
Monocytes and Macrophages in Spondyloarthritis: Functional Roles and Effects of Current Therapies
by
 , and
, and
Sara Martínez-Ramos
1,2,†, 
Carlos Rafael-Vidal
1,2,†,
José M. Pego-Reigosa
1,2 and
Samuel García
1,2,* 1
Rheumatology & Immuno-Mediated Diseases Research Group (IRIDIS), Galicia Sur Health Research Institute (IIS Galicia Sur), SERGAS-UVIGO, 36312 Vigo, Spain
2
Rheumatology Department, University Hospital Complex of Vigo, 36214 Vigo, Spain
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Cells 2022, 11(3), 515; https://doi.org/10.3390/cells11030515
Submission received: 30 December 2021
/
Revised: 27 January 2022
/
Accepted: 28 January 2022
/
Published: 2 February 2022
(This article belongs to the Special Issue Innate Immune System in Immune-Mediated Inflammatory Diseases (IMIDs))
Abstract
:Spondyloarthritis (SpA) is a family of chronic inflammatory diseases, being the most prevalent ankylosing spondylitis (AS) and psoriatic arthritis (PsA). These diseases share genetic, clinical and immunological features, such as the implication of human leukocyte antigen (HLA) class I molecule 27 (HLA-B27), the inflammation of peripheral, spine and sacroiliac joints and the presence of extra-articular manifestations (psoriasis, anterior uveitis, enthesitis and inflammatory bowel disease). Monocytes and macrophages are essential cells of the innate immune system and are the first line of defence against external agents. In rheumatic diseases including SpA, the frequency and phenotypic and functional characteristics of both cell types are deregulated and are involved in the pathogenesis of these diseases. In fact, monocytes and macrophages play key roles in the inflammatory processes characteristics of SpA. The aim of this review is analysing the characteristics and functional roles of monocytes and macrophages in these diseases, as well as the impact of different current therapies on these cell types.
1. Introduction
Spondyloarthritis (SpA) are defined as a group of chronic inflammatory diseases that affect mainly the spine and joints, being the sacroiliac joint the most typically involved. These diseases, which affect to approximately 1% of the worldwide population, cause serious disorders, pain and disabilities, leading to significant health and socioeconomic problems. In contrast to other rheumatic diseases such as rheumatoid arthritis (RA) and systemic lupus erythematosus (SLE), the prevalence of spondylarthritis is similar between males and females and the onset of the disease (third-fourth decade) is earlier than other rheumatic joint diseases [1,2,3,4,5].
The diseases that are framed within the SpA are ankylosing spondylitis (AS), undifferentiated spondyloarthritis (USpA), axial spondyloarthritis (axSpA), psoriatic arthritis (PsA), reactive arthritis (ReA), enteropathic arthritis and peripheral SpA, being the most predominant AS and PsA [1,6,7,8]. These diseases can be grouped in axial or peripheral depending on the regions of the body that are affected [9], and they share genetic, clinical and immunological features, including joint inflammation (peripheral and axial skeleton), extra-articular manifestations and the absence of diagnostic autoantibodies [10]. The joint affectation, mainly the sacroiliac joint, is a common sign in SpA, but some particularities can be related to the different diagnosis. In fact, low back pain is typically found in AS and UspA [11]; the conjunctivitis-urethritis-polyarthritis triad characterize ReA [12]; enteropathic arthritis patients present chronic bowel inflammation [13]; axial skeleton (spine and the sacroiliac joints) involvement defines axSpA; psoriasis is the hallmark of PsA [14] and peripheral arthritis with dactylitis is distinctive of both peripheral SpA and PsA [11,15]. Some of the most common extraarticular signs are psoriasis, anterior uveitis, enthesitis and inflammatory bowel disease, in order of frequency. Other comorbidities like osteoporosis, cardiovascular events, depression, hypertension and diabetes are also frequent in SpA patients [16,17,18,19]. It is also important to emphasize that, compared with the general population, PsA patients have higher mortality rates, being tumours, vascular disease and respiratory disorders the major causes of death [19].
Cells of the innate immune system represent the first barrier of defence against pathogens. Among these cell types are mucosal-associated invariant T (MAIT) cells, invariant natural killer T (iNKT) cells, gamma delta T cells (γδ T cells), innate lymphoid cells (ILCs), neutrophils, mast cells, eosinophils and monocytes and macrophages [20]. Monocytes and macrophages have an essential role in the activation of innate immune system and after recognition of pathogens release inflammatory cytokines such as TNF, IL-6, IL-1β and chemokines that activate and attract other immune cells to the inflammation sites. In addition, macrophages play an essential role in the defence against bacteria and external agents to degrade them to peptides, performing antigenic presentation at MHC I and II [21]. In addition, these cell types also play important roles in tissue repair and angiogenesis. Rheumatic diseases such as RA, SLE and SpA are characterized by an aberrant activation of immune cells, both from innate and adaptive system [21,22,23,24]. Given the relevance of monocytes and macrophages in the pathogenesis of SpA, this review aims to analyse the role and function of these cell types in these diseases, as well as the impact of different clinical treatments on monocyte and macrophage activity.
2. Genetic Factors
Genetic relationship has been established in the pathogenesis of SpA. The strongest association was found in the HLA-B27 gene, which encodes the human leukocyte antigen (HLA) class I molecule 27 (HLA-B27), which is the major risk factor for the development of SpA, especially AS and USpA [7,25]. Human leukocyte antigens (HLA), also known as Major Histocompatibility Complex (MHC), are responsible for the presentation of intracellular and extracellular peptides to immune system cells for subsequent activation. The HLA-B27 molecule belongs to MHC class I molecules, which present intracellular peptides to CD8+ T cells [26]. In addition, several HLA-B27 subtypes are identified with SpA disease phenotypes [27]. For example, HLA–B27 overexpression and the predominance of its subtypes B*2702, B*2705, and B*2707 are described in AS [28]. Remarkably, the presence of HLA-B27 gene is usually employed as a biomarker for early detection [29] and SpA patients HLA-B27 positive show more radiological signs of joint damage [26]
Besides that, HLA-C gene, which codifies HLA-C glycoprotein, is also involved in SpA genetics. HLA-C promotes, among others, the production of cytokines, osteoclast differentiation and monocyte activation. Additionally, single nucleotide polymorphisms (SNPs), such as rs10484554, rs13191343 or rs12191877, and risk alleles in HLA-C have been associated to PsA and its variable clinical manifestations. Higher levels of HLA-C protein are related to risk of suffering inflammatory bowel disease (IBD) [30,31,32,33].
Other genetic associations have been found between non-MHC genes and SpA, such as the endoplasmic reticulum aminopeptidase 1 gene (ERAP1), IL23 receptor (IL23R) and STAT3, which has been associated with AS by genome wide association studies (GWAS) [34]. ERAP1 polymorphisms could determine ERAP-1 enzymatic activity, thereby modifying HLA–B27 peptidome and regulating its implications in SpA [35]. Also, genes that encodes for pro-inflammatory genes such as IL12 and IL23A have been linked to PsA [36,37].
3. Epigenetics and Environmental Factors
Several epigenetic disbalances have been also associated to SpA pathogenesis. Methylation is one of the major epigenetic mechanisms involved in a wide range of diseases, including SpA. Aberrant methylation is consequence of an increased addition of methyl groups in certain gene regions. Thus, hypermethylation of promoter regions induces a decreased expression of genes, usually enhancing pathologic phenotypes [27]. GWAS studies have found differentially methylated positions in the HLA-DQB1 gene in AS patients. In addition, several hypermethylated genes have been described in SpA with respect to controls. Among them, hypermethylation of BCL11B, IRF8 and DNA methyltransferase genes such as DNMT1 was found in patients with AS with respect to healthy controls [38,39,40].
MicroRNAs (miRNAs) are also important epigenetic factors and multiple studies have reported dysregulation of different miRNAs in SpA patients. One of the most important works has found a signature of 13 miRNAs deregulated in monocytes, as well as 11 miRNAs deregulated in CD4+ T cells, in patients with axSpA compared to controls, which are implicated in the pathogenesis of SpA [41]. In this regard, correlations have been described between some miRNAs and certain clinical parameters of these diseases. For example, differential expression of miR-146a-5p, miR-125a-5p and miR-22-3p has been correlated in serum samples from SpA patients with TNF and C-reactive protein levels [42].
In addition, other related epigenetic marks have been reported in SpA. This is the case of histone modifications, as AS patients treated with a TNF inhibitor showed increased activity of the enzyme histone acetyl transferase, indicating increased acetylation and thus increased gene expression in these acetylated regions [43]. Also, increased levels of Histone Deacetylase 3 (HDAC3) were found in the peripheral blood of AS patients [44,45].
Environmental factors are also involved in SpA and, for instance, previous infections have been associated with the development of some SpA. A link has been found between the gut flora microbiota and inflammation in these diseases as well [7,46,47]. In fact, a rat model of SpA has shown that intestinal inflammation, dependent of microbiota, enhances bone erosive potential in monocytes. Besides, bowel disruption promoted systemic inflammation, osteoclastogenesis and joint destruction [48]. Conversely, in vitro studies revealed that anti-IL-17 drugs contribute to dysbiosis and gut inflammation through inhibition of IL-17 pathway [49,50]. Finally, mechanical stress seems to be a determining factor in the appearance and development of SpA [51].
4. Etiopathogenesis
Despite the numerous studies in the field, the molecular mechanisms of action involved in the pathogenesis of SpA are still unclear [52]. There are currently different hypotheses, which can occur in combination and have in common that the trigger for autoinflammatory processes are mechanisms mediated by the HLA-B27 antigen. First hypothesis postulates that certain HLA-27 subclasses bind to peptides that are recognized by CD8+ cells, which leads to the activation of autoreactive T cells [46,53]. The second hypothesis suggests that defective HLA-B27 folding, occurring at the endoplasmic reticulum of immune cells, trigger the activation of the unfolding protein response (UPR) pathway, which induces the translocation and therefore activation of transcription factor NF-kB to the nucleus, leading to the production of cytokines involved in the pathogenesis of the disease by different inflammatory cells [46]. Related to this, it has been recently seen in macrophages that HLA–B27 impaired ubiquitination might promote the accumulation of misfolded HLA–B27 dimers, which are involved in the pathogenesis of SpA [54]. Moreover, murine models have revealed SpA-associated unconventional HLA-B27 molecules, detected in monocytes, resident and infiltrating macrophages [55]. The third hypothesis postulates that, since HLA-B27 tends to form homodimers, these are recognized as self-antigens by T cells and natural killer cells, leading to increase the production of cytokines such as IL-17 and IL-23 [46,52,56]. Also, a fourth hypothesis indicates that different polymorphisms in the aminopeptidases ERAP1 and ERAP2 genes would be involved in the production of aberrant forms of HLA-B27 and in the modification of peptides bound to HLA-B27. Finally, the most recent hypothesis links gut inflammation and dysbiosis with HLA-B27 and susceptibility to the development of SpA [52].
Overall, the mechanisms of action of these diseases involve the production of autoreactive T cells, through the activation of CD8+ T cells and through a key role of antigen presenting cells such as dendritic cells and macrophages. Following this activation, CD8+ T cells activate and perpetuate the inflammatory process, mainly through the release of key cytokines such as IL-17, IL-23, TNF, and IL-6; chemokines and other mediators such as Receptor Activator of Nuclear Factor-κB (RANK). The final consequences of these processes are the inflammation of the affected tissues (joints, skin, eyes, gut) and the destruction of the joints and spine, due to impaired osteoclastogenesis and bone remodelling [57].
5. Cell Pathology: Monocytes, Macrophages and Osteoclasts in Spondyloarthritis
5.1. Monocytes
Cells of the innate immune system are involved in the onset and development of SpA and, within this group, monocytes have been shown to play a key role in the pathogenesis of these diseases [58]. According with the established literature, there are three classes of monocytes, defined by the expression of the surface markers CD14 and CD16. The classical monocytes, which are the most abundant population (around 90%), show high CD14 expression but no CD16 (CD14++ CD16−), meanwhile the non-classical express a low level of CD14 together with high CD16 (CD14+CD16++). Finally, the intermediate monocytes present high levels of CD14 and low CD16 expression (CD14++CD16+) and are considered a transitional population between the classical and non-classical monocyte subsets [58,59]. However, these 3 populations of monocytes reveal unique characteristics in healthy individuals, as they are defined by different transcriptional profiles, possess different repertoires of cell surface receptor genes and distinct cytokine production patterns revealed by LPS activation [59].
Similarly to other diseases, including RA, there is a disbalance in the frequency of three monocyte subsets in SpA patients [60]. In contrast to RA patients, who showed an increase of intermediated monocytes [61], an increment in the frequency of classical monocytes has been observed in SpA [62]. In addition, another study found an expansion of CD14++ CD16+ CCR9+ CX3CR1+ CD59+ monocytes in the peripheral blood, synovial fluid, synovial tissue and bone marrow of AS patients. Importantly, this population positively correlated with disease activity parameters and levels of C-reactive protein, and these cells displayed more pronounced phagocytic activity in AS patients than in controls [63]. Besides the frequency of monocyte subset, the phenotypic characteristics of monocytes are also altered in SpA patients. For instance, monocytes from AS patients exhibit higher pro-inflammatory phenotypes secrete higher amounts of pro-inflammatory cytokines and proteomic analysis showed a higher activation of leucocyte extravasation vascular, endothelial growth factor, Janus kinase/signal transducer and activator of protein (JAK/STAT) and TLR pathways [64,65]. Related to this, the levels of TLR4 were higher in monocytes from SpA and PsA patients compared to healthy controls [66]. Moreover, monocytes from PsA patients show higher expression of the calcium-binding proteins S100A8/A9, which have a central role in controlling leukocyte trafficking and the metabolic processes of arachidonic acid [67].
On another note, the monocytes to lymphocytes ratio (MLR) was increased in patients with AS compared with non-radiographic axSpA, another of the findings that highlights the role of monocytes in SpA. Furthermore, this ratio was correlated with C-reactive protein (CRP) levels, erythrocyte sedimentation rate (ESR) levels and spine movements [68].
Lastly, it is important to highlight that the monocytes are of great importance in these diseases because they are precursors of two key cell types in SpA: macrophages and osteoclasts, as we will discuss in the next sections [69].
5.2. Macrophages
Macrophages are other cells belonging to innate immune system that are present in all tissues and body compartments and serve as the first line of defence against infection. They are the main phagocytic cells, but they are also antigen presenters and secrete cytokines involved in the immune system activation. Macrophages play also key roles in the maintenance of tissue homeostasis and are critical cells in the orchestration of chronic inflammation observed in different diseases, including SpA [24,60,70,71]. Regarding the ontology of this cell type, it was accepted during decades that all macrophages originate from circulating adult blood monocytes. However, works from last years have shown that tissue macrophages, including microglia, Kupffer cells, Langerhans cells and kidney, alveolar, heart and synovial macrophages, originate during embryonic development from the yolk sac, fetal liver or hematopoietic stem cells [72,73].
The phenotypic characteristics and functional capacities of macrophages are defined by the environmental factors, such as cytokines and pathogens that they are exposed during the differentiation process. Historically, macrophages have been classified into classically activated pro-inflammatory M1 or alternatively activated M2 wound-healing and immunosuppressive macrophages [71]. M1 macrophages, which are induced by IFN-γ, LPS and GM-CSF, are key components of host defence and are characterized by the expression of the pro-inflammatory cytokines IL-1β, IL-6, IL-12, IL-23 and TNF. The Th2 cytokines IL-4 and IL-13 induce the differentiation of wound-healing macrophages, which secrete components of the extracellular matrix and are involved in the tissue homeostasis. Finally, IL-10 drives immunosuppressive M2 macrophages that dampen the immune response and limit inflammation, playing this manner a regulatory role [21,70,71]. In addition to the different functional characteristics, M1 and M2 macrophages also express specific surface markers, such as CD80 and CD64 (M1) and CD200R and CD163 (M2) [74]. This is a useful classification for the in vitro differentiation of macrophages; however in vivo distinction is not easy, as macrophages can show characteristic of both M1 and M2 macrophages and they show a broad heterogeneity. In fact, recent studies have reported the existence of several macrophage subsets in the synovium of RA patients [21,75,76].
Phenotypic characterization of the SpA lining and sublining layers has shown an increased expression of the M2 macrophage marker CD163 compared to RA synovium [77,78]. Importantly, CD163 was also increased in the colonic mucosa of SpA and Crohn’s patients versus ulcerative colitis patients and healthy controls [78,79]. The expression of M1 macrophages is more controversial, as an initial work found a reduction of CD80 and CD86 expression in the synovium of SpA compared to RA patients [78], but further works have not validated this finding, neither difference in the expression of CD14, CD68 CD64 or CD200R [77,80]. In addition, CD163 synovial expression was correlated with clinical disease parameters, such as swollen joint count (SJC), serum CRP and ESR [78]. In vitro studies also support these findings, as the expression of the M2 markers CD163 and CD200R is induced in peripheral blood monocytes by the stimulation with the synovial fluid of SpA, but not RA patients. In addition, the synovial fluid of SpA patients showed a reduced expression of TNF, IL-1 and CXCL-10, mediators secreted by M1 macrophages [81]. Altogether, these data suggest that in SpA M2 macrophages predominate over M1 phenotype. These M2-like phenotypic characteristics may be responsible of the differences in the synovium of SpA and RA patients, such as the reduced infiltration of B and T cells, higher frequency of Th17 cells and the increased vascularity, with more tortuous vessels found in SpA patients [78,82].
However, despite the M2-like phenotype of SpA macrophages, these cells are able to produce inflammatory cytokines. CD163+ SpA macrophages express high levels of HLA-DR and secrete TNF, but not IL-10 [78]. Moreover, TNF induced the expression of inflammatory mediators by monocyte-derived macrophages from blood and synovial fluid (SF) of PsA patients, as well from monocytes of healthy controls differentiated with the SF of PsA patients. Interestingly, activation of Tie2 signalling enhanced the TNF-dependent expression of these inflammatory mediators [83,84]. As Tie2 is also involved in angiogenesis and Angiopoietin-2, one of the Tie2 receptors, is elevated in the synovium of SpA patients, macrophage Tie2 signalling may be essential for the pathogenesis of SpA [85,86].
Importantly, IL-17, a key cytokine in SpA pathogenesis, stimulates the production and expression of proinflammatory cytokines by human macrophages [87]. Macrophages also express mediators that are elevated in SpA and are involved in the perpetuation of inflammation, such as Ca2+ binding proteins S100A8 and S100A9 and HMGB1 (high mobility group box 1 proteins) [88,89], and angiogenic factors (vascular endothelial growth factor –VEGF- and basic fibroblast growth factor –BFGF-) that are highly expressed in the synovium of early PsA patients [85]. Finally, macrophages also contribute to the joint destruction through the production of matrix metalloproteinases, including MMP-2, MMP-3, MMP-7 and MMP-9 [90,91].
5.3. Osteoclasts
Homeostatic bone tissue remodelling is consequence of a tight balance between the levels of bone formation (mediated by osteoblasts) and bone resorption (induced by osteoclasts). In SpA there is an imbalance in bone tissue remodelling, which leads to bone destruction and resorption, mainly at the peripheral joints, but also to new bone formation in the spine triggering disk fusion.
Osteoblasts have a mesenchymal origin, meanwhile osteoclasts are multinucleated cells of hematopoietic origin. Osteoclast development is controlled by the interaction of TNF superfamily receptor-ligand pair known as Receptor Activator of Nuclear Factor-κB (RANK) and RANK ligand (RANKL), which are both necessary and sufficient requirement for osteoclast formation [92]. RANKL is over-expressed in SpA cells, notably in macrophages and memory T lymphocytes, but also in synovial fibroblasts and osteoblasts [57]. This enhanced expression of RANKL is mediated by different cytokines, such as TNF, IL-17, TGF-β and IL-22 [93,94]. Besides the IL-17 mediated RANKL expression, IL-17 also plays a direct role in bone resorption through the expression of RANK in osteoclast precursors [93,95]. The role of IL-23 on osteoclastogenesis seems more controversial and different studies have shown that its implication in this process is due to the induction of Th17 cells and Il-17 secretion, rather than a direct effect [95]. Interestingly, sera from patients with axSpA, which showed higher levels of TNF and IL-17 compared to healthy control, promoted the expression of RANK during the osteoclastogenesis process, highlighting the prominent role of both cytokines [96].
Other mediators have been involved in bone remodelling. For example, Macrophage-colony stimulating factor (M-CSF) and IL-34, which maintain macrophages homeostasis and regulates osteoclasts, are 1raised in PsA serum and these levels are associated with bone erosion [97]. In addition, inhibition of the share receptor (CSF-R1), reduced the severity of arthritis and the bone destruction in an arthritis mouse model [98].
6. SpA Treatments and Effect on Monocyte/Macrophage Function
The European Alliance of Associations for Rheumatology (EULAR) and the American College of Rheumatology (ACR) have recently stablished medication guidelines for treating Spondyloarthritis. To date, the first line of pharmacological treatment of axSpA is the non-steroidal anti-inflammatory drugs (NSAIDs) therapy, followed by glucocorticoids, while second and third lines correspond to Biological disease-modifying antirheumatic drugs (DMARDs), relegating Non-Biological DMARDs to the last option. According to guidelines, Biologicals DMARDs employed are TNF inhibitors, IL-17 inhibitors and JAK inhibitors, in order of preference. In PsA, the use of Non-Biological DMARDs prevails over NSAIDs and the administration of IL-12/23 inhibitors is suggested [99,100,101,102]. Nevertheless, new-targeted medications are being studied. Below these lines, we analyse their effects on monocytes and macrophages populations. Also, the mechanism of action and effects of treatments in monocytes and macrophages are summarized in Table 1.
6.1. NSAIDs
Despite non-steroidal anti-inflammatory drugs (NSAIDs) are the first-line drug treatment for SpA, not much evidence is found about the effect of these drugs in SpA monocytes/macrophages. NSAIDs inhibit the activity of cyclooxygenase (COX), which modulates prostaglandin E2 (PGE2) expression. PGE2 is an important early mediator of enthesitis, the hallmark of SpA, and is involved in the differentiation of Th17 cells [116], and, therefore, could activate monocytes/macrophages in an indirect manner. In addition, previous studies showed an immune modulatory role of NSAIDs, as these drugs attenuated inflammatory processes induced by macrophages and T cells, both key cell types in the pathogenesis of SpA [103].
However, a study in axSpA patients found that NSAID treatment did not modulate the monocytes secretion of IL-1β, IL-6, and TNF, in contrast with the effect of inhibitors (TNFi). These results suggest that NSAIDs are not important modulators of monocytes/macrophages function [65]. In contrast to this lack of action on dampening inflammation, NSAIDs minimize radiographic signs of spinal damage progression in axSpA [117,118] and this effect might be due to the reduced COX/PGE2 activity, which would reduce Th17 differentiation, leading to decreased osteoclastogenesis. Nevertheless, further studies are needed for validating this hypothesis.
6.2. Glucocorticoids
The administration of glucocorticoids is another line of treatment for SpA patients, but the utilization is controversial because of their adverse effects, as osteoporosis and increase of cardiovascular risk. However, a recent systematic review has demonstrated the efficacy of using glucocorticoids in the short term (≤6 months) in SpA. Importantly, no deaths or major adverse events were reported [119]. Glucocorticoids signal through the glucocorticoid receptor (GCR), which activate cellular pathways that modulate the activity of the transcription factors C/EBPβ, PPARs and NFκB. These transcription factors promote the expression of anti-inflammatory mediators [120] and inhibit the expression of inflammatory mediators, including prostaglandins (departing from COX2 transcription blocking) [119,121] and Phospholipase A2 in the arachidonic acid pathway [122]. The overall consequence is the resolution of the inflammation [123,124,125].
In monocytes, glucocorticoids increase IL-10 secretion [126] and deplete the proinflammatory CD16+ cells [104]. It has been proven that glucocorticoids also mitigate some of the effects of macrophage activation, such as the production of important macrophage mediators, including TNF and IL-6. However, macrophages produce large amounts of pro-inflammatory cytokines, and doses of corticosteroids that are potentially toxic if sustained for more than few days are needed for reducing this production [127]. TNF also takes part in intervertebral disc degeneration, suggesting that corticosteroids could reduce pain and, remarkably, the damage in intervertebral discs via reducing monocyte/macrophage TNF secretion [128]. Finally, it has recently been reported that glucocorticoids may act on macrophages inducing a phenotype involved in tissue repair, but this effect needs to be demosntrated in SpA pathogenesis [120].
6.3. Non-Biological Disease-Modifying Anti-Rheumatic Drugs
Non-Biologicals disease-modifying antirheumatic drugs (DMARDs) are widely used for the treatment of rheumatic diseases, including SpA. Methotrexate (MTX) is the most common DMARD and, due to its effect modulating cell-specific signalling pathways, inhibits important pro-inflammatory properties of different cell types involved in the pathogenesis of rheumatic diseases, including T cells, macrophages, endothelial cells and fibroblast-like synoviocytes [105,129]. In the context of this review, MTX induces apoptosis of monocytes and reduces the expression of IL-1β by monocyte precursors and the expression of Fcγ receptors on monocytes of RA patients, demonstrating an anti-inflammatory role [105,106].
In addition, MTX induces release of adenosine by different cell types and the binding of adenosine to the adenosine receptor (ADORA) 2a and 3 reduces the monocyte secretion of TNF and IL-6, and induces the polarization of macrophages towards an anti-inflammatory M2 phenotype. These data suggest an anti-inflammatory effect of MTX on monocyte/macrophages in an indirect manner [129,130]. However, another study showed that MTX dose-dependently induced the expression of IL-1, IL-6 and TNF in a monocytic cell line, which could be implicated in the adverse effect of MTX use in some rheumatic patients [131]. Finally, a study in 10 PsA patients showed that MTX treatment reduced the number of CD68+ macrophages in the synovial tissue, as well the expression of inflammatory mediators secreted by monocytes/macrophages, such as IL-1β, IL-8, TNF and MMP-3 [132].
6.4. Anti-TNF Treatments
TNF is one of the key cytokines in the pathogenesis of SpA and is involved in different pathogenic processes of the disease, like inflammation, angiogenesis and osteoclastogenesis. TNF signals through TNF receptors (TNFR), mainly TNFR1 and TNFR2. After receptor binding, TNF activates two different pathways. On one hand, TNF activates MAP kinase pathways, such as c-Jun N-terminal kinases (JNKs), which induces the translocation of the transcription factor AP-1 to the nucleus. On the other hand, TNF activates another signalling pathway leading to the phosphorylation and degradation of IκBα, which promote the nuclear translocation of NF-κB. Both transcription factors ultimately trigger the expression of pro-inflammatory, anti-apoptotic, angiogenic and cell proliferation genes, which most of them are involved in the pathogenesis of SpA [133]. For that reason, there are currently several approved anti-TNF drugs for the treatment of these diseases [134].
Etanercept was the first anti-TNF therapy approved for therapeutic use in SpA. This drug is a TNF receptor fusion protein and is approved in different spondyloarthropathies: PsA, AS and non-radiographic axSpA. Regarding its function in monocytes and macrophages, recent studies has shown that etanercept skewed macrophage polarization towards a M2 phenotype and that etanercept reduced the expression of LPS-induced expression of NF-κB target genes and the LPS-induced MMP9 activity in SpA monocytes [107,108,109].
Adalimumab and Infliximab are recombinant human monoclonal antibodies that blocks TNF. They are currently approved for the treatment of several SpA, specifically PsA, plaque psoriasis, AS, ulcerative colitis and Crohn’s disease [9,134]. Both TNF inhibitors (TNFi) restrict pathological angiogenesis and inflammatory cell infiltration in the synovium, including macrophage infiltration of the sublining layer [135]. In PsA patients, Adalimumab leads to a decrease in the number of tissue resident macrophages (CD163+), infiltrating macrophages (MRP8+) and early stage differentiated macrophages (MRP14+) [136], while Infliximab significantly reduces the CD68+ macrophage levels in synovial tissue of PsA patients [137,138]. Moreover, both TNFi also enhance in vitro nonclassical monocytes, decrease classical monocytes [139] and modulate macrophage polarization to M2 phenotype [109]. In vitro research studies have shown that both Adalimumab and Infliximab inhibit IL-12/IL-23 production in M1 macrophages through the formation of immune complexes, demonstrating a functional effect of these TNFi in these cell types [140]. Finally, Infliximab also inhibits the osteoclast resorptive activity in AS patients [141].
On the other hand, Certolizumab pegol and Golimumab are also two anti-TNF monoclonal antibody-based treatments used and approved for SpA. Previous research has shown that Certolizumab, as well as Infliximab and Adalimumab, induces differentiation of an immunosuppressive macrophage subtype that inhibits T-cell proliferation [142].
6.5. Anti-IL-17 Treatments
Since IL-17 is an essential cytokine involved in the pathogenesis of SpA, therapies against this cytokine have been developed in the last years. There are currently two approved anti-IL-17 treatments for SpA: Secukinumab and Ixekizumab. Secukinumab is a treatment whose target is the cytokine IL-17A and its structure is a fully human monoclonal IgG1k antibody. It has been validated in several SpA diseases, such as AS [143] and PsA [144]. Likewise, Ixekizumab is an anti-IL-17A drug and is a humanized IgG4 monoclonal antibody. Its efficacy has been validated in different SpA types, such as PsA [145] or AS [146]. Besides that, there are currently several anti-IL-17 drugs of clinical interest in development, such as Bimekizumab [147] and Afasevikumab [148].
Different macrophage subpopulations express several subunits of the IL-17 receptor (IL-17R). Macrophage IL-17 signalling is mediated through IL-17R and downstream signalling leads to the activation of C/EBP proteins and the nuclear translocation of NF-kB. This results in the release of a group of key cytokines such as IL12p70, GM-CSF, IL-3 and IL-9, representing a specific cytokine response profile [149,150]. Thus, direct blockade of IL-17 by binding to an anti-IL17 antibody prevents the ligation to IL-17R, inhibiting this manner the release of the aforementioned inflammatory mediators and therefore the pathogenic pathways that trigger macrophage activation and stimulation. The functional consequences in the context of SpA would be the abrogation of monocyte/macrophage activation and the reduction of IL-17-mediated osteoclastogenesis [110,111]. In fact, recent studies reveal that anti-IL17 drugs, in particular Secukinumab, reduce macrophage infiltration and MMP-3 expression, besides controlling disease signs, all without compromising systemic immune response [112].
6.6. Anti-IL-12/Anti-IL-23 Therapy
IL-12 and IL-23 are cytokines involved in the pathogenesis of autoimmune and immune-mediated diseases. Both cytokines present the p40 subunit and play a key role in immune cell regulation. IL-12 primarily mediates Th1 responses, enhancing IFN-γ production by NK cells and T cells, leading ultimately to the skewing towards M1 macrophages. On the other hand, IL-23 is crucial for the Th17 differentiation and is involved in the production of IL-17A and IL-17F by NK cells. In terms of signalling mechanisms, both IL-12 and IL-23 share similar pathways, such as JAK2, TYK2, STAT1, STAT3, STAT4 and STAT5 [151,152].
Ustekinumab is a monoclonal antibody against IL-12 and IL-23 approved in ulcerative colitis, Crohn’s disease and PsA. Its mechanism of action consists in the binding of the antibody to the p40 subunit present in both IL-12 and IL-23, inhibiting both cytokines. Inhibition of this subunit is effective in SpA because the IL-23/IL-17 axis plays a key role in the pathogenesis of these diseases, and macrophages are involved in this process, as they are the main producers of this cytokine. Thus, on one hand, IL-23 blockade inhibits Th17 differentiation and a decrease in IL-17 levels. And, on the other hand, the blockade of IL-12 impairs Th1-lymphocytes differentiation [153].
In addition, a recent functional study of the effect of Ustekinumab on PsA has yielded important results. In this study, a significantly lower infiltration of CD68+ macrophages in the synovial sublining layer was found, which ameliorates the pathogenesis of the disease. Regarding expression of inflammatory mediators, Ustekinumab reduced the levels of IL-23p19 and MMP3 the in synovial tissue biopsies of these patients [113].
6.7. JAK Inhibitors
Janus kinase (JAK)/STAT pathways are essential in the inflammatory processes observed in rheumatic diseases [154]. There are different JAK family members (JAK1-JAK3 and TYK2), being JAK1 the most important. In the context of monocyte/macrophages, JAK/STAT pathways are involved in different processes. On one hand, JAK/STAT signals the response to cytokines involved in macrophages differentiation [155]. For example, the binding of IFN-γ to its receptor activates JAK/STAT pathways, leading to M1 macrophages differentiation. On the other hand, IFN-γ, alone or in combination with IL-12, signal through JAK2-Tyrosine kinase 2 (TYK2) or JAK1-JAK2 pathways and induce the release of TNF by macrophages, which contributes to the development of the disease [154].
Due to the prominent role of JAK/STAT pathways in the production of inflammatory mediators, JAK inhibitors (JAKi) have emerged in the last years as therapeutic options of great relevance for the treatment of several immune-mediated inflammatory diseases. JAKi reduce the production of IL-12 and IFN-γ, which promote a decrease in the levels of TNF. Besides that, JAK inhibition also can cause the reduction of other cytokine production directly or indirectly, such as IL-17, IL-23, IL-18, IL-1, IL-6, IL-7, IFN-α and IFN-β, improving the inflammatory status [114]. However, and despite multiple inflammatory cytokines signal through JAK/STAT, the JAKi do not appear to have a direct effect on cytokine targets such as TNF or IL-17. The mechanism of action of JAKi in rheumatic diseases might be the interaction with alternative cytokine pathways [154].
Since JAK receptors are also involved in different proinflammatory signalling pathways observed in the pathogenesis of SpA, JAKi are promising therapeutic options [156]. In fact, Tofacitinib, a JAK1 and JAK3 inhibitor, is a drug already approved in PsA [157] and with promising results in a phase III trial in AS [158]. However, the effect of Tofacitinib on SpA monocyte/macrophages activation is still unknown and further research is needed in this context.
6.8. Other Biological Therapies
6.8.1. CTLA4-Ig (Abatacept)
Abatacept, an immunoglobulin against Cytotoxic T-Lymphocyte Antigen 4 (CTLA4-Ig), is a selective modulator of T cells employed in SpA treatment [159]. Abatacept inhibits CD4+ T cell activation, but also it has been observed that Abatacept reduces the migratory capacity of monocytes from RA patients [115,160]. Moreover, Abatacept downregulates the macrophages TNF production induced by activated T cells [161] and a recent paper has shown that Abatacept is able to directly skew RA macrophages towards a M2 phenotype [162].
6.8.2. IL-6 Inhibitors
Despite the implications of IL-6 in SpA pathogenesis, its inhibition has completely failed [163,164], except in anti-TNF-refractory aggressive SpA [165]. The treatment weakness could happen, according to mouse model experiments, because the pathological mechanisms in tissue resident cells of only few SpA patients would be dependent of IL-6 [166].
6.9. Directed Therapies: From Monocytes and Macrophages to Disease Management
Directly targeting monocytes and macrophages or their pathways is a potential novel therapeutic in SpA, as they are both implicated in the pathophysiology of the diseases. Therefore, they are promising therapeutic approaches, but they are still not approved for the clinical use.
6.9.1. Granulocyte–Monocyte Colony Stimulating Factor (GM-CSF) Inhibition
In chronic inflammation, hematopoietic stem cells are redirected to myelopoiesis for granulocyte-monocyte progenitors (GMPs) formation. Particularly, in SpA, these GMPs gather in bone marrow, spleen and affected joints, contributing to disease progression. Furthermore, secreted granulocyte-monocyte colony stimulating factor (GM-CSF) is a cytokine essential for the proliferation and differentiation of myeloid cells, including monocytes. Innate lymphoid cells, IL-17A+ T cells and mast cells secrete granulocyte-monocyte colony stimulating factor (GM-CSF). As these cell types are elevated in SpA patients, consequently the levels of this cytokine are also elevated [167,168,169]. Functionally, GM-CSF also induces monocyte inflammatory activity [170]. Taking this into account, antibody blocking of GM-CSF reveals potential therapeutic value in SpA, as it is being tested in other rheumatic diseases [169,171].
In animal models, neutralizing antibodies reduces arthritis signs [172,173]. Some anti-GM- CSF has been already assessed in several rheumatic processes, including SpA, as Mavrilimumab [171,174], Namilumab [175,176] or Otilimab [177], while new molecules are now being tested in randomized controlled trials (ClinicalTrials.gov Identifier: NCT03622658; NCT04205851).
6.9.2. Apheresis
Granulocyte and monocyte apheresis, an extracorporeal therapy consisting of selective removal of monocytes and macrophages from blood, is a safe procedure that could have an application in SpA, as it is propose for PsA and other rheumatic diseases [178,179]. However, further studies are needed to fully elucidate the therapeutic use in SpA.
7. Concluding Remarks
In this review, we have summarized and analyzed the main molecular mechanisms involved in the pathogenesis of spondyloarthritis and the function of monocytes and macrophages in these diseases (Figure 1). As it has been observed, approved drugs for the treatment of spondyloarthropathies have a clear involvement at the molecular level with the mechanisms of action of monocytes and macrophages in these diseases. This indicates a preponderant role of monocytes and macrophages in SpA and highlights these cell types as a promising target for the development of new therapies.
Author Contributions
Conceptualization, S.G.; writing—review and editing, S.M.-R., C.R.-V., J.M.P.-R. and S.G. All authors have read and agreed to the published version of the manuscript.
Funding
C.R.-V. and S.M.-R. are supported by predoctoral fellowships from Xunta de Galicia (IN606A-2020/043 and IN606A-2021/024, respectively). S.G. is supported by the Miguel Servet program (CP19/00005) from the Instituto de Salud Carlos III (ISCIII) and the European Social Fund (“Investing in your future”).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The graphical abstract figure was created with BioRender.com.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Vanhoof, J.; Declerck, K.; Geusens, P. Prevalence of Rheumatic Diseases in a Rheumatological Outpatient Practice. Ann. Rheum. Dis. 2002, 61, 453–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braun, J.; Sieper, J. Ankylosing Spondylitis. Lancet 2007, 369, 1379–1390. [Google Scholar] [CrossRef]
- Capelusnik, D.; Ramiro, S.; Schneeberger, E.E.; Citera, G. Peripheral Arthritis and Higher Disease Activity Lead to More Functional Impairment in Axial Spondyloarthritis: Longitudinal Analysis from ESPAXIA. Semin. Arthritis Rheum. 2021, 51, 553–558. [Google Scholar] [CrossRef] [PubMed]
- Paramarta, J.E.; de Rycke, L.; Ambarus, C.A.; Tak, P.P.; Baeten, D. Undifferentiated Spondyloarthritis vs Ankylosing Spondylitis and Psoriatic Arthritis: A Real-Life Prospective Cohort Study of Clinical Presentation and Response to Treatment. Rheumatology 2013, 52, 1873–1878. [Google Scholar] [CrossRef] [Green Version]
- Boel, A.; López-Medina, C.; van der Heijde, D.; van Gaalen, F.A. Age at Onset in Axial Spondyloarthritis around the World: Data from the International ASAS-PERSPA Study. Ann. Rheum. Dis. 2021, 80, 735. [Google Scholar] [CrossRef]
- Van Tubergen, A. The Changing Clinical Picture and Epidemiology of Spondyloarthritis. Nat. Rev. Rheumatol. 2015, 11, 110–118. [Google Scholar] [CrossRef]
- Ehrenfeld, M. Geoepidemiology: The Environment and Spondyloarthropathies. Autoimmun. Rev. 2010, 9, A325–A329. [Google Scholar] [CrossRef]
- Fragoulis, G.E.; Liava, C.; Daoussis, D.; Akriviadis, E.; Garyfallos, A.; Dimitroulas, T. Inflammatory Bowel Diseases and Spondyloarthropathies: From Pathogenesis to Treatment. World J. Gastroenterol. 2019, 25, 2162–2176. [Google Scholar] [CrossRef]
- Sieper, J.; Poddubnyy, D. New Evidence on the Management of Spondyloarthritis. Nat. Rev. Rheumatol. 2016, 12, 282–295. [Google Scholar] [CrossRef]
- Lories, R.J. Advances in Understanding the Pathophysiology of Spondyloarthritis. Best Pract. Res. Clin. Rheumatol. 2018, 32, 331–341. [Google Scholar] [CrossRef]
- Rojas-Vargas, M.; Munoz-Gomariz, E.; Escudero, A.; Font, P.; Zarco, P.; Almodovar, R.; Gratacos, J.; Mulero, J.; Juanola, X.; Montilla, C.; et al. First Signs and Symptoms of Spondyloarthritis—Data from an Inception Cohort with a Disease Course of Two Years or Less (REGISPONSER-Early). Rheumatology 2009, 48, 404–409. [Google Scholar] [CrossRef] [Green Version]
- Wu, I.B.; Schwartz, R.A. Reiter’s Syndrome: The Classic Triad and More. J. Am. Acad. Dermatol. 2008, 59, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Holden, W.; Orchard, T.; Wordsworth, P. Enteropathic Arthritis. Rheum. Dis. Clin. North Am. 2003, 29, 513–530. [Google Scholar] [CrossRef]
- Coates, L.C.; Kavanaugh, A.; Mease, P.J.; Soriano, E.R.; Laura Acosta-Felquer, M.; Armstrong, A.W.; Bautista-Molano, W.; Boehncke, W.-H.; Campbell, W.; Cauli, A.; et al. Group for Research and Assessment of Psoriasis and Psoriatic Arthritis 2015 Treatment Recommendations for Psoriatic Arthritis. Arthritis Rheumatol. 2016, 68, 1060–1071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carron, P.; de Craemer, A.-S.; van den Bosch, F. Peripheral Spondyloarthritis: A Neglected Entity—State of the Art. RMD Open 2020, 6, e001136. [Google Scholar] [CrossRef] [PubMed]
- Mathieu, S.; Motreff, P.; Soubrier, M. Spondyloarthropathies: An Independent Cardiovascular Risk Factor? Jt. Bone Spine 2010, 77, 542–545. [Google Scholar] [CrossRef] [PubMed]
- Maguire, S.; Gallagher, P.; O’Shea, F. The Negative Impact of Depression in Women with Axial Spondyloarthropathy. Jt. Bone Spine 2022, 89, 105261. [Google Scholar] [CrossRef] [PubMed]
- Curtis, J.R.; Winthrop, K.; Bohn, R.L.; Suruki, R.; Siegel, S.; Stark, J.L.; Xie, F.; Yun, H.; Chen, L.; Deodhar, A. The Annual Diagnostic Prevalence of Ankylosing Spondylitis and Axial Spondyloarthritis in the United States Using Medicare and MarketScan Databases. ACR Open Rheumatol. 2021, 3, 743–752. [Google Scholar] [CrossRef]
- Colaco, K.; Widdifield, J.; Luo, J.; Rosen, C.F.; Alhusayen, R.; Paterson, J.M.; Campbell, W.; Tu, K.; Bernatsky, S.; Gladman, D.D.; et al. Trends in Mortality and Cause-Specific Mortality among Patients with Psoriasis and Psoriatic Arthritis in Ontario, Canada. J. Am. Acad. Dermatol. 2021, 84, 1302–1309. [Google Scholar] [CrossRef]
- Rosine, N.; Miceli-Richard, C. Innate Cells: The Alternative Source of IL-17 in Axial and Peripheral Spondyloarthritis? Front. Immunol. 2021, 11, 3206. [Google Scholar] [CrossRef]
- Ross, E.A.; Devitt, A.; Johnson, J.R. Macrophages: The Good, the Bad, and the Gluttony. Front. Immunol. 2021, 12, 3234. [Google Scholar] [CrossRef] [PubMed]
- Roszkowski, L.; Ciechomska, M. Tuning Monocytes and Macrophages for Personalized Therapy and Diagnostic Challenge in Rheumatoid Arthritis. Cells 2021, 10, 1860. [Google Scholar] [CrossRef] [PubMed]
- Tsokos, G.C.; Lo, M.S.; Reis, P.C.; Sullivan, K.E. New Insights into the Immunopathogenesis of Systemic Lupus Erythematosus. Nat. Rev. Rheumatol. 2016, 12, 716–730. [Google Scholar] [CrossRef] [PubMed]
- Ambarus, C.; Yeremenko, N.; Tak, P.P.; Baeten, D. Pathogenesis of Spondyloarthritis. Curr. Opin. Rheumatol. 2012, 24, 351–358. [Google Scholar] [CrossRef]
- McMichael, A.; Bowness, P. HLA-B27: Natural Function and Pathogenic Role in Spondyloarthritis. Arthritis Res. 2002, 4, S153–S158. [Google Scholar] [CrossRef]
- Bodis, G.; Toth, V.; Schwarting, A. Role of Human Leukocyte Antigens (HLA) in Autoimmune Diseases. Rheumatol. Ther. 2018, 5, 5–20. [Google Scholar] [CrossRef] [Green Version]
- Kavadichanda, C.G.; Geng, J.; Bulusu, S.N.; Negi, V.S.; Raghavan, M. Spondyloarthritis and the Human Leukocyte Antigen (HLA)-B*27 Connection. Front. Immunol. 2021, 12, 497. [Google Scholar] [CrossRef]
- Jeanty, C.; Sourisce, A.; Noteuil, A.; Jah, N.; Wielgosik, A.; Fert, I.; Breban, M.; André, C. HLA-B27 Subtype Oligomerization and Intracellular Accumulation Patterns Correlate with Predisposition to Spondyloarthritis. Arthritis Rheumatol. 2014, 66, 2113–2123. [Google Scholar] [CrossRef]
- Prajzlerová, K.; Grobelná, K.; Pavelka, K.; Šenolt, L.; Filková, M. An Update on Biomarkers in Axial Spondyloarthritis. Autoimmun. Rev. 2016, 15, 501–509. [Google Scholar] [CrossRef]
- Apps, R.; Qi, Y.; Carlson, J.M.; Chen, H.; Gao, X.; Thomas, R.; Yuki, Y.; del Prete, G.Q.; Goulder, P.; Brumme, Z.L.; et al. Influence of HLA-C Expression Level on HIV Control. Science 2013, 340, 87–91. [Google Scholar] [CrossRef] [Green Version]
- Chandran, V.; Bull, S.B.; Pellett, F.J.; Ayearst, R.; Pollock, R.A.; Gladman, D.D. Killer-Cell Immunoglobulin-like Receptor Gene Polymorphisms and Susceptibility to Psoriatic Arthritis. Rheumatology 2014, 53, 233–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siegel, R.J.; Bridges, S.L.; Ahmed, S. HLA-C: An Accomplice in Rheumatic Diseases. ACR Open Rheumatol. 2019, 1, 571–579. [Google Scholar] [CrossRef] [PubMed]
- Stuart, P.E.; Tejasvi, T.; Shaiq, P.A.; Kullavanijaya, P.; Qamar, R.; Raja, G.K.; Li, Y.; Voorhees, J.J.; Abecasis, G.R.; Elder, J.T.; et al. A Single SNP Surrogate for Genotyping HLA-C*06:02 in Diverse Populations. J. Investig. Dermatol. 2015, 135, 1177–1180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cortes, A.; Pulit, S.L.; Leo, P.J.; Pointon, J.J.; Robinson, P.C.; Weisman, M.H.; Ward, M.; Gensler, L.S.; Zhou, X.; Garchon, H.J.; et al. Major Histocompatibility Complex Associations of Ankylosing Spondylitis Are Complex and Involve Further Epistasis with ERAP1. Nat. Commun. 2015, 6, 7146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costantino, F.; Talpin, A.; Evnouchidou, I.; Kadi, A.; Leboime, A.; Said-Nahal, R.; Bonilla, N.; Letourneur, F.; Leturcq, T.; Ka, Z.; et al. ERAP1 Gene Expression Is Influenced by Nonsynonymous Polymorphisms Associated with Predisposition to Spondyloarthritis. Arthritis Rheumatol. 2015, 67, 1525–1534. [Google Scholar] [CrossRef] [PubMed]
- Costantino, F.; Breban, M.; Garchon, H.J. Genetics and Functional Genomics of Spondyloarthritis. Front. Immunol. 2018, 9, 2933. [Google Scholar] [CrossRef] [PubMed]
- Reveille, J.D. The Genetic Basis of Spondyloarthritis. Ann. Rheum. Dis. 2011, 70, i44–i50. [Google Scholar] [CrossRef]
- International Genetics of Ankylosing Spondylitis Consortium (IGAS). Identification of Multiple Risk Variants for Ankylosing Spondylitis through High-Density Genotyping of Immune-Related Loci. Nat. Genet. 2013, 45, 730–738. [Google Scholar] [CrossRef] [Green Version]
- Vecellio, M.; Roberts, A.R.; Cohen, C.J.; Cortes, A.; Knight, J.C.; Bowness, P.; Wordsworth, B.P. The Genetic Association of RUNX3 with Ankylosing Spondylitis Can Be Explained by Allele-Specific Effects on IRF4 Recruitment that Alter Gene Expression. Ann. Rheum. Dis. 2016, 75, 1534–1540. [Google Scholar] [CrossRef] [Green Version]
- Roberts, A.R.; Vecellio, M.; Chen, L.; Ridley, A.; Cortes, A.; Knight, J.C.; Bowness, P.; Cohen, C.J.; Wordsworth, B.P. An Ankylosing Spondylitis-Associated Genetic Variant in the IL23R-IL12RB2 Intergenic Region Modulates Enhancer Activity and Is Associated with Increased Th1-Cell Differentiation. Ann. Rheum. Dis. 2016, 75, 2150–2156. [Google Scholar] [CrossRef] [Green Version]
- Fogel, O.; Bugge Tinggaard, A.; Fagny, M.; Sigrist, N.; Roche, E.; Leclere, L.; Deleuze, J.F.; Batteux, F.; Dougados, M.; Miceli-Richard, C.; et al. Deregulation of MicroRNA Expression in Monocytes and CD4+ T Lymphocytes from Patients with Axial Spondyloarthritis. Arthritis Res. Ther. 2019, 21, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez-Sanchez, C.; Font-Ugalde, P.; Ruiz-Limon, P.; Lopez-Pedrera, C.; Castro-Villegas, M.C.; Abalos-Aguilera, M.C.; Barbarroja, N.; de la Rosa, I.A.; Lopez-Montilla, M.D.; Escudero-Contreras, A.; et al. Circulating MicroRNAs as Potential Biomarkers of Disease Activity and Structural Damage in Ankylosing Spondylitis Patients. Hum. Mol. Genet. 2018, 27, 875–890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toussirot, E.; Abbas, W.; Khan, K.A.; Tissot, M.; Jeudy, A.; Baud, L.; Bertolini, E.; Wendling, D.; Herbein, G. Imbalance between HAT and HDAC Activities in the PBMCs of Patients with Ankylosing Spondylitis or Rheumatoid Arthritis and Influence of HDAC Inhibitors on TNF Alpha Production. PLoS ONE 2013, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Wang, L. Role of Histone Deacetylase 3 in Ankylosing Spondylitis via Negative Feedback Loop with MicroRNA-130a and Enhancement of Tumor Necrosis Factor-1α Expression in Peripheral Blood Mononuclear Cells. Mol. Med. Rep. 2016, 13, 35–40. [Google Scholar] [CrossRef]
- Cherqaoui, B.; Crémazy, F.; Hue, C.; Garchon, H.J.; Breban, M.; Costantino, F. Epigenetics of Spondyloarthritis. Jt. Bone Spine 2020, 87, 565–571. [Google Scholar] [CrossRef]
- Dougados, M.; Baeten, D. Spondyloarthritis. Lancet 2011, 377, 2127–2137. [Google Scholar] [CrossRef]
- Asquith, M.J.; Stauffer, P.; Davin, S.; Mitchell, C.; Lin, P.; Rosenbaum, J.T. Perturbed Mucosal Immunity and Dysbiosis Accompany Clinical Disease in a Rat Model of Spondyloarthritis. Arthritis Rheumatol. 2016, 68, 2151–2162. [Google Scholar] [CrossRef] [Green Version]
- Ansalone, C.; Utriainen, L.; Milling, S.; Goodyear, C.S. Role of Gut Inflammation in Altering the Monocyte Compartment and Its Osteoclastogenic Potential in HLA–B27–Transgenic Rats. Arthritis Rheumatol. 2017, 69, 1807–1815. [Google Scholar] [CrossRef] [Green Version]
- Manasson, J.; Wallach, D.S.; Guggino, G.; Stapylton, M.; Badri, M.H.; Solomon, G.; Reddy, S.M.; Coras, R.; Aksenov, A.A.; Jones, D.R.; et al. Interleukin-17 Inhibition in Spondyloarthritis Is Associated with Subclinical Gut Microbiome Perturbations and a Distinctive Interleukin-25–Driven Intestinal Inflammation. Arthritis Rheumatol. 2020, 72, 645–657. [Google Scholar] [CrossRef]
- Wang, L.; Wang, Y.; Zhang, P.; Song, C.; Pan, F.; Li, G.; Peng, L.; Yang, Y.; Wei, Z.; Huang, F. Gut Microbiota Changes in Patients with Spondyloarthritis: A Systematic Review. Semin. Arthritis Rheum. 2021, in press. [Google Scholar] [CrossRef]
- Cambré, I.; Gaublomme, D.; Burssens, A.; Jacques, P.; Schryvers, N.; De Muynck, A.; Meuris, L.; Lambrecht, S.; Carter, S.; de Bleser, P.; et al. Mechanical Strain Determines the Site-Specific Localization of Inflammation and Tissue Damage in Arthritis. Nat. Commun. 2018, 9, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharip, A.; Kunz, J. Understanding the Pathogenesis of Spondyloarthritis. Biomolecules 2020, 10, 1461. [Google Scholar] [CrossRef] [PubMed]
- Stoll, M.L. Interactions of the Innate and Adaptive Arms of the Immune System in the Pathogenesis of Spondyloarthritis. Clin. Exp. Rheumatol. 2011, 29, 322–330. [Google Scholar] [PubMed]
- Navid, F.; Layh-Schmitt, G.; Sikora, K.A.; Cougnoux, A.; Colbert, R.A. The Role of Autophagy in the Degradation of Misfolded HLA-B27 Heavy Chains. Arthritis Rheumatol. 2018, 70, 746–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rysnik, O.; McHugh, K.; van Duivenvoorde, L.; van Tok, M.; Guggino, G.; Taurog, J.; Kollnberger, S.; Ciccia, F.; Baeten, D.; Bowness, P. Non-Conventional Forms of HLA-B27 Are Expressed in Spondyloarthritis Joints and Gut Tissue. J. Autoimmun. 2016, 70, 12–21. [Google Scholar] [CrossRef] [Green Version]
- Generali, E.; Bose, T.; Selmi, C.; Voncken, J.W.; Damoiseaux, J.G.M.C. Nature versus Nurture in the Spectrum of Rheumatic Diseases: Classification of Spondyloarthritis as Autoimmune or Autoinflammatory. Autoimmun. Rev. 2018, 17, 935–941. [Google Scholar] [CrossRef]
- Crotti, T.N.; Smith, M.D.; Weedon, H.; Ahern, M.J.; Findlay, D.M.; Kraan, M.; Tak, P.P.; Haynes, D.R. Receptor Activator NF-KappaB Ligand (RANKL) Expression in Synovial Tissue from Patients with Rheumatoid Arthritis, Spondyloarthropathy, Osteoarthritis, and from Normal Patients: Semiquantitative and Quantitative Analysis. Ann. Rheum. Dis. 2002, 61, 1047–1054. [Google Scholar] [CrossRef] [Green Version]
- Wong, K.L.; Tai, J.J.Y.; Wong, W.C.; Han, H.; Sem, X.; Yeap, W.H.; Kourilsky, P.; Wong, S.C. Gene Expression Profiling Reveals the Defining Features of the Classical, Intermediate, and Nonclassical Human Monocyte Subsets. Blood 2011, 118, 16–31. [Google Scholar] [CrossRef] [Green Version]
- Ziegler-Heitbrock, L.; Ancuta, P.; Crowe, S.; Dalod, M.; Grau, V.; Hart, D.N.; Leenen, P.J.M.; Liu, Y.J.; MacPherson, G.; Randolph, G.J.; et al. Nomenclature of Monocytes and Dendritic Cells in Blood. Blood 2010, 116, e74–e80. [Google Scholar] [CrossRef]
- Locati, M.; Curtale, G.; Mantovani, A. Diversity, Mechanisms, and Significance of Macrophage Plasticity. Annu. Rev. Pathol. Mech. Dis. 2020, 15, 123–147. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Zhang, L.; Yu, C.; Yang, X.-F.; Wang, H. Monocyte and Macrophage Differentiation: Circulation Inflammatory Monocyte as Biomarker for Inflammatory Diseases. Biomark. Res. 2014, 2, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Surdacki, A.; Sulicka, J.; Korkosz, M.; Mikoałjczyk, T.; Telesinśka-Jasiówka, D.; Klimek, E.; Kierzkowska, I.; Guzik, T.; Grodzicki, T.K. Blood Monocyte Heterogeneity and Markers of Endothelial Activation in Ankylosing Spondylitis. J. Rheumatol. 2014, 41, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Ciccia, F.; Guggino, G.; Zeng, M.; Thomas, R.; Ranganathan, V.; Rahman, A.; Alessandro, R.; Rizzo, A.; Saieva, L.; Macaluso, F.; et al. Proinflammatory CX3CR1+CD59+Tumor Necrosis Factor–Like Molecule 1A+Interleukin-23+ Monocytes Are Expanded in Patients with Ankylosing Spondylitis and Modulate Innate Lymphoid Cell 3 Immune Functions. Arthritis Rheumatol. 2018, 70, 2003–2013. [Google Scholar] [CrossRef] [PubMed]
- Wright, C.; Edelmann, M.; diGleria, K.; Kollnberger, S.; Kramer, H.; McGowan, S.; McHugh, K.; Taylor, S.; Kessler, B.; Bowness, P. Ankylosing Spondylitis Monocytes Show Upregulation of Proteins Involved in Inflammation and the Ubiquitin Proteasome Pathway. Ann. Rheum. Dis. 2009, 68, 1626–1632. [Google Scholar] [CrossRef] [PubMed]
- Conrad, K.; Wu, P.; Sieper, J.; Syrbe, U. In Vivo Pre-Activation of Monocytes in Patients with Axial Spondyloarthritis. Arthritis Res. Ther. 2015, 17, 179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Rycke, L.; Vandooren, B.; Kruithof, E.; de Keyser, F.; Veys, E.M.; Baeten, D. Tumor Necrosis Factor α Blockade Treatment Down-Modulates the Increased Systemic and Local Expression of Toll-like Receptor 2 and Toll-like Receptor 4 in Spondylarthropathy. Arthritis Rheum. 2005, 52, 2146–2158. [Google Scholar] [CrossRef]
- Aochi, S.; Tsuji, K.; Sakaguchi, M.; Huh, N.; Tsuda, T.; Yamanishi, K.; Komine, M.; Iwatsuki, K. Markedly Elevated Serum Levels of Calcium-Binding S100A8/A9 Proteins in Psoriatic Arthritis Are Due to Activated Monocytes/Macrophages. J. Am. Acad. Dermatol. 2011, 64, 879–887. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Deng, W.; Zheng, S.; Feng, F.; Huang, Z.; Huang, Q.; Guo, X.; Huang, Z.; Huang, X.; Pan, X.; et al. Relationship between Monocytes to Lymphocytes Ratio and Axial Spondyloarthritis. Int. Immunopharmacol. 2018, 57, 43–46. [Google Scholar] [CrossRef]
- Gulino, G.R.; Van Mechelen, M.; Lories, R. Cellular and Molecular Diversity in Spondyloarthritis. Semin. Immunol. 2021, 101521. [Google Scholar] [CrossRef]
- Sica, A.; Mantovani, A. Macrophage Plasticity and Polarization: In Vivo Veritas. J. Clin. Investig. 2012, 122, 787–795. [Google Scholar] [CrossRef]
- Mosser, D.M.; Edwards, J.P. Exploring the Full Spectrum of Macrophage Activation. Nat. Rev. Immunol. 2008, 8, 958–969. [Google Scholar] [CrossRef] [PubMed]
- Epelman, S.; Lavine, K.J.; Randolph, G.J. Origin and Functions of Tissue Macrophages. Immunity 2014, 41, 21–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Culemann, S.; Grüneboom, A.; Nicolás-Ávila, J.Á.; Weidner, D.; Lämmle, K.F.; Rothe, T.; Quintana, J.A.; Kirchner, P.; Krljanac, B.; Eberhardt, M.; et al. Locally Renewing Resident Synovial Macrophages Provide a Protective Barrier for the Joint. Nature 2019, 572, 670–675. [Google Scholar] [CrossRef] [PubMed]
- Ambarus, C.A.; Krausz, S.; van Eijk, M.; Hamann, J.; Radstake, T.R.D.J.; Reedquist, K.A.; Tak, P.P.; Baeten, D.L.P. Systematic Validation of Specific Phenotypic Markers for in Vitro Polarized Human Macrophages. J. Immunol. Methods 2012, 375, 196–206. [Google Scholar] [CrossRef]
- Tardito, S.; Martinelli, G.; Soldano, S.; Paolino, S.; Pacini, G.; Patane, M.; Alessandri, E.; Smith, V.; Cutolo, M. Macrophage M1/M2 Polarization and Rheumatoid Arthritis: A Systematic Review. Autoimmun. Rev. 2019, 18, 102397. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Wei, K.; Slowikowski, K.; Fonseka, C.Y.; Rao, D.A.; Kelly, S.; Goodman, S.M.; Tabechian, D.; Hughes, L.B.; Salomon-Escoto, K.; et al. Defining Inflammatory Cell States in Rheumatoid Arthritis Joint Synovial Tissues by Integrating Single-Cell Transcriptomics and Mass Cytometry. Nat. Immunol. 2019, 20, 928–942. [Google Scholar] [CrossRef]
- Ambarus, C.A.; Noordenbos, T.; de Hair, M.J.H.; Tak, P.P.; Baeten, D.L.P. Intimal Lining Layer Macrophages but Not Synovial Sublining Macrophages Display an IL-10 Polarized-like Phenotype in Chronic Synovitis. Arthritis Res. Ther. 2012, 14, R74. [Google Scholar] [CrossRef] [Green Version]
- Baeten, D.; Demetter, P.; Cuvelier, C.A.; Kruithof, E.; van Damme, N.; de Vos, M.; Veys, E.M.; de Keyser, F. Macrophages Expressing the Scavenger Receptor CD163: A Link between Immune Alterations of the Gut and Synovial Inflammation in Spondyloarthropathy. J. Pathol. 2002, 196, 343–350. [Google Scholar] [CrossRef] [Green Version]
- Rudwaleit, M.; Baeten, D. Ankylosing Spondylitis and Bowel Disease. Best Practice & Research Clinical Rheumatology 2006, 20, 451–471. [Google Scholar] [CrossRef]
- Alivernini, S.; Bruno, D.; Tolusso, B.; Bui, L.; Petricca, L.; Gigante, M.R.; Birra, D.; Fedele, A.L.; Peluso, G.; Federico, F.; et al. Differential Synovial Tissue Biomarkers among Psoriatic Arthritis and Rheumatoid Factor/Anti-Citrulline Antibody-Negative Rheumatoid Arthritis. Arthritis Res. Ther. 2019, 21, 116. [Google Scholar] [CrossRef] [Green Version]
- Vandooren, B.; Noordenbos, T.; Ambarus, C.; Krausz, S.; Cantaert, T.; Yeremenko, N.; Boumans, M.; Lutter, R.; Tak, P.P.; Baeten, D. Absence of a Classically Activated Macrophage Cytokine Signature in Peripheral Spondylarthritis, Including Psoriatic Arthritis. Arthritis Rheum. 2009, 60, 966–975. [Google Scholar] [CrossRef] [PubMed]
- Veale, D.J.; Fearon, U. What Makes Psoriatic and Rheumatoid Arthritis so Different? RMD Open 2015, 1, e000025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García Pérez, S.; Malvar Fernández, B.; Tak, P.P.; Baeten, D.L.; Reedquist, K.A. THU0517 Tie2 Signaling Induces A Pro-Inflammatory Phenotype in Rheumatoid Arthritis and Psoriatic Arthritis Macrophages. Ann. Rheum. Dis. 2014, 73, 362. [Google Scholar] [CrossRef]
- Kabala, P.A.; Malvar-Fernández, B.; Lopes, A.P.; Carvalheiro, T.; Hartgring, S.A.Y.; Tang, M.W.; Conde, C.; Baeten, D.L.; Sleeman, M.; Tak, P.P.; et al. Promotion of Macrophage Activation by Tie2 in the Context of the Inflamed Synovia of Rheumatoid Arthritis and Psoriatic Arthritis Patients. Rheumatology 2020, 59, 426–438. [Google Scholar] [CrossRef] [Green Version]
- Fearon, U.; Griosios, K.; Fraser, A.; Reece, R.; Emery, P.; Jones, P.F.; Veale, D.J. Angiopoietins, Growth Factors, and Vascular Morphology in Early Arthritis. J. Rheumatol. 2003, 30, 260–268. [Google Scholar]
- Van de Sande, M.G.H.; de Launay, D.; de Hair, M.J.H.; García, S.; van de Sande, G.P.M.; Wijbrandts, C.A.; Gerlag, D.M.; Reedquist, K.A.; Tak, P.P. Local Synovial Engagement of Angiogenic TIE-2 Is Associated with the Development of Persistent Erosive Rheumatoid Arthritis in Patients with Early Arthritis. Arthritis Rheum. 2013, 65, 3073–3083. [Google Scholar] [CrossRef]
- Jovanovic, D.V.; di Battista, J.A.; Martel-Pelletier, J.; Jolicoeur, F.C.; He, Y.; Zhang, M.; Mineau, F.; Pelletier, J.P. IL-17 Stimulates the Production and Expression of Proinflammatory Cytokines, IL-Beta and TNF-Alpha, by Human Macrophages. J. Immunol. 1998, 160, 3513–3521. [Google Scholar]
- Wang, S.; Song, R.; Wang, Z.; Jing, Z.; Wang, S.; Ma, J. S100A8/A9 in Inflammation. Front. Immunol. 2018, 9, 1298. [Google Scholar] [CrossRef]
- Taniguchi, N.; Kawakami, Y.; Maruyama, I.; Lotz, M. HMGB Proteins and Arthritis. Hum. Cell 2018, 31, 1–9. [Google Scholar] [CrossRef]
- Schinocca, C.; Rizzo, C.; Fasano, S.; Grasso, G.; La Barbera, L.; Ciccia, F.; Guggino, G. Role of the IL-23/IL-17 Pathway in Rheumatic Diseases: An Overview. Front. Immunol. 2021, 12, 7–10. [Google Scholar] [CrossRef]
- Franco, G.C.N.; Kajiya, M.; Nakanishi, T.; Ohta, K.; Pedro, L.; Groppo, F.C.; Ernst, C.W.O.; Boyesen, J.L.; Bartlett, J.D.; Stashenko, P.; et al. Inhibition of Matrix Metalloproteinase-9 Activity by Doxycycline Ameliorates RANK Ligand-Induced Osteoclast Differentiation in Vitro and in Vivo. Exp. Cell Res. 2012, 317, 1454–1464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, W.; Schwarz, E.M. Mechanisms of Bone Resorption and New Bone Formation in Spondyloarthropathies. Curr. Rheumatol. Rep. 2002, 4, 513–517. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.A.; Colbert, R.A. The Interleukin-23/Interleukin-17 Axis in Spondyloarthritis Pathogenesis: Th17 and Beyond. Arthritis Rheumatol. 2014, 66, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Adamopoulos, I.E.; Chao, C.; Geissler, R.; Laface, D.; Blumenschein, W.; Iwakura, Y.; McClanahan, T.; Bowman, E.P. Interleukin-17A Upregulates Receptor Activator of NF-ΚB on Osteoclast Precursors. Arthritis Res. Ther. 2010, 12, R29. [Google Scholar] [CrossRef] [Green Version]
- Gravallese, E.M.; Schett, G. Effects of the IL-23–IL-17 Pathway on Bone in Spondyloarthritis. Nat. Rev. Rheumatol. 2018, 14, 631–640. [Google Scholar] [CrossRef]
- Korkosz, M.; Czepiel, M.; Guła, Z.; Stec, M.; Wȩglarczyk, K.; Rutkowska-Zapała, M.; Gruca, A.; Lenart, M.; Baran, J.; Gasowski, J.; et al. Sera of Patients with Axial Spondyloarthritis (AxSpA) Enhance Osteoclastogenic Potential of Monocytes Isolated from Healthy Individuals. BMC Musculoskelet. Disord. 2018, 19, 1–9. [Google Scholar] [CrossRef]
- Dalbeth, N.; Pool, B.; Smith, T.; Callon, K.E.; Lobo, M.; Taylor, W.J.; Jones, P.B.; Cornish, J.; McQueen, F.M. Circulating Mediators of Bone Remodeling in Psoriatic Arthritis: Implications for Disordered Osteoclastogenesis and Bone Erosion. Arthritis Res. Ther. 2010, 12, R164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia, S.; Hartkamp, L.M.; Malvar-Fernandez, B.; van Es, I.E.; Lin, H.; Wong, J.; Long, L.; Zanghi, J.A.; Rankin, A.L.; Masteller, E.L.; et al. Colony-Stimulating Factor (CSF) 1 Receptor Blockade Reduces Inflammation in Human and Murine Models of Rheumatoid Arthritis. Arthritis Res. Ther. 2016, 18, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Heijde, D.; Ramiro, S.; Landewé, R.; Baraliakos, X.; van den Bosch, F.; Sepriano, A.; Regel, A.; Ciurea, A.; Dagfinrud, H.; Dougados, M.; et al. 2016 Update of the ASAS-EULAR Management Recommendations for Axial Spondyloarthritis. Ann. Rheum. Dis. 2017, 76, 978–991. [Google Scholar] [CrossRef]
- Singh, J.A.; Guyatt, G.; Ogdie, A.; Gladman, D.D.; Deal, C.; Deodhar, A.; Dubreuil, M.; Dunham, J.; Husni, M.E.; Kenny, S.; et al. 2018 American College of Rheumatology/National Psoriasis Foundation Guideline for the Treatment of Psoriatic Arthritis. Arthritis Rheumatol. 2019, 71, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Ward, M.M.; Deodhar, A.; Gensler, L.S.; Dubreuil, M.; Yu, D.; Khan, M.A.; Haroon, N.; Borenstein, D.; Wang, R.; Biehl, A.; et al. 2019 Update of the American College of Rheumatology/Spondylitis Association of America/Spondyloarthritis Research and Treatment Network Recommendations for the Treatment of Ankylosing Spondylitis and Nonradiographic Axial Spondyloarthritis. Arthritis Rheumatol. 2019, 71, 1599–1613. [Google Scholar] [CrossRef]
- Gossec, L.; Baraliakos, X.; Kerschbaumer, A.; de Wit, M.; McInnes, I.; Dougados, M.; Primdahl, J.; McGonagle, D.G.; Aletaha, D.; Balanescu, A.; et al. EULAR Recommendations for the Management of Psoriatic Arthritis with Pharmacological Therapies: 2019 Update. Ann. Rheum. Dis. 2020, 79, 700–712. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.Y. Immunomodulatory Effect of Nonsteroidal Anti-Inflammatory Drugs (NSAIDs) at the Clinically Available Doses. Arch. Pharmacal Res. 2007, 30, 64–74. [Google Scholar] [CrossRef] [PubMed]
- Dayyani, F.; Belge, K.-U.; Frankenberger, M.; Mack, M.; Berki, T.; Ziegler-Heitbrock, L. Mechanism of Glucocorticoid-Induced Depletion of Human CD14+ CD16+ Monocytes. J. Leukoc. Biol. 2003, 74, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Wijngaarden, S.; van Roon, J.A.G.; van de Winkel, J.G.J.; Bijlsma, J.W.J.; Lafeber, F.P.J.G. Down-Regulation of Activating Fcγ Receptors on Monocytes of Patients with Rheumatoid Arthritis upon Methotrexate Treatment. Rheumatology 2005, 44, 729–734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seitz, M.; Zwicker, M.; Loetscher, P. Effects of Methotrexate on Differentiation of Monocytes and Production of Cytokine Inhibitors by Monocytes. Arthritis Rheum. 1998, 41, 2032–2038. [Google Scholar] [CrossRef]
- Obeng, J.A.; Amoruso, A.; Camaschella, G.L.E.; Sola, D.; Brunelleschi, S.; Fresu, L.G. Modulation of Human Monocyte/Macrophage Activity by Tocilizumab, Abatacept and Etanercept: An in Vitro Study. Eur. J. Pharmacol. 2016, 780, 33–37. [Google Scholar] [CrossRef]
- Zhao, J.; Yuan, W.; Tao, C.; Sun, P.; Yang, Z.; Xu, W. M2 Polarization of Monocytes in Ankylosing Spondylitis and Relationship with Inflammation and Structural Damage. APMIS 2017, 125, 1070–1075. [Google Scholar] [CrossRef]
- Menegatti, S.; Guillemot, V.; Latis, E.; Yahia-Cherbal, H.; Mittermüller, D.; Rouilly, V.; Mascia, E.; Rosine, N.; Koturan, S.; Millot, G.A.; et al. Immune Response Profiling of Patients with Spondyloarthritis Reveals Signalling Networks Mediating TNF-Blocker Function in Vivo. Ann. Rheum. Dis. 2021, 80, 475–486. [Google Scholar] [CrossRef]
- Garcia-Montoya, L.; Marzo-Ortega, H. The Role of Secukinumab in the Treatment of Psoriatic Arthritis and Ankylosing Spondylitis. Ther. Adv. Musculoskelet. Dis. 2018, 10, 169–180. [Google Scholar] [CrossRef]
- Patel, D.D.; Lee, D.M.; Kolbinger, F.; Antoni, C. Effect of IL-17A Blockade with Secukinumab in Autoimmune Diseases. Ann. Rheum. Dis. 2013, 72, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Mens, L.J.J.; Sande, M.G.H.; Menegatti, S.; Chen, S.; Blijdorp, I.C.J.; Jong, H.M.; Fluri, I.A.; Latuhihin, T.E.; Kuijk, A.W.R.; Rogge, L.; et al. Brief Report: Interleukin-17 Blockade with Secukinumab in Peripheral Spondyloarthritis Impacts Synovial Immunopathology without Compromising Systemic Immune Responses. Arthritis Rheumatol. 2018, 70, 1994–2002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiechter, R.H.; de Jong, H.M.; van Mens, L.J.J.; Fluri, I.A.; Tas, S.W.; Baeten, D.L.P.; Yeremenko, N.G.; van de Sande, M.G.H. IL-12p40/IL-23p40 Blockade with Ustekinumab Decreases the Synovial Inflammatory Infiltrate through Modulation of Multiple Signaling Pathways Including MAPK-ERK and Wnt. Front. Immunol. 2021, 12, 1–18. [Google Scholar] [CrossRef]
- McInnes, I.B.; Szekanecz, Z.; McGonagle, D.; Maksymowych, W.P.; Pfeil, A.; Lippe, R.; Song, I.-H.; Lertratanakul, A.; Sornasse, T.; Biljan, A.; et al. A Review of JAK–STAT Signalling in the Pathogenesis of Spondyloarthritis and the Role of JAK Inhibition. Rheumatology 2021, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Bonelli, M.; Ferner, E.; Savitskaya, A.; Bluml, S.; Steiner, C.-W.; Rath, E.; Smolen, J.S.; Scheinecker, C. Effects of Abatacept on Monocytes in Patients with Rheumatoid Arthritis. Ann. Rheum. Dis. 2010, 69, A67. [Google Scholar] [CrossRef] [Green Version]
- Maseda, D.; Johnson, E.M.; Nyhoff, L.E.; Baron, B.; Kojima, F.; Wilhelm, A.J.; Ward, M.R.; Woodward, J.G.; Brand, D.D.; Crofford, L.J. MPGES1-Dependent Prostaglandin E2 (PGE2) Controls Antigen-Specific Th17 and Th1 Responses by Regulating T Autocrine and Paracrine PGE2 Production. J. Immunol. 2018, 200, 725–736. [Google Scholar] [CrossRef] [Green Version]
- Poddubnyy, D.; Rudwaleit, M.; Haibel, H.; Listing, J.; Märker-Hermann, E.; Zeidler, H.; Braun, J.; Sieper, J. Effect of Non-Steroidal Anti-Inflammatory Drugs on Radiographic Spinal Progression in Patients with Axial Spondyloarthritis: Results from the German Spondyloarthritis Inception Cohort. Ann. Rheum. Dis. 2012, 71, 1616–1622. [Google Scholar] [CrossRef] [Green Version]
- Kroon, F.P.; van der Burg, L.R.; Ramiro, S.; Landewé, R.B.; Buchbinder, R.; Falzon, L.; van der Heijde, D. Non-Steroidal Anti-Inflammatory Drugs (NSAIDs) for Axial Spondyloarthritis (Ankylosing Spondylitis and Non-Radiographic Axial Spondyloarthritis). Cochrane Database Syst. Rev. 2015. [Google Scholar] [CrossRef]
- Dhir, V.; Mishra, D.; Samanta, J. Glucocorticoids in Spondyloarthritis—Systematic Review and Real-World Analysis. Rheumatology 2021, 60, 4463–4475. [Google Scholar] [CrossRef]
- Desgeorges, T.; Caratti, G.; Mounier, R.; Tuckermann, J.; Chazaud, B. Glucocorticoids Shape Macrophage Phenotype for Tissue Repair. Front. Immunol. 2019, 10, 1591. [Google Scholar] [CrossRef]
- Rhen, T.; Cidlowski, J.A. Antiinflammatory Action of Glucocorticoids—New Mechanisms for Old Drugs. N. Engl. J. Med. 2005, 353, 1711–1723. [Google Scholar] [CrossRef] [Green Version]
- Gijón, M.A.; Leslie, C.C. Regulation of Arachidonic Acid Release and Cytosolic Phospholipase A2 Activation. J. Leukoc. Biol. 1999, 65, 330–336. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; McEachin, R.C.; States, D.J. Computationally Identifying Novel NF-ΚB-Regulated Immune Genes in the Human Genome. Genome Res. 2003, 13, 654–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knezevic, N.N.; Jovanovic, F.; Voronov, D.; Candido, K.D. Do Corticosteroids Still Have a Place in the Treatment of Chronic Pain? Front. Pharmacol. 2018, 9, 1229. [Google Scholar] [CrossRef]
- Sasse, S.K.; Gruca, M.; Allen, M.A.; Kadiyala, V.; Song, T.; Gally, F.; Gupta, A.; Pufall, M.A.; Dowell, R.D.; Gerber, A.N. Nascent Transcript Analysis of Glucocorticoid Crosstalk with TNF Defines Primary and Cooperative Inflammatory Repression. Genome Res. 2019, 29, 1753–1765. [Google Scholar] [CrossRef] [PubMed]
- Mozo, L.; Suarez, A.; Gutierrez, C. Glucocorticoids Up-Regulate Constitutive Interleukin-10 Production by Human Monocytes. Clin. Exp. Allergy 2004, 34, 406–412. [Google Scholar] [CrossRef] [PubMed]
- Udalova, I.A.; Mantovani, A.; Feldmann, M. Macrophage Heterogeneity in the Context of Rheumatoid Arthritis. Nat. Rev. Rheumatol. 2016, 12, 472–485. [Google Scholar] [CrossRef]
- Risbud, M.V.; Shapiro, I.M. Role of Cytokines in Intervertebral Disc Degeneration: Pain and Disc Content. Nat. Rev. Rheumatol. 2014, 10, 44–56. [Google Scholar] [CrossRef]
- Cronstein, B.N.; Aune, T.M. Methotrexate and Its Mechanisms of Action in Inflammatory Arthritis. Nat. Rev. Rheumatol. 2020, 16, 145–154. [Google Scholar] [CrossRef]
- Cronstein, B.N.; Sitkovsky, M. Adenosine and Adenosine Receptors in the Pathogenesis and Treatment of Rheumatic Diseases. Nat. Rev. Rheumatol. 2017, 13, 41–51. [Google Scholar] [CrossRef]
- Olsen, N.J.; Spurlock, C.F.; Aune, T.M. Methotrexate Induces Production of IL-1 and IL-6 in the Monocytic Cell Line U937. Arthritis Res. Ther. 2014, 16, R17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kane, D.; Gogarty, M.; O’Leary, J.; Silva, I.; Bermingham, N.; Bresnihan, B.; FitzGerald, O. Reduction of Synovial Sublining Layer Inflammation and Proinflammatory Cytokine Expression in Psoriatic Arthritis Treated with Methotrexate. Arthritis Rheum. 2004, 50, 3286–3295. [Google Scholar] [CrossRef] [PubMed]
- Parameswaran, N.; Patial, S. Tumor Necrosis Factor-α Signaling in Macrophages. Crit. Rev. Eukaryot. Gene Expr. 2010, 20, 87–103. [Google Scholar] [CrossRef]
- Menegatti, S.; Bianchi, E.; Rogge, L. Anti-TNF Therapy in Spondyloarthritis and Related Diseases, Impact on the Immune System and Prediction of Treatment Responses. Front. Immunol. 2019, 10, 382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baeten, D.; Kruithof, E.; van den Bosch, F.; Demetter, P.; van Damme, N.; Cuvelier, C.; de Vos, M.; Mielants, H.; Veys, E.M.; de Keyser, F. Immunomodulatory Effects of Anti-Tumor Necrosis Factor? Therapy on Synovium in Spondylarthropathy: Histologic Findings in Eight Patients from an Open-Label Pilot Study. Arthritis Rheum. 2001, 44, 186–195. [Google Scholar] [CrossRef]
- Van Kuijk, A.W.R.; Gerlag, D.M.; Vos, K.; Wolbink, G.; De Groot, M.; De Rie, M.A.; Zwinderman, A.H.; Dijkmans, B.A.C.; Tak, P.P. A Prospective, Randomised, Placebo-Controlled Study to Identify Biomarkers Associated with Active Treatment in Psoriatic Arthritis: Effects of Adalimumab Treatment on Synovial Tissue. Ann. Rheum. Dis. 2009, 68, 1303–1309. [Google Scholar] [CrossRef] [PubMed]
- Cañete, J.D.; Pablos, J.L.; Sanmartí, R.; Mallofré, C.; Marsal, S.; Maymó, J.; Gratacós, J.; Mezquita, J.; Mezquita, C.; Cid, M.C. Antiangiogenic Effects of Anti-Tumor Necrosis Factor α Therapy with Infliximab in Psoriatic Arthritis. Arthritis Rheum. 2004, 50, 1636–1641. [Google Scholar] [CrossRef]
- Kruithof, E.; Baeten, D.; van den Bosch, F.; Mielants, H.; Veys, E.M.; de Keyser, F. Histological Evidence that Infliximab Treatment Leads to Downregulation of Inflammation and Tissue Remodelling of the Synovial Membrane in Spondyloarthropathy. Ann. Rheum. Dis. 2005, 64, 529–536. [Google Scholar] [CrossRef] [Green Version]
- Batko, B.; Schramm-Luc, A.; Skiba, D.; Mikolajczyk, T.; Siedlinski, M. TNF-α Inhibitors Decrease Classical CD14hiCD16− Monocyte Subsets in Highly Active, Conventional Treatment Refractory Rheumatoid Arthritis and Ankylosing Spondylitis. Int. J. Mol. Sci. 2019, 20, 291. [Google Scholar] [CrossRef] [Green Version]
- Bloemendaal, F.M.; Koelink, P.J.; Van Schie, K.A.; Rispens, T.; Peters, C.P.; Buskens, C.J.; Van Der Bilt, J.D.; Bemelman, W.A.; Korf, H.; Sabino, J.G.; et al. TNF-Anti-TNF Immune Complexes Inhibit IL-12/IL-23 Secretion by Inflammatory Macrophages via an Fc-Dependent Mechanism. J. Crohn’s Colitis 2018, 12, 1122–1130. [Google Scholar] [CrossRef]
- Gengenbacher, M.; Sebald, H.-J.; Villiger, P.M.; Hofstetter, W.; Seitz, M. Infliximab Inhibits Bone Resorption by Circulating Osteoclast Precursor Cells in Patients with Rheumatoid Arthritis and Ankylosing Spondylitis. Ann. Rheum. Dis. 2007, 67, 620–624. [Google Scholar] [CrossRef]
- Vos, A.C.W.; Wildenberg, M.E.; Duijvestein, M.; Verhaar, A.P.; Van Den Brink, G.R.; Hommes, D.W. AntiTumor Necrosis Factor-α Antibodies Induce Regulatory Macrophages in an Fc Region-Dependent Manner. Gastroenterology 2011, 140, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Baeten, D.; Sieper, J.; Braun, J.; Baraliakos, X.; Dougados, M.; Emery, P.; Deodhar, A.; Porter, B.; Martin, R.; Andersson, M.; et al. Secukinumab, an Interleukin-17A Inhibitor, in Ankylosing Spondylitis. N. Engl. J. Med. 2015, 373, 2534–2548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McInnes, I.B.; Mease, P.J.; Kirkham, B.; Kavanaugh, A.; Ritchlin, C.T.; Rahman, P.; Van Der Heijde, D.; Landewé, R.; Conaghan, P.G.; Gottlieb, A.B.; et al. Secukinumab, a Human Anti-Interleukin-17A Monoclonal Antibody, in Patients with Psoriatic Arthritis (FUTURE 2): A Randomised, Double-Blind, Placebo-Controlled, Phase 3 Trial. Lancet 2015, 386, 1137–1146. [Google Scholar] [CrossRef] [Green Version]
- Mease, P.J.; Smolen, J.S.; Behrens, F.; Nash, P.; Liu Leage, S.; Li, L.; Tahir, H.; Gooderham, M.; Krishnan, E.; Liu-Seifert, H.; et al. A Head-to-Head Comparison of the Efficacy and Safety of Ixekizumab and Adalimumab in Biological-Naïve Patients with Active Psoriatic Arthritis: 24-Week Results of a Randomised, Open-Label, Blinded-Assessor Trial. Ann. Rheum. Dis. 2020, 79, 123–131. [Google Scholar] [CrossRef] [Green Version]
- Van der Heijde, D.; Cheng-Chung Wei, J.; Dougados, M.; Mease, P.; Deodhar, A.; Maksymowych, W.P.; Van den Bosch, F.; Sieper, J.; Tomita, T.; Landewé, R.; et al. Ixekizumab, an Interleukin-17A Antagonist in the Treatment of Ankylosing Spondylitis or Radiographic Axial Spondyloarthritis in Patients Previously Untreated with Biological Disease-Modifying Anti-Rheumatic Drugs (COAST-V): 16 Week Results of a Phase 3 Ra. Lancet 2018, 392, 2441–2451. [Google Scholar] [CrossRef] [Green Version]
- Wasilewska, A.; Winiarska, M.; Olszewska, M.; Rudnicka, L. Interleukin-17 Inhibitors. A New Era in Treatment of Psoriasis and Other Skin Diseases. Postepy Dermatol. I Alergol. 2016, 33, 247–252. [Google Scholar] [CrossRef] [Green Version]
- Chiricozzi, A.; De Simone, C.; Fossati, B.; Peris, K. Emerging Treatment Options for the Treatment of Moderate to Severe Plaque Psoriasis and Psoriatic Arthritis: Evaluating Bimekizumab and Its Therapeutic Potential. Psoriasis Targets Ther. 2019, 9, 73–74. [Google Scholar] [CrossRef] [Green Version]
- Rafael-Vidal, C.; Pérez, N.; Altabás, I.; Garcia, S.; Pego-Reigosa, J.M. Blocking IL-17: A Promising Strategy in the Treatment of Systemic Rheumatic Diseases. Int. J. Mol. Sci. 2020, 21, 7100. [Google Scholar] [CrossRef]
- Barin, J.G.; Baldeviano, G.C.; Talor, M.V.; Wu, L.; Ong, S.; Quader, F.; Chen, P.; Zheng, D.; Caturegli, P.; Rose, N.R.; et al. Macrophages Participate in IL-17-Mediated Inflammation. Eur. J. Immunol. 2012, 42, 726–736. [Google Scholar] [CrossRef] [Green Version]
- Bastos, K.R.B.; Marinho, C.R.F.; Barboza, R.; Russo, M.; Álvarez, J.M.; D’Império Lima, M.R. What Kind of Message Does IL-12/IL-23 Bring to Macrophages and Dendritic Cells? Microbes Infect. 2004, 6, 630–636. [Google Scholar] [CrossRef] [PubMed]
- McKenzie, B.S.; Kastelein, R.A.; Cua, D.J. Understanding the IL-23–IL-17 Immune Pathway. Trends Immunol. 2006, 27, 17–23. [Google Scholar] [CrossRef]
- Benson, J.M.; Peritt, D.; Scallon, B.J.; Heavner, G.A.; Shealy, D.J.; Giles-Komar, J.M.; Mascelli, M.A. Discovery and Mechanism of Ustekinumab: A Human Monoclonal Antibody Targeting Interleukin-12 and Interleukin-23 for Treatment of Immune-Mediated Disorders. mAbs 2011, 3, 535–545. [Google Scholar] [CrossRef]
- Spinelli, F.R.; Colbert, R.A.; Gadina, M. JAK1: Number One in the Family; Number One in Inflammation? Rheumatology 2021, 60, ii3–ii10. [Google Scholar] [CrossRef] [PubMed]
- Morris, R.; Kershaw, N.J.; Babon, J.J. The Molecular Details of Cytokine Signaling via the JAK/STAT Pathway. Protein Sci. 2018, 27, 1984–2009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malyshev, I.; Malyshev, Y. Current Concept and Update of the Macrophage Plasticity Concept: Intracellular Mechanisms of Reprogramming and M3 Macrophage “Switch” Phenotype. BioMed Res. Int. 2015, 2015, 1–22. [Google Scholar] [CrossRef] [Green Version]
- EMA announcement Tofacitinib for Psoriatic Arthritis (PsA) Is Now Approved by the European Commission. Rheumatology 2018, 57, e55. [CrossRef] [Green Version]
- Deodhar, A.; Sliwinska-Stanczyk, P.; Xu, H.; Baraliakos, X.; Gensler, L.S.; Fleishaker, D.; Wang, L.; Wu, J.; Menon, S.; Wang, C.; et al. Tofacitinib for the Treatment of Ankylosing Spondylitis: A Phase III, Randomised, Double-Blind, Placebo-Controlled Study. Ann. Rheum. Dis. 2021, 80, 1004–1013. [Google Scholar] [CrossRef]
- Olivieri, I.; D’Angelo, S.; Mennillo, G.A.; Pistone, G.; Scarano, E.; Padula, A. Abatacept in Spondyloarthritis Refractory to Tumour Necrosis Factor α Inhibition: Figure 1. Ann. Rheum. Dis. 2009, 68, 151–152. [Google Scholar] [CrossRef]
- Bonelli, M.; Ferner, E.; Göschl, L.; Blüml, S.; Hladik, A.; Karonitsch, T.; Kiener, H.P.; Byrne, R.; Niederreiter, B.; Steiner, C.W.; et al. Abatacept (CTLA-4IG) Treatment Reduces the Migratory Capacity of Monocytes in Patients with Rheumatoid Arthritis. Arthritis Rheum. 2013, 65, 599–607. [Google Scholar] [CrossRef]
- Wenink, M.H.; Santegoets, K.C.M.; Platt, A.M.; van den Berg, W.B.; van Riel, P.L.C.M.; Garside, P.; Radstake, T.R.D.J.; McInnes, I.B. Abatacept Modulates Proinflammatory Macrophage Responses upon Cytokine-Activated T Cell and Toll-like Receptor Ligand Stimulation. Ann. Rheum. Dis. 2012, 71, 80–83. [Google Scholar] [CrossRef] [PubMed]
- Cutolo, M.; Soldano, S.; Gotelli, E.; Montagna, P.; Campitiello, R.; Paolino, S.; Pizzorni, C.; Sulli, A.; Smith, V.; Tardito, S. CTLA4-Ig Treatment Induces M1–M2 Shift in Cultured Monocyte-Derived Macrophages from Healthy Subjects and Rheumatoid Arthritis Patients. Arthritis Res. Ther. 2021, 23, 306. [Google Scholar] [CrossRef] [PubMed]
- Sieper, J.; Porter-Brown, B.; Thompson, L.; Harari, O.; Dougados, M. Assessment of Short-Term Symptomatic Efficacy of Tocilizumab in Ankylosing Spondylitis: Results of Randomised, Placebo-Controlled Trials. Ann. Rheum. Dis. 2014, 73, 95–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sieper, J.; Braun, J.; Kay, J.; Badalamenti, S.; Radin, A.R.; Jiao, L.; Fiore, S.; Momtahen, T.; Yancopoulos, G.D.; Stahl, N.; et al. Sarilumab for the Treatment of Ankylosing Spondylitis: Results of a Phase II, Randomised, Double-Blind, Placebo-Controlled Study (ALIGN). Ann. Rheum. Dis. 2015, 74, 1051–1057. [Google Scholar] [CrossRef] [Green Version]
- Merashli, M.; de Marco, G.; Podgorski, M.; McGonagle, D.; Marzo-Ortega, H. Evidence of Response to IL-6 Inhibition in Some Cases of Refractory Spondyloarthritis-Associated Peripheral Synovitis. Ann. Rheum. Dis. 2016, 75, 1418–1420. [Google Scholar] [CrossRef]
- De Wilde, K.; Martens, A.; Lambrecht, S.; Jacques, P.; Drennan, M.B.; Debusschere, K.; Govindarajan, S.; Coudenys, J.; Verheugen, E.; Windels, F.; et al. A20 Inhibition of STAT1 Expression in Myeloid Cells: A Novel Endogenous Regulatory Mechanism Preventing Development of Enthesitis. Ann. Rheum. Dis. 2017, 76, 585–592. [Google Scholar] [CrossRef]
- Al-Mossawi, M.H.; Chen, L.; Fang, H.; Ridley, A.; de Wit, J.; Yager, N.; Hammitzsch, A.; Pulyakhina, I.; Fairfax, B.P.; Simone, D.; et al. Unique Transcriptome Signatures and GM-CSF Expression in Lymphocytes from Patients with Spondyloarthritis. Nat. Commun. 2017, 8, 1510. [Google Scholar] [CrossRef]
- Blijdorp, I.C.J.; Menegatti, S.; van Mens, L.J.J.; van de Sande, M.G.H.; Chen, S.; Hreggvidsdottir, H.S.; Noordenbos, T.; Latuhihin, T.E.; Bernink, J.H.; Spits, H.; et al. Expansion of Interleukin-22– and Granulocyte–Macrophage Colony-Stimulating Factor–Expressing, but Not Interleukin-17A–Expressing, Group 3 Innate Lymphoid Cells in the Inflamed Joints of Patients With Spondyloarthritis. Arthritis Rheumatol. 2019, 71, 392–402. [Google Scholar] [CrossRef]
- Regan-Komito, D.; Swann, J.W.; Demetriou, P.; Cohen, E.S.; Horwood, N.J.; Sansom, S.N.; Griseri, T. GM-CSF Drives Dysregulated Hematopoietic Stem Cell Activity and Pathogenic Extramedullary Myelopoiesis in Experimental Spondyloarthritis. Nat. Commun. 2020, 11, 155. [Google Scholar] [CrossRef]
- Shi, H.; Chen, L.; Ridley, A.; Zaarour, N.; Brough, I.; Caucci, C.; Smith, J.E.; Bowness, P. GM-CSF Primes Proinflammatory Monocyte Responses in Ankylosing Spondylitis. Front. Immunol. 2020, 11, 1520. [Google Scholar] [CrossRef]
- Cook, A.D.; Hamilton, J.A. Investigational Therapies Targeting the Granulocyte Macrophage Colony-Stimulating Factor Receptor-α in Rheumatoid Arthritis: Focus on Mavrilimumab. Ther. Adv. Musculoskelet. Dis. 2018, 10, 29–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cook, A.D.; Braine, E.L.; Campbell, I.K.; Rich, M.J.; Hamilton, J.A. Blockade of Collagen-Induced Arthritis Post-Onset by Antibody to Granulocyte-Macrophage Colony-Stimulating Factor (GM-CSF): Requirement for GM-CSF in the Effector Phase of Disease. Arthritis Res. 2001, 3, 293. [Google Scholar] [CrossRef] [PubMed]
- Cook, A.D.; Lee, M.-C.; Saleh, R.; Khiew, H.-W.; Christensen, A.D.; Achuthan, A.; Fleetwood, A.J.; Lacey, D.C.; Smith, J.E.; Förster, I.; et al. TNF and Granulocyte Macrophage-Colony Stimulating Factor Interdependence Mediates Inflammation via CCL17. JCI Insight 2018, 3, e99249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burmester, G.R.; McInnes, I.B.; Kremer, J.; Miranda, P.; Korkosz, M.; Vencovsky, J.; Rubbert-Roth, A.; Mysler, E.; Sleeman, M.A.; Godwood, A.; et al. A Randomised Phase IIb Study of Mavrilimumab, a Novel GM–CSF Receptor Alpha Monoclonal Antibody, in the Treatment of Rheumatoid Arthritis. Ann. Rheum. Dis. 2017, 76, 1020–1030. [Google Scholar] [CrossRef]
- Huizinga, T.W.J.; Batalov, A.; Stoilov, R.; Lloyd, E.; Wagner, T.; Saurigny, D.; Souberbielle, B.; Esfandiari, E. Phase 1b Randomized, Double-Blind Study of Namilumab, an Anti-Granulocyte Macrophage Colony-Stimulating Factor Monoclonal Antibody, in Mild-to-Moderate Rheumatoid Arthritis. Arthritis Res. Ther. 2017, 19, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, P.C.; Saurigny, D.; Vencovsky, J.; Takeuchi, T.; Nakamura, T.; Matsievskaia, G.; Hunt, B.; Wagner, T.; Souberbielle, B. Efficacy and Safety of Namilumab, a Human Monoclonal Antibody against Granulocyte-Macrophage Colony-Stimulating Factor (GM-CSF) Ligand in Patients with Rheumatoid Arthritis (RA) with Either an Inadequate Response to Background Methotrexate Therapy or an Inadequate Response or Intolerance to an Anti-TNF (Tumour Necrosis Factor) Biologic Therapy: A Randomized, Controlled Trial. Arthritis Res. Ther. 2019, 21, 101. [Google Scholar] [CrossRef] [Green Version]
- Behrens, F.; Tak, P.P.; Østergaard, M.; Stoilov, R.; Wiland, P.; Huizinga, T.W.; Berenfus, V.Y.; Vladeva, S.; Rech, J.; Rubbert-Roth, A.; et al. MOR103, a Human Monoclonal Antibody to Granulocyte–Macrophage Colony-Stimulating Factor, in the Treatment of Patients with Moderate Rheumatoid Arthritis: Results of a Phase Ib/IIa Randomised, Double-Blind, Placebo-Controlled, Dose-Escalation Trial. Ann. Rheum. Dis. 2015, 74, 1058–1064. [Google Scholar] [CrossRef] [Green Version]
- Kanekura, T.; Seishima, M.; Honma, M.; Etou, T.; Eto, H.; Okuma, K.; Okubo, Y.; Yamaguchi, Y.; Kambara, T.; Mabuchi, T.; et al. Therapeutic Depletion of Myeloid Lineage Leukocytes by Adsorptive Apheresis for Psoriatic Arthritis: Efficacy of a Non-Drug Intervention for Patients Refractory to Pharmacologics. J. Dermatol. 2017, 44, 1353–1359. [Google Scholar] [CrossRef]
- Kanekura, T. Clinical and Immunological Effects of Adsorptive Myeloid Lineage Leukocyte Apheresis in Patients with Immune Disorders. J. Dermatol. 2018, 45, 943–950. [Google Scholar] [CrossRef]
Figure 1.
Schematic overview of monocytes and macrophage roles in spondyloarthritis and effect of current therapies. MTX (methrotrexate), GC (glucocorticoids), JAKi (JAK inhibitors).
Figure 1.
Schematic overview of monocytes and macrophage roles in spondyloarthritis and effect of current therapies. MTX (methrotrexate), GC (glucocorticoids), JAKi (JAK inhibitors).


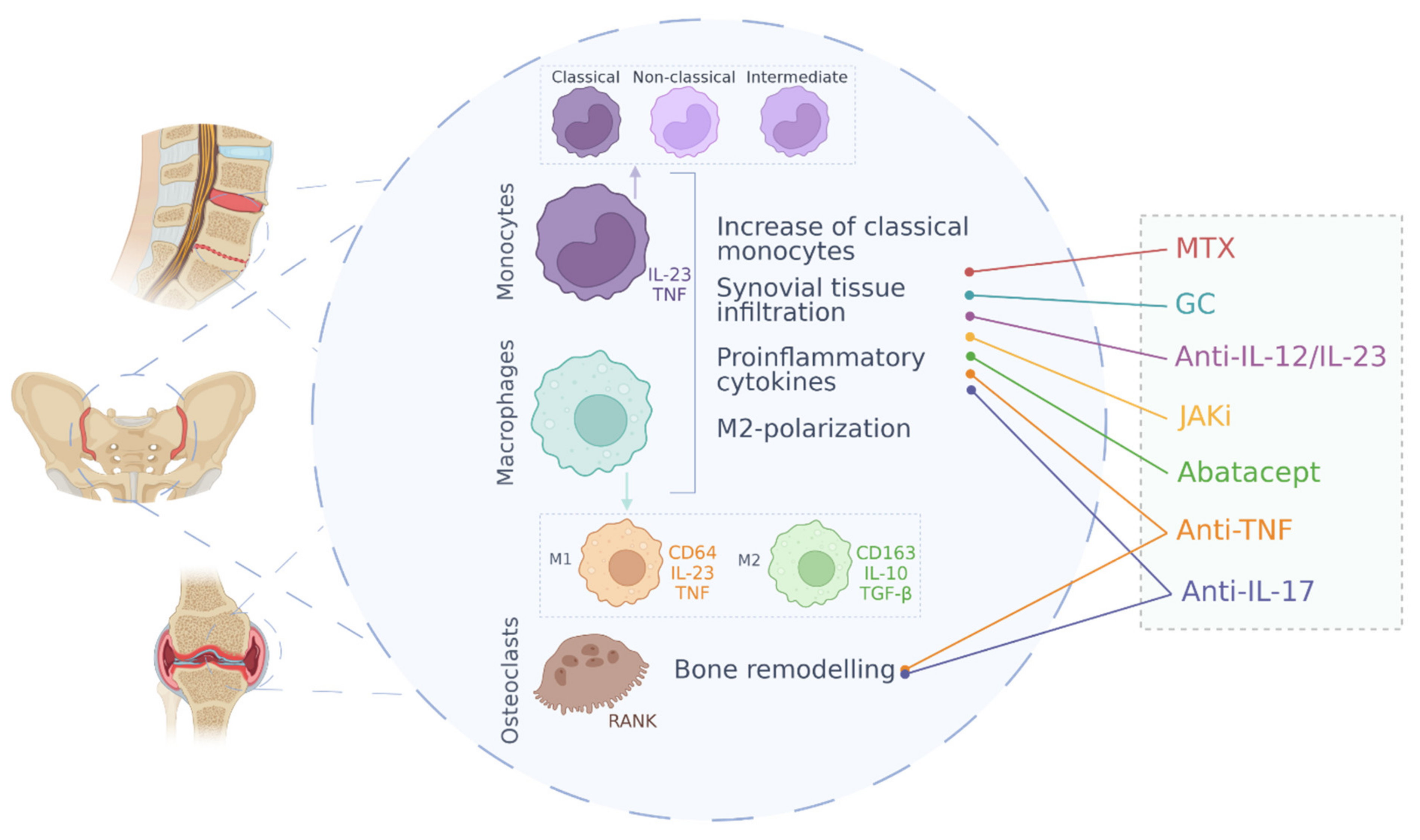{kind=link}
Table 1.
Treatments in SpA, mechanisms of action and its effects on monocytes and macrophages.
| Treatment Strategy | Drug | Mechanism of Action | Main Effects on Monocytes/Macrophages |
|---|---|---|---|
| NSAIDs | Meloxicam, Ibuprofen, Meclofenamate, etc. | Cyclooxygenase activity inhibition | Decreased activation of macrophages [103] |
| Glucocorticoids | Prednisolone, methylprednisolone | Phospholipase A2 suppression; NF-κB antagonization | Increased IL-10 secretion by monocytes; CD16+ cells depletion; restrained activity of pro-inflammatory cytokines [104] |
| Non-biological DMARDs | Methotrexate | Down-regulation of monocytes and macrophages activation; production of cytokine inhibitors by monocytes and IL-1β suppression [105,106] | |
| Anti-TNF | Adalimumab, Infliximab, Certolizumab pegol, Golimumab and Etanercept | TNF antibody blocking and TNF receptor blocking | Decrease of macrophage infiltration in the synovial tissue; decrease of expression levels of metalloproteinases; differentiation of immunosuppressive macrophages [107,108,109] |
| Anti-IL-17 | Secukinumab, Ixekizumab, Bimekizumab and Afasevikumab | IL-17 antibody blocking | Decrease of macrophages infiltration and MMPs expression; reduction of IL-17-mediated osteoclastogenesis [110,111,112] |
| Anti-IL-12/IL-23 | Ustekinumab | Antibody blocking of IL-12 and IL-23 cytokines | Lower infiltration of CD68+ macrophages in the synovial sublining layer [113] |
| JAK inhibitors | Tofacitinib | JAK1 and JAK3 inhibition | Decreased production of pro-inflammatory cytokines by macrophages [114] |
| CTLA4-Ig | Abatacept | Immunoglobulin against Cytotoxic T-Lymphocyte Antigen 4 | Decrease of TNF production; regulation of migratory capacity [115] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Martínez-Ramos, S.; Rafael-Vidal, C.; Pego-Reigosa, J.M.; García, S. Monocytes and Macrophages in Spondyloarthritis: Functional Roles and Effects of Current Therapies. Cells 2022, 11, 515. https://doi.org/10.3390/cells11030515
AMA Style
Martínez-Ramos S, Rafael-Vidal C, Pego-Reigosa JM, García S. Monocytes and Macrophages in Spondyloarthritis: Functional Roles and Effects of Current Therapies. Cells. 2022; 11(3):515. https://doi.org/10.3390/cells11030515
Chicago/Turabian StyleMartínez-Ramos, Sara, Carlos Rafael-Vidal, José M. Pego-Reigosa, and Samuel García. 2022. "Monocytes and Macrophages in Spondyloarthritis: Functional Roles and Effects of Current Therapies" Cells 11, no. 3: 515. https://doi.org/10.3390/cells11030515
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.