
Species Delimitation of Southeast Pacific Angel Sharks (Squatina spp.) Reveals Hidden Diversity through DNA Barcoding

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
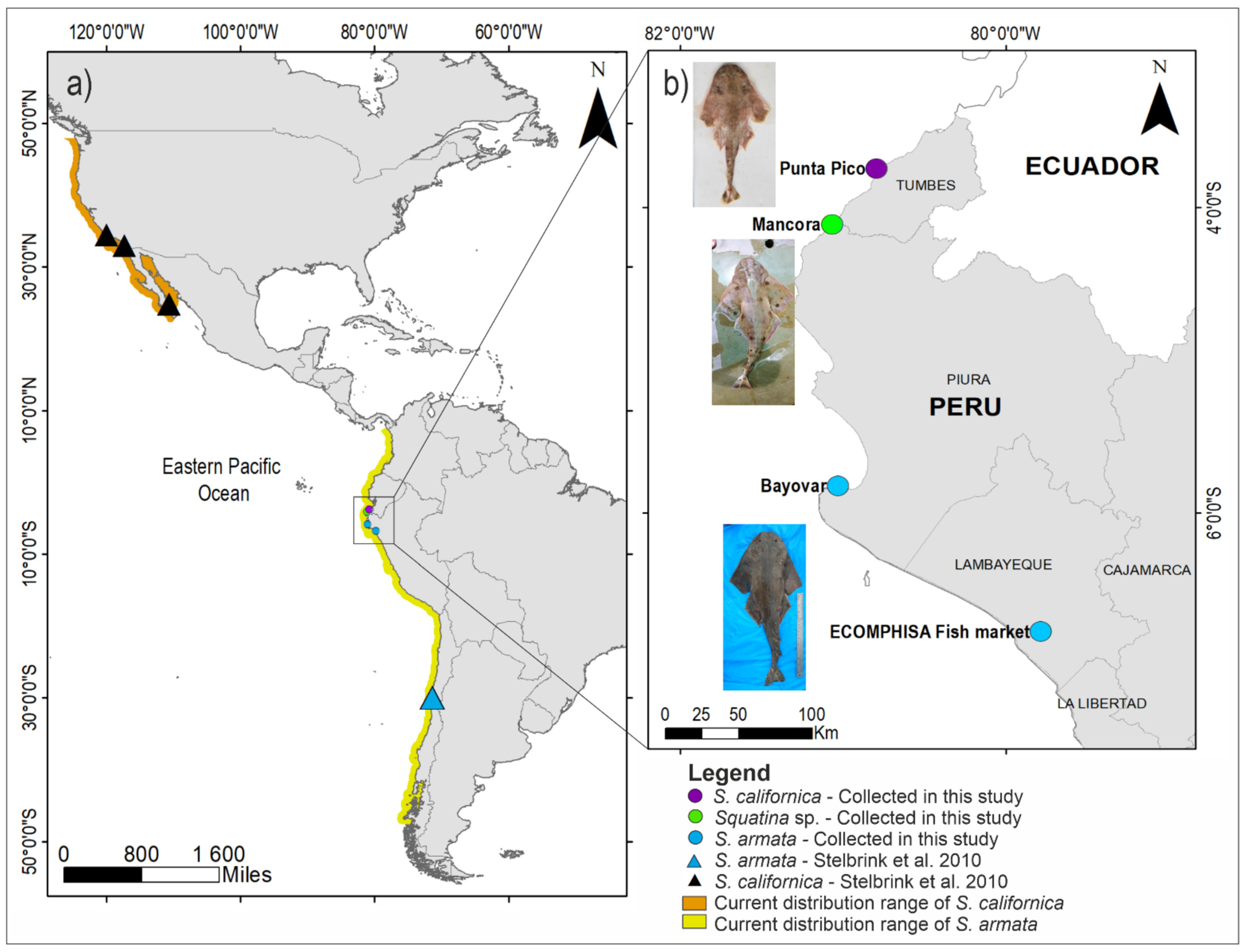
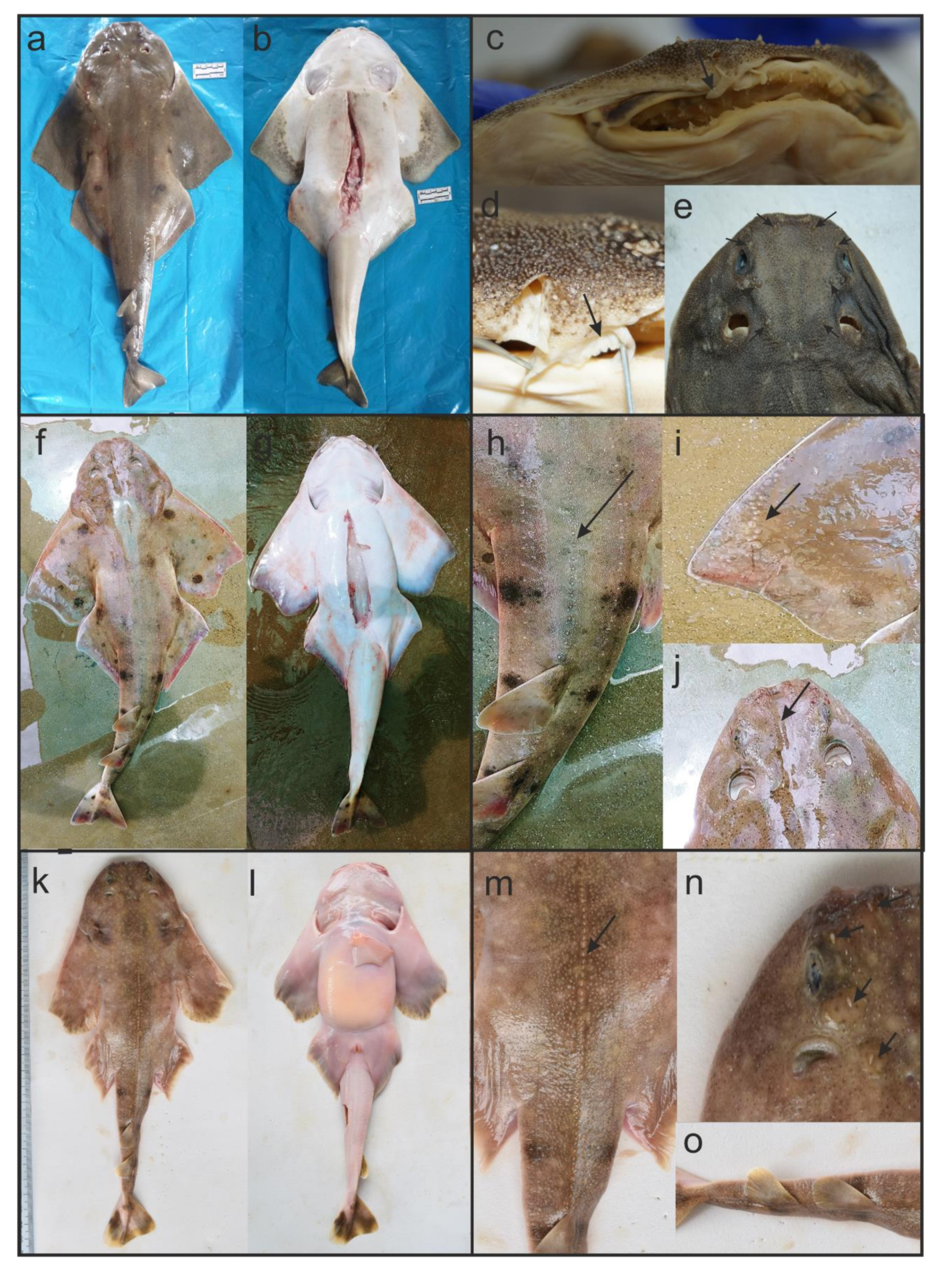
2.1. Morphological Identification and Sample Collection
2.2. DNA Extraction, Amplification and Sequencing
2.3. Species Delimitation Methods
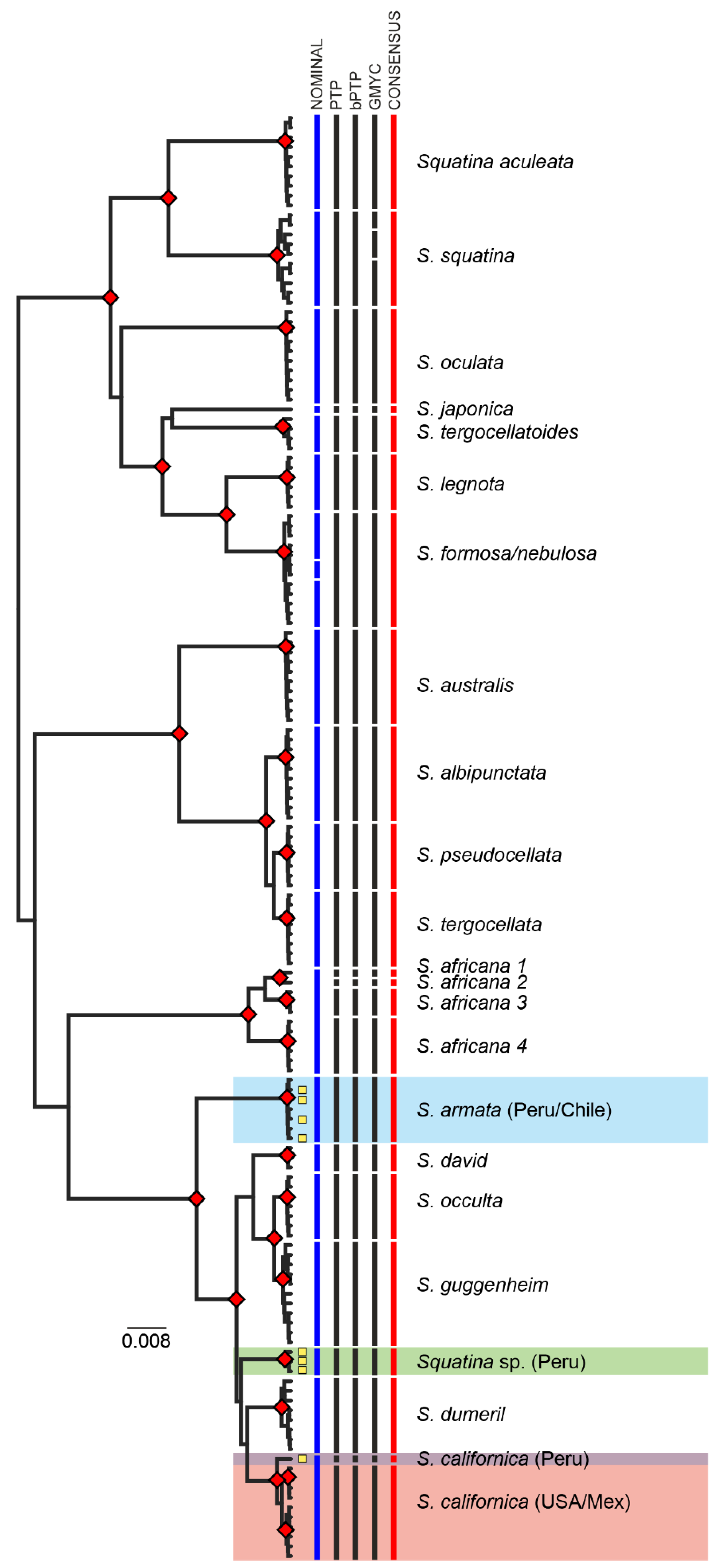
3. Results
3.1. Taxonomic Identification
3.2. MOTU Delimitation Analyses
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Compagno, L.J.V. FAO Species Catalogue—Volume 4, Sharks of the World: An Annotated and Illustrated Catalogue of Shark Species Known to Date. Part 1—Hexanchiformes to Lamniformes; FAO: Rome, Italy, 1984; pp. 503–512. [Google Scholar]
- Compagno, L.J.V. Sharks of the World: An Annotated and Illustrated Catalogue of Shark Species Known to Date—Volume 2, Bullhead, Mackerel and Carpet Sharks (Heterodontiformes, Lamniformes and Orectolobiformes); FAO Species Catalogue for Fishery Purposes: Rome, Italy, 2001; Volume 2, p. 278. [Google Scholar]
- Vélez-Zuazo, X.; Agnarsson, I. Molecular Phylogenetics and Evolution Shark tales: A molecular species-level phylogeny of sharks (Selachimorpha, Chondrichthyes). Mol. Phylogenet. Evol. 2011, 58, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Stelbrink, B.; Von Rintelen, T.; Cliff, G.; Kriwet, J. Molecular systematics and global phylogeography of angel sharks (genus Squatina). Mol. Phylogenet. Evol. 2010, 54, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Acero, P.A.; Tavera, J.J.; Anguila, R.; Hernández, L. A New Southern Caribbean Species of Angel Shark (Chondrichthyes, Squaliformes, Squatinidae), Including Phylogeny and Tempo of Diversification of American Species. Copeia 2016, 104, 577–585. [Google Scholar] [CrossRef]
- Vaz, D.F.B.; De Carvalho, M.R. New Species of Squatina (Squatiniformes: Squatinidae) from Brazil, with Comments on the Taxonomy of Angel Sharks from the Central and Northwestern Atlantic. Copeia 2018, 106, 144–160. [Google Scholar] [CrossRef]
- Weigmann, S. Annotated checklist of the living sharks, batoids and chimaeras (Chondrichthyes) of the world, with a focus on biogeographical diversity. J. Fish Biol. 2016, 88, 837–1037. [Google Scholar] [CrossRef]
- Castro-Aguirre, J.L.; Pérez, H.E.; Campos, L.H. Dos nuevas especies del género Squatina (Chondrichthyes: Squatinidae) del Golfo de México. Rev. Biol. Trop. 2006, 54, 1031–1040. [Google Scholar] [PubMed]
- Fricke, R.; Eschmeyer, W.N.; Van Der Laan, R. (Eds.) Eschmeyer’s Catalog of Fishes: Genera, Species, References. Available online: http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp (accessed on 5 January 2021).
- Ebert, D.A.; Fowler, S.L.; Compagno, L.J. Sharks of the World: A Fully Illustrated Guide; Wild Nature Press: Plymouth, UK, 2013. [Google Scholar]
- Del Moral-Flores, L.F.; Morrone, J.J.; Durand, J.A.; Espinosa-Pérez, H.; De León Pérez-Ponce, G. Lista patrón de los tiburones, rayas y quimeras de México. Arx. Miscellánia Zoológica 2015, 13, 47–163. [Google Scholar] [CrossRef] [Green Version]
- Ehemann, N.R.; del González-González, L.V.; Chollet-Villalpando, J.G.; De La Cruz-Agüero, J. Updated checklist of the extant Chondrichthyes within the Exclusive Economic Zone of Mexico. ZooKeys 2018, 774, 17–39. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Amaro, S.; Ramírez-Macías, D.; Vázquez-Juárez, R.; Flores-Ramírez, S.; Galván-Magaña, F.; Gutiérrez-Rivera, J.N. Estructura poblacional del tiburón ángel del Pacífico (Squatina californica) a lo largo de la costa noroccidental de México con base en la regíon control del ADN mitochondrial. Ciencias Mar. 2017, 43, 69–80. [Google Scholar] [CrossRef] [Green Version]
- Jacquet, J.; Alava, J.J.; Pramod, G.; Henderson, S.; Zeller, D. In hot soup: Sharks captured in Ecuador’s waters. Environ. Sci. 2008, 5, 269–283. [Google Scholar] [CrossRef] [Green Version]
- Chirichigno, N.; Cornejo, R. Catalogo Comentado Peces Marinos del Perú; Instituto del Mar del Peru: Callao, Peru, 2001; p. 217. [Google Scholar]
- Bustamante, C.; Vargas-Caro, C.; Bennett, M.B. Not all fish are equal: Functional biodiversity of cartilaginous fishes (Elasmobranchii and Holocephali) in Chile. J. Fish Biol. 2014, 85, 1617–1633. [Google Scholar] [CrossRef] [PubMed]
- Cornejo, R.; Vélez-Zuazo, X.; González-Pestana, A.; Kouri, C.; Mucientes, G. An updated checklist of Chondrichthyes from the southeast Pacific off Peru. Check List 2015, 17. [Google Scholar] [CrossRef] [Green Version]
- Calle-Morán, M.D.; Béarez, P. Updated checklist of marine cartilaginous fishes from continental and insular Ecuador (Tropical Eastern Pacific Ocean). Cybium Rev. Int. d’Ichtyologie. 2020, 44, 239–250. [Google Scholar]
- Mejía-Gallegos, J.; Flores-Portugal, L.A.; Segura, G. Exploración sobre Recursos Costeros y Recursos Demersales; Crucero 7104 B/I SNP 1; Instituto del Mar del Peru: Callao, Peru, 1971; p. 16. [Google Scholar]
- Espinoza, M.; Diaz, E.; Angulo, A.; Hernández, S.; Clarke, T.M. Chondrichthyan Diversity, Conservation Status, and Management Challenges in Costa Rica. Front. Mar. Sci. 2018, 5, 1–15. [Google Scholar] [CrossRef]
- Cailliet, G.M.; Chabot, C.L.; Nehmens, M.C.; Carlsle, A.B. Squatina californica (amended version of 2016 assessment). Available online: https://www.iucnredlist.org/species/39328/177163701 (accessed on 20 February 2021).
- Dulvy, N.K.; Acuña, E.; Bustamante, C.; Cavallos, A.; Herman, K.; Navia, A.F.; Pardo, S.A.; Velez-Zuazo, X. Squatina armata. Available online: https://www.iucnredlist.org/species/44571/116831653 (accessed on 20 February 2021).
- Chirichigno, N.; Velez, J. Clave para Identificar los Peces Marinos del Perú; Instituto del Mar del Perú: Callao, Peru, 1998; p. 31. [Google Scholar]
- Last, P.R.; White, W.T. Three new angel sharks (Chondrichthyes: Squatinidae) from the Indo-Australian region. Zootaxa 2008, 1734, 1–26. [Google Scholar] [CrossRef]
- Kato, S.; Springer, S.; Wagner, M.H. Field Guide to Eastern Pacific and Hawaiian Sharks; United States Department of the Interior: Washington, DC, USA, 1967; Volume 271, 47p.
- Ayres, W.O. On new fishes of the California coast. Proc. Calif. Acad. Sci. 1859, 2, 25–32. [Google Scholar]
- Philippi, R.A. Sobre los tiburones y algunos otros peces de Chile. An. Univ. Chile 1887, 71, 535–574. [Google Scholar]
- Ellis, J.R.; Barker, J.; McCully Phillips, S.R.; Meyers, E.K.M.; Heupel, M. Angel sharks (Squatinidae): A review of biological knowledge and exploitation. J. Fish Biol. 2021, 98, 592–621. [Google Scholar] [CrossRef]
- Domingues, R.R.; Hilsdorf, A.W.S.; Gadig, O.B.F. The importance of considering genetic diversity in shark and ray conservation policies. Conserv. Genet. 2017, 19, 501–525. [Google Scholar] [CrossRef] [Green Version]
- Hosegood, J.; Humble, E.; Ogden, R.; De Bruyn, M.; Creer, S.; Stevens, G.M.W.; Abudaya, M.; Bassos-Hull, K.; Bonfil, R.; Fernando, D.; et al. Phylogenomics and species delimitation for effective conservation of manta and devil rays. Mol. Ecol. 2020, 29, 4783–4796. [Google Scholar] [CrossRef]
- Walsh, J.H.; Ebert, D.A. A review of the systematics of western North Pacific angel sharks, genus Squatina, with redescriptions of Squatina formosa, S. japonica, and S. nebulosa (Chondrichthyes: Squatiniformes, Squatinidae). Zootaxa 2007, 1551, 31–47. [Google Scholar] [CrossRef]
- Walsh, J.H.; Ebert, D.A.; Compagno, L.J.V. Squatina caillieti sp. nov., a new species of angel shark (Chondrichthyes: Squat-iniformes: Squatinidae) from the Philippine Islands. Zootaxa 2011, 59, 49–59. [Google Scholar] [CrossRef] [Green Version]
- Theiss, S.M.; Ebert, D.A. Lost and found: Recovery of the holotype of the ocellated angelshark, Squatina tergocellatoides Chen, 1963 (Squatinidae), with comments on western Pacific squatinids. Zootaxa 2013, 3752, 73–85. [Google Scholar] [CrossRef] [Green Version]
- Vaz, D.F.B.; De Carvalho, M.R. Morphological and taxonomic revision of species of Squatina from the Southwestern Atlantic Ocean (Chondrichthyes: Squatiniformes: Squatinidae). Zootaxa 2013, 3695, 1–81. [Google Scholar] [CrossRef] [PubMed]
- Ward, R.D.; Zemlak, T.S.; Innes, B.H.; Last, P.R.; Hebert, P.D. DNA barcoding Australia’s fish species. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2005, 360, 1847–1857. [Google Scholar] [CrossRef]
- Ward, R.D.; Holmes, B.H.; White, W.T.; Last, P.R. DNA barcoding Australasian chondrichthyans: Results and potential uses in conservation. Mar. Freshw. Res. 2008, 59, 57–71. [Google Scholar] [CrossRef]
- Velez-Zuazo, X.; Alfaro-Shigueto, J.; Mangel, J.; Papa, R.; Agnarsson, I. What barcode sequencing reveals about the shark fishery in Peru. Fish. Res. 2015, 161, 34–41. [Google Scholar] [CrossRef]
- Borsa, P.; Arlyza, I.S.; Hoareau, T.B.; Shen, K.-N. Diagnostic description and geographic distribution of four new cryptic species of the blue-spotted maskray species complex (Myliobatoidei: Dasyatidae; Neotrygon spp.) based on DNA sequences. J. Oceanol. Limnol. 2017, 36, 827–841. [Google Scholar] [CrossRef] [Green Version]
- Smart, J.J.; Chin, A.; Baje, L.; Green, M.E.; Appleyard, S.A.; Tobin, A.J.; Simpfendorfer, C.A.; White, W.T. Effects of Including Misidentified Sharks in Life History Analyses: A Case Study on the Grey Reef Shark Carcharhinus amblyrhynchos from Papua New Guinea. PLoS ONE 2016, 11, e0153116. [Google Scholar] [CrossRef]
- Cariani, A.; Messinetti, S.; Ferrari, A.; Arculeo, M.; Bonello, J.J.; Bonnici, L.; Cannas, R.; Carbonara, P.; Cau, A.; Charilaou, C.; et al. Improving the Conservation of Mediterranean Chondrichthyans: The ELASMOMED DNA Barcode Reference Library. PLoS ONE 2017, 12, e0170244. [Google Scholar] [CrossRef]
- Bernardo, C.; Corrêa de Lima Adachi, A.M.; Paes da Cruz, V.; Foresti, F.; Loose, R.H.; Bornatowski, H. The label “Cação” is a shark or a ray and can be a threatened species! Elasmobranch trade in Southern Brazil unveiled by DNA barcoding. Mar. Policy 2020, 116, 103920. [Google Scholar] [CrossRef]
- Bunholi, I.V.; da Silva Ferrette, B.L.; De Biasi, J.B.; de Oliveira Magalhães, C.D.; Rotundo, M.M.; Oliveira, C.; Foresti, F.; Mendonça, F.F. The fishing and illegal trade of the angelshark: DNA barcoding against misleading identifications. Fish. Res. 2018, 206, 193–197. [Google Scholar] [CrossRef] [Green Version]
- Vella, A.; Vella, N.; Schembri, S. A molecular approach towards taxonomic identification of elasmobranch species from Maltese fisheries landings. Mar. Genom. 2017, 36, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Navia, A.F.; Mejía-Falla, P. Checklist of marine elasmobranchs of Colombia. Univ. Sci. 2019, 24, 241–276. [Google Scholar] [CrossRef] [Green Version]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets Brief Communication. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Stoeckle, M.Y.; Das Mishu, M.; Charlop-Powers, Z. GoFish: A streamlined environmental DNA presence/absence assay for marine vertebrates. bioRixv 2018, 331322. [Google Scholar] [CrossRef]
- Steinke, D.; Connell, A.D.; Hebert, P.D. Linking adults and immatures of South African marine fishes. Genome 2016, 59, 959–967. [Google Scholar] [CrossRef] [Green Version]
- Almerón-Souza, F.; Sperb, C.; Castilho, C.L.; Figueiredo, P.I.C.C.; Gonçalves, L.T.; Machado, R.; Oliveira, L.R.; Valiati, V.H.; Fagundes, N.J.R. Molecular Identification of Shark Meat from Local Markets in Southern Brazil Based on DNA Barcoding: Evidence for Mislabeling and Trade of Endangered Species. Front. Genet. 2018, 9, 1–12. [Google Scholar] [CrossRef]
- Fitzpatrick, C.K.; Finnegan, K.A.; Osaer, F.; Narváez, K.; Shivji, M.S. The complete mitochondrial genome of the Critically Endangered Angelshark, Squatina squatina. Mitochondrial DNA Part B Resour. 2017, 2, 212–213. [Google Scholar] [CrossRef]
- Moftah, M.; Aziz, S.H.A.; Elramah, S.; Favereaux, A. Classification of Sharks in the Egyptian Mediterranean Waters Using Morphological and DNA Barcoding Approaches. PLoS ONE 2011, 6, e27001. [Google Scholar] [CrossRef]
- Gao, Y.; Liu, T.; Wei, T.; Geng, X.; Wang, J.; Ma, H. Complete mitochondrial genome of Clouded angelshark (Squatina nebulosa). Mitochondrial DNA Part A 2016, 27, 1599–1600. [Google Scholar] [CrossRef]
- De Oliveira Ribeiro, A.; Caires, R.A.; Mariguela, T.C.; Pereira, L.H.G.; Hanner, R.; Oliveira, C. DNA barcodes identify marine fishes of São Paulo State, Brazil. Mol. Ecol. Resour. 2012, 12, 1012–1020. [Google Scholar] [CrossRef]
- Keskin, E.; Atar, H.H. DNA barcoding commercially important fish species of Turkey. Mol. Ecol. Resour. 2013, 13, 788–797. [Google Scholar] [CrossRef]
- Wang, Z.-D.; Guo, Y.-S.; Liu, X.-M.; Fan, Y.-B.; Liu, C.-W. DNA barcoding South China Sea fishes. Mitochondrial DNA 2012, 23, 405–410. [Google Scholar] [CrossRef]
- Sembiring, A.; Pertiwi, N.P.D.; Mahardini, A.; Wulandari, R.; Kurniasih, E.M.; Kuncoro, A.W.; Cahyani, N.D.; Anggoro, A.W.; Ulfa, M.; Madduppa, H.; et al. DNA barcoding reveals targeted fisheries for endangered sharks in Indonesia. Fish. Res. 2015, 164, 130–134. [Google Scholar] [CrossRef]
- Fujisawa, T.; Barraclough, T.G. Delimiting Species Using Single-Locus Data and the Generalized Mixed Yule Coalescent Approach: A Revised Method and Evaluation on Simulated Data Sets. Syst. Biol. 2013, 62, 707–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yule, G.U. A Mathematical Theory of Evolution, Based on the Conclusions of Dr. J.C. Willis, F.R.S. Philos. Trans. R. Soc. London Ser. B Contain. Pap. Biol. Character 1925, 213, 21–87. [Google Scholar]
- Hudson, R.R. Gene genealogies and the coalescent process. Oxford Surv. Evol. Biol. 1990, 7, 44. [Google Scholar]
- Zhang, J.; Kapli, P.; Pavlidis, P.; Stamatakis, A. A general species delimitation method with applications to phylogenetic placements. Bioinformatics 2013, 29, 2869–2876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouckaert, R.; Heled, J.; Kühnert, D.; Vaughan, T.; Wu, C.-H.; Xie, D.; Suchard, M.A.; Rambaut, A.; Drummond, A.J. BEAST 2: A Software Platform for Bayesian Evolutionary Analysis. PLoS Comput. Biol. 2014, 10, e1003537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), IEEE, New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar] [CrossRef] [Green Version]
- Gaida, I.H. Evolutionary Aspects of Gene Expression in the Pacific Angel Shark, Squatina california (Squatiniformes: Squatinidae). Copeia 1995, 532–554. [Google Scholar] [CrossRef]
- Gonzalez-Pestana, A.; Kouri, C.; Velez-Zuazo, X. Shark fisheries in the Southeast Pacific: A 61-year analysis from Peru. F1000Research 2014, 3, 164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- IMARPE. Resultados Preliminares del Primer Crucero de Exploración Pesquera del SNP-1 6901; Instituto del Mar del Perú: Callao, Peru, 1969; pp. 11–12. [Google Scholar]
- Chirichigno, N. Clave para identificar los peces marinos del Perú; Instituto del Mar del Perú: Callao, Peru, 1974; p. 30. [Google Scholar]
- Bartholomew, D.C.; Mangel, J.C.; Alfaro-Shigueto, J.; Pingo, S.; Jimenez, A.; Godley, B.J. Remote electronic monitoring as a potential alternative to on-board observers in small-scale fisheries. Biol. Conserv. 2018, 219, 35–45. [Google Scholar] [CrossRef]
- Cashion, M.S.; Bailly, N.; Pauly, D. Official catch data underrepresent shark and ray taxa caught in Mediterranean and Black Sea fisheries. Mar. Policy 2019, 105, 1–9. [Google Scholar] [CrossRef]
- Leet, W.S.; Dewees, C.M.; Klingbeil, R.; Larson, E.J. California’s Living Marine Resources: A Status Report 2001. Available online: https://wildlife.ca.gov/Conservation/Marine/Status/2001 (accessed on 18 January 2021).
- Pondella, D.J., II; Allen, L.G. The nearshore fish assemblage of Santa Catalina Island. In Proceedings of the Fifth California Islands Symposium, Santa Rosa Island, CA, USA, 29 March–1 April 1999; pp. 394–400. [Google Scholar]
- Bizzarro, J.J.; Smith, W.D.; Hueter, R.E.; Villavicencio–Garayzar, C.J. Activities and Catch Composition of Artisanal Elasmobranch Fishing Sites on the Eastern Coast of Baja California Sur, Mexico. Bull. South Calif. Acad. Sci. 2009, 108, 137–151. [Google Scholar] [CrossRef] [Green Version]
- Ambily, M.N.; Zacharia, P.U.; Najmudeen, T.M.; Ambily, L.; Sunil, K.T.S.; Radhakrishnan, M.; Kishor, T.G. First Record of African Angel Shark, Squatina africana (Chondricthyes: Squatinidae) in Indian Waters, Confirmed by DNA Barcoding. J. Ichthyol. 2018, 58, 312–317. [Google Scholar] [CrossRef]
- Von Der Heyden, S. Why do we need to integrate population genetics into South African marine protected area planning? Afr. J. Mar. Sci. 2009, 31, 263–269. [Google Scholar] [CrossRef]
- Spalding, M.D.; Fox, H.E.; Allen, G.R.; Davidson, N.; Ferdaña, Z.A.; Finlayson, M.; Halpern, B.S.; Jorge, M.A.; Lombana, A.; Lourie, S.A.; et al. Marine Ecoregions of the World: A Bioregionalization of Coastal and Shelf Areas. Bioscience 2007, 57, 573–583. [Google Scholar] [CrossRef] [Green Version]
- Von Der Heyden, S.; Prochazka, K.; Bowie, R.C.K.; Prochazka, K. Significant population structure and asymmetric gene flow patterns amidst expanding populations of Clinus cottoides (Perciformes, Clinidae): Application of molecular data to marine conservation planning in South Africa. Mol. Ecol. 2008, 17, 4812–4826. [Google Scholar] [CrossRef] [PubMed]
- Van der Merwe, A.E.B.; Gledhill, K.S. Molecular species identification and population genetics of chondrichthyans in South Africa: Current challenges, priorities and progress. Afr. Zoöl 2015, 50, 205–217. [Google Scholar] [CrossRef]
- Ramirez, J.L.; Rosas-Puchuri, U.; Cañedo, R.M.; Alfaro-Shigueto, J.; Ayon, P.; Zelada-Mázmela, E.; Siccha-Ramirez, R.; Velez-Zuazo, X. DNA barcoding in the Southeast Pacific marine realm: Low coverage and geographic representation despite high diversity. PLoS ONE 2020, 15, e0244323. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Species | Pemar Code | BOLD Code | Voucher Code | Sampling Site | Sex and Total Length |
|---|---|---|---|---|---|
| Squatina sp. | Pemar_V0209 | PMVTB124-21 | - | Piura/Mancora | Female, unknown TL |
| Squatina sp. | Pemar_V0210 | PMVTB125-21 | - | Piura/Mancora | Male, unknown TL |
| Squatina sp. | Pemar_V0211 | PMVTB126-21 | - | Piura/Mancora | Male, unknown TL |
| Squatina armata | Pemar_V0083 | PMVTB046-20 | - | Lambayeque/Terminal Pesquero ECOMPHISA | Female, 81 cm TL |
| Squatina armata | Pemar_V0173 | PMVTB067-20 | - | Piura/close to Bayovar Port | Female, 64.9 cm TL |
| Squatina armata | Pemar_V0174 | PMVTB068-20 | MUSM 65818 | Piura/close to Bayovar Port | Male, 37.2 cm TL |
| Squatina armata | Pemar_V0086 | PMVTB047-20 | - | Lambayeque/Terminal Pesquero ECOMPHISA | Female, 93.4 cm TL |
| Squatina californica | LCT_2160 | FMCT1223-19 | Tumbes/805 m off Punta Pico coast | Female, 23.7 cm TL |
| Extant Species | Distribution | BOLD System | |
|---|---|---|---|
| N° COI Sequences Used in This Study | Sampling Site (Country) | ||
| Squatina aculeata Cuvier 1829 | Mediterranean Sea; eastern Atlantic: southern Portugal south to Namibia, including Selvagens Islands (Portugal) | 10 | Malta, Senegal, Turkey |
| Squatina africana Regan 1908 | Western Indian Ocean: East Africa, South Africa to Madagascar | 11 | South Africa and Indian Ocean |
| Squatina albipunctata Last and White 2008 * | Australia: Queensland to Victoria | 10 | Australia |
| Squatina australis Regan 1906 | Southeastern Indian Ocean: Victoria, Tasmania, South Australia and Western Australia | 10 | Australia |
| Squatina armata (Philippi 1887) ** | Southeastern Pacific: Ecuador south to Chile | 3 | Chile |
| Squatina californica Ayres 1859 | Eastern Pacific: Puget Sound (Washington, DC, USA.) south to Pacific coast of Baja California Sur (Mexico); Ecuador south to Chile (needs confirmation); questionable from Alaska (USA) | 10 | Mexico, United States |
| Squatina david Acero P., Tavera, Anguila and Hernández 2016 *** | Western Atlantic: Panama, Colombia, Venezuela (southern Caribbean) | 3 | Colombia |
| Squatina dumeril Lesueur 1818 | Western Atlantic (including Caribbean Sea) | 8 | United States |
| Squatina formosa Shen and Ting 1972 | Western North Pacific | 10 | Taiwan, Japan |
| Squatina guggenheim Marini 1936**** | Southwestern Atlantic: Brazil south to Argentina | 11 | Brazil, Argentina |
| Squatina japonica Bleeker 1858 | Northwestern Pacific | 1 | Japan |
| Squatina legnota Last and White 2008 | Off southern Indonesia | 6 | Indonesia |
| Squatina nebulosa Regan 1906 | Western North Pacific | 2 | China, South China Sea |
| Squatina occulta Vooren and da Silva 1991 | Southwestern Atlantic: Brazil, Uruguay and Argentina | 7 | Brazil |
| Squatina oculata Bonaparte 1840 | Mediterranean Sea; eastern Atlantic: Portugal south to Namibia | 10 | Malta, Senegal, Turkey |
| Squatina pseudocellata Last and White 2008 | Australia: Western Australia | 7 | Australia |
| Squatina squatina (Linnaeus 1758) | Western Baltic Sea; North Sea; Mediterranean Sea; Black Sea; eastern Atlantic: Norway south to Western Sahara, including Canary Islands | 10 | Egypt, Ireland, Spain (Canary Islands), Turkey |
| Squatina tergocellata McCulloch 1914 | Southern and western Australia | 8 | Australia |
| Squatina tergocellatoides Chen 1963 | North Pacific: Taiwan Straits, Vietnam, Hong Kong, Malaysia | 4 | Malaysia, Vietnan |
| Squatina argentina (Marini 1930) | Southwestern Atlantic: Brazil to Uruguay and Argentina | no sequences | |
| Squatina caillieti Walsh, Ebert and Compagno 2011 | Philippines | no sequences | |
| Squatina varii Vaz and Carvalho 2018 | Brazil | no sequences | |
| Mean Intra- | Maximum Intra- | Distance to NN | NN | |
|---|---|---|---|---|
| Nominal | ||||
| Squatina aculeata | 0.06 | 0.16 | 6.22 | Squatina legnota |
| Squatina africana | 1.39 | 2.51 | 7.34 | Squatina sp. |
| Squatina albipunctata | 0 | 0 | 0.99 | Squatina pseudocellata |
| Squatina armata | 0 | 0 | 2.85 | Squatina guggenheim |
| Squatina australis | 0.03 | 0.16 | 5.33 | Squatina albipunctata |
| Squatina californica | 0.45 | 0.85 | 1.99 | Squatina david |
| Squatina david | 0.11 | 0.16 | 1.33 | Squatina guggenheim |
| Squatina dumeril | 0.43 | 0.99 | 2.52 | Squatina david |
| Squatina formosa | 0.13 | 0.33 | 0 | Squatina nebulosa |
| Squatina guggenheim | 0.31 | 0 | 6.99 | Squatina occulta |
| Squatina japonica | NaN | 0 | 6.99 | Squatina formosa |
| Squatina legnota | 0 | 0 | 2.51 | Squatina formosa |
| Squatina nebulosa | 0 | 0 | 0 | Squatina formosa |
| Squatina occulta | 0 | 0 | 0.50 | Squatina guggenheim |
| Squatina oculata | 0 | 0 | 5.87 | Squatina legnota |
| Squatina pseudocellata | 0 | 0 | 0.99 | Squatina albipunctata |
| Squatina squatina | 0.52 | 0.99 | 6.61 | Squatina aculeata |
| Squatina tergocellata | 0 | 0 | 0.99 | Squatina pseudocellata |
| Squatina tergocellatoides | 0.25 | 0.50 | 4.44 | Squatina legnota |
| Squatina sp. | 0.22 | 0.33 | 1.50 | Squatina guggenheim |
| Squalus acanthias | NaN | 0 | 7.70 | Squalus cubensis |
| Squalus cubensis | NaN | 0 | 7.70 | Squalus acanthias |
| Consensus MOTUs | ||||
| Squatina aculeata | 0.06 | 0.16 | 6.22 | Squatina legnota |
| Squatina albipunctata | 0 | 0 | 0.99 | Squatina pseudocellata |
| Squatina armata | 0 | 0 | 2.85 | Squatina guggeheim |
| Squatina australis | 0.03 | 0.17 | 5.33 | Squatina albipunctata |
| Squatina david | 0.11 | 0.16 | 1.33 | Squatina guggeheim |
| Squatina dumeril | 0.43 | 0.99 | 2.52 | Squatina david |
| Squatina guggeheim | 0.31 | 0.66 | 0.50 | Squatina occulta |
| Squatina japonica | NaN | 0 | 6.99 | Squatina formosa/nebulosa |
| Squatina legnota | 0 | 0 | 2.51 | Squatina formosa/nebulosa |
| Squatina occulta | 0 | 0 | 0.50 | Squatina guggeheim |
| Squatina oculata | 0 | 0 | 5.87 | Squatina legnota |
| Squatina pseudocellata | 0 | 0 | 0.99 | Squatina albipunctata |
| Squatina tergocellata | 0 | 0 | 0.99 | Squatina pseudocellata |
| Squatina tergocellatoides | 0.25 | 0.50 | 4.44 | Squatina legnota |
| Squatina sp. | 0.22 | 0.33 | 1.50 | Squatina guggeheim |
| Squatina squatina | 0.52 | 0.99 | 6.61 | Squatina aculeata |
| Squatina californica USA/Mex | 0.41 | 0.85 | 0.49 | Squatina californica Per |
| Squatina californica Per | NaN | 0 | 0.50 | Squatina californica USA/Mex |
| Squatina africana 1 | NaN | 0 | 0.83 | Squatina africana 2 |
| Squatina africana 2 | NaN | 0 | 0.83 | Squatina africana 1 |
| Squatina africana 3 | 0.11 | 0.17 | 0.83 | Squatina africana 2 |
| Squatina africana 4 | 0 | 0 | 2.00 | Squatina africana 2 |
| Squatina formosa/nebulosa | 0.15 | 0.33 | 2.51 | Squatina legnota |
| Squalus acanthias | NaN | 0 | 7.70 | Squalus cubensis |
| Squalus cubensis | NaN | 0 | 7.70 | Squalus acanthias |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cañedo-Apolaya, R.M.; Ortiz-Alvarez, C.; Alfaro-Cordova, E.; Alfaro-Shigueto, J.; Velez-Zuazo, X.; Mangel, J.C.; Siccha-Ramirez, R.; Yamashiro, C.; Ramirez, J.L. Species Delimitation of Southeast Pacific Angel Sharks (Squatina spp.) Reveals Hidden Diversity through DNA Barcoding. Diversity 2021, 13, 177. https://doi.org/10.3390/d13050177
Cañedo-Apolaya RM, Ortiz-Alvarez C, Alfaro-Cordova E, Alfaro-Shigueto J, Velez-Zuazo X, Mangel JC, Siccha-Ramirez R, Yamashiro C, Ramirez JL. Species Delimitation of Southeast Pacific Angel Sharks (Squatina spp.) Reveals Hidden Diversity through DNA Barcoding. Diversity. 2021; 13(5):177. https://doi.org/10.3390/d13050177
Chicago/Turabian StyleCañedo-Apolaya, Rosa M., Clara Ortiz-Alvarez, Eliana Alfaro-Cordova, Joanna Alfaro-Shigueto, Ximena Velez-Zuazo, Jeffrey C. Mangel, Raquel Siccha-Ramirez, Carmen Yamashiro, and Jorge L. Ramirez. 2021. "Species Delimitation of Southeast Pacific Angel Sharks (Squatina spp.) Reveals Hidden Diversity through DNA Barcoding" Diversity 13, no. 5: 177. https://doi.org/10.3390/d13050177