By-Products of Agri-Food Industry as Tannin-Rich Sources: A Review of Tannins’ Biological Activities and Their Potential for Valorization

 , ,
, ,  , , , ,
, , , ,  and
and 
Abstract
:
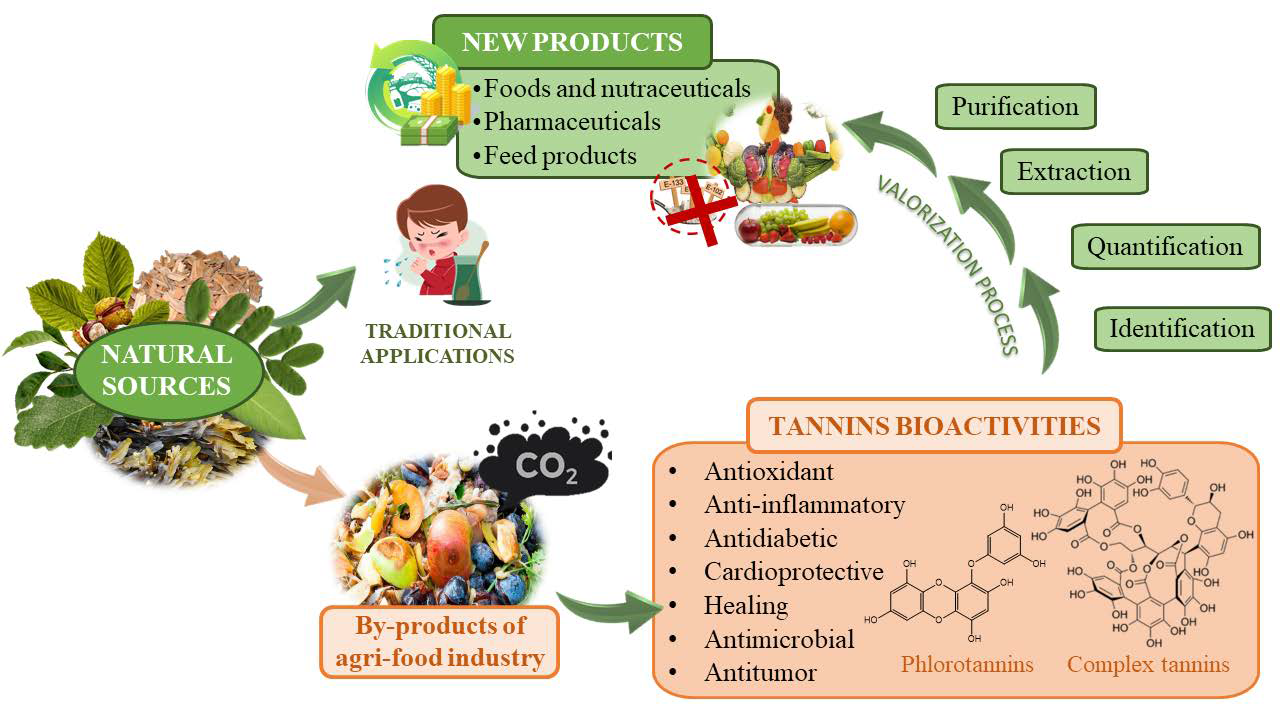
1. Introduction
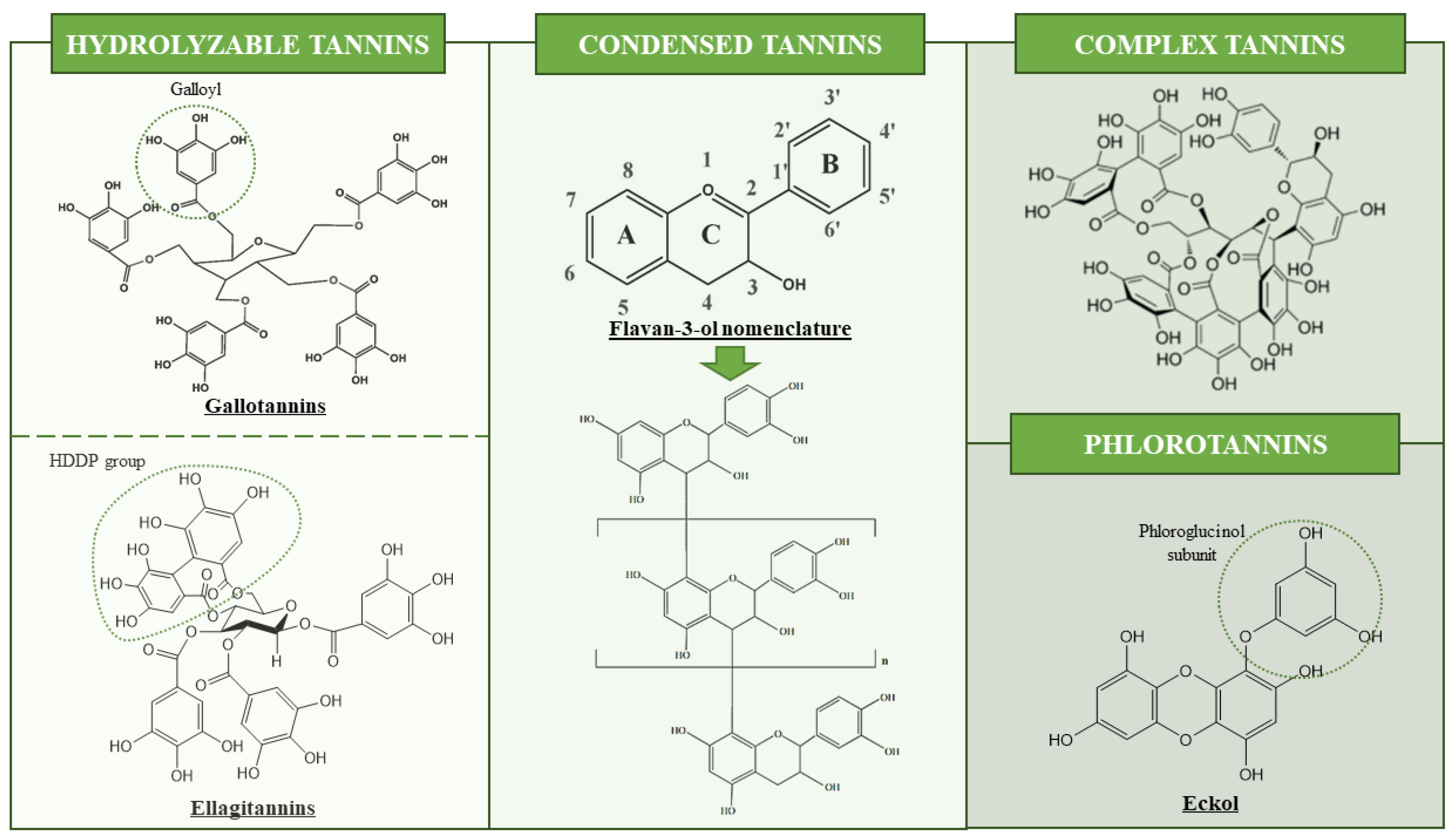
1.1. Tannins as Target Compounds
1.2. Circular Economy and Exploitation of By-Products
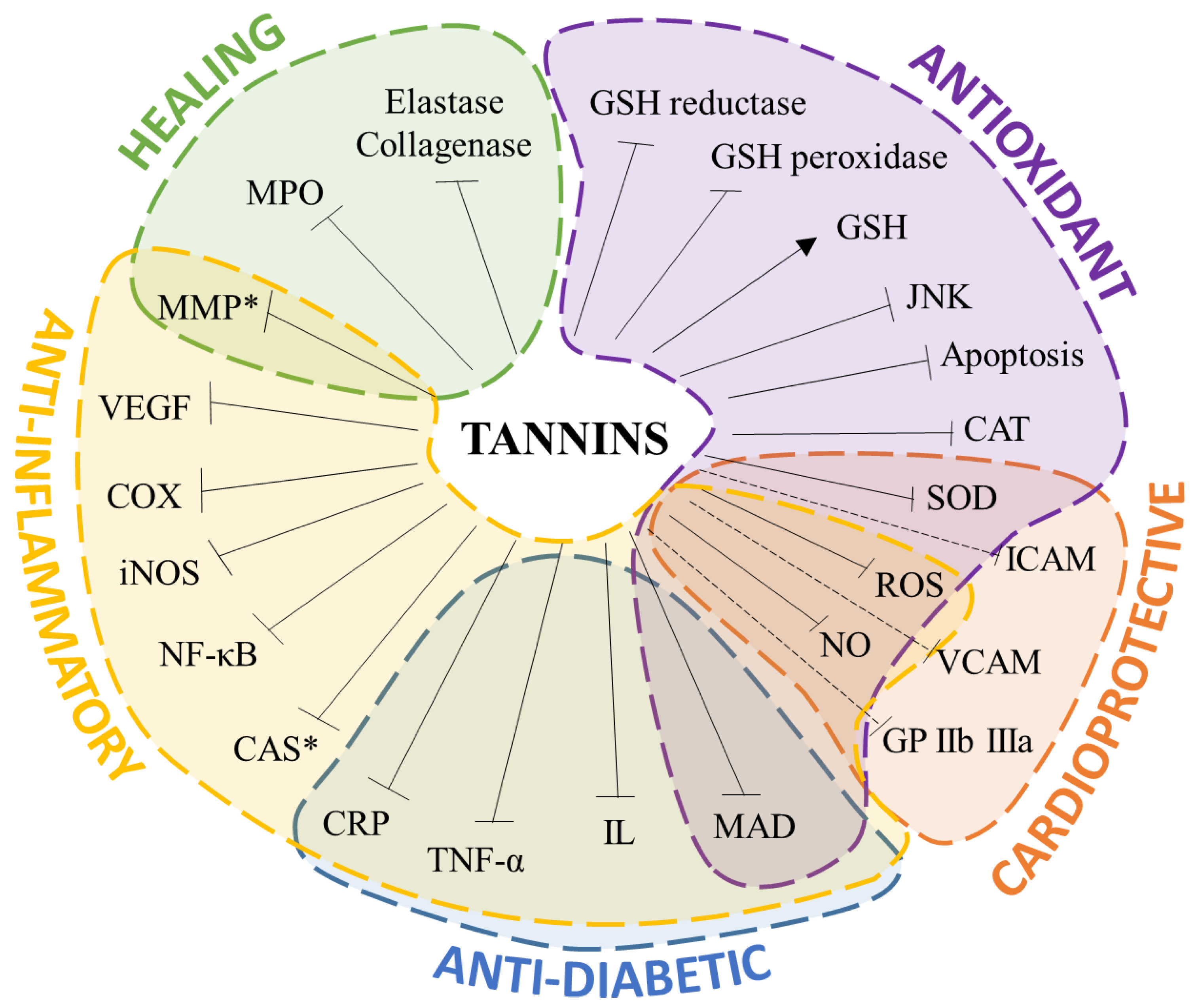
2. Biological Activities of Tannin-Rich Extracts
2.1. Antioxidant
2.2. Anti-Inflammatory
2.3. Antidiabetic
2.4. Cardioprotection and Blood Circulation Improvement
2.5. Wound Healing
2.6. Antimicrobial
2.7. Other Beneficial Applications of Tannins
2.7.1. Human Beings
2.7.2. Veterinarians
2.7.3. Botanical
2.7.4. Food Additives
3. Valorization Approach and Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Tannins | |
| CoT | Complex tannin |
| CT | Condensed tannin |
| EA | Ellagic acid |
| ET | Ellagitannin |
| GA | Gallic acid |
| GT | Gallotannin |
| HHDP | Hexahydroxydiphenol |
| HT | Hydrolysable tannin |
| PC | Procyanidin |
| PD | Prodelphinidin |
| PG | Phloroglucinol |
| PGG | Pentagalloylglucose |
| PoGG | Polygalloylglucose |
| PT | Phlorotannin |
| TGG | Trigalloylglucose |
| Bioactivities and Assays | |
| ADP | Adenosine diphosphate |
| Bcl-2 | Apoptosis inhibitor gen |
| CAS | Caspase |
| CAT | Catalase |
| COX | Cyclooxygenase |
| CRP | C-reactive protein |
| DM | Diabetes mellitus |
| DNA | Deoxyribonucleic acid |
| GABA | Gamma-aminobutyric acid |
| GSH | Glutathione (reduced) |
| HBeAG | E antigen of the hepatitis B virus |
| HBV | Hepatitis B virus |
| HIV | Human immunodeficiency virus |
| ICAM | Intercellular adhesion molecule |
| IL | Interleukin |
| iNOS | Nitric oxide synthase |
| IZ | Inhibition zone |
| JNK | C-Jun N-terminal kinase |
| LPS | Lipopolysaccharide |
| MAD | Malondialdehyde |
| MBC | Minimum bactericidal concentration |
| MIC | Minimal inhibitory concentration |
| MMP | Matrix metalloproteinase |
| mRNA | Messenger ribonucleic acid |
| MRSA | Methicillin-resistant Staphylococcus aureus |
| NADPH | Nicotinamide adenine dinucleotide (reduced) |
| NF-κB | Nuclear factor-κB |
| NO | Nitric oxide |
| NSAID | Nonsteroidal anti-inflammatory drug |
| RNA | Ribonucleic acid |
| ROS | Reactive oxygen species |
| SOD | Superoxide dismutase |
| TNF-α | Tumor necrosis factor-α |
| VCAM | Vascular cell adhesion protein |
| VEGF | Vascular endothelial growth factor |
References
- Barbehenn, R.V.; Peter Constabel, C. Tannins in plant-herbivore interactions. Phytochemistry 2011, 72, 1551–1565. [Google Scholar] [CrossRef] [PubMed]
- De Hoyos-Martínez, P.L.; Merle, J.; Labidi, J. Charrier–El Bouhtoury, F. Tannins extraction: A key point for their valorization and cleaner production. J. Clean. Prod. 2019, 206, 1138–1155. [Google Scholar] [CrossRef] [Green Version]
- Shirmohammadli, Y.; Efhamisisi, D.; Pizzi, A. Tannins as a sustainable raw material for green chemistry: A review. Ind. Crop. Prod. 2018, 126, 316–332. [Google Scholar] [CrossRef]
- Vuolo, M.M.; Lima, V.S.; Maróstica Junior, M.R. Phenolic compounds: Structure, classification, and antioxidant power. In Bioactive Compounds: Health Benefits and Potential Applications; Elsevier Inc.: Amsterdam, The Netherlands, 2018; pp. 33–50. ISBN 9780128147757. [Google Scholar]
- Ky, I.; Le Floch, A.; Zeng, L.; Pechamat, L.; Jourdes, M.; Teissedre, P.L. Tannins. Encycl. Food Health 2015, 7, 247–255. [Google Scholar]
- Okuda, T.; Ito, H. Tannins of constant structure in medicinal and food plants-hydrolyzable tannins and polyphenols related to tannins. Molecules 2011, 16, 2191–2217. [Google Scholar] [CrossRef]
- Macáková, K.; Kolečkář, V.; Cahlíková, L.; Chlebek, J.; Hoštálková, A.; Kuča, K.; Jun, D.; Opletal, L. Tannins and their influence on health. In Recent Advances in Medicinal Chemistry; Rahman, A., Choudhary, M., Perry, G., Eds.; Bentham Science Publishers: Sarja, UAE, 2014; Volume 1, pp. 159–208. ISBN 9780128039618. [Google Scholar]
- Molino, S.; Casanova, N.A.; Henares, J.Á.R.; Miyakawa, M.E.F. Natural tannin wood extracts as a potential food ingredient in the food industry. J. Agric. Food Chem. 2020, 68, 2836–2848. [Google Scholar] [CrossRef]
- Shahidi, F.; Ambigaipalan, P. Phenolics and polyphenolics in foods, beverages and spices: Antioxidant activity and health effects—A review. J. Funct. Foods 2015, 18, 820–897. [Google Scholar] [CrossRef]
- Fraga-Corral, M.; García-Oliveira, P.; Pereira, A.G.; Lourenço-Lopes, C.; Jimenez-Lopez, C.; Prieto, M.A.; Simal-Gandara, J. Technological application of tannin-based extracts. Molecules 2020, 25, 614. [Google Scholar] [CrossRef] [Green Version]
- Jiménez, N.; Esteban-Torres, M.; Mancheño, J.M.; de las Rivas, B.; Muñoza, R. Tannin degradation by a novel tannase enzyme present in some Lactobacillus plantarum strains. Appl. Environ. Microbiol. 2014, 80, 2991–2997. [Google Scholar] [CrossRef] [Green Version]
- Arapitsas, P. Hydrolyzable tannin analysis in food. Food Chem. 2012, 135, 1708–1717. [Google Scholar] [CrossRef]
- Hagerman, A.E. Hydrolyzable tannin structural chemistry. Tann. Handb. 2010, 1–8. Available online: http//www.users.muohio.edu/hagermae/tannin.pdf (accessed on 11 October 2020).
- Rousserie, P.; Rabot, A.; Geny-Denis, L. From flavanols biosynthesis to wine tannins: What place for grape seeds? J. Agric. Food Chem. 2019, 67, 1325–1343. [Google Scholar] [CrossRef] [PubMed]
- Sieniawska, E.; Baj, T. Tannins. In Pharmacognosy; Elsevier: Amsterdam, The Netherlands, 2017; pp. 199–232. [Google Scholar]
- Venkatesan, J.; Keekan, K.K.; Anil, S.; Bhatnagar, I.; Kim, S.-K. Phlorotannins. Encycl. Food Chem. 2019, 515–527. [Google Scholar] [CrossRef]
- Erpel, F.; Mateos, R.; Pérez-Jiménez, J.; Pérez-Correa, J.R. Phlorotannins: From isolation and structural characterization, to the evaluation of their antidiabetic and anticancer potential. Food Res. Int. 2020, 137, 109589. [Google Scholar] [CrossRef]
- Salminen, J.P.; Karonen, M. Chemical ecology of tannins and other phenolics: We need a change in approach. Funct. Ecol. 2011, 25, 325–338. [Google Scholar] [CrossRef]
- Huang, Q.; Liu, X.; Zhao, G.; Hu, T.; Wang, Y. Potential and challenges of tannins as an alternative to in-feed antibiotics for farm animal production. Anim. Nutr. 2018, 4, 137–150. [Google Scholar] [CrossRef]
- Sharma, K.; Kumar, V.; Kaur, J.; Tanwar, B.; Goyal, A.; Sharma, R.; Gat, Y.; Kumar, A. Health effects, sources, utilization and safety of tannins: A critical review. Toxin Rev. 2019, 1–13. [Google Scholar] [CrossRef]
- Sharma, K.; Guleria, S.; Razdan, V.K.; Babu, V. Synergistic antioxidant and antimicrobial activities of essential oils of some selected medicinal plants in combination and with synthetic compounds. Ind. Crop. Prod. 2020, 154, 112569. [Google Scholar] [CrossRef]
- Rana, J.; Paul, J. Consumer behavior and purchase intention for organic food: A review and research agenda. J. Retail. Consum. Serv. 2017, 38, 157–165. [Google Scholar] [CrossRef]
- Winans, K.; Kendall, A.; Deng, H. The history and current applications of the circular economy concept. Renew. Sustain. Energy Rev. 2017, 68, 825–833. [Google Scholar] [CrossRef]
- Jimenez-Lopez, C.; Fraga-Corral, M.; Carpena, M.; García-Oliveira, P.; Echave, J.; Pereira, A.G.; Lourenço-Lopes, C.; Prieto, M.A.; Simal-Gandara, J. Agriculture waste valorisation as a source of antioxidant phenolic compounds within a circular and sustainable bioeconomy. Food Funct. 2020, 11, 4853–4877. [Google Scholar] [CrossRef] [PubMed]
- Aires, A.; Carvalho, R.; Saavedra, M.J. Valorization of solid wastes from chestnut industry processing: Extraction and optimization of polyphenols, tannins and ellagitannins and its potential for adhesives, cosmetic and pharmaceutical industry. Waste Manag. 2016, 48, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Grenda, K.; Arnold, J.; Hunkeler, D.; Gamelas, J.A.F.; Rasteiro, M.G. Tannin-based coagulants from laboratory to pilot plant scales for coloured wastewater treatment. BioResources 2018, 13, 2727–2747. [Google Scholar] [CrossRef] [Green Version]
- Cassani, L.; Gomez-zavaglia, A.; Jimenez-lopez, C.; Prieto, M.A.; Simal-gandara, J. Seaweed-based natural ingredients: Stability of phlorotannins during extraction, storage, passage through the gastrointestinal tract and potential incorporation into functional foods. Food Res. Int. 2020, 109676. [Google Scholar] [CrossRef] [PubMed]
- Das, A.K.; Islam, M.N.; Faruk, M.O.; Ashaduzzaman, M.; Dungani, R. Review on tannins: Extraction processes, applications and possibilities. S. Afr. J. Bot. 2020, 135, 58–70. [Google Scholar] [CrossRef]
- Mirabella, N.; Castellani, V.; Sala, S. Current options for the valorization of food manufacturing waste: A review. J. Clean. Prod. 2014, 65, 28–41. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, A.; Baenas, N.; Dominguez-Perles, R.; Barros, A.; Rosa, E.; Moreno, D.A.; Garcia-Viguera, C. Natural bioactive compounds from winery by-products as health promoters: A review. Int. J. Mol. Sci. 2014, 15, 15638–15678. [Google Scholar] [CrossRef] [Green Version]
- Smeriglio, A.; Barreca, D.; Bellocco, E.; Trombetta, D. Proanthocyanidins and hydrolysable tannins: Occurrence, dietary intake and pharmacological effects. Br. J. Pharmacol. 2017, 174, 1244–1262. [Google Scholar] [CrossRef] [Green Version]
- Serrano, J.; Puupponen-Pimiä, R.; Dauer, A.; Aura, A.-M.; Saura-Calixto, F. Tannins: Current knowledge of food sources, intake, bioavailability and biological effects. Mol. Nutr. Food Res. 2009, 53, S310–S329. [Google Scholar] [CrossRef] [Green Version]
- Xiong, J.; Grace, M.H.; Esposito, D.; Wang, F.; Lila, M.A. Phytochemical characterization and anti-inflammatory properties of Acacia mearnsii leaves. Nat. Prod. Commun. 2016, 11. [Google Scholar] [CrossRef] [Green Version]
- dos Santos, C.; Vargas, Á.; Fronza, N.; dos Santos, J.H.Z. Structural, textural and morphological characteristics of tannins from Acacia mearnsii encapsulated using sol-gel methods: Applications as antimicrobial agents. Colloids Surf. B Biointerfaces 2017, 151, 26–33. [Google Scholar] [CrossRef]
- Rather, L.J.; Shahid-ul-Islam; Mohammad, F. Acacia nilotica (L.): A review of its traditional uses, phytochemistry, and pharmacology. Sustain. Chem. Pharm. 2015, 2, 12–30. [Google Scholar] [CrossRef]
- Safari, V.Z.; Kamau, J.K.; Nthiga, P.M.; Ngugi, M.P.; Orinda, G.; Njagi, E.M. Antipyretic, antiinflammatory and antinociceptive activities of aqueous bark extract of Acacia nilotica (L.) Delile in albino mice. Pain Manag. Med. 2016, 2, 2. [Google Scholar]
- Brizi, C.; Santulli, C.; Micucci, M.; Budriesi, R.; Chiarini, A.; Aldinucci, C.; Frosini, M. Neuroprotective Effects of Castanea sativa Mill. bark extract in human neuroblastoma cells subjected to oxidative stress. J. Cell. Biochem. 2016, 117, 510–520. [Google Scholar] [CrossRef] [PubMed]
- Chiocchio, I.; Prata, C.; Mandrone, M.; Ricciardiello, F.; Marrazzo, P.; Tomasi, P.; Angeloni, C.; Fiorentini, D.; Malaguti, M.; Poli, F.; et al. Leaves and spiny burs of Castanea Sativa from an experimental chestnut grove: Metabolomic analysis and anti-neuroinflammatory activity. Metabolites 2020, 10, 408. [Google Scholar] [CrossRef]
- Cardullo, N.; Muccilli, V.; Saletti, R.; Giovando, S.; Tringali, C. A mass spectrometry and 1H NMR study of hypoglycemic and antioxidant principles from a Castanea sativa tannin employed in oenology. Food Chem. 2018, 268, 585–593. [Google Scholar] [CrossRef] [PubMed]
- Chiarini, A.; Micucci, M.; Malaguti, M.; Budriesi, R.; Ioan, P.; Lenzi, M.; Fimognari, C.; Gallina Toschi, T.; Comandini, P.; Hrelia, S. Sweet chestnut (Castanea sativa Mill.) bark extract: Cardiovascular activity and myocyte protection against oxidative damage. Oxidative Med. Cell. Longev. 2013, 2013, 471790. [Google Scholar] [CrossRef] [Green Version]
- Silva, V.; Falco, V.; Dias, M.I.; Barros, L.; Silva, A.; Capita, R.; Alonso-Calleja, C.; Amaral, J.S.; Igrejas, G.; Ferreira, I.C.F.R.; et al. Evaluation of the phenolic profile of Castanea sativa Mill. by-products and their antioxidant and antimicrobial activity against multiresistant bacteria. Antioxidants 2020, 9, 87. [Google Scholar] [CrossRef] [Green Version]
- Štumpf, S.; Hostnik, G.; Primožič, M.; Leitgeb, M.; Salminen, J.-P.; Bren, U. The effect of growth medium strength on minimum inhibitory concentrations of tannins and tannin extracts against E. coli. Molecules 2020, 25, 2947. [Google Scholar] [CrossRef]
- Reggi, S.; Giromini, C.; Dell’Anno, M.; Baldi, A.; Rebucci, R.; Rossi, L. In vitro digestion of chestnut and quebracho tannin extracts: Antimicrobial effect, antioxidant capacity and cytomodulatory activity in swine intestinal IPEC-J2 cells. Animals 2020, 10, 195. [Google Scholar] [CrossRef] [Green Version]
- Esposito, T.; Celano, R.; Pane, C.; Piccinelli, A.L.; Sansone, F.; Picerno, P.; Zaccardelli, M.; Aquino, R.P.; Mencherini, T. Chestnut (Castanea sativa miller.) burs extracts and functional compounds: Uhplc-uv-hrms profiling, antioxidant activity, and inhibitory effects on phytopathogenic fungi. Molecules 2019, 24, 302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonelli, F.; Turini, L.; Sarri, G.; Serra, A.; Buccioni, A.; Mele, M. Oral administration of chestnut tannins to reduce the duration of neonatal calf diarrhea. BMC Vet. Res. 2018, 14, 4–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regueiro, J.; Sánchez-González, C.; Vallverdú-Queralt, A.; Simal-Gándara, J.; Lamuela-Raventós, R.; Izquierdo-Pulido, M. Comprehensive identification of walnut polyphenols by liquid chromatography coupled to linear ion trap-Orbitrap mass spectrometry. Food Chem. 2014, 152, 340–348. [Google Scholar] [CrossRef] [PubMed]
- Papoutsi, Z.; Kassi, E.; Chinou, I.; Halabalaki, M.; Skaltsounis, L.A.; Moutsatsou, P. Walnut extract (Juglans regia L.) and its component ellagic acid exhibit anti-inflammatory activity in human aorta endothelial cells and osteoblastic activity in the cell line KS483. Br. J. Nutr. 2008, 99, 715–722. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Qi, G.; Li, D.; Meng, H.; Zhu, Z.; Zhao, Y.; Qi, Y.; Zhang, X. Walnut (Juglans regia L.) kernel extracts protect against isoproterenol-induced myocardial infarction in rats. Rejuvenation Res. 2018, 22, 306–312. [Google Scholar] [CrossRef]
- Meagher, L.P.; Lane, G.; Sivakumaran, S.; Tavendale, M.H.; Fraser, K. Characterization of condensed tannins from Lotus species by thiolytic degradation and electrospray mass spectrometry. Anim. Feed Sci. Technol. 2004, 117, 151–163. [Google Scholar] [CrossRef]
- Hedqvist, H.; Mueller-Harvey, I.; Reed, J.D.; Krueger, C.G.; Murphy, M. Characterisation of tannins and in vitro protein digestibility of several Lotus corniculatus varieties. Anim. Feed Sci. Technol. 2000, 87, 41–56. [Google Scholar] [CrossRef]
- Min, B.R.; Fernandez, J.M.; Barry, T.N.; McNabb, W.C.; Kemp, P.D. The effect of condensed tannins in Lotus corniculatus upon reproductive efficiency and wool production in ewes during autumn. Anim. Feed Sci. Technol. 2001, 92, 185–202. [Google Scholar] [CrossRef]
- Raitanen, J.-E.; Järvenpää, E.; Korpinen, R.; Mäkinen, S.; Hellström, J.; Kilpeläinen, P.; Liimatainen, J.; Ora, A.; Tupasela, T.; Jyske, T. Tannins of conifer bark as nordic piquancy—Sustainable preservative and aroma? Molecules 2020, 25, 567. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Cai, D.; Zhang, L.; Tang, W.; Yan, R.; Guo, H.; Chen, X. Identification of hydrolyzable tannins (punicalagin, punicalin and geraniin) as novel inhibitors of hepatitis B virus covalently closed circular DNA. Antiviral Res. 2016, 134, 97–107. [Google Scholar] [CrossRef] [Green Version]
- Arunkumar, J.; Rajarajan, S. Study on antiviral activities, drug-likeness and molecular docking of bioactive compounds of Punica granatum L. to Herpes simplex virus-2 (HSV-2). Microb. Pathog. 2018, 118, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Muccilli, V.; Cardullo, N.; Spatafora, C.; Cunsolo, V.; Tringali, C. α-Glucosidase inhibition and antioxidant activity of an oenological commercial tannin. Extraction, fractionation and analysis by HPLC/ESI-MS/MS and 1H NMR. Food Chem. 2017, 215, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Horvathova, M.; Orszaghova, Z.; Laubertova, L.; Vavakova, M.; Sabaka, P.; Rohdewald, P.; Durackova, Z.; Muchova, J. Effect of the French oak wood extract robuvit on markers of oxidative stress and activity of antioxidant enzymes in healthy volunteers: A pilot study. Oxidative Med. Cell. Longev. 2014, 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Natella, F.; Leoni, G.; Maldini, M.; Natarelli, L.; Comitato, R.; Schonlau, F.; Virgili, F.; Canali, R. Absorption, metabolism, and effects at transcriptome level of a standardized french oak wood extract, Robuvit, in healthy volunteers: Pilot study. J. Agric. Food Chem. 2014, 62, 443–453. [Google Scholar] [CrossRef] [PubMed]
- Khalilpour, S.; Sangiovanni, E.; Piazza, S.; Fumagalli, M.; Beretta, G.; Dell’Agli, M. In vitro evidences of the traditional use of Rhus coriaria L. fruits against skin inflammatory conditions. J. Ethnopharmacol. 2019, 238, 111829. [Google Scholar] [CrossRef]
- Isik, S.; Tayman, C.; Cakir, U.; Koyuncu, I.; Taskin Turkmenoglu, T.; Cakir, E. Sumac (Rhus coriaria) for the prevention and treatment of necrotizing enterocolitis. J. Food Biochem. 2019, 43, e13068. [Google Scholar] [CrossRef]
- Monforte, M.T.; Smeriglio, A.; Germanò, M.P.; Pergolizzi, S.; Circosta, C.; Galati, E.M. Evaluation of antioxidant, antiinflammatory, and gastroprotective properties of Rubus fruticosus L. fruit juice. Phytother. Res. 2018, 32, 1404–1414. [Google Scholar] [CrossRef]
- Mirazi, N.; Hosseini, A. Attenuating properties of Rubus fruticosus L. on oxidative damage and inflammatory response following streptozotocin-induced diabetes in the male Wistar rats. J. Diabetes Metab. Disord. 2020. [Google Scholar] [CrossRef]
- Li, Y.; Fu, X.; Duan, D.; Liu, X.; Xu, J.; Gao, X. Extraction and identification of phlorotannins from the brown alga, Sargassum fusiforme (Harvey) Setchell. Mar. Drugs 2017, 15, 49. [Google Scholar] [CrossRef] [Green Version]
- Casas, M.P.; Rodríguez-Hermida, V.; Pérez-Larrán, P.; Conde, E.; Liveri, M.T.; Ribeiro, D.; Fernandes, E.; Domínguez, H. In vitro bioactive properties of phlorotannins recovered from hydrothermal treatment of Sargassum muticum. Sep. Purif. Technol. 2016, 167, 117–126. [Google Scholar] [CrossRef]
- Venter, P.B.; Sisa, M.; Van Der Merwe, M.J.; Bonnet, S.L.; van der Westhuizen, J.H. Analysis of commercial proanthocyanidins. Part 1: The chemical composition of quebracho (Schinopsis lorentzii and Schinopsis balansae) heartwood extract. Phytochemistry 2012, 73, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Cardullo, N.; Muccilli, V.; Cunsolo, V.; Tringali, C. Mass spectrometry and 1H-NMR study of Schinopsis lorentzii (Quebracho) tannins as a source of hypoglycemic and antioxidant principles. Molecules 2020, 25, 3257. [Google Scholar] [CrossRef] [PubMed]
- Fruet, A.P.B.; Giotto, F.M.; Fonseca, M.A.; Nörnberg, J.L.; de Mello, A.S. Effects of the incorporation of tannin extract from quebracho colorado wood on color parameters, lipid oxidation, and sensory attributes of beef patties. Foods 2020, 9, 667. [Google Scholar] [CrossRef] [PubMed]
- Falcão, L.; Araújo, M.E.M. Vegetable tannins used in the manufacture of historic leathers. Molecules 2018, 23, 1081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, M.H.; Ali, Z.; Khan, I.A.; Khan, S.I. Anti-inflammatory activity of constituents isolated from Terminalia chebula. Nat. Prod. Commun. 2014, 9. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Pérez, C.; García-Villanova, B.; Guerra-Hernández, E.; Verardo, V. Grape seeds proanthocyanidins: An overview of in vivo bioactivity in animal models. Nutrients 2019, 11, 2435. [Google Scholar] [CrossRef] [Green Version]
- Sieniawska, E. Activities of Tannins—From in vitro studies to clinical trials. Nat. Prod. Commun. 2015, 10. [Google Scholar] [CrossRef] [Green Version]
- Daré, R.G.; Nakamura, C.V.; Ximenes, V.F.; Lautenschlager, S.O.S. Tannic acid, a promising anti-photoaging agent: Evidences of its antioxidant and anti-wrinkle potentials, and its ability to prevent photodamage and MMP-1 expression in L929 fibroblasts exposed to UVB. Free Radic. Biol. Med. 2020, 160, 342–355. [Google Scholar] [CrossRef]
- Okuda, T.; Kimura, Y.; Yoshida, T.; Hatano, T.; Okuda, H.; Arichi, S. Studies on the activities of tannins and related compounds from medicinal plants and drugs. I. Inhibitory effects on lipid peroxidation in mitochondria and microsomes of liver. Chem. Pharm. Bull. 1983, 31, 1625–1631. [Google Scholar] [CrossRef] [Green Version]
- Iwata, S.; Fukaya, Y.; Nakazawa, K.; Okuda, T. Effects of tannins on the oxidative damage of mouse ocular lens I. using the oxidative damage model induced by the xanthine-xanthine oxidase system. J. Ocul. Pharmacol. Ther. 1987, 3, 227–238. [Google Scholar] [CrossRef]
- Beninger, C.W.; Hosfield, G.L. Antioxidant activity of extracts, condensed tannin fractions, and pure flavonoids from Phaseolus vulgaris L. seed coat color genotypes. J. Agric. Food Chem. 2003, 51, 7879–7883. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Zou, B.; Li, C.; Yang, J.; Xu, S.; Hagerman, A.E. High molecular weight persimmon tannin is a potent antioxidant both ex vivo and in vivo. Food Res. Int. 2012, 45, 26–30. [Google Scholar] [CrossRef]
- Huang, W.; Niu, H.; Xue, X.; Li, J.; Li, C. Robinetinidol-(4β→8)-epigallocatechin 3-O-gallate, a galloyl dimer prorobinetinidin from Acacia mearnsii De Wild, effectively protects human neuroblastoma SH-SY5Y cells against acrolein-induced oxidative damage. J. Alzheimer′s Dis. 2010, 21, 493–506. [Google Scholar] [CrossRef] [PubMed]
- Terra, X.; Montagut, G.; Bustos, M.; Llopiz, N.; Ardèvol, A.; Bladé, C.; Fernández-Larrea, J.; Pujadas, G.; Salvadó, J.; Arola, L.; et al. Grape-seed procyanidins prevent low-grade inflammation by modulating cytokine expression in rats fed a high-fat diet. J. Nutr. Biochem. 2009, 20, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Park, M.; Cho, H.; Jung, H.; Lee, H.; Hwang, K.T. Antioxidant and anti-inflammatory activities of tannin fraction of the extract from black raspberry seeds compared to grape seeds. J. Food Biochem. 2014. [Google Scholar] [CrossRef]
- Kowalska, K.; Olejnik, A.; Zielińska-Wasielica, J.; Olkowicz, M. Raspberry (Rubus idaeus L.) fruit extract decreases oxidation markers, improves lipid metabolism and reduces adipose tissue inflammation in hypertrophied 3T3-L1 adipocytes. J. Funct. Foods 2019, 62, 103568. [Google Scholar] [CrossRef]
- Li, D.Q.; Zhao, J.; Xie, J.; Li, S.P. A novel sample preparation and on-line HPLC-DAD-MS/MS-BCD analysis for rapid screening and characterization of specific enzyme inhibitors in herbal extracts: Case study of α-glucosidase. J. Pharm. Biomed. Anal. 2014, 88, 130–135. [Google Scholar] [CrossRef]
- Senthilkumar, G.P.; Subramanian, S.P. Biochemical studies on the effect of Terminalia chebula on the levels of glycoproteins in streptozotocin-induced experimental diabetes in rats. J. Appl. Biomed. 2008, 6, 105–115. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, K.; Yokoyama, S. ichiro Induction of uncoupling protein-1 and -3 in brown adipose tissue by kaki-tannin in type 2 diabetic NSY/Hos mice. Food Chem. Toxicol. 2012, 50, 184–190. [Google Scholar] [CrossRef]
- Dudzinska, D.; Bednarska, K.; Boncler, M.; Luzak, B.; Watala, C. The influence of Rubus idaeus and Rubus caesius leaf extracts on platelet aggregation in whole blood. Cross-talk of platelets and neutrophils. Platelets 2016, 27, 433–439. [Google Scholar] [CrossRef]
- Ram, H.; Jatwa, R.; Purohit, A. Antiatherosclerotic and cardioprotective potential of Acacia senegal seeds in diet-induced atherosclerosis in rabbits. Biochem. Res. Int. 2014, 2014, 436848. [Google Scholar] [CrossRef] [PubMed]
- Tonin, T.D.; Thiesen, L.C.; de Oliveira Nunes, M.L.; Broering, M.F.; Donato, M.P.; Goss, M.J.; Petreanu, M.; Niero, R.; Machado, I.D.; Santin, J.R. Rubus imperialis (Rosaceae) extract and pure compound niga-ichigoside F1: Wound healing and anti-inflammatory effects. Naunyn. Schmiedeberg′s. Arch. Pharmacol. 2016, 389, 1235–1244. [Google Scholar] [CrossRef] [PubMed]
- Gabr, S.A.; Alghadir, A.H. Evaluation of the biological effects of lyophilized hydrophilic extract of rhus coriaria on Myeloperoxidase (MPO) activity, wound healing, and microbial infections of skin wound tissues. Evid. Based Complement. Altern. Med. 2019, 2019, 5861537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quave, C.L.; Plano, L.R.W.; Pantuso, T.; Bennett, B.C. Effects of extracts from Italian medicinal plants on planktonic growth, biofilm formation and adherence of methicillin-resistant Staphylococcus aureus. J. Ethnopharmacol. 2008, 118, 418–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saraiva, A.M.; Saraiva, C.L.; Cordeiro, R.P.; Soares, R.R.; Xavier, H.S.; Caetano, N. Atividade antimicrobiana e sinérgica das frações das folhas de Schinopsis brasiliensis Engl. frente a clones multirresistentes de Staphylococcus aureus. Rev. Bras. Plantas Med. 2013, 15, 199–207. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Feng, M.; Yang, K.; Cao, Y.; Zhang, J.; Xu, J.; Hernández, S.H.; Wei, X.; Fan, M. Transcriptomic and metabolomic analyses reveal antibacterial mechanism of astringent persimmon tannin against Methicillin-resistant Staphylococcus aureus isolated from pork. Food Chem. 2020, 309, 125692. [Google Scholar] [CrossRef]
- Martini, S.; D’Addario, C.; Colacevich, A.; Focardi, S.; Borghini, F.; Santucci, A.; Figura, N.; Rossi, C. Antimicrobial activity against Helicobacter pylori strains and antioxidant properties of blackberry leaves (Rubus ulmifolius) and isolated compounds. Int. J. Antimicrob. Agents 2009, 34, 50–59. [Google Scholar] [CrossRef] [Green Version]
- Hajaji, S.; Jabri, M.-A.; Sifaoui, I.; López-Arencibia, A.; Reyes-Batlle, M.; B’chir, F.; Valladares, B.; Pinero, J.E.; Lorenzo-Morales, J.; Akkari, H. Amoebicidal, antimicrobial and in vitro ROS scavenging activities of Tunisian Rubus ulmifolius Schott, methanolic extract. Exp. Parasitol. 2017, 183, 224–230. [Google Scholar] [CrossRef]
- Sisti, M.; De Santi, M.; Fraternale, D.; Ninfali, P.; Scoccianti, V.; Brandi, G. Antifungal activity of Rubus ulmifolius Schott standardized in vitro culture. LWT Food Sci. Technol. 2008, 41, 946–950. [Google Scholar] [CrossRef]
- Elizondo, A.M.; Mercado, E.C.; Rabinovitz, B.C.; Fernandez-Miyakawa, M.E. Effect of tannins on the in vitro growth of Clostridium perfringens. Vet. Microbiol. 2010, 145, 308–314. [Google Scholar] [CrossRef]
- Khalifa, I.; Zhu, W.; Mohammed, H.H.H.; Dutta, K.; Li, C. Tannins inhibit SARS-CoV-2 through binding with catalytic dyad residues of 3CLpro: An in silico approach with 19 structural different hydrolysable tannins. J. Food Biochem. 2020, 44, e13432. [Google Scholar] [CrossRef] [PubMed]
- Subbotina, M.D.; Timchenko, V.N.; Vorobyov, M.M.; Konunova, Y.S.; Aleksandrovih, Y.S.; Shushunov, S. Effect of oral administration of tormentil root extract (Potentilla tormentilla) on rotavirus diarrhea in children: A randomized, double blind, controlled trial. Pediatr. Infect. Dis. J. 2003, 22, 706–711. [Google Scholar] [CrossRef] [PubMed]
- Russo, M.; Coppola, V.; Giannetti, E.; Buonavolontà, R.; Piscitelli, A.; Staiano, A. Oral administration of tannins and flavonoids in children with acute diarrhea: A pilot, randomized, control-case study. Ital. J. Pediatrics 2018, 44, 4–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molino, S.; Fernández-Miyakawa, M.; Giovando, S.; Rufián-Henares, J.Á. Study of antioxidant capacity and metabolization of quebracho and chestnut tannins through in vitro gastrointestinal digestion-fermentation. J. Funct. Foods 2018, 49, 188–195. [Google Scholar] [CrossRef]
- Noce, A.; Marrone, G.; Bernini, R.; Campo, M.; Di Lauro, M.; Pietroboni Zaitseva, A.; Romani, A.; di Daniele, N. Impact of tannins as food supplement in a ckd population with recurrent urinary tract infections: Preliminary data. Nephrol. Dial. Transplant. 2020, 35, 1264. [Google Scholar] [CrossRef]
- Chandrasekhar, Y.; Phani Kumar, G.; Navya, K.; Ramya, E.M.; Anilakumar, K.R. Tannins from Terminalia chebula fruits attenuates GABA antagonist-induced anxiety-like behaviour via modulation of neurotransmitters. J. Pharm. Pharmacol. 2018, 70, 1662–1674. [Google Scholar] [CrossRef]
- Fagundes, G.M.; Benetel, G.; Santos, K.C.; Welter, K.C.; Melo, F.A.; Muir, J.P.; Bueno, I.C.S. Tannin-rich plants as natural manipulators of rumen fermentation in the livestock industry. Molecules 2020, 25, 2943. [Google Scholar] [CrossRef]
- El-Saber Batiha, G.; Beshbishy, A.M.; Adeyemi, O.S.; Nadwa, E.H.; Rashwan, E.K.M.; Alkazmi, L.M.; Elkelish, A.A.; Igarashi, I. Phytochemical screening and antiprotozoal effects of the methanolic berberis vulgaris and acetonic rhus coriaria extracts. Molecules 2020, 25, 550. [Google Scholar] [CrossRef] [Green Version]
- Quattrucci, A.; Ovidi, E.; Tiezzi, A.; Vinciguerra, V.; Balestra, G.M. Biological control of tomato bacterial speck using Punica granatum fruit peel extract. Crop Prot. 2013, 46, 18–22. [Google Scholar] [CrossRef]
- Li, R.; Dai, T.; Zhou, W.; Fu, G.; Wan, Y.; McClements, D.J.; Li, J. Impact of pH, ferrous ions, and tannic acid on lipid oxidation in plant-based emulsions containing saponin-coated flaxseed oil droplets. Food Res. Int. 2020, 136, 109618. [Google Scholar] [CrossRef]
- Al-Hijazeen, M.; Lee, E.J.; Mendonca, A.; Ahn, D.U. Effects of tannic acid on lipid and protein oxidation, color, and volatiles of raw and cooked chicken breast meat during storage. Antioxidants 2016, 5, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engelberth, A.S. Evaluating economic potential of food waste valorization: Onward to a diverse feedstock biorefinery. Curr. Opin. Green Sustain. Chem. 2020, 26, 100385. [Google Scholar] [CrossRef]
- Makkar, H.P.S.; Singh, B.; Negi, S.S. Tannin levels and their degree of polymerisation and specific activity in some agro-industrial by-products. Biol. Wastes 1990, 31, 137–144. [Google Scholar] [CrossRef]
- Si Heung Sung Antibacterial and antioxidant activities of tannins extracted from agricultural by-products. J. Med. Plants Res. 2012, 6, 3072–3079. [CrossRef]
- Tavares, C.S.; Martins, A.; Miguel, M.G.; Carvalheiro, F.; Duarte, L.C.; Gameiro, J.A.; Figueiredo, A.C.; Roseiro, L.B. Bioproducts from forest biomass II. Bioactive compounds from the steam-distillation by-products of Cupressus lusitanica Mill. and Cistus ladanifer L. wastes. Ind. Crop. Prod. 2020, 158, 112991. [Google Scholar] [CrossRef]
- Giovando, S.; Koch, G.; Romagnoli, M.; Paul, D.; Vinciguerra, V.; Tamantini, S.; Marini, F.; Zikeli, F.; Mugnozza, G.S. Spectro-topochemical investigation of the location of polyphenolic extractives (tannins) in chestnut wood structure and ultrastructure. Ind. Crop. Prod. 2019, 141, 111767. [Google Scholar] [CrossRef]
- Bhoite, R.N.; Murthy, P.S. Biodegradation of coffee pulp tannin by Penicillium verrucosum for production of tannase, statistical optimization and its application. Food Bioprod. Process. 2015, 94, 727–735. [Google Scholar] [CrossRef]
- Arbenz, A.; Avérous, L. Chemical modification of tannins to elaborate aromatic biobased macromolecular architectures. Green Chem. 2015, 17, 2626–2646. [Google Scholar] [CrossRef] [Green Version]
- Liang, X.; Jiang, Y.; Guo, Z.; Fang, S. Separation, UPLC-QTOF-MS/MS analysis, and antioxidant activity of hydrolyzable tannins from water caltrop (Trapa quadrispinosa) pericarps. LWT 2020, 133, 110010. [Google Scholar] [CrossRef]
- Wong-Paz, J.E.; Guyot, S.; Aguilar-Zárate, P.; Muñiz-Márquez, D.B.; Contreras-Esquivel, J.C.; Aguilar, C.N. Structural characterization of native and oxidized procyanidins (condensed tannins) from coffee pulp (Coffea arabica) using phloroglucinolysis and thioglycolysis-HPLC-ESI-MS. Food Chem. 2021, 340, 127830. [Google Scholar] [CrossRef]
- Hayder, Z.; Elfalleh, W.; Othman, K.; Benabderrahim, M.A.; Hannachi, H. Modeling of polyphenols extraction from pomegranate by-product using rotatable central composite design of experiments. Acta Ecol. Sin. 2020. In Press. [Google Scholar] [CrossRef]
- Ping, L.; Pizzi, A.; Guo, Z.D.; Brosse, N. Condensed tannins from grape pomace: Characterization by FTIR and MALDI TOF and production of environment friendly wood adhesive. Ind. Crop. Prod. 2012, 40, 13–20. [Google Scholar] [CrossRef]
- Hashim, N.; Paramasivam, M.; Tan, J.S.; Kernain, D.; Hussin, M.H.; Brosse, N.; Gambier, F.; Raja, P.B. Green mode synthesis of silver nanoparticles using Vitis vinifera’s tannin and screening its antimicrobial activity / apoptotic potential versus cancer cells. Mater. Today Commun. 2020, 25, 101511. [Google Scholar] [CrossRef]
- Cerulli, A.; Napolitano, A.; Masullo, M.; Hošek, J.; Pizza, C.; Piacente, S. Chestnut shells (Italian cultivar “Marrone di Roccadaspide” PGI): Antioxidant activity and chemical investigation with in depth LC-HRMS/MSn rationalization of tannins. Food Res. Int. 2020, 129, 108787. [Google Scholar] [CrossRef]
- Saad, H.; Charrier-El Bouhtoury, F.; Pizzi, A.; Rode, K.; Charrier, B.; Ayed, N. Characterization of pomegranate peels tannin extractives. Ind. Crop. Prod. 2012, 40, 239–246. [Google Scholar] [CrossRef]
- Maran, J.P.; Manikandan, S.; Priya, B.; Gurumoorthi, P. Box-Behnken design based multi-response analysis and optimization of supercritical carbon dioxide extraction of bioactive flavonoid compounds from tea (Camellia sinensis L.) leaves. J. Food Sci. Technol. 2015, 52, 92–104. [Google Scholar] [CrossRef]
- Naima, R.; Oumam, M.; Hannache, H.; Sesbou, A.; Charrier, B.; Pizzi, A.; El Bouhtoury, F.C. Comparison of the impact of different extraction methods on polyphenols yields and tannins extracted from Moroccan Acacia mollissima barks. Ind. Crop. Prod. 2015, 70, 245–252. [Google Scholar] [CrossRef]
- Dahmoune, F.; Nayak, B.; Moussi, K.; Remini, H.; Madani, K. Optimization of microwave-assisted extraction of polyphenols from Myrtus communis L. leaves. Food Chem. 2015, 166, 585–595. [Google Scholar] [CrossRef]
- Politi, F.A.S.; de Mello, J.C.P.; Migliato, K.F.; Nepomuceno, A.L.A.; Moreira, R.R.D.; Pietro, R.C.L.R. Antimicrobial, cytotoxic and antioxidant activities and determination of the total tannin content of bark extracts Endopleura uchi. Int. J. Mol. Sci. 2011, 12, 2757–2768. [Google Scholar] [CrossRef] [Green Version]
- Kemppainen, K.; Siika-aho, M.; Pattathil, S.; Giovando, S.; Kruus, K. Spruce bark as an industrial source of condensed tannins and non-cellulosic sugars. Ind. Crop. Prod. 2014, 52, 158–168. [Google Scholar] [CrossRef]
- Talmaciu, A.I.; Ravber, M.; Volf, I.; Knez, Ž.; Popa, V.I. Isolation of bioactive compounds from spruce bark waste using sub- and supercritical fluids. J. Supercrit. Fluids 2016, 117, 243–251. [Google Scholar] [CrossRef]
- Liu, Z.; Chen, Z.; Han, F.; Kang, X.; Gu, H.; Yang, L. Microwave-assisted method for simultaneous hydrolysis and extraction in obtaining ellagic acid, gallic acid and essential oil from Eucalyptus globulus leaves using Brönsted acidic ionic liquid [HO3S(CH2)4mim]HSO4. Ind. Crop. Prod. 2016, 81, 152–161. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Tannins | Properties | Patent No. |
|---|---|---|
| Punicalin, punicalagin, pedunculagin, tellimagrandin, corilagin, granatine a and b, terminalin | Treatment or prevention of cognitive and neurodegenerative disorders, metabolic syndrome, type 2 diabetes, dyslipidemia or obesity. | US20190000867A1 |
| Punicalagins | Functional food and beverage with increased antioxidant capacity for preventing or treating hypercholesterolemia and/or hypertension | EP2033526A1 |
| Chestnut tannins | Antioxidant or anti-microbial additive, or agent for reducing nitrosamines or mycotoxins | EP2904910B1 |
| Ellagitannins | Treatment of bacterial infections | US20110105421A1 |
| GA, EA, isoquercitrin, tellimagrandin I and II, pedunculagin, TGGs, PGG and di-galloyl-hexahydroxydiphenoyl-D-glucose | Inhibition or prevention of obesity, lipid storage (reducing blood triglyceride levels), hyperlipemia, arteriosclerosis and thrombosis | US7687085B2 |
| Gallotannins and ellagitannins | Regulation of the synthesis and secretion of cytokines, including TNF-α and IL-1β | US20080070850A1 |
| Ellagitannins | Anti-inflammatory or anti-allergic agent by the inhibition of histamine release from mast cells. Regular oral administration of product can ameliorate or prevent rhinitis, atopic dermatitis or asthma | EP0727218A3 |
| 1,3,4-tri-galloylquinic acid, galloylshikimic acid derivatives strictinin, corilagin, castalagin, vescalagin, chebulinic acid, chebulagic acid, punicalin, punicalagin, punicacortein C, cannamtannin B2 | Inhibition of the propagation in human cells of a human retrovirus (HIV) | CA2001898A1 |
| Tellimagrandin | Inhibition of Gram-positive bacteria (Staphylococcus aureus) growth, anti-inflammation and leukemia treatment | US8975234B2 |
| Source | Species | Classification | Compounds | Bioactivities | Ref. |
|---|---|---|---|---|---|
| Acacia sp. | A. mearnsii | CT | Epi-FIS derivatives | Antioxidant, anti-inflammatory, antimicrobial | [33,34] |
| A. nilotica | CT | PoGG, EA, GA, diGA, epi/gallocatechin, dicatechin derivatives | Antinociceptive, anti-inflammatory and antipyretic | [35,36] | |
| Castanea sp. | C. sativa | HT | CAST, VES, EA, chestanin | Antioxidant, anti-inflammatory, antidiabetic, cardioprotective, antimicrobial, antifungal, antidiarrheal (vet.) | [37,38,39,40,41,42,43,44,45] |
| Juglans sp. | J. regia | HT | EA, pedunculagin, casuariin | Antiplatelet, cardioprotective, antiatherogenic and anti-inflammatory | [46,47,48] |
| Lotus sp. | L. corniculatus | CT | Heteropolymers PC: PD | Improvement of animal performance | [49,50,51] |
| L. pedunculatus | CT | ||||
| Picea sp. | P. abies | CT | - | Antioxidant (food preservative) | [52] |
| Punica sp. | P. granatum | HT | Punicalagin, punicalin, geraniin | Antiviral (herpes simplex-2, hepatitis B) | [53,54] |
| Quercus sp. | Q. robur | HT | Castalin, vescalin, CAST, VES, GA, EA, PoGG | Antioxidant, antidiabetic | [55,56,57] |
| Rhus sp. | R. coriaria | CT HT | GA, QUERG, CYANG derivatives | Antimicrobial, anti-inflammatory, immunomodulatory, antiapoptotic and healing | [58,59] |
| Rubus sp | R. fruticosus | CT | CYANG, GA, malvidin-3-galactoside, vanillic acid | Antioxidant, anti-inflammatory, antidiabetic and gastroprotective | [60,61] |
| Sargassum sp. | S. fusiforme | PT | Eckol, dieckol, fuhalols | Antioxidant | [62] |
| S. muticum | PT | PG, diphlorethol, bi- and tri-fuhalol A, B | Antioxidant, antibacterial, antiproliferative, anti-inflammatory | [63] | |
| Schinopsis sp. | S. lorentzii | CT HT | FIS-catechin polymers TGG, PGG, quinic acid-GA esters | Antioxidant, antimicrobial, anthelmintic | [64,65,66,67] |
| S. balansae | CT | ProFIS polymers | Antioxidant, antimicrobial, anthelmintic | [1,64,65,66,67,68,69,70,71,72,73,74,75,76,77,78,79,80,81,82,83,84,85,86,87,88,89,90,91,92,93,94,95,96,97,98,99,100,101,102,103,104,105,106,107,108] | |
| Terminalia sp. | T. chebula | HT | Chebulinic acid, TGG | Anti-inflammatory | [68] |
| Vitis sp. | V. vinifera | CT | Galloylated PC, PC, PD | Antioxidant, anti-inflammatory, antiobesity | [69] |
| Species | Tannin | By-Product | Extraction Techniques | Experimental Conditions | Activity | Ref. |
|---|---|---|---|---|---|---|
| Trapa quadrispinosa | HT | Pericarps | UAE | Et/W (60/40, v/v), 30 min, 40 °C, L/S ratio 40 mL/g | Antioxidant (DPPH) | [112] |
| Cupressus lusitanica and Cistus ladanife | TTC | Waste distilled after steam distillation | UAE | Et S/L ratio 1:20, 30 min, 30 °C, 70% A | Antioxidant (ABTS) | [108] |
| Coffee (Coffea arabica) | Procyanidins (CT) | Pulp | UAE | W/A extract, 20 min, RT | - | [113] |
| Pomegranate (P. granatum var. Gabsi) | TTC | Peels | UAE | 2.63 g/100g dw, 55.46% E, 30 min | Antioxidant (DPPH and ABTS) | [114] |
| Red grape variety (Vitis vinifera | CT | Pomace | HAE | NaOH, Na2CO3 or NaHCO3) and Na2SO3 (2.5% or 5% (w/w). S/L ratio 1:8, 120 min, 100 °C | Production of environmentally friendly wood adhesive | [115] |
| Silver NPs, antimicrobial and apoptotic potential | [116] | |||||
| Chestnut (Castanea sativa) | TTC | Shells | Maceration | Et (20 mL × 3 days × 3 times) or Et/W 7:3 v/v (20 mL × 3 days × 3 times) | Antioxidant (DPPH and TEAC) | [117] |
| Pomegranate (Punica granatum L.) | TTC | Peels | HAE | W, 2% SS and 0.5% SB, S/L ratio 1:5, 7 h, 80 ± 5 °C | - | [118] |
| Tea (Camellia sinensis L.) | TTC | Leaves | SFC-CO2 | Supercritical CO2 flow rate 8 g/min, 188 bar, 50 °C, co-solvent flow rate 2.94 g/min | Antioxidant (ABTS) | [119] |
| Acacia mollissima | HT and CT | Bark | HAE and MAE | HAE: M (2h, 20 °C and 60 °C). MAE (1 min, 300W or 5 min, 150W) | - | [120] |
| Myrtus communis L. | TTC | Leaves | MAE | Et 42% (60 s, 500 W, S/L ratio 32 mL/g) | Antioxidant (DPPH, TEAC and ORAC) | [121] |
| Endopleura uchi | TTC | Bark | Maceration | Et/W 50% | Antimicrobial, cytotoxic and antioxidant | [122] |
| Norway spruce (Picea abies) | CT | Bark | Hot water extraction | 10% solid content, 2% SS, 0.5% SC, (75 °C, 120 min) | - | [123] |
| Spruce (Picea abies) | TTC | Bark | SFC-CO2 | Solvent consumption 2.5 kg CO2/kg product and 24.94 kg Et 70/kg product, 100 bar, 40 °C | Antioxidant (DPPH) | [124] |
| Eucalyptus globulus | EA and GA | Leaves | BMSHE | 1.0 M [HO3S(CH2)4mim] HSO4, L/S ratio 30 mL/g. MAE: 20 min, 385 W) | - | [125] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fraga-Corral, M.; Otero, P.; Echave, J.; Garcia-Oliveira, P.; Carpena, M.; Jarboui, A.; Nuñez-Estevez, B.; Simal-Gandara, J.; Prieto, M.A. By-Products of Agri-Food Industry as Tannin-Rich Sources: A Review of Tannins’ Biological Activities and Their Potential for Valorization. Foods 2021, 10, 137. https://doi.org/10.3390/foods10010137
Fraga-Corral M, Otero P, Echave J, Garcia-Oliveira P, Carpena M, Jarboui A, Nuñez-Estevez B, Simal-Gandara J, Prieto MA. By-Products of Agri-Food Industry as Tannin-Rich Sources: A Review of Tannins’ Biological Activities and Their Potential for Valorization. Foods. 2021; 10(1):137. https://doi.org/10.3390/foods10010137
Chicago/Turabian StyleFraga-Corral, María, Paz Otero, Javier Echave, Paula Garcia-Oliveira, Maria Carpena, Amira Jarboui, Bernabé Nuñez-Estevez, Jesus Simal-Gandara, and Miguel A. Prieto. 2021. "By-Products of Agri-Food Industry as Tannin-Rich Sources: A Review of Tannins’ Biological Activities and Their Potential for Valorization" Foods 10, no. 1: 137. https://doi.org/10.3390/foods10010137