Molecular Characterisation of Canine Osteosarcoma in High Risk Breeds

 , , and
, , and
Abstract
:1. Introduction
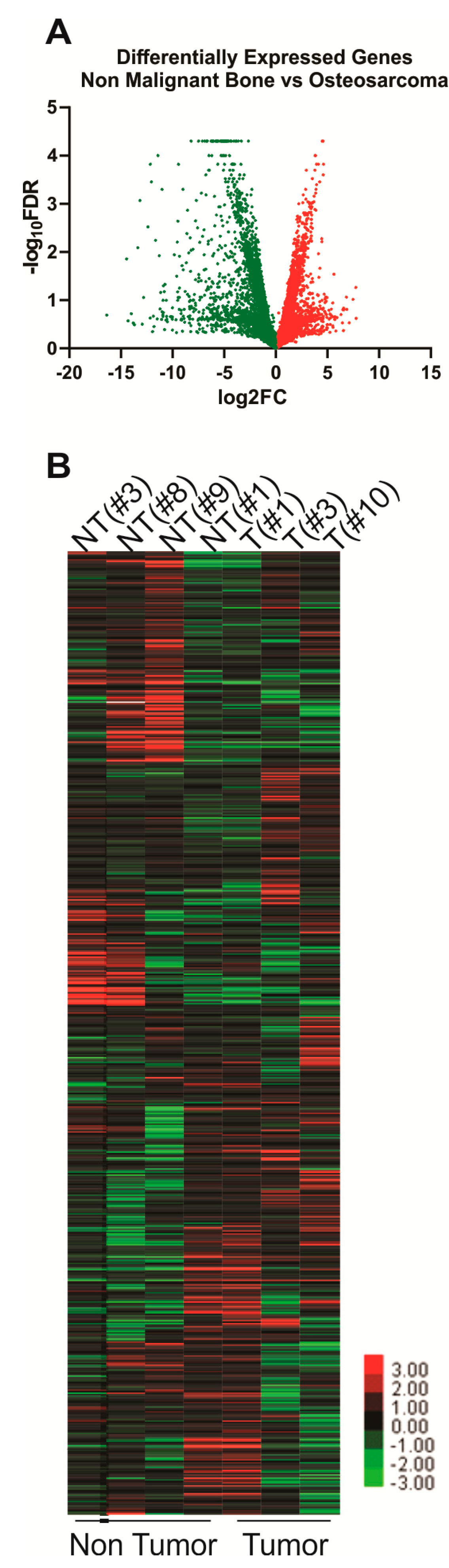
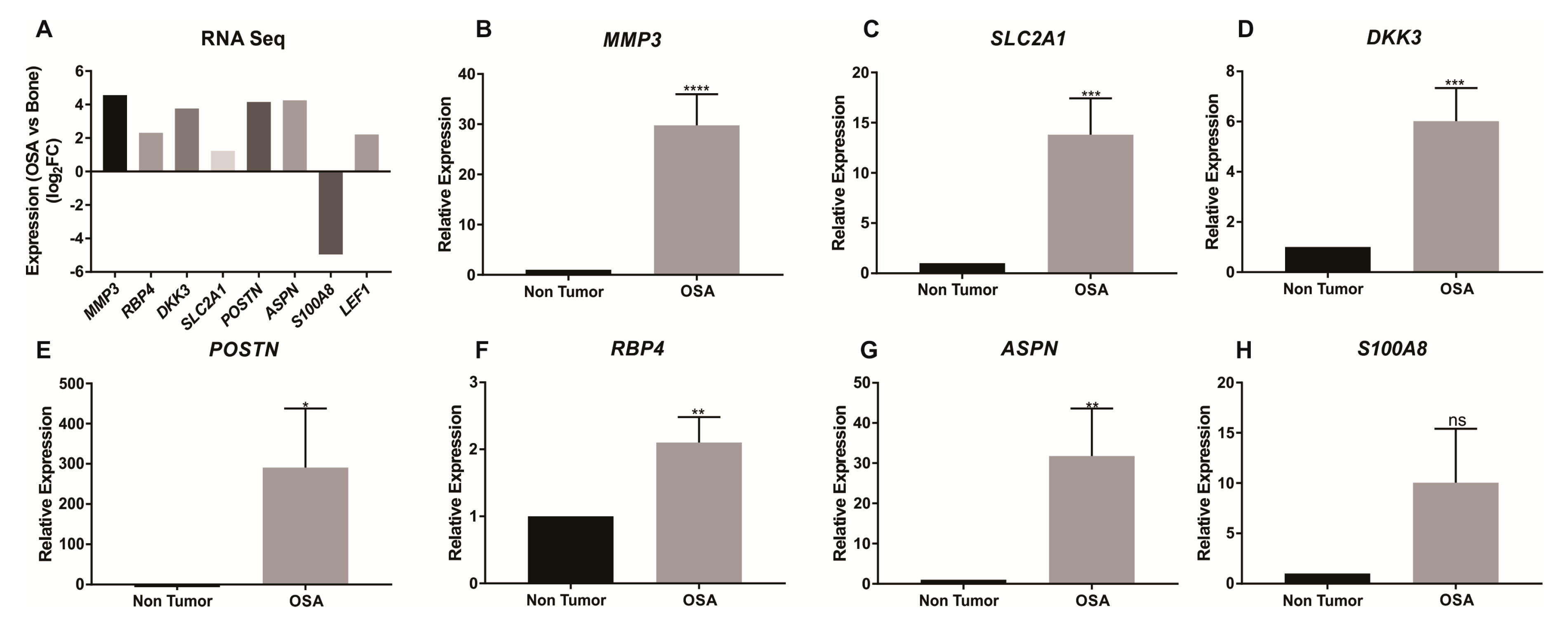
2. Results
2.1. RNAseq Gene Expression
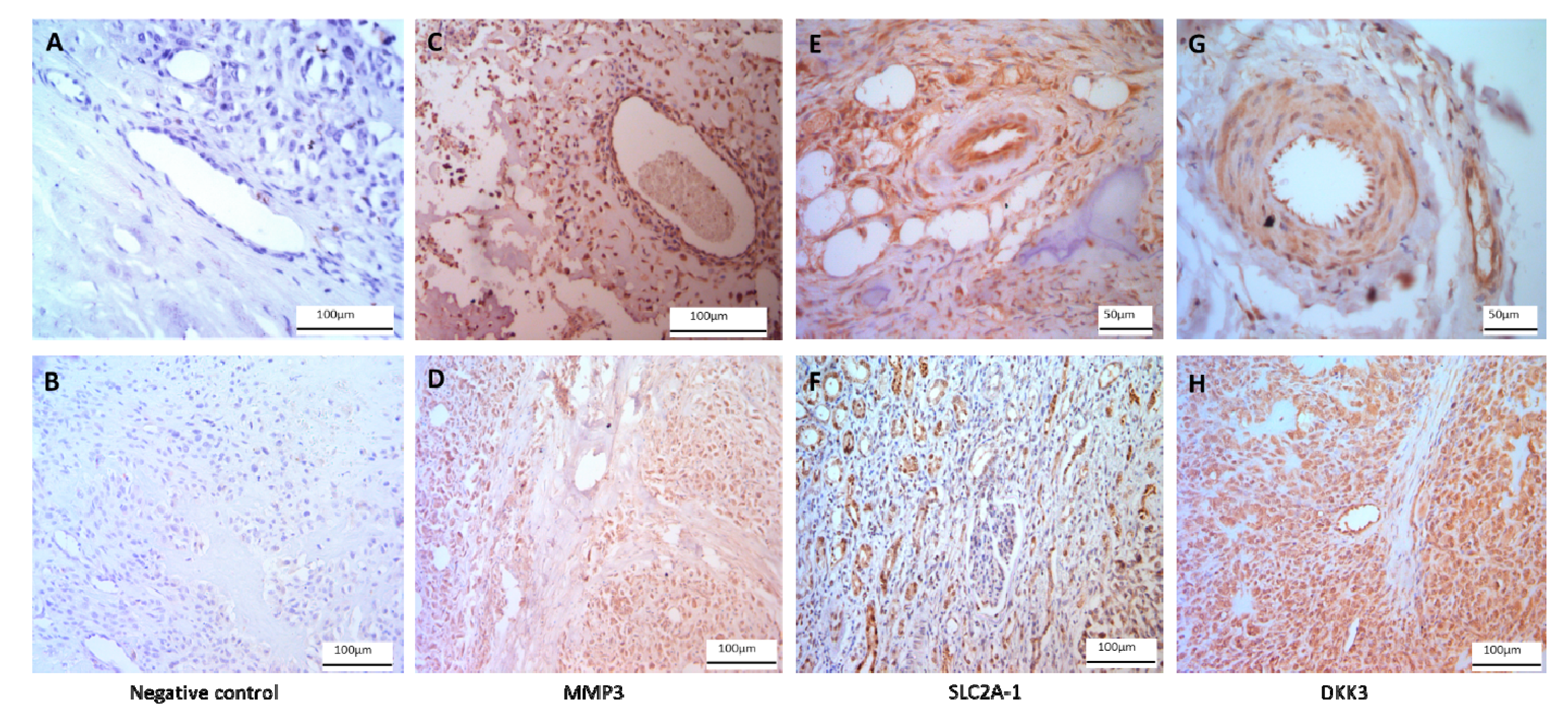
2.2. qRT-PCR and Immunohistochemistry Validation
2.3. Cohort Epidemiology
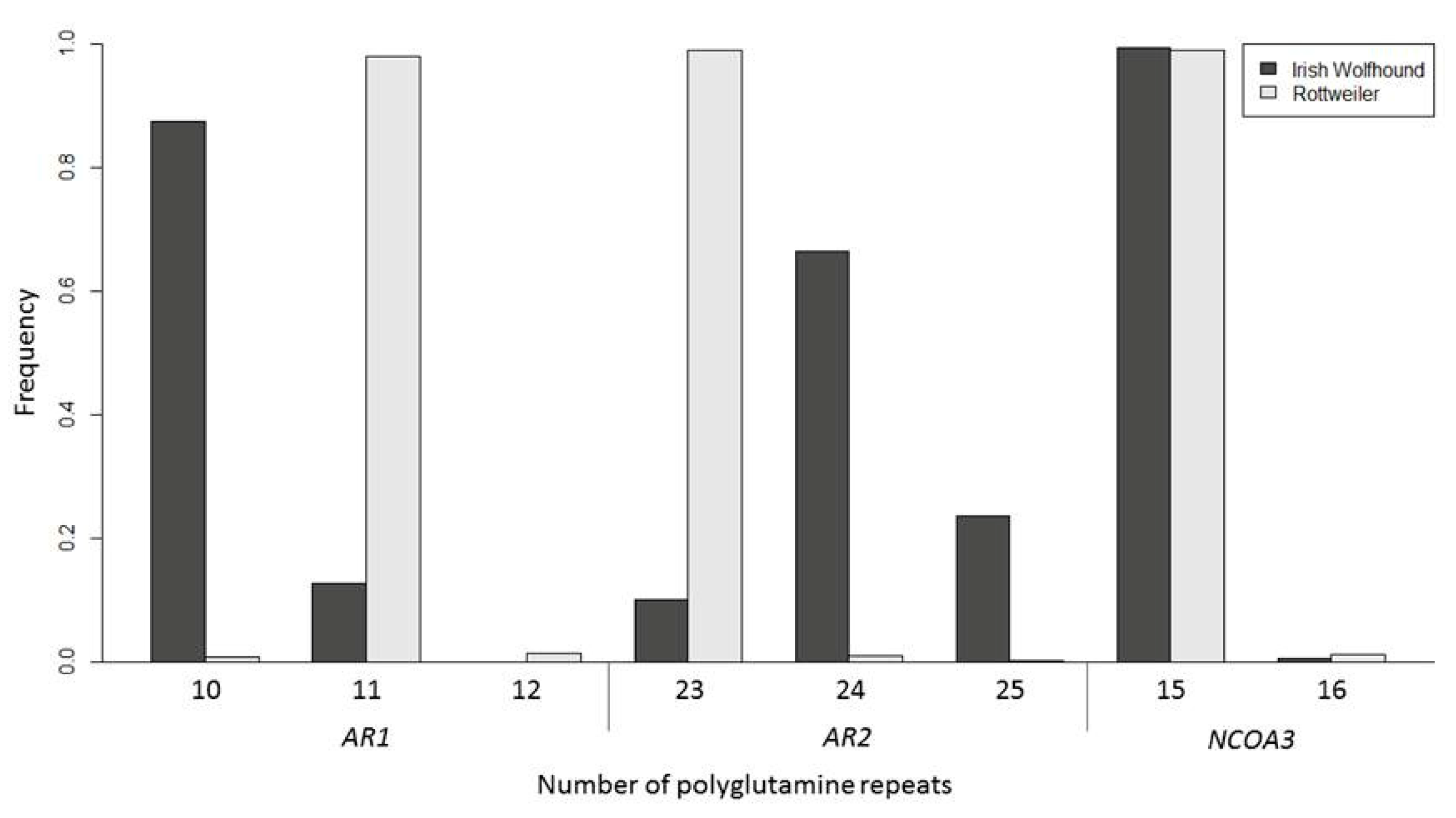
2.4. Androgen polyQ Analysis
3. Discussion
4. Materials and Methods
4.1. Samples and Ethics
4.2. RNA Extraction and Next Generation Sequencing
4.3. Quantitative Reverse Transcriptase PCR
4.4. Immunohistochemistry
4.5. PolyQ Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mirabello, L.; Troisi, R.J.; Savage, S.A. International osteosarcoma incidence patterns in children and adolescents, middle ages and elderly persons. Int. J. Cancer 2009, 125, 229–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siegel, R.; Naishadham, D. jemal A: Cancer statistics, 2013. CA Cancer J. Clin. 2013, 63, 11–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirabello, L.; Troisi, R.J.; Savage, S.A. Osteosarcoma incidence and survival rates from 1973 to 2004: Data from the Surveillance, Epidemiology, and End Results Program. Cancer 2009, 115, 1531–1543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fenger, J.M.; London, C.A.; Kisseberth, W.C. Canine Osteosarcoma: A Naturally Occurring Disease to Inform Pediatric Oncology. ILAR J. 2014, 55, 69–85. [Google Scholar] [CrossRef] [Green Version]
- Tinkey, P.T.; Lembo, T.M.; Evans, G.R.D.; Cundiff, J.H.; Gray, K.N.; Price, R.E. Postirradiation sarcomas in Sprague-Dawley rats. Radiat. Res. 1998, 149, 401–404. [Google Scholar] [CrossRef]
- Jacks, T.; Remington, L.; Williams, B.O.; Schmitt, E.M.; Halachmi, S.; Bronson, R.T.; Weinberg, R.A. Tumor spectrum analysis in p53-mutant mice. Curr. Biol. 1994, 4, 1–7. [Google Scholar] [CrossRef]
- Berlin, O.; Samid, D.; Donthineni-Rao, R.; Akeson, W.; Amiel, D.; Woods, V.L. Development of a novel spontaneous metastasis model of human osteosarcoma transplanted orthotopically into bone of athymic mice. Cancer Res. 1993, 53, 4890–4895. [Google Scholar]
- Parkin, D.M.; Ferlay, J.; Bray, F.; Edwards, B.; Forman, D.; Curado, M.P.; Shin, H.-R. Fifty years of cancer incidence: CI5 I-IX. Int. J. Cancer 2010, 127, 2918–2927. [Google Scholar] [CrossRef]
- Egenvall, A.; Nødtvedt, A.; Von Euler, H. Bone tumors in a population of 400 000 insured Swedish dogs up to 10 y of age: Incidence and survival. Can. J. Vet. Res. 2007, 71, 292–299. [Google Scholar]
- Frimberger, A.E.; Chan, C.M.; Moore, A.S. Canine Osteosarcoma Treated by Post-Amputation Sequential Accelerated Doxorubicin and Carboplatin Chemotherapy: 38 Cases. J. Am. Anim. Hosp. Assoc. 2016, 52, 149–156. [Google Scholar] [CrossRef]
- Moore, A.S.; Dernell, W.S.; Ogilvie, G.K.; Kristal, O.; Elmslie, R.; Kitchell, B.; Susaneck, S.; Rosenthal, R.; Klein, M.K.; Obradovoich, J.; et al. Doxorubicin and BAY 12–9566 for the Treatment of Osteosarcoma in Dogs: A Randomized, Double-Blind, Placebo-Controlled Study. J. Vet. Intern. Med. 2007, 21, 783–790. [Google Scholar] [CrossRef] [PubMed]
- Straw, R.C.; Withrow, S.J.; Richter, S.L.; Powers, B.E.; Klein, M.K.; Postorino, N.C.; LaRue, S.M.; Ogilvie, G.K.; Vail, D.M.; Morrison, W.B.; et al. Amputation and Cisplatin for Treatment of Canine Osteosarcoma. J. Vet. Intern. Med. 1991, 5, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Savage, S.A.; Mirabello, L.; Wang, Z.; Gastier-Foster, J.M.; Gorlick, R.; Khanna, C.; Flanagan, A.M.; Tirabosco, R.; Andrulis, I.L.; Wunder, J.S.; et al. Genome-wide association study identifies two susceptibility loci for osteosarcoma. Nat. Genet. 2013, 45, 799–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirabello, L.; Pfeiffer, R.; Murphy, G.; Daw, N.C.; Patiño-García, A.; Troisi, R.J.; Hoover, R.N.; Douglass, C.; Schüz, J.; Craft, A.W.; et al. Height at diagnosis and birth-weight as risk factors for osteosarcoma. Cancer Causes Control. 2011, 22, 899–908. [Google Scholar] [CrossRef] [Green Version]
- McNeill, C.J.; Overley, B.; Shofer, F.S.; Kent, M.S.; Clifford, C.A.; Samluk, M.; Haney, S.; Van Winkle, T.J.; Sorenmo, K.U. Characterization of the biological behaviour of appendicular osteosarcoma in Rottweilers and a comparison with other breeds: A review of 258 dogs. Vet. Comp. Oncol. 2007, 5, 90–98. [Google Scholar] [CrossRef]
- Ru, G.; Terracini, B.; Glickman, L. Host related risk factors for canine osteosarcoma. Vet. J. 1998, 156, 31–39. [Google Scholar] [CrossRef]
- Yang, Y.C.; Banuelos, C.A.; Mawji, N.R.; Wang, J.; Kato, M.; Haile, S.; McEwan, I.J.; Plymate, S.; Sadar, M.D. Targeting Androgen Receptor Activation Function-1 with EPI to Overcome Resistance Mechanisms in Castration-Resistant Prostate Cancer. Clin. Cancer Res. 2016, 22, 4466–4477. [Google Scholar] [CrossRef] [Green Version]
- Mongan, N.P.; Tadokoro-Cuccaro, R.; Bunch, T.; Hughes, I.A. Androgen insensitivity syndrome. Best Pract. Res. Clin. Endocrinol. Metab. 2015, 29, 569–580. [Google Scholar] [CrossRef]
- Lu, N.Z.; Wardell, S.E.; Burnstein, K.L.; DeFranco, D.; Fuller, P.J.; Giguère, V.; Hochberg, R.B.; McKay, L.; Renoir, J.-M.; Weigel, N.L.; et al. International Union of Pharmacology. LXV. The Pharmacology and Classification of the Nuclear Receptor Superfamily: Glucocorticoid, Mineralocorticoid, Progesterone, and Androgen Receptors. Pharmacol. Rev. 2006, 58, 782–797. [Google Scholar] [CrossRef]
- Fang, H.; Tong, W.; Branham, W.S.; Moland, C.L.; Dial, S.L.; Hong, H.; Xie, Q.; Perkins, R.; Owens, W.; Sheehan, D.M. Study of 202 Natural, Synthetic, and Environmental Chemicals for Binding to the Androgen Receptor. Chem. Res. Toxicol. 2003, 16, 1338–1358. [Google Scholar] [CrossRef]
- McKenna, N.J.; O’Malley, B.W. Combinatorial Control of Gene Expression by Nuclear Receptors and Coregulators. Cell 2002, 108, 465–474. [Google Scholar] [CrossRef] [Green Version]
- Dart, D.A.; Waxman, J.; Aboagye, E.O.; Bevan, C.L. Visualising Androgen Receptor Activity in Male and Female Mice. PLoS ONE 2013, 8, e71694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Infante, J.B.; Alvelos, M.I.; Bastos, M.; Carrilho, F.; Lemos, M.C. Complete androgen insensitivity syndrome caused by a novel splice donor site mutation and activation of a cryptic splice donor site in the androgen receptor gene. J. Steroid Biochem. Mol. Biol. 2016, 155, 63–66. [Google Scholar] [CrossRef] [PubMed]
- Gottlieb, B.; Beitel, L.K.; Nadarajah, A.; Paliouras, M.; Trifiro, M. The androgen receptor gene mutations database: 2012 update. Hum. Mutat. 2012, 33, 887–894. [Google Scholar] [CrossRef]
- Batch, J.A.; Williams, D.M.; Davies, H.R.; Brown, B.D.; Evans, B.A.J.; Hughes, I.A.; Patterson, M.N. Androgen receptor gene mutations identified by SSCP in fourteen subjects with androgen insensitivity syndrome. Hum. Mol. Genet. 1992, 1, 497–503. [Google Scholar] [CrossRef]
- Brown, T.R.; Lubahn, D.B.; Wilson, E.M.; Joseph, D.R.; French, F.S.; Migeon, C.J. Deletion of the steroid-binding domain of the human androgen receptor gene in one family with complete androgen insensitivity syndrome: Evidence for further genetic heterogeneity in this syndrome. Proc. Natl. Acad. Sci. USA 1988, 85, 8151–8155. [Google Scholar] [CrossRef] [Green Version]
- Kazemi-Esfarjani, P.; Trifiro, M.A.; Pinsky, L. Evidence for a repressive function of the long polyglutamine tract in the human androgen receptor: Possible pathogenetic relevance for the (CAG)n-expanded neuronopathies. Hum. Mol. Genet. 1995, 4, 523–527. [Google Scholar] [CrossRef]
- Chamberlain, N.L.; Driver, E.D.; Miesfeld, R.L. The length and location of CAG trinucleotide repeats in the androgen receptor N-terminal domain affect transactivation function. Nucleic Acids Res. 1994, 22, 3181–3186. [Google Scholar] [CrossRef] [Green Version]
- Ibáñez, L.; Ong, K.K.; Mongan, N.P.; Jääskeläinen, J.; Marcos, M.V.; Hughes, I.A.; De Zegher, F.; Dunger, D.B. Androgen Receptor Gene CAG Repeat Polymorphism in the Development of Ovarian Hyperandrogenism. J. Clin. Endocrinol. Metab. 2003, 88, 3333–3338. [Google Scholar] [CrossRef] [Green Version]
- Hsing, A.W.; Gao, Y.T.; Wu, G.; Wang, X.; Deng, J.; Chen, Y.L.; Sesterhenn, I.A.; Mostofi, F.K.; Benichou, J.; Chang, C. Polymorphic CAG and GGN repeat lengths in the androgen receptor gene and prostate cancer risk: A population-based case-control study in China. Cancer Res. 2000, 60, 5111–5116. [Google Scholar]
- Rebbeck, T.R.; Kantoff, P.W.; Krithivas, K.; Neuhausen, S.; Blackwood, M.A.; Godwin, A.K.; Daly, M.B.; Narod, S.A.; Garber, J.E.; Lynch, H.T.; et al. Modification of BRCA1-associated breast cancer risk by the polymorphic androgen-receptor CAG repeat. Am. J. Hum. Genet. 1999, 64, 1371–1377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giovannucci, E.; Stampfer, M.J.; Krithivas, K.; Brown, M.; Brufsky, A.; Talcott, J.; Hennekens, C.H.; Kantoff, P.W. The CAG repeat within the androgen receptor gene and its relationship to prostate cancer. Proc. Natl. Acad. Sci. USA 1997, 94, 3320–3323. [Google Scholar] [CrossRef] [Green Version]
- La Spada, A.R.; Wilson, E.M.; Lubahn, D.B.; Harding, A.E.; Fischbeck, K.H. Androgen receptor gene mutations in X-linked spinal and bulbar muscular atrophy. Nature 1991, 352, 77–79. [Google Scholar] [CrossRef] [PubMed]
- Yepuru, M.; Wu, Z.; Kulkarni, A.; Yin, F.; Barrett, C.M.; Kim, J.; Steiner, M.S.; Miller, D.D.; Dalton, J.T.; Narayanan, R. Steroidogenic Enzyme AKR1C3 Is a Novel Androgen Receptor-Selective Coactivator that Promotes Prostate Cancer Growth. Clin. Cancer Res. 2013, 19, 5613–5625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boorjian, S.A.; Heemers, H.V.; Frank, I.; Farmer, S.A.; Schmidt, L.J.; Sebo, T.J.; Tindall, D.J. Expression and significance of androgen receptor coactivators in urothelial carcinoma of the bladder. Endocr. Relat. Cancer 2009, 16, 123–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ngan, E.S.-W.; Hashimoto, Y.; Ma, Z.-Q.; Tsai, M.-J.; Tsai, S.Y. Overexpression of Cdc25B, an androgen receptor coactivator, in prostate cancer. Oncogene 2003, 22, 734–739. [Google Scholar] [CrossRef]
- Gnanapragasam, V.J.; Leung, H.Y.; Pulimood, A.S.; Neal, D.E.; Robson, C.N. Expression of RAC 3, a steroid hormone receptor co-activator in prostate cancer. Br. J. Cancer 2001, 85, 1928–1936. [Google Scholar] [CrossRef]
- Mongan, N.; Jääskeläinen, J.; Bhattacharyya, S.; Leu, R.; Hughes, I.; Mongan, N.P. Steroid receptor coactivator-3 glutamine repeat polymorphism and the androgen insensitivity syndrome. Eur. J. Endocrinol. 2003, 148, 277–279. [Google Scholar] [CrossRef] [Green Version]
- Tan, J.-A.; Hall, S.H.; Petrusz, P.; French, F.S. Thyroid Receptor Activator Molecule, TRAM-1, Is an Androgen Receptor Coactivator. Endocrinology 2000, 141, 3440–3450. [Google Scholar] [CrossRef]
- Han, G.; Xie, S.; Fang, H.; Li, G.; Han, Y.; Qin, Z. The AIB1 gene polyglutamine repeat length polymorphism contributes to risk of epithelial ovarian cancer risk: A case–control study. Tumor Biol. 2015, 36, 371–374. [Google Scholar] [CrossRef]
- Burandt, E.; Jens, G.; Holst, F.; Jänicke, F.; Müller, V.; Quaas, A.; Choschzick, M.; Wilczak, W.; Terracciano, L.; Simon, R.; et al. Prognostic relevance of AIB1 (NCoA3) amplification and overexpression in breast cancer. Breast Cancer Res. Treat. 2013, 137, 745–753. [Google Scholar] [CrossRef] [PubMed]
- Luo, F.; Li, W.; Zhang, J.; Huang, K.; Fu, J.; Xie, Z. Overexpression of steroid receptor coactivator-3 in bone cancers: An in vivo immunohistochemical study with tissue microarray. Pathol. Res. Pract. 2013, 209, 790–796. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Fang, Z.-Y.; Ding, Y.; Yao, W.-T.; Yang, Y.; Zhu, Z.-Q.; Wang, W.; Zhang, Q.-X. Amplifications of NCOA3 gene in colorectal cancers in a Chinese population. World J. Gastroenterol. 2012, 18, 855–860. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Benyamin, B.; McEvoy, B.P.; Gordon, S.; Henders, A.K.; Nyholt, D.R.; Madden, P.A.; Heath, A.C.; Martin, N.G.; Montgomery, G.; et al. Common SNPs explain a large proportion of the heritability for human height. Nat. Genet. 2010, 42, 565–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villani, A.; Tabori, U.; Schiffman, J.; Shlien, A.; Beyene, J.; Druker, H.; Novokmet, A.; Finlay, J.; Malkin, D. Biochemical and imaging surveillance in germline TP53 mutation carriers with Li-Fraumeni syndrome: A prospective observational study. Lancet Oncol. 2011, 12, 559–567. [Google Scholar] [CrossRef]
- Limacher, J.-M.; Frebourg, T.; Natarajan-Ame, S.; Bergerat, J.-P. Two metachronous tumors in the radiotherapy fields of a patient with Li-Fraumeni syndrome. Int. J. Cancer 2001, 96, 238–242. [Google Scholar] [CrossRef]
- Stinco, G.; Governatori, G.; Mattighello, P.; Patrone, P. Multiple cutaneous neoplasms in a patient with Rothmund–Thomson syndrome: Case report and published work review. J. Dermatol. 2008, 35, 154–161. [Google Scholar] [CrossRef]
- Ognjanovic, S.; Olivier, M.; Bergemann, T.L.; Hainaut, P. Sarcomas in TP53 germline mutation carriers. Cancer 2012, 118, 1387–1396. [Google Scholar] [CrossRef]
- Thomas, D.M.; Ballinger, M.L. Etiologic, environmental and inherited risk factors in sarcomas. J. Surg. Oncol. 2015, 111, 490–495. [Google Scholar] [CrossRef]
- Poos, K.; Smida, J.; Nathrath, M.; Maugg, D.; Baumhoer, D.; Neumann, A.; Korsching, E. Structuring osteosarcoma knowledge: An osteosarcoma-gene association database based on literature mining and manual annotation. Database 2014, 2014, bau042. [Google Scholar] [CrossRef] [Green Version]
- De Bruin, E.C.; McGranahan, N.; Mitter, R.; Salm, M.; Wedge, D.C.; Yates, L.; Jamal-Hanjani, M.; Shafi, S.; Murugaesu, N.; Rowan, A.J.; et al. Spatial and temporal diversity in genomic instability processes defines lung cancer evolution. Science 2014, 346, 251–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galanos, P.S.; Vougas, K.; Walter, D.; Polyzos, A.; Maya-Mendoza, A.; Haagensen, E.J.; Kokkalis, A.; Roumelioti, F.-M.; Gagos, S.; Tzetis, M.; et al. Chronic p53-independent p21 expression causes genomic instability by deregulating replication licensing. Nat. Cell Biol. 2016, 18, 777–789. [Google Scholar] [CrossRef] [PubMed]
- Al-Romaih, K.; Bayani, J.; Vorobyova, J.; Karaskova, J.; Park, P.; Zielenska, M.; Squire, J.A. Chromosomal instability in osteosarcoma and its association with centrosome abnormalities. Cancer Genet. Cytogenet. 2003, 144, 91–99. [Google Scholar] [CrossRef]
- Carter, S.L.; Eklund, A.C.; Kohane, I.S.; Harris, L.N.; Szállási, Z. A signature of chromosomal instability inferred from gene expression profiles predicts clinical outcome in multiple human cancers. Nat. Genet. 2006, 38, 1043–1048. [Google Scholar] [CrossRef]
- Phillips, J.C.; Stephenson, B.; Hauck, M.; Dillberger, J. Heritability and segregation analysis of osteosarcoma in the Scottish deerhound. Genomics 2007, 90, 354–363. [Google Scholar] [CrossRef] [Green Version]
- Vonholdt, B.M.; Pollinger, J.P.; Lohmueller, K.E.; Han, E.; Parker, H.G.; Quignon, P.; Degenhardt, J.D.; Boyko, A.R.; Earl, D.A.; Auton, A.; et al. Genome-wide SNP and haplotype analyses reveal a rich history underlying dog domestication. Nature 2010, 464, 898–902. [Google Scholar] [CrossRef] [Green Version]
- Phillips, J.C.; Lembcke, L.; Chamberlin, T. A novel locus for canine osteosarcoma (OSA1) maps to CFA34, the canine orthologue of human 3q26. Genomics 2010, 96, 220–227. [Google Scholar] [CrossRef] [Green Version]
- Karlsson, E.K.; Sigurdsson, S.; Ivansson, E.; Thomas, R.; Elvers, I.; Curtis, J.L.; Howald, C.; Tonomura, N.; Perloski, M.; Swofford, R.; et al. Genome-wide analyses implicate 33 loci in heritable dog osteosarcoma, including regulatory variants near CDKN2A/B. Genome Biol. 2013, 14, R132. [Google Scholar] [CrossRef]
- Millanta, F.; Asproni, P.; Cancedda, S.; Vignoli, M.; Poli, A. Immunohistochemical Expression of COX-2, mPGES and EP2 Receptor in Normal and Reactive Canine Bone and in Canine Osteosarcoma. J. Comp. Pathol. 2012, 147, 153–160. [Google Scholar] [CrossRef] [PubMed]
- McCleese, J.K.; Bear, M.D.; Kulp, S.K.; Mazcko, C.; Khanna, C.; London, C. Met interacts with EGFR and Ron in canine osteosarcoma. Vet. Comp. Oncol. 2013, 11, 124–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, L.; Argyle, S.A.; Kamida, A.; Morrison, K.O.; Argyle, D.J. The long-acting COX-2 inhibitor mavacoxib (Trocoxil™) has anti-proliferative and pro-apoptotic effects on canine cancer cell lines and cancer stem cells in vitro. BMC Vet. Res. 2014, 10, 184. [Google Scholar] [CrossRef] [Green Version]
- Shahi, M.H.; York, D.; Gandour-Edwards, R.; Withers, S.S.; Holt, R.; Rebhun, R.B. BMI1 Is Expressed in Canine Osteosarcoma and Contributes to Cell Growth and Chemotherapy Resistance. PLoS ONE 2015, 10, e0131006. [Google Scholar] [CrossRef] [PubMed]
- Selvarajah, G.T.; Kirpensteijn, J.; Van Wolferen, M.E.; Rao, N.A.; Fieten, H.; Mol, J.A. Gene expression profiling of canine osteosarcoma reveals genes associated with short and long survival times. Mol. Cancer 2009, 8, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selvarajah, G.T.; Verheije, M.H.; Kik, M.; Slob, A.; Rottier, P.J.; Mol, J.A.; Kirpensteijn, J. Expression of epidermal growth factor receptor in canine osteosarcoma: Association with clinicopathological parameters and prognosis. Vet. J. 2012, 193, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Selvarajah, G.T.; Bonestroo, F.A.S.; Kirpensteijn, J.; Kik, M.J.L.; Van Der Zee, R.; Van Eden, W.; Timmermans-Sprang, E.P.M.; Slob, A.; Mol, J.A. Heat shock protein expression analysis in canine osteosarcoma reveals HSP60 as a potentially relevant therapeutic target. Cell Stress Chaperon. 2013, 18, 607–622. [Google Scholar] [CrossRef] [Green Version]
- Maniscalco, L.; Iussich, S.; Morello, E.; Martano, M.; Gattino, F.; Miretti, S.; Biolatti, B.; Accornero, P.; Martignani, E.; Sánchez-Céspedes, R.; et al. Increased expression of insulin-like growth factor-1 receptor is correlated with worse survival in canine appendicular osteosarcoma. Vet. J. 2015, 205, 272–280. [Google Scholar] [CrossRef]
- Urfer, S. How the past affects the present: A genetic history of the Irish wolfhound. In Dogs: Biology, Behavior and Health Disorders; DeGiovine, V.M., Ed.; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2011; pp. 69–91. [Google Scholar]
- Dillberger, J.E.; McAtee, S.A. Osteosarcoma inheritance in two families of Scottish deerhounds. Canine Genet. Epidemiol. 2017, 4, 1–12. [Google Scholar] [CrossRef] [Green Version]
- The Cancer Genome Atlas Research Network. Comprehensive molecular characterization of urothelial bladder carcinoma. Nature 2014, 507, 315–322. [Google Scholar] [CrossRef] [Green Version]
- Calon, A.; Lonardo, E.; Berenguer, A.; Espinet, E.; Hernando-Momblona, X.; Iglesias, M.; Sevillano, M.; Palomo-Ponce, S.; Tauriello, D.V.F.; Byrom, D.; et al. Stromal gene expression defines poor-prognosis subtypes in colorectal cancer. Nat. Genet. 2015, 47, 320–329. [Google Scholar] [CrossRef] [Green Version]
- Pfefferle, A.D.; Agrawal, Y.N.; Koboldt, D.C.; Kanchi, K.L.; Herschkowitz, J.I.; Mardis, E.R.; Rosen, J.M.; Perou, C.M. Genomic profiling of murine mammary tumors identifies potential personalized drug targets for p53-deficient mammary cancers. Dis. Model. Mech. 2016, 9, 749–757. [Google Scholar] [CrossRef] [Green Version]
- Van ’t Veer, L.J.; Dai, H.; van de Vijver, M.J.; He, Y.D.; Hart, A.A.M.; Mao, M.; Peterse, H.L.; van der Kooy, K.; Marton, M.J.; Witteveen, A.T.; et al. Gene expression profiling predicts clinical outcome of breast cancer. Nature 2002, 415, 530–536. [Google Scholar] [CrossRef] [Green Version]
- Fehrmann, R.S.N.; Karjalainen, J.M.; Krajewska, M.; Westra, H.-J.; Maloney, D.; Simeonov, A.; Pers, T.H.; Hirschhorn, J.N.; Jansen, R.C.; Schultes, E.; et al. Gene expression analysis identifies global gene dosage sensitivity in cancer. Nat. Genet. 2015, 47, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Gillet, J.-P.; Andersen, J.B.; Madigan, J.P.; Varma, S.; Bagni, R.K.; Powell, K.; Burgan, W.E.; Wu, C.-P.; Calcagno, A.M.; Ambudkar, S.V.; et al. A Gene Expression Signature Associated with Overall Survival in Patients with Hepatocellular Carcinoma Suggests a New Treatment Strategy. Mol. Pharmacol. 2016, 89, 263–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okada, Y.; Nagase, H.; Harris, E.D. A metalloproteinase from human rheumatoid synovial fibroblasts that digests connective tissue matrix components. Purification and characterization. J. Biol. Chem. 1986, 261, 14245–14255. [Google Scholar]
- Kurahara, S.-I.; Shinohara, M.; Ikebe, T.; Nakamura, S.; Beppu, M.; Hiraki, A.; Takeuchi, H.; Shirasuna, K. Expression of MMPS, MT-MMP, and TIMPs in squamous cell carcinoma of the oral cavity: Correlations with tumor invasion and metastasis. Head Neck 1999, 21, 627–638. [Google Scholar] [CrossRef]
- Mehner, C.; Miller, E.; Nassar, A.; Bamlet, W.R.; Radisky, E.S.; Radisky, D.C. Tumor cell expression of MMP3 as a prognostic factor for poor survival in pancreatic, pulmonary, and mammary carcinoma. Genes Cancer 2015, 6, 480–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banik, D.; Netherby, C.S.; Bogner, P.N.; Abrams, S.I. MMP3-Mediated tumor progression is controlled transcriptionally by a novel IRF8-MMP3 interaction. Oncotarget 2015, 6, 15164–15179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bielack, S.S.; Kempf-Bielack, B.; Delling, G.; Exner, G.U.; Flege, S.; Helmke, K.; Kotz, R.; Salzer-Kuntschik, M.; Werner, M.; Winkelmann, W.; et al. Prognostic Factors in High-Grade Osteosarcoma of the Extremities or Trunk: An Analysis of 1,702 Patients Treated on Neoadjuvant Cooperative Osteosarcoma Study Group Protocols. J. Clin. Oncol. 2002, 20, 776–790. [Google Scholar] [CrossRef] [PubMed]
- Reddy, K.I.A.; Wafa, H.; Gaston, C.L.; Grimer, R.J.; Abudu, A.T.; Jeys, L.M.; Carter, S.; Tillman, R.M.; Zuiderbaan, H.A.; Khamaisy, S.; et al. Does amputation offer any survival benefit over limb salvage in osteosarcoma patients with poor chemonecrosis and close margins? Bone Jt. J. 2015, 97, 115–120. [Google Scholar] [CrossRef]
- Fray, M.J.; Dickinson, R.P.; Huggins, J.P.; Occleston, N.L. A Potent, Selective Inhibitor of Matrix Metalloproteinase-3 for the Topical Treatment of Chronic Dermal Ulcers. J. Med. Chem. 2003, 46, 3514–3525. [Google Scholar] [CrossRef]
- Bramhall, S.; Schulz, J.; Nemunaitis, J.; Brown, P.D.; Baillet, M.; Buckels, J.A.C. A double-blind placebo-controlled, randomised study comparing gemcitabine and marimastat with gemcitabine and placebo as first line therapy in patients with advanced pancreatic cancer. Br. J. Cancer 2002, 87, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Davies, B.; Brown, P.D.; East, N.; Crimmin, M.J.; Balkwill, F.R. A synthetic matrix metalloproteinase inhibitor decreases tumor burden and prolongs survival of mice bearing human ovarian carcinoma xenografts. Cancer Res. 1993, 53, 2087–2091. [Google Scholar] [PubMed]
- Watson, S.A.; Morris, T.M.; Collins, H.M.; Bawden, L.J.; Hawkins, K.; Bone, E.A. Inhibition of tumour growth by marimastat in a human xenograft model of gastric cancer: Relationship with levels of circulating CEA. Br. J. Cancer 1999, 81, 19–23. [Google Scholar] [CrossRef] [Green Version]
- Shepherd, F.A.; Giaccone, G.; Seymour, L.; Debruyne, C.; Bezjak, A.; Hirsh, V.; Smylie, M.; Rubin, S.; Martins, H.; Lamont, A.; et al. Prospective, Randomized, Double-Blind, Placebo-Controlled Trial of Marimastat After Response to First-Line Chemotherapy in Patients With Small-Cell Lung Cancer: A Trial of the National Cancer Institute of Canada-Clinical Trials Group and the European Organization for Research and Treatment of Cancer. J. Clin. Oncol. 2002, 20, 4434–4439. [Google Scholar] [CrossRef]
- Tsai, H.-C.; Su, H.-L.; Huang, C.-Y.; Fong, Y.-C.; Hsu, C.-J.; Tang, C.-H. CTGF increases matrix metalloproteinases expression and subsequently promotes tumor metastasis in human osteosarcoma through down-regulating miR-519d. Oncotarget 2014, 5, 3800–3812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.-F.; Du, W.-X.; Chen, J.-J. Elevated expression of matrix metalloproteinase-3 in human osteosarcoma and its association with tumor metastasis. J. BUON 2016, 21, 235–243. [Google Scholar]
- Shows, T.B.; Eddy, R.L.; Byers, M.G.; Fukushima, Y.; Dehaven, C.R.; Murray, J.C.; Bell, G.I. Polymorphic Human Glucose Transporter Gene (GLUT) is on Chromosome 1p31.3→p35. Diabetes 1987, 36, 546–549. [Google Scholar] [CrossRef] [Green Version]
- Younes, M.; Brown, R.W.; Stephenson, M.; Gondo, M.; Cagle, P.T. Overexpression of Glut1 and Glut3 in stage I nonsmall cell lung carcinoma is associated with poor survival. Cancer 1997, 80, 1046–1051. [Google Scholar] [CrossRef]
- Kunkel, M.; Reichert, T.E.; Benz, P.; Lehr, H.-A.; Jeong, J.-H.; Wieand, S.; Bartenstein, P.; Wagner, W.; Whiteside, T.L. Overexpression of Glut-1 and increased glucose metabolism in tumors are associated with a poor prognosis in patients with oral squamous cell carcinoma. Cancer 2003, 97, 1015–1024. [Google Scholar] [CrossRef]
- Smeland, E.; Kilvaer, T.; Sorbye, S.; Valkov, A.; Andersen, S.; Bremnes, R.M.; Busund, L.-T.; Donnem, T. Prognostic Impacts of Hypoxic Markers in Soft Tissue Sarcoma. Sarcoma 2012, 2012, 541650. [Google Scholar] [CrossRef]
- Kubo, T.; Shimose, S.; Fujimori, J.; Furuta, T.; Arihiro, K.; Ochi, M. Does Expression of Glucose Transporter Protein-1 Relate to Prognosis and Angiogenesis in Osteosarcoma? Clin. Orthop. Relat. Res. 2015, 473, 305–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacEwen, E.G.; Kurzman, I.D. Canine osteosarcoma: Amputation and chemoimmunotherapy. Vet. Clin. N. Am. Small Anim. Pract. 1996, 26, 123–133. [Google Scholar] [CrossRef]
- Klepper, J.; Flörcken, A.; Fischbarg, J.; Voit, T. Effects of anticonvulsants on GLUT1-mediated glucose transport in GLUT1 deficiency syndrome in vitro. Eur. J. Nucl. Med. Mol. Imaging 2003, 162, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Shuralyova, I.; Tajmir, P.; Bilan, P.J.; Sweeney, G.; Coe, I.R. Inhibition of glucose uptake in murine cardiomyocyte cell line HL-1 by cardioprotective drugs dilazep and dipyridamole. Am. J. Physiol. Circ. Physiol. 2004, 286, H627–H632. [Google Scholar] [CrossRef]
- Louters, L.L.; Stehouwer, N.; Rekman, J.; Tidball, A.M.; Cok, A.; Holstege, C.P. Verapamil Inhibits the Glucose Transport Activity of GLUTJ. Med. Toxicol. 2010, 6, 100–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rufino, A.; Rosa, S.C.; Judas, F.; Mobasheri, A.; Lopes, M.; Mendes, A.F. Expression and function of K(ATP) channels in normal and osteoarthritic human chondrocytes: Possible role in glucose sensing. J. Cell. Biochem. 2013, 114, 1879–1889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woo, D.K.; Kim, H.S.; Lee, H.S.; Kang, Y.H.; Yang, H.-K.; Kim, W.H. Altered expression and mutation of β-catenin gene in gastric carcinomas and cell lines. Int. J. Cancer 2001, 95, 108–113. [Google Scholar] [CrossRef]
- Dellinger, T.; Planutis, K.; Jandial, D.D.; Eskander, R.N.; Martinez, M.E.; Zi, X.; Monk, B.J.; Holcombe, R.F. Expression of the Wnt antagonist Dickkopf-3 is associated with prognostic clinicopathologic characteristics and impairs proliferation and invasion in endometrial cancer. Gynecol. Oncol. 2012, 126, 259–267. [Google Scholar] [CrossRef] [Green Version]
- Dey, N.; Barwick, B.G.; Moreno, C.S.; Ordanic-Kodani, M.; Chen, Z.; Oprea-Ilies, G.; Tang, W.; Catzavelos, C.; Kerstann, K.F.; Sledge, J.G.W.; et al. Wnt signaling in triple negative breast cancer is associated with metastasis. BMC Cancer 2013, 13, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Voloshanenko, O.; Erdmann, G.; Dubash, T.D.; Augustin, I.; Metzig, M.; Moffa, G.; Hundsrucker, C.; Kerr, G.; Sandmann, T.; Anchang, B.; et al. Wnt secretion is required to maintain high levels of Wnt activity in colon cancer cells. Nat. Commun. 2013, 4, 2610. [Google Scholar] [CrossRef] [Green Version]
- Baehs, S.; Herbst, A.; Thieme, S.E.; Perschl, C.; Behrens, A.; Scheel, S.; Jung, A.; Brabletz, T.; Göke, B.; Blum, H.; et al. Dickkopf-4 is frequently down-regulated and inhibits growth of colorectal cancer cells. Cancer Lett. 2009, 276, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Hall, C.L.; Zhang, H.; Baile, S.; Ljungman, M.; Kuhstoss, S.; Keller, E.T. p21(CIP-1/WAF-1) Induction Is Required to Inhibit Prostate Cancer Growth Elicited by Deficient Expression of the Wnt Inhibitor Dickkopf-1 (DKK-1). Cancer Res. 2010, 70, 9916–9926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, X.-L.; Qin, X.-R.; Zhang, S.; Ye, L.-H. Downregulation of Dickkopf-1 is responsible for high proliferation of breast cancer cells via losing control of Wnt/β-catenin signaling. Acta Pharmacol. Sin. 2010, 31, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, S.Y.; Hsieh, P.S.; Chiu, C.T.; Chen, W.Y. Dickkopf-3/REIC functions as a suppressor gene of tumor growth. Oncogene 2004, 23, 9183–9189. [Google Scholar] [CrossRef] [Green Version]
- Lee, E.-J.; Jo, M.; Rho, S.B.; Park, K.; Yoo, Y.-N.; Park, J.; Chae, M.; Zhang, W.; Lee, J.-H. Dkk3, downregulated in cervical cancer, functions as a negative regulator of β-catenin. Int. J. Cancer 2009, 124, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Xiang, T.; Li, L.; Yin, X.; Zhong, L.; Peng, W.; Qiu, Z.; Ren, G.; Tao, Q. Epigenetic silencing of the WNT antagonist Dickkopf 3 disrupts normal Wnt/β-catenin signalling and apoptosis regulation in breast cancer cells. J. Cell. Mol. Med. 2013, 17, 1236–1246. [Google Scholar] [CrossRef]
- Lin, C.H.; Guo, Y.; Ghaffar, S.; McQueen, P.; Pourmorady, J.; Christ, A.B.; Rooney, K.; Ji, T.; Eskander, R.; Zi, X.; et al. Dkk-3, a Secreted Wnt Antagonist, Suppresses Tumorigenic Potential and Pulmonary Metastasis in Osteosarcoma. Sarcoma 2013, 2013, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.; Kurenbekova, L.; Gao, Y.; Roos, A.; Creighton, C.J.; Rao, P.; Hicks, J.; Man, T.-K.; Lau, C.; Brown, A.M.C.; et al. NKD2, a negative regulator of Wnt signaling, suppresses tumor growth and metastasis in osteosarcoma. Oncogene 2015, 34, 5069–5079. [Google Scholar] [CrossRef] [Green Version]
- Kessenbrock, K.; Dijkgraaf, G.J.P.; Lawson, D.A.; Littlepage, L.E.; Shahi, P.; Pieper, U.; Werb, Z. A role for matrix metalloproteinases in regulating mammary stem cell function via the Wnt signaling pathway. Cell Stem Cell 2013, 13, 300–313. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.J.; Kang, G.J.; Kim, E.J.; Park, M.K.; Byun, H.J.; Nam, S.; Lee, H.; Lee, C.H. Novel effects of sphingosylphosphorylcholine on invasion of breast cancer: Involvement of matrix metalloproteinase-3 secretion leading to WNT activation. Biochim. Biophys. Acta (BBA)—Mol. Basis Dis. 2016, 1862, 1533–1543. [Google Scholar] [CrossRef]
- Fiorito, V.; Chiabrando, D.; Petrillo, S.; Bertino, F.; Tolosano, E. The Multifaceted Role of Heme in Cancer. Front. Oncol. 2019, 9, 1540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morell, H.; Savoie, J.C.; London, I.M. The biosynthesis of heme and the incorporation of glycine into globin in rabbit bone marrow in vitro. J. Biol. Chem. 1958, 233, 923–929. [Google Scholar] [PubMed]
- Mölzer, C.; Pfleger, B.; Putz, E.; Roßmann, A.; Schwarz, U.; Wallner, M.; Bulmer, A.C.; Wagner, K.-H. In vitro DNA-damaging effects of intestinal and related tetrapyrroles in human cancer cells. Exp. Cell Res. 2013, 319, 536–545. [Google Scholar] [CrossRef]
- Jansen, T.; Hortmann, M.; Oelze, M.; Opitz, B.; Steven, S.; Schell, R.; Knorr, M.; Karbach, S.; Schuhmacher, S.; Wenzel, P.; et al. Conversion of biliverdin to bilirubin by biliverdin reductase contributes to endothelial cell protection by heme oxygenase-1—Evidence for direct and indirect antioxidant actions of bilirubin. J. Mol. Cell. Cardiol. 2010, 49, 186–195. [Google Scholar] [CrossRef]
- Adamopoulos, C.; Gargalionis, A.N.; Basdra, E.K.; Papavassiliou, A.G. Deciphering signaling networks in osteosarcoma pathobiology. Exp. Biol. Med. 2016, 241, 1296–1305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins-Neves, S.R.; Corver, W.E.; Paiva-Oliveira, D.I.; van den Akker, B.E.; Briaire-de-Bruijn, I.H.; Bovee, J.V.; Gomes, C.M.; Cleton-Jansen, A.M. Osteosarcoma Stem Cells Have Active Wnt/beta-catenin and Overexpress SOX2 and KLFJ4. J. Cell Physiol. 2016, 231, 876–886. [Google Scholar] [CrossRef]
- Martins-Neves, S.R.; Paiva-Oliveira, D.I.; Fontes-Ribeiro, C.; Bovee, J.; Cleton-Jansen, A.M.; Gomes, C.M.F. IWR-1, a tankyrase inhibitor, attenuates Wnt/beta-catenin signaling in cancer stem-like cells and inhibits in vivo the growth of a subcutaneous human osteosarcoma xenograft. Cancer Lett. 2018, 414, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Shakya, A.; Goren, A.; Shalek, A.K.; German, C.N.; Snook, J.; Kuchroo, V.K.; Yosef, N.; Chan, R.C.; Regev, A.; Williams, M.A.; et al. Oct1 and OCA-B are selectively required for CD4 memory T cell function. J. Exp. Med. 2015, 212, 2115–2131. [Google Scholar] [CrossRef]
- Reya, T.; O’Riordan, M.X.D.; Okamura, R.; Devaney, E.; Willert, K.; Nusse, R.; Grosschedl, R. Wnt Signaling Regulates B Lymphocyte Proliferation through a LEF-1 Dependent Mechanism. Immunity 2000, 13, 15–24. [Google Scholar] [CrossRef] [Green Version]
- Huelsken, J.; Behrens, J. The Wnt signalling pathway. J. Cell Sci. 2002, 115, 3977–3978. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, R.E.I.; Hackam, A.S. Analysis of Dickkopf3 interactions with Wnt signaling receptors. Growth Factors 2010, 28, 232–242. [Google Scholar] [CrossRef] [PubMed]
- Behrens, J.; Von Kries, J.P.; Kühl, M.; Bruhn, L.; Wedlich, D.; Grosschedl, R.; Birchmeier, W. Functional interaction of β-catenin with the transcription factor LEF-1. Nature 1996, 382, 638–642. [Google Scholar] [CrossRef] [PubMed]
- Hsu, S.-C.; Galceran, J.; Grosschedl, R. Modulation of Transcriptional Regulation by LEF-1 in Response to Wnt-1 Signaling and Association with β-Catenin. Mol. Cell. Biol. 1998, 18, 4807–4818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simpson, S.; Edwards, J.; Ferguson-Mignan, T.F.N.; Cobb, M.; Mongan, N.P.; Rutland, C.S. Genetics of Human and Canine Dilated Cardiomyopathy. Int. J. Genom. 2015, 2015, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Simpson, S.; Dunning, M.D.; Brownlie, S.; Patel, J.; Godden, M.; Cobb, M.; Mongan, N.P.; Rutland, C.S. Multiple Genetic Associations with Irish Wolfhound Dilated Cardiomyopathy. BioMed Res. Int. 2016, 2016, 1–14. [Google Scholar] [CrossRef]
- Lai, C.-L.; L’Eplattenier, H.; Ham, R.V.D.; Verseijden, F.; Jagtenberg, A.; Mol, J.; Teske, E. Androgen Receptor CAG Repeat Polymorphisms in Canine Prostate Cancer. J. Vet. Intern. Med. 2008, 22, 1380–1384. [Google Scholar] [CrossRef]
- Konno, A.; Inoue-Murayama, M.; Hasegawa, T. Androgen receptor gene polymorphisms are associated with aggression in Japanese Akita Inu. Biol. Lett. 2011, 7, 658–660. [Google Scholar] [CrossRef] [Green Version]
- Maejima, M.; Inoue-Murayama, M.; Koshimura, A.; Kato, S.; Nara, H.; Randi, E.; Kitagawa, H.; Iwasaki, T.; Murayama, Y. Allelic variation of two poly-glutamine repeats in the canine androgen receptor gene. J. Anim. Genet. 2005, 32, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Wright, S. The Genetical Structure of Populations. Ann. Eugen. 1949, 15, 323–354. [Google Scholar] [CrossRef]
- Slatkin, M. A Measure of Population Subdivision Based on Microsatellite Allele Frequencies. Genetics 1995, 139, 457–462. [Google Scholar]
- Fu, Y.X. Statistical Tests of Neutrality of Mutations against Population Growth, Hitchhiking and Background Selection. Genetics 1997, 147, 915–925. [Google Scholar] [PubMed]
- R CoreTeam. R: A Language and Environment for Statistical Computing; R Foundation: Vienna, Austria, 2013. [Google Scholar]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14, R13–R36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trapnell, C.; Hendrickson, D.G.; Sauvageau, M.; Goff, L.; Rinn, J.L.; Pachter, L. Differential analysis of gene regulation at transcript resolution with RNA-seq. Nat. Biotechnol. 2013, 31, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Wang, J.; Jaehnig, E.J.; Shi, Z.; Zhang, B. WebGestalt 2019: Gene set analysis toolkit with revamped UIs and APIs. Nucleic Acids Res. 2019, 47, W199–W205. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Kirov, S.; Snoddy, J. WebGestalt: An integrated system for exploring gene sets in various biological contexts. Nucleic Acids Res. 2005, 33, W741–W748. [Google Scholar] [CrossRef]
- Wang, J.; Duncan, D.; Shi, Z.; Zhang, B. WEB-based GEne SeT AnaLysis Toolkit (WebGestalt): Update 2013. Nucleic Acids Res. 2013, 41, W77–W83. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, E.M.; Laursen, K.B.; Whitchurch, J.; McWilliam, A.; Ødum, N.; Persson, J.L.; Heery, D.; Gudas, L.J.; Mongan, N.P. MiR137 is an androgen regulated repressor of an extended network of transcriptional coregulators. Oncotarget 2015, 6, 35710–35725. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene_ID | Gene | Log2 Fold Change | p Value |
|---|---|---|---|
| ENSCAFG00000007134 | MYBPC1 | −14.4441 | 0.01395 |
| ENSCAFG00000001705 | MB | −13.3627 | 0.0081 |
| ENSCAFG00000013875 | MYL1 | −13.1689 | 0.00085 |
| ENSCAFG00000008253 | ACTA1 | −12.3844 | 0.003 |
| ENSCAFG00000011103 | NRAP | −12.1537 | 0.00015 |
| ENSCAFG00000005717 | ENSCAFG00000005717 | −12.0616 | 0.00035 |
| ENSCAFG00000014281 | PYGM | −11.6411 | 0.0057 |
| ENSCAFG00000011619 | MYH7 | −11.4064 | 0.0001 |
| ENSCAFG00000002620 | TNNT1 | −11.3052 | 0.0119 |
| ENSCAFG00000001016 | TRDN | −11.034 | 0.0005 |
| ENSCAFG00000009333 | CESDD1 | −10.7269 | 0.04655 |
| ENSCAFG00000005896 | RYR1 | −9.83487 | 0.01135 |
| ENSCAFG00000009404 | TNNC1 | −9.61599 | 0.0317 |
| ENSCAFG00000015475 | DES | −9.54008 | 0.0062 |
| ENSCAFG00000005358 | HHATL | −9.49779 | 0.0023 |
| ENSCAFG00000008707 | CKMT2 | −9.43202 | 0.00015 |
| ENSCAFG00000017254 | ATP2A1 | −9.34349 | 0.01225 |
| ENSCAFG00000001140 | MYOT | −9.08389 | 0.0191 |
| ENSCAFG00000012927 | ALPK3 | −9.01117 | 0.0005 |
| ENSCAFG00000005343 | KLHL40 | −8.96012 | 0.0234 |
| ENSCAFG00000002982 | MYBPC2 | −8.66855 | 0.01675 |
| ENSCAFG00000012432 | MYOZ2 | −8.62006 | 0.00415 |
| ENSCAFG00000011638 | SPTA1 | −8.56014 | 0.00135 |
| ENSCAFG00000001161 | OBSCN | −8.44333 | 0.01045 |
| ENSCAFG00000011418 | HFE2 | −8.31185 | 0.0051 |
| Gene_ID | Gene | Log2 Fold Change | p Value |
|---|---|---|---|
| ENSCAFG00000010484 | PKP2 | 3.54183 | 0.00095 |
| ENSCAFG00000005510 | EOMES | 3.56165 | 0.04155 |
| ENSCAFG00000012983 | GPR64 | 3.66911 | 0.0009 |
| ENSCAFG00000013448 | COLGALT2 | 3.69448 | 0.0007 |
| ENSCAFG00000007845 | DKK3 | 3.76815 | 0.0013 |
| ENSCAFG00000009722 | TOX3 | 3.77935 | 0.0002 |
| ENSCAFG00000023615 | MMP-12 | 3.78547 | 0.0001 |
| ENSCAFG00000002589 | COL9A1 | 3.84351 | 0.0001 |
| ENSCAFG00000003503 | ARHGEF5 | 3.84945 | 0.0006 |
| ENSCAFG00000005101 | RPSA | 3.86821 | 0.00135 |
| ENSCAFG00000019918 | FOXF1 | 3.8936 | 0.0005 |
| ENSCAFG00000017309 | RPL4 | 3.92678 | 0.00015 |
| ENSCAFG00000018123 | ARSI | 3.92752 | 0.0004 |
| ENSCAFG00000007280 | RNF180 | 4.12335 | 0.00015 |
| ENSCAFG00000010205 | ID4 | 4.12517 | 0.0108 |
| ENSCAFG00000006073 | POSTN | 4.15174 | 0.03365 |
| ENSCAFG00000009756 | ELOVL2 | 4.23619 | 0.00035 |
| ENSCAFG00000002307 | ASPN | 4.25607 | 0.02175 |
| ENSCAFG00000009135 | SAA1 | 4.42549 | 0.0053 |
| ENSCAFG00000019036 | LIPG | 4.44654 | 0.0061 |
| ENSCAFG00000016731 | MSX2 | 4.46357 | 0.0337 |
| ENSCAFG00000015063 | MMP3 | 4.56236 | 0.0337 |
| ENSCAFG00000018597 | LAMA1 | 4.58776 | 0.00025 |
| ENSCAFG00000029131 | HAPLN1 | 4.61072 | 0.00015 |
| ENSCAFG00000031443 | EREG | 5.61347 | 0.02875 |
| Gene Set | Database | Description | Size | Expect | Ratio | False Discovery Rate (FDR) |
|---|---|---|---|---|---|---|
| hsa05414 | KEGG | Dilated cardiomyopathy | 90 | 5.9888 | 2.6717 | 0.04748 |
| hsa05410 | KEGG | Hypertrophic cardiomyopathy | 83 | 5.523 | 2.7159 | 0.04748 |
| hsa04020 | KEGG | Calcium signaling pathway | 183 | 12.177 | 2.053 | 0.04748 |
| P02746 | Panther | Heme biosynthesis | 12 | 0.96911 | 7.2231 | 0.001603 |
| R-HSA-397014 | Reactome | Muscle contraction | 206 | 13.605 | 2.7932 | 0.0075 |
| R-HSA-189451 | Reactome | Heme biosynthesis | 11 | 0.72645 | 11.012 | 0.00 |
| R-HSA-390522 | Reactome | Striated Muscle Contraction | 36 | 2.3775 | 5.468 | 0.000151 |
| R-HSA-189445 | Reactome | Metabolism of porphyrins | 17 | 1.1227 | 8.0164 | 0.00017 |
| R-HSA-109582 | Reactome | Hemostasis | 620 | 40.946 | 1.6363 | 0.014273 |
| R-HSA-5578775 | Reactome | Ion homeostasis | 56 | 3.6983 | 3.5151 | 0.018011 |
| R-HSA-5576891 | Reactome | Cardiac conduction | 141 | 9.3118 | 2.3626 | 0.037904 |
| OSA Cases and Diagnosis | IWH (n = 379) | Rottweiler (n = 1019) |
|---|---|---|
| Number of OSA cases (%) | 0.53 | 3.04 |
| Male age of diagnosis (mean, years) | 5.34 | 7.65 |
| Female age of diagnosis (mean, years) | 7.00 ** | 8.50 * |
| Breed | Group | Diagnosis and Age | AR1 Q-Repeat | AR2 Q-Repeat | AR Q-Repeat Number (Mean ± SEM) |
|---|---|---|---|---|---|
| IWH | Male | OSA | 10.25 ± 0.25 (4) | 24.50 ± 0.29 (4) | 34.75 ± 0.25 (4) |
| Non OSA over 6.5 years | 10.05 ± 0.05 (19) | 24.31 ± 0.12 (16) | 34.31 ± 0.12 (16) | ||
| Female | OSA | 10.13 ± 0.13 (8) | 24.57 ± 0.13 (7) | 34.57 ± 0.13 (7) | |
| Non OSA over 9 years | 10.04 ± 0.04 (13) | 24.19 ± 0.16 (13) | 34.23 ± 0.15 (13) | ||
| Rottweiler | Male | OSA | 11.00 ± 0.00 (12) | 23.00 ± 0.00 (12) | 34.00 ± 0.00 (12) |
| Non OSA over 9 years | 11.00 ± 0.00 (49) | 23.00 ± 0.00 (50) | 34.00 ± 0.00 (49) | ||
| Female | OSA | 11.03 ± 0.03 (19) | 23.00 ± 0.00 (18) | 34.03 ± 0.03 (18) | |
| Non OSA over 10 years | 11.00 ± 0.00 (43) | 22.98 ± 0.02 (45) | 34.00 ± 0.00 (43) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Simpson, S.; Dunning, M.; de Brot, S.; Alibhai, A.; Bailey, C.; Woodcock, C.L.; Mestas, M.; Akhtar, S.; Jeyapalan, J.N.; Lothion-Roy, J.; et al. Molecular Characterisation of Canine Osteosarcoma in High Risk Breeds. Cancers 2020, 12, 2405. https://doi.org/10.3390/cancers12092405
Simpson S, Dunning M, de Brot S, Alibhai A, Bailey C, Woodcock CL, Mestas M, Akhtar S, Jeyapalan JN, Lothion-Roy J, et al. Molecular Characterisation of Canine Osteosarcoma in High Risk Breeds. Cancers. 2020; 12(9):2405. https://doi.org/10.3390/cancers12092405
Chicago/Turabian StyleSimpson, Siobhan, Mark Dunning, Simone de Brot, Aziza Alibhai, Clara Bailey, Corinne L. Woodcock, Madeline Mestas, Shareen Akhtar, Jennie N. Jeyapalan, Jennifer Lothion-Roy, and et al. 2020. "Molecular Characterisation of Canine Osteosarcoma in High Risk Breeds" Cancers 12, no. 9: 2405. https://doi.org/10.3390/cancers12092405