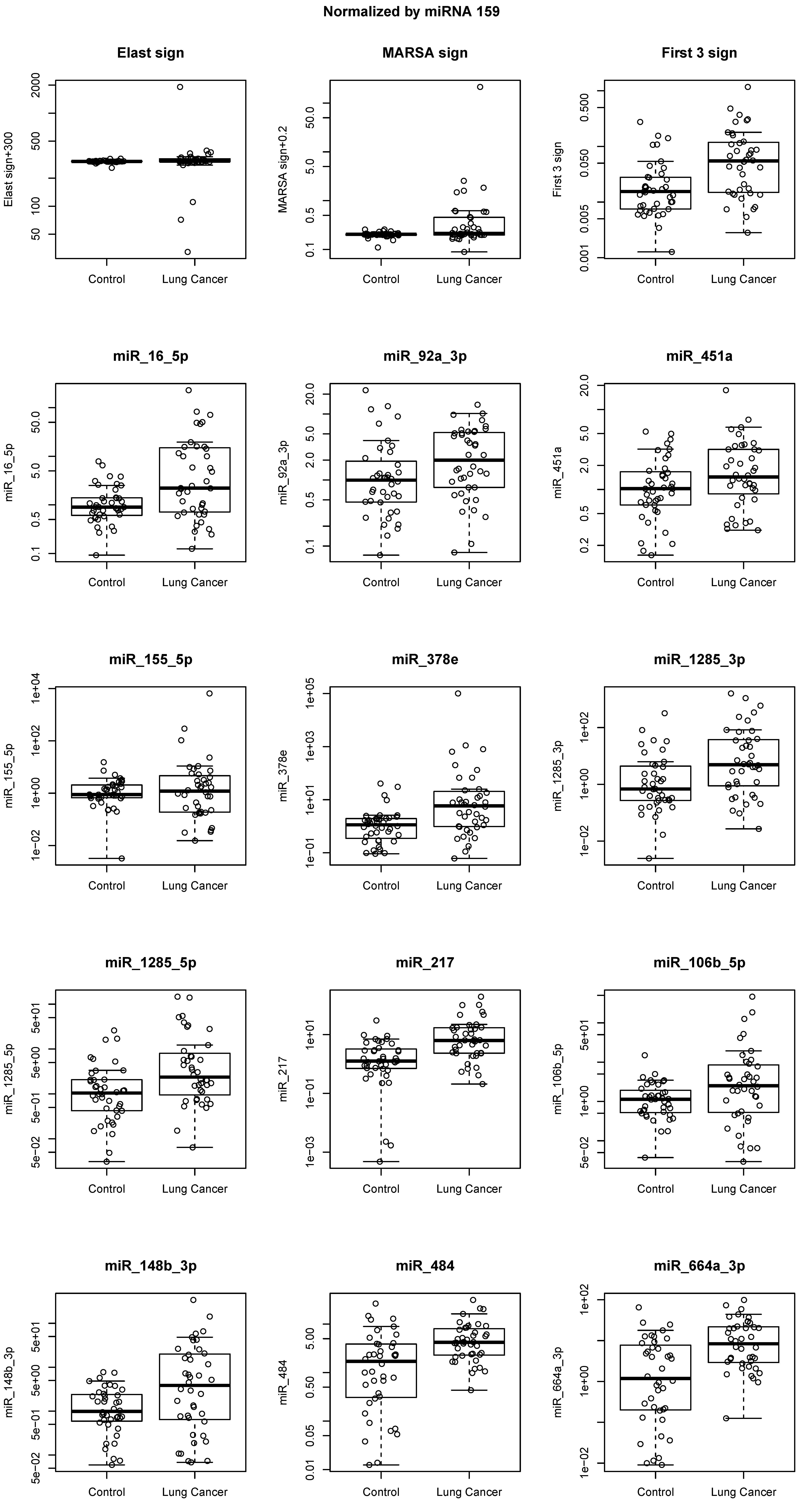
2.3. miRNA Signatures and Individual miRNA Levels Were Verified in a Separate Set of Plasma Samples
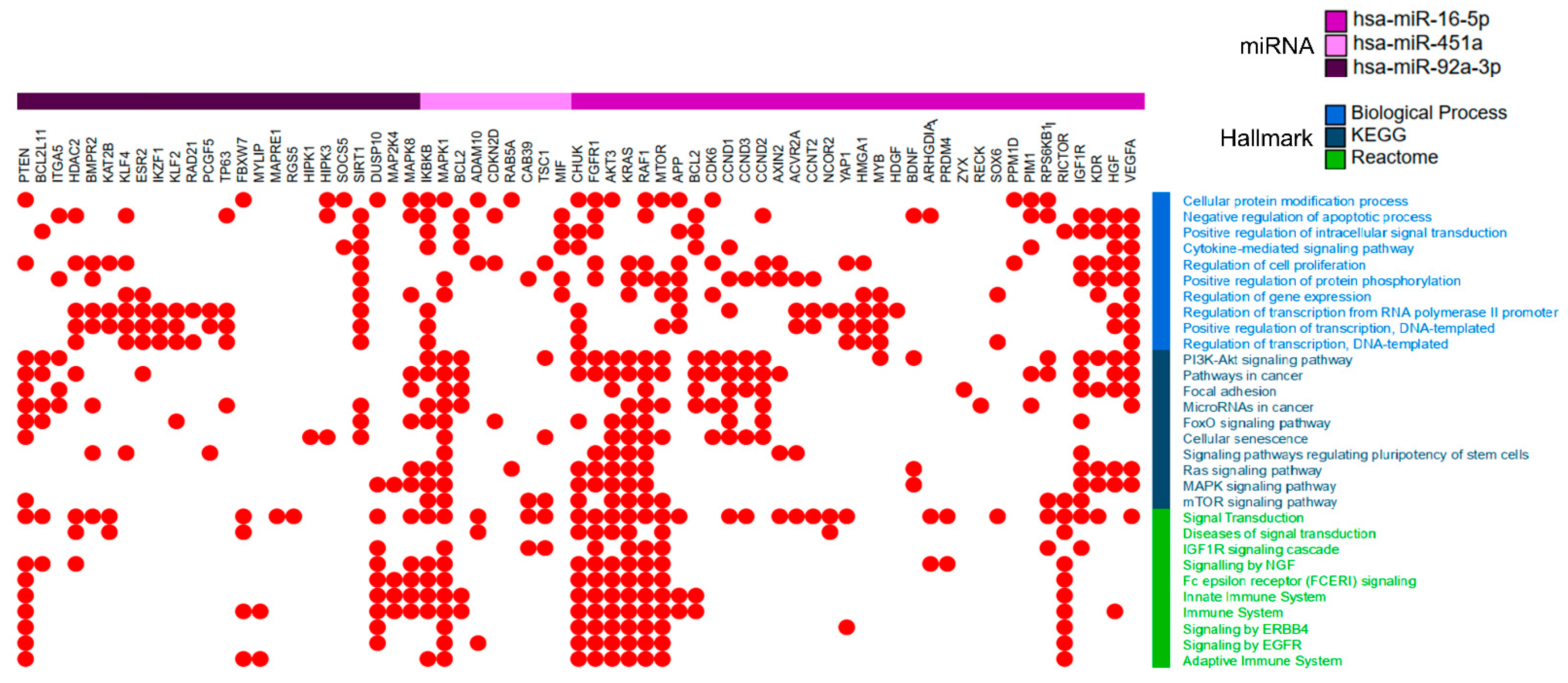
2.4. miR-16-5p, miR-92a-3p and miR-451a Target Genes are Associated with Lung Tumorigenesis Pathways
3. Discussion
4. Material and Methods
4.1. Study Population
4.2. RNA Extraction
4.3. Quantitative MiRNA Expression Analysis by Nanostring NCounter® Assay
4.4. Validation of MiRNA Expression by TaqMan® Quantitative Real-Time PCR
4.5. Statistical Analyses
4.6. Computational Data Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2018. CA. Cancer J. Clin. 2018, 68, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Krol, J.; Loedige, I.; Filipowicz, W. The widespread regulation of microRNA biogenesis, function and decay. Nat. Rev. Genet. 2010, 11, 597–610. [Google Scholar] [CrossRef] [PubMed]
- Iorio, M.V.; Croce, C.M. MicroRNA dysregulation in cancer: Diagnostics, monitoring and therapeutics. A comprehensive review. EMBO Mol. Med. 2012, 4, 143–159. [Google Scholar] [CrossRef] [PubMed]
- Schwarzenbach, H.; Nishida, N.; Calin, G.A.; Pantel, K. Clinical relevance of circulating cell-free microRNAs in cancer. Nat. Rev. Clin. Oncol. 2014, 11, 145–156. [Google Scholar] [CrossRef]
- Anfossi, S.; Fu, X.; Nagvekar, R.; Calin, G.A. MicroRNAs, regulatory messengers inside and outside cancer cells. In Advances in Experimental Medicine and Biology; Springer: New York, NY, USA, 2018; Volume 1056, pp. 87–108. [Google Scholar]
- Anfossi, S.; Babayan, A.; Pantel, K.; Calin, G.A. Clinical utility of circulating non-coding RNAs—An update. Nat. Rev. Clin. Oncol. 2018, 15, 541–563. [Google Scholar] [CrossRef]
- Wu, L.; Hu, B.; Zhao, B.; Liu, Y.; Yang, Y.; Zhang, L.; Chen, J. Circulating microRNA-422a is associated with lymphatic metastasis in lung cancer. Oncotarget 2017, 8, 42173–42188. [Google Scholar] [CrossRef] [Green Version]
- Cinegaglia, N.C.; Andrade, S.C.S.; Tokar, T.; Pinheiro, M.; Severino, F.E.; Oliveira, R.A.; Hasimoto, E.N.; Cataneo, D.C.; Cataneo, A.J.M.; Defaveri, J.; et al. Integrative transcriptome analysis identifies deregulated microRNA-transcription factor networks in lung adenocarcinoma. Oncotarget 2016, 7, 28920–28934. [Google Scholar] [CrossRef] [Green Version]
- Enfield, K.S.S.; Pikor, L.A.; Martinez, V.D.; Lam, W.L. Mechanistic Roles of Noncoding RNAs in Lung Cancer Biology and Their Clinical Implications. Genet. Res. Int. 2012, 2012, 737416. [Google Scholar] [CrossRef] [Green Version]
- Arab, A.; Karimipoor, M.; Irani, S.; Kiani, A.; Zeinali, S.; Tafsiri, E.; Sheikhy, K. Potential circulating miRNA signature for early detection of NSCLC. Cancer Genet. 2017, 216, 150–158. [Google Scholar] [CrossRef]
- Aushev, V.N.; Zborovskaya, I.B.; Laktionov, K.K.; Girard, N.; Cros, M.-P.; Herceg, Z.; Krutovskikh, V. Comparisons of microRNA Patterns in Plasma before and after Tumor Removal Reveal New Biomarkers of Lung Squamous Cell Carcinoma. PLoS ONE 2013, 8, e78649. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, F.; Nicassio, F.; Marzi, M.; Belloni, E.; Dall’Olio, V.; Bernard, L.; Pelosi, G.; Maisonneuve, P.; Veronesi, G.; Di Fiore, P.P. A serum circulating miRNA diagnostic test to identify asymptomatic high-risk individuals with early stage lung cancer. EMBO Mol. Med. 2011, 3, 495–503. [Google Scholar] [CrossRef] [PubMed]
- Boeri, M.; Verri, C.; Conte, D.; Roz, L.; Modena, P.; Facchinetti, F.; Calabrò, E.; Croce, C.M.; Pastorino, U.; Sozzi, G. MicroRNA signatures in tissues and plasma predict development and prognosis of computed tomography detected lung cancer. Proc. Natl. Acad. Sci. USA 2011, 108, 3713–3718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Hu, Z.; Wang, W.; Ba, Y.; Ma, L.; Zhang, C.; Wang, C.; Ren, Z.; Zhao, Y.; Wu, S.; et al. Identification of ten serum microRNAs from a genome-wide serum microRNA expression profile as novel noninvasive biomarkers for nonsmall cell lung cancer diagnosis. Int. J. Cancer 2012, 130, 1620–1628. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Qi, H.; Teng, J.; Su, B.; Chen, H.; Wang, C.; Xia, Q. Identification of serum miRNAs by nano-quantum dots microarray as diagnostic biomarkers for early detection of non-small cell lung cancer. Tumor Biol. 2016, 37, 7777–7784. [Google Scholar] [CrossRef]
- Halvorsen, A.R.; Bjaanæs, M.; LeBlanc, M.; Holm, A.M.; Bolstad, N.; Rubio, L.; Peñalver, J.C.; Cervera, J.; Mojarrieta, J.C.; López-Guerrero, J.A.; et al. A unique set of 6 circulating microRNAs for early detection of non-small cell lung cancer. Oncotarget 2016, 7, 37250–37259. [Google Scholar] [CrossRef] [Green Version]
- Leng, Q.; Wang, Y.; Jiang, F. A Direct Plasma miRNA Assay for Early Detection and Histological Classification of Lung Cancer. Transl. Oncol. 2018, 11, 883–889. [Google Scholar] [CrossRef] [PubMed]
- Powrózek, T.; Krawczyk, P.; Kowalski, D.M.; Kuźnar-Kamińska, B.; Winiarczyk, K.; Olszyna-Serementa, M.; Batura-Gabryel, H.; Milanowski, J. Application of plasma circulating microRNA-448, 506, 4316, and 4478 analysis for non-invasive diagnosis of lung cancer. Tumour Biol. 2016, 37, 2049–2055. [Google Scholar] [CrossRef]
- Powrózek, T.; Krawczyk, P.; Kowalski, D.M.; Winiarczyk, K.; Olszyna-Serementa, M.; Milanowski, J. Plasma circulating microRNA-944 and microRNA-3662 as potential histologic type-specific early lung cancer biomarkers. Transl. Res. 2015, 166, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Sestini, S.; Boeri, M.; Marchiano, A.; Pelosi, G.; Galeone, C.; Verri, C.; Suatoni, P.; Sverzellati, N.; La Vecchia, C.; Sozzi, G.; et al. Circulating microRNA signature as liquid-biopsy to monitor lung cancer in low-dose computed tomography screening. Oncotarget 2015, 6, 32868–32877. [Google Scholar] [CrossRef] [Green Version]
- Sozzi, G.; Boeri, M.; Rossi, M.; Verri, C.; Suatoni, P.; Bravi, F.; Roz, L.; Conte, D.; Grassi, M.; Sverzellati, N.; et al. Clinical utility of a plasma-based miRNA signature classifier within computed tomography lung cancer screening: A correlative MILD trial study. J. Clin. Oncol. 2014, 32, 768–773. [Google Scholar] [CrossRef] [PubMed]
- Su, J.; Leng, Q.; Lin, Y.; Ma, J.; Jiang, F.; Lee, C.-J.; Fang, H.; Jiang, F. Integrating Circulating Immunological and Sputum Biomarkers for the Early Detection of Lung Cancer. Biomark. Cancer 2018, 10, 1179299X18759297. [Google Scholar] [CrossRef] [PubMed]
- Su, K.; Zhang, T.; Wang, Y.; Hao, G. Diagnostic and prognostic value of plasma microRNA-195 in patients with non-small cell lung cancer. World J. Surg. Oncol. 2016, 14, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Mei, H.; Xu, C.; Tang, H.; Wei, W. Circulating microRNA-339-5p and -21 in plasma as an early detection predictors of lung adenocarcinoma. Pathol. Res. Pract. 2018, 214, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Tai, M.C.; Yanagisawa, K.; Nakatochi, M.; Hotta, N.; Hosono, Y.; Kawaguchi, K.; Naito, M.; Taniguchi, H.; Wakai, K.; Yokoi, K.; et al. Blood-borne miRNA profile-based diagnostic classifier for lung adenocarcinoma. Sci. Rep. 2016, 6, 31389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, J.; Gao, W.; Zhu, C.J.; Liu, Y.Q.; Mei, Z.; Cheng, T.; Shu, Y.Q. Identification of plasma microRNA-21 as a biomarker for early detection and chemosensitivity of non-small cell lung cancer. Chin. J. Cancer 2011, 30, 407–414. [Google Scholar] [CrossRef] [Green Version]
- Wozniak, M.B.; Scelo, G.; Muller, D.C.; Mukeria, A.; Zaridze, D.; Brennan, P. Circulating MicroRNAs as Non-Invasive Biomarkers for Early Detection of Non-Small-Cell Lung Cancer. PLoS ONE 2015, 10, e0125026. [Google Scholar] [CrossRef] [Green Version]
- Zheng, D.; Haddadin, S.; Wang, Y.; Gu, L.Q.; Perry, M.C.; Freter, C.E.; Wang, M.X. Plasma micrornas as novel biomarkers for early detection of lung cancer. Int. J. Clin. Exp. Pathol. 2011, 4, 575–586. [Google Scholar]
- Zou, J.G.; Ma, L.F.; Li, X.; Xu, F.L.; Fei, X.Z.; Liu, Q.; Bai, Q.L.; Dong, Y.L. Circulating microRNA array (miR-182, 200b and 205) for the early diagnosis and poor prognosis predictor of non-small cell lung cancer. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 1108–1115. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Tokar, T.; Pastrello, C.; Rossos, A.E.M.; Abovsky, M.; Hauschild, A.C.; Tsay, M.; Lu, R.; Jurisica, I. mirDIP 4.1-integrative database of human microRNA target predictions. Nucleic Acids Res. 2018, 46, D360–D370. [Google Scholar] [CrossRef] [PubMed]
- Lonsdale, J.; Thomas, J.; Salvatore, M.; Phillips, R.; Lo, E.; Shad, S.; Hasz, R.; Walters, G.; Garcia, F.; Young, N.; et al. The Genotype-Tissue Expression (GTEx) project. Nat. Genet. 2013, 45, 580–585. [Google Scholar] [CrossRef] [PubMed]
- Ow, G.S.; Kuznetsov, V.A. Multiple signatures of a disease in potential biomarker space: Getting the signatures consensus and identification of novel biomarkers. BMC Genomics 2015, 16, S2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hennessey, P.T.; Sanford, T.; Choudhary, A.; Mydlarz, W.W.; Brown, D.; Adai, A.T.; Ochs, M.F.; Ahrendt, S.A.; Mambo, E.; Califano, J.A. Serum microrna biomarkers for detection of non-small cell lung cancer. PLoS ONE 2012, 7, e32307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montani, F.; Jacopo Marzi, M.; Dezi, F.; Dama, E.; Mary Carletti, R.; Bonizzi, G.; Bertolotti, R.; Bellomi, M.; Rampinelli, C.; Maisonneuve, P.; et al. miR-Test: A Blood Test for Lung Cancer Early Detection. J. Natl. Cancer Inst. 2015, 107, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marzi, M.J.; Montani, F.; Carletti, R.M.; Dezi, F.; Dama, E.; Bonizzi, G.; Sandri, M.T.; Rampinelli, C.; Bellomi, M.; Maisonneuve, P.; et al. Optimization and standardization of circulating MicroRNA detection for clinical application: The miR-test case. Clin. Chem. 2016, 62, 743–754. [Google Scholar] [CrossRef]
- Geiss, G.K.; Bumgarner, R.E.; Birditt, B.; Dahl, T.; Dowidar, N.; Dunaway, D.L.; Fell, H.P.; Ferree, S.; George, R.D.; Grogan, T.; et al. Direct multiplexed measurement of gene expression with color-coded probe pairs. Nat. Biotechnol. 2008, 26, 317–325. [Google Scholar] [CrossRef]
- Reis, P.P.; Waldron, L.; Perez-Ordonez, B.; Pintilie, M.; Galloni, N.N.; Xuan, Y.; Cervigne, N.K.; Warner, G.C.; Makitie, A.A.; Simpson, C.; et al. A gene signature in histologically normal surgical margins is predictive of oral carcinoma recurrence. BMC Cancer 2011, 11, 437. [Google Scholar] [CrossRef] [Green Version]
- Veldman-Jones, M.H.; Brant, R.; Rooney, C.; Geh, C.; Emery, H.; Harbron, C.G.; Wappett, M.; Sharpe, A.; Dymond, M.; Barrett, J.C.; et al. Evaluating robustness and sensitivity of the nanostring technologies ncounter platform to enable multiplexed gene expression analysis of clinical samples. Cancer Res. 2015, 75, 2587–2593. [Google Scholar] [CrossRef] [Green Version]
- Reis, P.P.; Tokar, T.; Goswami, R.S.; Xuan, Y.; Sukhai, M.; Seneda, A.L.; Móz, L.E.S.; Perez-Ordonez, B.; Simpson, C.; Goldstein, D.; et al. A 4-gene signature from histologically normal surgical margins predicts local recurrence in patients with oral carcinoma: Clinical validation. Sci. Rep. 2020, 10, 1–8. [Google Scholar] [CrossRef]
- Stewart, R.L.; Updike, K.L.; Factor, R.E.; Henry, N.L.; Boucher, K.M.; Bernard, P.S.; Varley, K.E. A multigene assay determines risk of recurrence in patients with triple-negative breast cancer. Cancer Res. 2019, 79, 3466–3478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ell, B.; Mercatali, L.; Ibrahim, T.; Campbell, N.; Schwarzenbach, H.; Pantel, K.; Amadori, D.; Kang, Y. Tumor-Induced Osteoclast miRNA Changes as Regulators and Biomarkers of Osteolytic Bone Metastasis. Cancer Cell 2013, 24, 542–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stückrath, I.; Rack, B.; Janni, W.; Jäger, B.; Pantel, K.; Schwarzenbach, H. Aberrant plasma levels of circulating miR-16, miR-107, miR-130a and miR-146a are associated with lymph node metastasis and receptor status of breast cancer patients. Oncotarget 2015, 6, 13387–13401. [Google Scholar] [CrossRef] [PubMed]
- Schwarzenbach, H.; Machado Da Silva, A.; Calin, G.; Pantel, K. Which is the accurate data normalization strategy for microRNA quantification? Clin. Chem. 2015, 61, 1333–1342. [Google Scholar] [CrossRef] [PubMed]
- Quintanal-Villalonga, A.; Molina-Pinelo, S.; Cirauqui, C.; Ojeda-Márquez, L.; Marrugal, Á.; Suarez, R.; Conde, E.; Ponce-Aix, S.; Enguita, A.B.; Carnero, A.; et al. FGFR1 Cooperates with EGFR in Lung Cancer Oncogenesis, and Their Combined Inhibition Shows Improved Efficacy. J. Thorac. Oncol. 2019, 14, 641–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, C.; Shan, Z.; Hong, J.; Yang, L. MicroRNA-92a promotes epithelial-mesenchymal transition through activation of PTEN/PI3K/AKT signaling pathway in non-small cell lung cancer metastasis. Int. J. Oncol. 2017, 51, 235–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choudhry, Z.; Tripathi, V.; Sutton, M.; Bao, B.; Mohammad, R.; Azmi, A. Regulation of KRAS-PAK4 Axis by MicroRNAs in Cancer. Curr. Pharm. Des. 2014, 20, 5275–5278. [Google Scholar] [CrossRef]
- Ma, Y.; Pan, X.; Xu, P.; Mi, Y.; Wang, W.; Wu, X.; He, Q.; Liu, X.; Tang, W.; An, H.X. Plasma microRNA alterations between EGFR-activating mutational NSCLC patients with and without primary resistance to TKI. Oncotarget 2017, 8, 88529–88536. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Zhang, Q.; Zhang, M.; Su, W.; Wang, Z.; Li, Y.; Zhang, J.; Beer, D.G.; Yang, S.; Chen, G. Serum microRNA signature is capable of early diagnosis for non-small cell lung cancer. Int. J. Biol. Sci. 2019, 15, 1712–1722. [Google Scholar] [CrossRef] [Green Version]
- Menezes, R.J.; Roberts, H.C.; Paul, N.S.; McGregor, M.; Chung, T.B.; Patsios, D.; Weisbrod, G.; Herman, S.; Pereira, A.; McGregor, A.; et al. Lung cancer screening using low-dose computed tomography in at-risk individuals: The Toronto experience. Lung Cancer 2010, 67, 177–183. [Google Scholar] [CrossRef]
- McWilliams, A.; Tammemagi, M.C.; Mayo, J.R.; Roberts, H.; Liu, G.; Soghrati, K.; Yasufuku, K.; Martel, S.; Laberge, F.; Gingras, M.; et al. Probability of cancer in pulmonary nodules detected on first screening CT. N. Engl. J. Med. 2013, 369, 910–919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, T.C.; Coenen-Stass, A.M.L.; Wood, M.J.A. Assessment of RT-qPCR normalization strategies for accurate quantification of extracellular microRNAs in murine Serum. PLoS ONE 2014, 9, e89237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarzenbach, H.; Da Silva, A.M.; Calin, G.; Pantel, K. Data normalization strategies for microRNA quantification. Clin. Chem. 2015, 61, 1333–1342. [Google Scholar] [CrossRef]
- Griffiths-Jones, S. miRBase: microRNA sequences, targets and gene nomenclature. Nucleic Acids Res. 2006, 34, D140–D144. [Google Scholar] [CrossRef] [PubMed]
- Kozomara, A.; Griffiths-Jones, S. miRBase: Integrating microRNA annotation and deep-sequencing data. Nucleic Acids Res. 2011, 39, D152–D157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozomara, A.; Griffiths-Jones, S. miRBase: Annotating high confidence microRNAs using deep sequencing data. Nucleic Acids Res. 2014, 42, D68–D73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wahid, F.; Shehzad, A.; Khan, T.; Kim, Y.Y. MicroRNAs: Synthesis, mechanism, function, and recent clinical trials. Biochim. Biophys. Acta-Mol. Cell Res. 2010, 1803, 1231–1243. [Google Scholar] [CrossRef] [Green Version]
- Sundarbose, K.; Kartha, R.; Subramanian, S. MicroRNAs as Biomarkers in Cancer. Diagnostics 2013, 3, 84–104. [Google Scholar] [CrossRef] [Green Version]
- Vigneron, N.; Meryet-Figuière, M.; Guttin, A.; Issartel, J.P.; Lambert, B.; Briand, M.; Louis, M.H.; Vernon, M.; Lebailly, P.; Lecluse, Y.; et al. Towards a new standardized method for circulating miRNAs profiling in clinical studies: Interest of the exogenous normalization to improve miRNA signature accuracy. Mol. Oncol. 2016, 10, 981–992. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling The False Discovery Rate - A Practical And Powerful Approach To Multiple Testing. J. R. Stat. Soc. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Benjamini, Y.; Yekutieli, D. The control of the false discovery rate in multiple testing under dependency. Ann. Stat. 2001, 29, 1165–1188. [Google Scholar]
- Friedman, J.; Hastie, T.; Tibshirani, R. Regularization Paths for Generalized Linear Models via Coordinate Descent. J. Stat. Softw. 2010, 33, 1–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simon, N.; Friedman, J.; Hastie, T.; Tibshirani, R. Regularization Paths for Cox’s Proportional Hazards Model via Coordinate Descent. J. Stat. Softw. 2011, 39, 1. [Google Scholar] [CrossRef] [PubMed]
- Tibshirani, R.; Bien, J.; Friedman, J.; Hastie, T.; Simon, N.; Taylor, J.; Tibshirani, R.J. Strong rules for discarding predictors in lasso-type problems. J. R. Stat. Soc. Ser. B Stat. Methodol. 2012, 74, 245–266. [Google Scholar] [CrossRef]
- Available online: http://web.stanford.edu/~hastie/glmnet_matlab/; https://web.stanford.edu/~hastie/Papers/Glmnet_Vignette.pdf (accessed on 19 May 2017).
- Zhu, C.-Q.; Strumpf, D.; Li, C.-Y.; Li, Q.; Liu, N.; Der, S.; Shepherd, F.A.; Tsao, M.-S.; Jurisica, I. Prognostic Gene Expression Signature for Squamous Cell Carcinoma of Lung. Clin. Cancer Res. 2010, 16, 5038–5047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, C.-Q.; Ding, K.; Strumpf, D.; Weir, B.A.; Meyerson, M.; Pennell, N.; Thomas, R.K.; Naoki, K.; Ladd-Acosta, C.; Liu, N.; et al. Prognostic and predictive gene signature for adjuvant chemotherapy in resected non-small-cell lung cancer. J. Clin. Oncol. 2010, 28, 4417–4424. [Google Scholar] [CrossRef]
- Bardou, P.; Mariette, J.; Escudié, F.; Djemiel, C.; Klopp, C. Jvenn: An interactive Venn diagram viewer. BMC Bioinforma. 2014, 15, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Chou, C.H.; Shrestha, S.; Yang, C.D.; Chang, N.W.; Lin, Y.L.; Liao, K.W.; Huang, W.C.; Sun, T.H.; Tu, S.J.; Lee, W.H.; et al. miRTarBase update 2018: A resource for experimentally validated microRNA-target interactions. Nucleic Acids Res. 2018, 46, D296–D302. [Google Scholar] [CrossRef]
- Raphael, B.J.; Hruban, R.H.; Aguirre, A.J.; Moffitt, R.A.; Yeh, J.J.; Stewart, C.; Robertson, A.G.; Cherniack, A.D.; Gupta, M.; Getz, G.; et al. Integrated Genomic Characterization of Pancreatic Ductal Adenocarcinoma. Cancer Cell 2017, 32, 185–203. [Google Scholar] [CrossRef]
- Fernandez, N.F.; Gundersen, G.W.; Rahman, A.; Grimes, M.L.; Rikova, K.; Hornbeck, P.; Maayan, A. Clustergrammer, a web-based heatmap visualization and analysis tool for high-dimensional biological data. Sci. Data 2017, 4, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Aran, D.; Sirota, M.; Butte, A.J. Systematic pan-cancer analysis of tumour purity. Nat. Commun. 2015, 6, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aran, D.; Camarda, R.; Odegaard, J.; Paik, H.; Oskotsky, B.; Krings, G.; Goga, A.; Sirota, M.; Butte, A.J. Comprehensive analysis of normal adjacent to tumor transcriptomes. Nat. Commun. 2017, 8, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Oliveira, G.; Paccielli Freire, P.; Santiloni Cury, S.; de Moraes, D.; Santos Oliveira, J.; Dal-Pai-Silva, M.; do Reis, P.P.; Francisco Carvalho, R. An Integrated Meta-Analysis of Secretome and Proteome Identify Potential Biomarkers of Pancreatic Ductal Adenocarcinoma. Cancers 2020, 12, 716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNA (Ranked by p-Value) | Patients (Cases) | Healthy (Controls) | FC | p Value of FC | FDR |
|---|---|---|---|---|---|
| Lung adenocarcinoma (LUAD) | |||||
| miR-16-5p | 1852.9 | 381.9 | 4.85 | 2.563 × 10−6 | 0.002 |
| miR-451a | 14,300.9 | 2518.2 | 5.68 | 2.319 × 10−4 | 0.035 |
| miR-92a-3p | 528.4 | 154.4 | 3.42 | 4.203 × 10−4 | 0.035 |
| miR-25-3p | 408.4 | 87.5 | 4.67 | 1.508 × 10−3 | 0.035 |
| miR-494-3p | 4293.8 | 327.5 | 13.11 | 2.918 × 10−3 | 0.035 |
| miR-1285-5p | 55.4 | 5.6 | 9.79 | 2.918 × 10−3 | 0.035 |
| miR-125b-5p | 42.0 | 8.5 | 4.90 | 2.918 × 10−3 | 0.035 |
| miR-448 | 16.0 | 1.3 | 11.73 | 3.587 × 10−3 | 0.035 |
| miR-155-5p | 112.5 | 15.1 | 7.45 | 3.587 × 10−3 | 0.035 |
| miR-2682-5p | 93.2 | 9.8 | 9.43 | 4.377 × 10−3 | 0.035 |
| Lung squamous cell carcinoma (LUSC) | |||||
| miR-16-5p | 2043.0 | 381.9 | 5.35 | 2.447 × 10−4 | 0.153 |
| miR-451a | 17,129.5 | 2518.2 | 6.80 | 2.676 × 10−3 | 0.153 |
| miR-92a-3p | 394.3 | 154.4 | 2.55 | 3.276 × 10−3 | 0.153 |
| miR-25-3p | 499.4 | 87.5 | 5.71 | 4.056 × 10−3 | 0.153 |
| miR-149-5p | 141.5 | 16.3 | 8.68 | 4.601 × 10−3 | 0.153 |
| miR-548ah-5p | 45.7 | 6.1 | 7.50 | 4.601 × 10−3 | 0.153 |
| miR-1285-5p | 36.3 | 5.6 | 6.42 | 4.601 × 10−3 | 0.153 |
| miR-155-5p | 89.4 | 15.1 | 5.92 | 4.601 × 10−3 | 0.153 |
| miR-3168 | 12.1 | 2.2 | 5.38 | 4.601 × 10−3 | 0.153 |
| miR-575 | 17.5 | 1.6 | 10.99 | 5.939 × 10−3 | 0.153 |
| MiRNA | Patients (Cases) | Healthy (Controls) | FC | p Value of FC | C-Statistics | Combined Score |
|---|---|---|---|---|---|---|
| miR-16-5p | 1886.09 | 381.98 | 4.94 | 1.98 × 10−6 | 0.97 | 2.00 |
| miR-451a | 14,890.94 | 2518.28 | 5.91 | 0.00018 | 0.91 | 2.33 |
| miR-1285-5p | 48.61 | 5.66 | 8.59 | 0.0013 | 0.86 | 3.00 |
| miR-92a-3p | 516.32 | 154.48 | 3.34 | 0.0007 | 0.91 | 3.67 |
| miR-155-5p | 108.73 | 15.12 | 7.19 | 0.0015 | 0.86 | 4.00 |
| miR-217 | 3.55 | 1.4 | 2.53 | 0.04 | 0.74 | 6.00 |
| miR-378e | 327.84 | 146.72 | 2.23 | 0.19 | 0.65 | 7.00 |
| miR-1285-3p | 6.6 | 3.61 | 1.83 | 0.54 | 0.57 | 8.33 |
| miR-106b-5p | 40.35 | 36.43 | 1.11 | 0.47 | 0.41 | 9.67 |
| miR-664a-3p | 10.01 | 9.2 | 1.08 | 0.84 | 0.53 | 9.67 |
| miR-484 | 26.6 | 25.81 | 1.03 | 0.90 | 0.52 | 11.00 |
| miR-148b-3p | 39.05 | 37.71 | 1.03 | 0.98 | 0.49 | 11.33 |
| MiRNAs | Patients (Cases) | Healthy (Control) | FC | C-Statistics | p Value of FC |
|---|---|---|---|---|---|
| Signatures | |||||
| C-Statistics | 0.0548 | 0.0155 | 0.727 | 0.00019 | |
| Elastic net | 5.4338 | 2.7423 | 0.644 | 0.013 | |
| MARSA | 0.0142 | 0.0042 | 0.644 | 0.013 | |
| Individual miRNAs | |||||
| miR-16-5p | 2.205 | 0.8973 | 2.0 | 0.676 | 0.0031 |
| miR-92a-3p | 1.9971 | 0.9947 | 2.4 | 0.653 | 0.009 |
| miR-451a | 1.4331 | 1.0229 | 1.8 | 0.629 | 0.024 |
| miR-378e | 5.8682 | 1.132 | 5.1 | 0.726 | 0.0002 |
| miR-1285-3p | 4.8923 | 0.6846 | 7.9 | 0.69 | 0.0016 |
| miR-1285-5p | 2.3703 | 1.0443 | 2.7 | 0.701 | 0.00086 |
| miR-217 | 6.2591 | 1.2573 | 9.5 | 0.766 | 0.000013 |
| miR-106b-5p | 2.5163 | 1.134 | 1.7 | 0.651 | 0.0098 |
| miR-148b-3p | 3.8442 | 0.9845 | 3.8 | 0.661 | 0.0063 |
| miR-484 | 4.2059 | 1.7054 | 2.6 | 0.735 | 0.00011 |
| miR-664a-3p | 8.3679 | 1.185 | 4.7 | 0.739 | 0.000084 |
| miR-155-5p | 1.1978 | 0.8908 | 0.8 | 0.508 | 0.45 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reis, P.P.; Drigo, S.A.; Carvalho, R.F.; Lopez Lapa, R.M.; Felix, T.F.; Patel, D.; Cheng, D.; Pintilie, M.; Liu, G.; Tsao, M.-S. Circulating miR-16-5p, miR-92a-3p, and miR-451a in Plasma from Lung Cancer Patients: Potential Application in Early Detection and a Regulatory Role in Tumorigenesis Pathways. Cancers 2020, 12, 2071. https://doi.org/10.3390/cancers12082071
Reis PP, Drigo SA, Carvalho RF, Lopez Lapa RM, Felix TF, Patel D, Cheng D, Pintilie M, Liu G, Tsao M-S. Circulating miR-16-5p, miR-92a-3p, and miR-451a in Plasma from Lung Cancer Patients: Potential Application in Early Detection and a Regulatory Role in Tumorigenesis Pathways. Cancers. 2020; 12(8):2071. https://doi.org/10.3390/cancers12082071
Chicago/Turabian StyleReis, Patricia P., Sandra A. Drigo, Robson F. Carvalho, Rainer Marco Lopez Lapa, Tainara F. Felix, Devalben Patel, Dangxiao Cheng, Melania Pintilie, Geoffrey Liu, and Ming-Sound Tsao. 2020. "Circulating miR-16-5p, miR-92a-3p, and miR-451a in Plasma from Lung Cancer Patients: Potential Application in Early Detection and a Regulatory Role in Tumorigenesis Pathways" Cancers 12, no. 8: 2071. https://doi.org/10.3390/cancers12082071