Spoilage Potential of Pseudomonas (P. fragi, P. putida) and LAB (Leuconostoc mesenteroides, Lactobacillus sakei) Strains and Their Volatilome Profile during Storage of Sterile Pork Meat Using GC/MS and Data Analytics

 ,
,  ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design and Preparation of Sterile Meat Samples
2.2. Preparation of Inoculum and Inoculation of the Sterile Pork Meat
2.3. Microbiological Analyses
2.4. Sensory Analysis
2.5. Determination of Volatile Compounds by Headspace SPME-GC/MS
2.6. Estimation of Growth Kinetic Parameters and Data Analysis
3. Results and Discussion
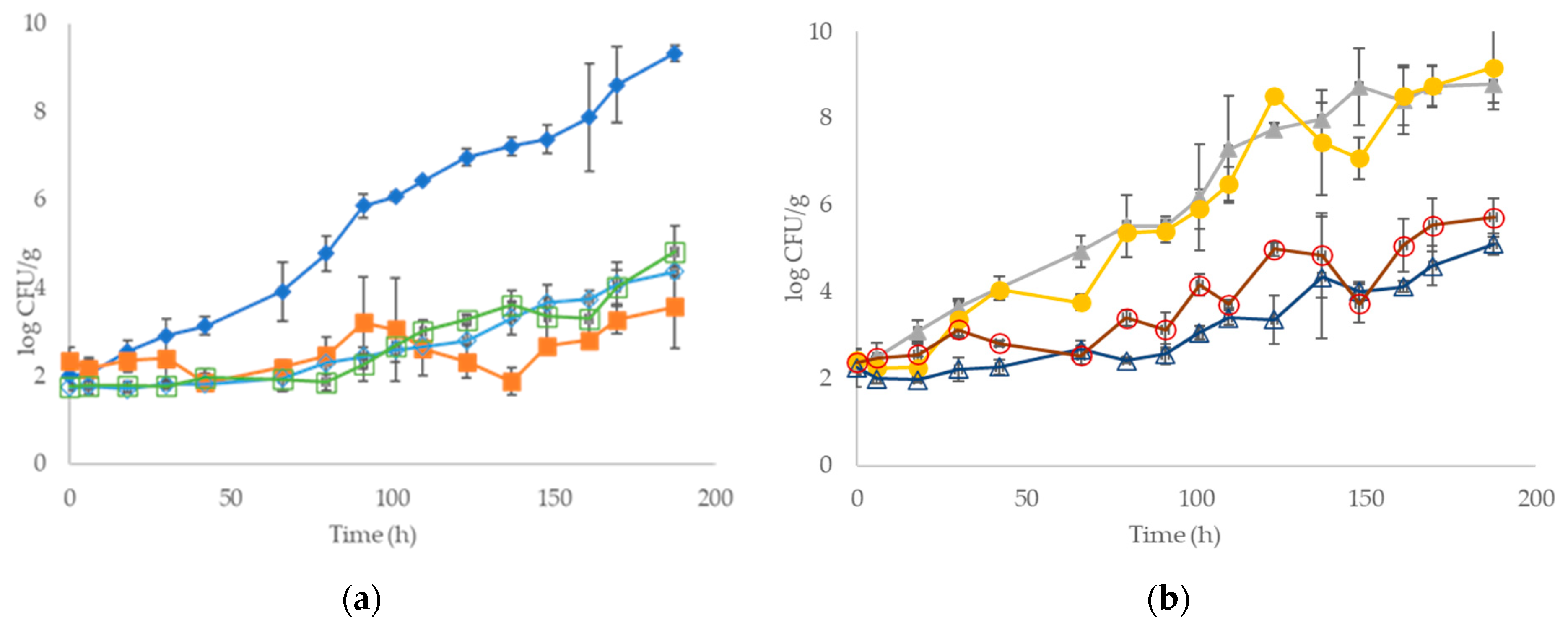
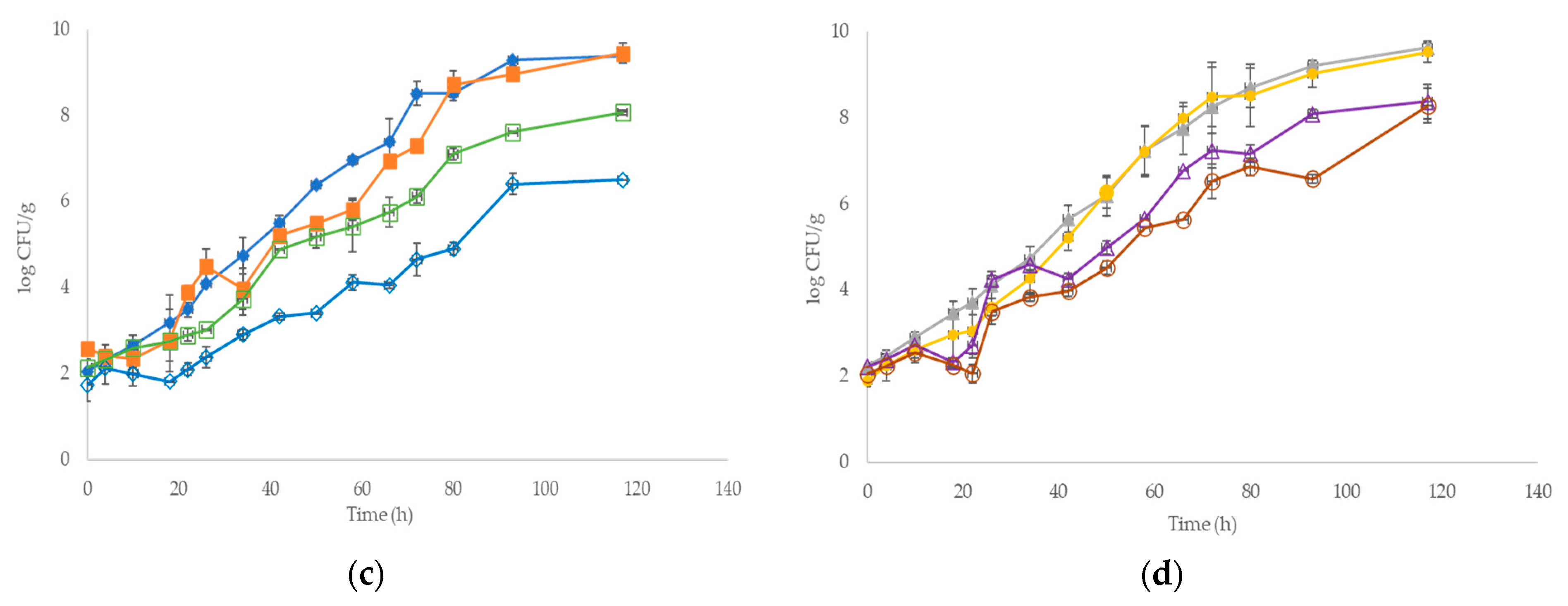
3.1. Development of Microbial Association and Sensory Analysis
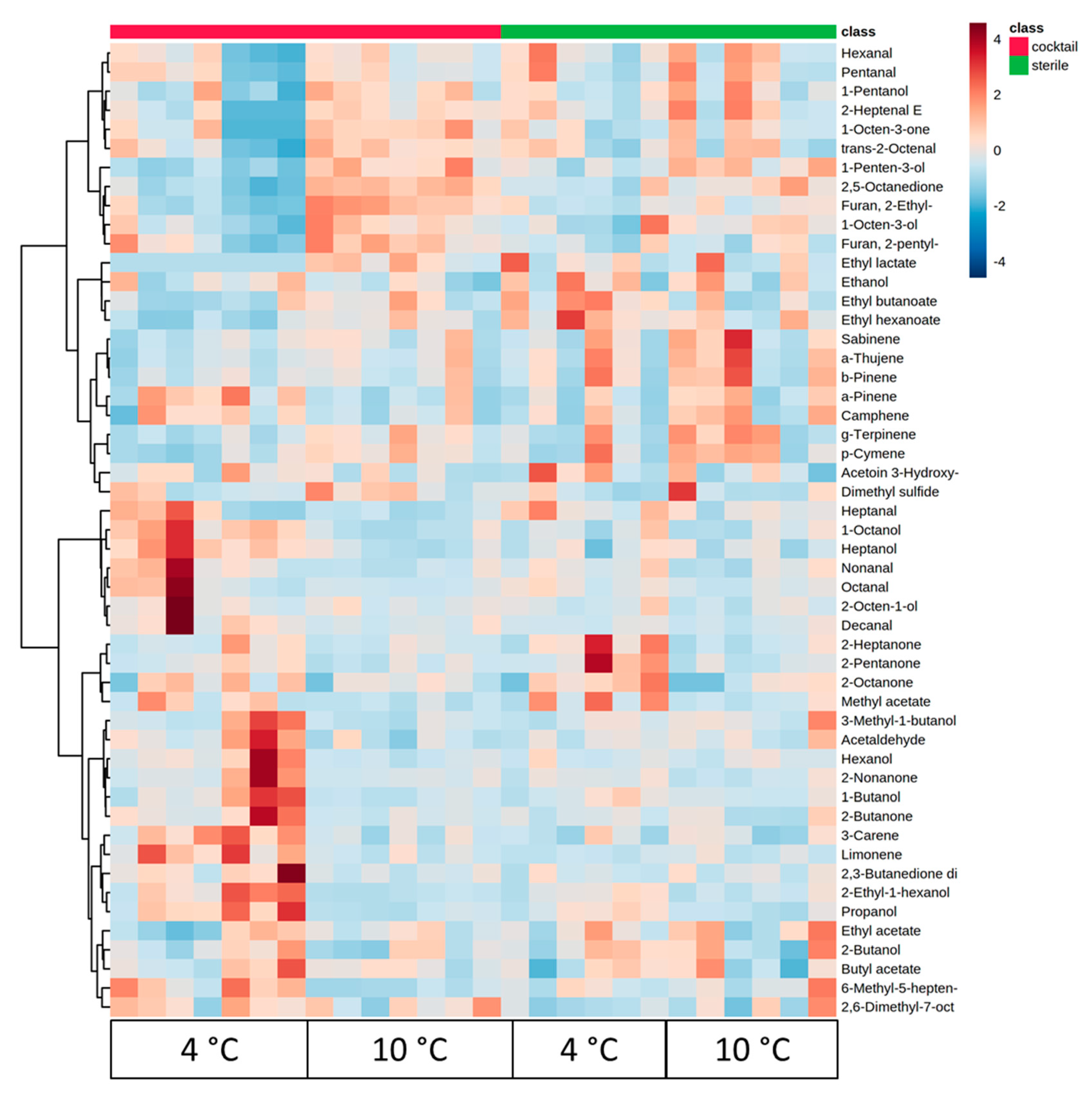
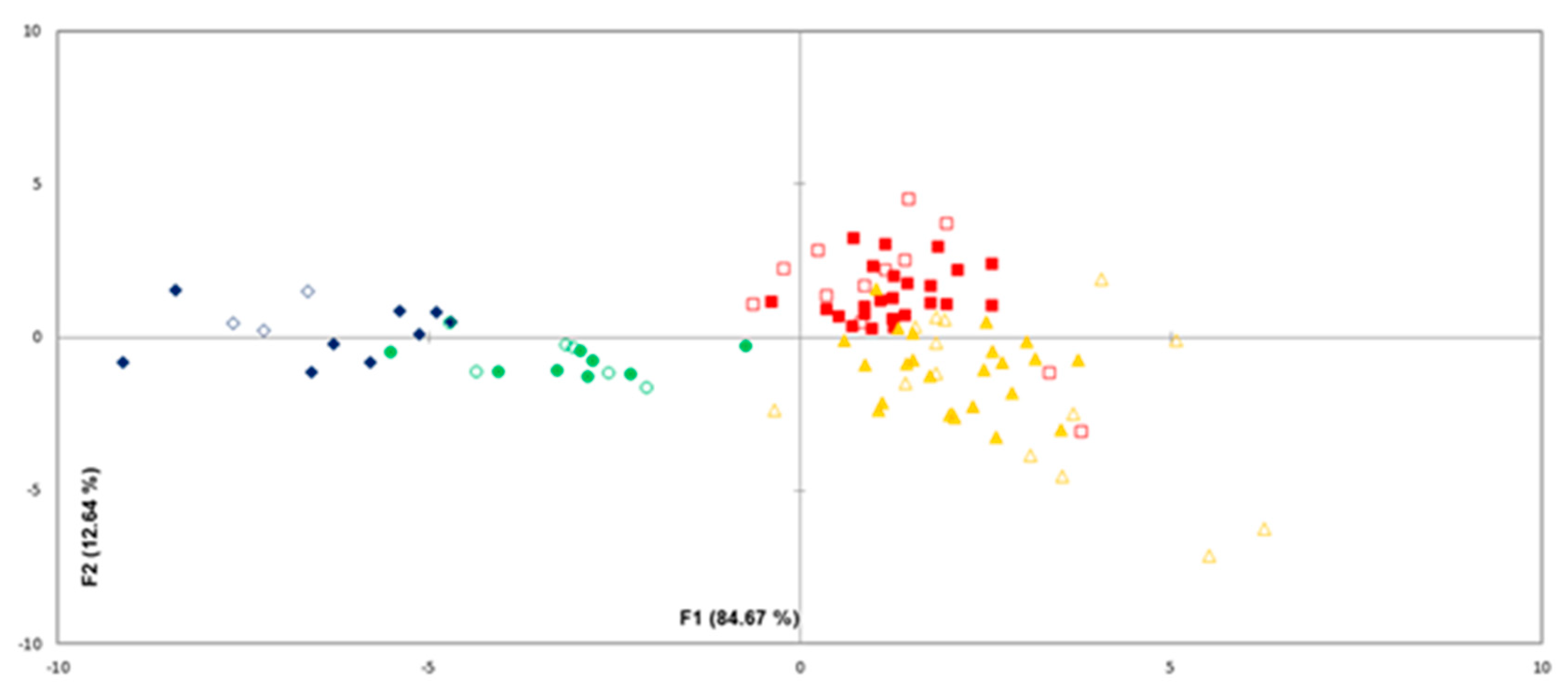
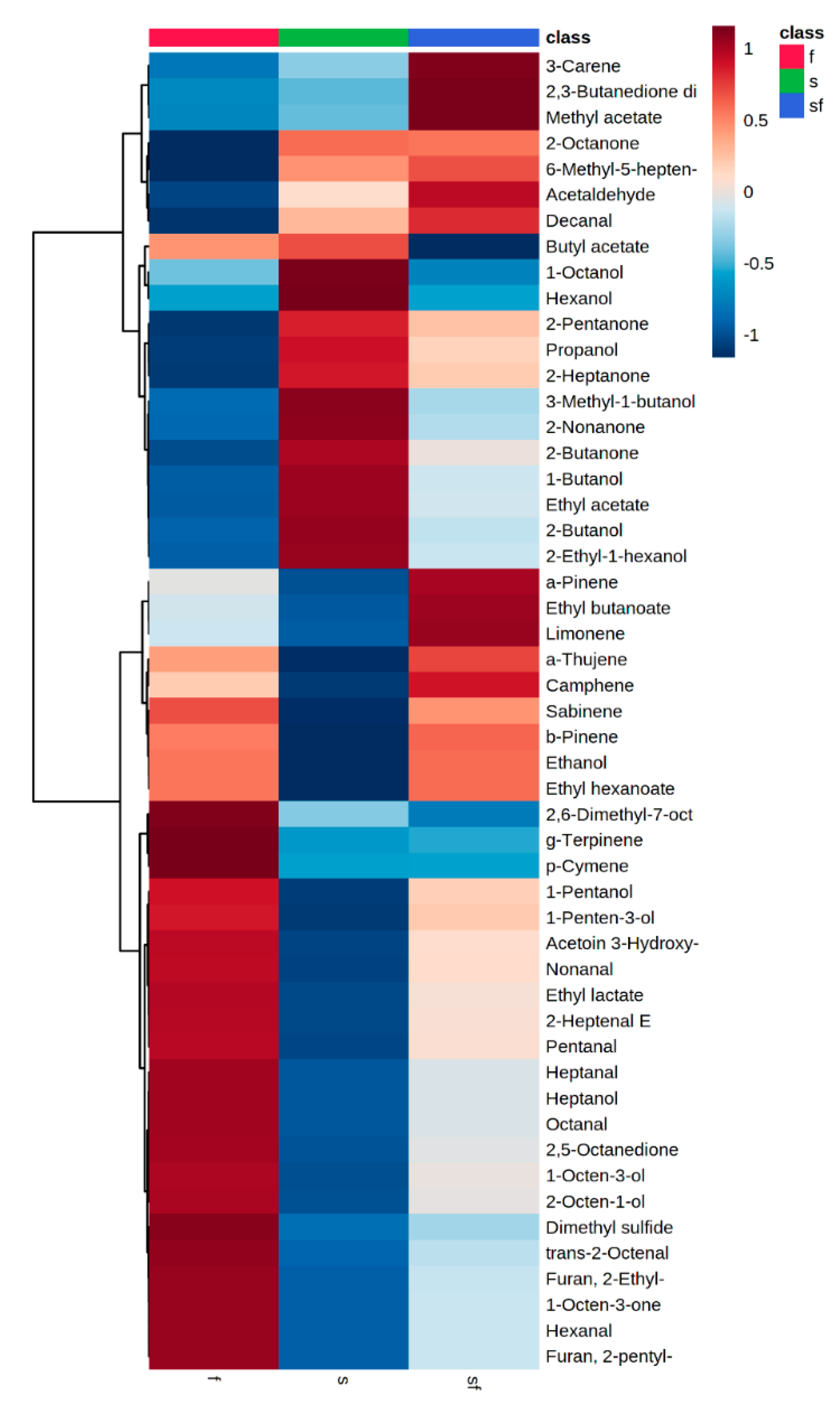
3.2. Discrimination of Sterile From Inoculated Meat Using Volatile Compounds
3.3. Correlation of Meat Sensory Classes with Volatile Compounds
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Casaburi, A.; Piombino, P.; Nychas, G.-J.; Villani, F.; Ercolini, D. Bacterial populations and the volatilome associated to meat spoilage. Int. J. Food Microbiol. 2015, 45, 83–102. [Google Scholar] [CrossRef] [PubMed]
- Doulgeraki, A.I.; Nychas, G.-J.E. Monitoring the succession of the biota grown on a selective medium for pseudomonads during storage of minced beef with molecular-based methods. Int. J. Food Microbiol. 2013, 34, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Ercolini, D.; Russo, F.; Torrieri, E.; Masi, P.; Villani, F. Changes in the spoilage-related microbiota of beef during refrigerated storage under different packaging conditions. Appl. Environ. Microbiol. 2006, 72, 4663–4671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ercolini, D.; Ferrocino, I.; Nasi, A.; Ndagijimana, M.; Vernocchi, P.; La Storia, A.; Laghi, L.; Mauriello, G.; Guerzoni, E.M.; Villani, F. Monitoring of Microbial Metabolites and Bacterial Diversity in Beef Stored under Different Packaging Conditions. Appl. Environ. Microbiol. 2011, 77, 7372–7381. [Google Scholar] [CrossRef] [Green Version]
- Odeyemi, O.A.; Alegbeleye, O.O.; Strateva, M.; Stratev, D. Understanding spoilage microbial community and spoilage mechanisms in foods of animal origin. Compr. Rev. Food Sci Food Saf. 2020, 19, 311–331. [Google Scholar] [CrossRef] [Green Version]
- Nychas, G.-J.E.; Marshall, D.L.; Sofos, J.N. Meat, Poultry, and Seafood. Microbiology: Fundamentals and Frontiers; Doyle, M.P., Beuchat, L.R., Eds.; ASM Press: Washington, DC, USA, 2007; pp. 105–140. [Google Scholar]
- Nychas, G.; Dillon, V.; Board, R. Glucose, the Key Substrate in the Microbiological Changes Occurring in Meat and Certain Meat Products. Biotechnol. Appl. Biochem. 1988, 10, 203–231. [Google Scholar]
- Doulgeraki, A.I.; Paramithiotis, S.; Kagkli, D.M.; Nychas, G.-J.E. Lactic acid bacteria population dynamics during minced beef storage under aerobic or modified atmosphere packaging conditions. Int. J. Food Microbiol. 2010, 27, 1028–1034. [Google Scholar] [CrossRef] [Green Version]
- Freiding, S.; Gutsche, K.A.; Ehrmann, M.A.; Vogel, R.F. Genetic screening of Lactobacillus sakei and Lactobacillus curvatus strains for their peptidolytic system and amino acid metabolism, and comparison of their volatilomes in a model system. Syst. Appl. Microbiol. 2011, 34, 311–320. [Google Scholar] [CrossRef]
- Nychas, G.-J.E.; Skandamis, P.N.; Tassou, C.C.; Koutsoumanis, K.P. Meat spoilage during distribution. Meat Sci. 2008, 78, 77–89. [Google Scholar] [CrossRef]
- Vasilopoulos, C.; De Maere, G.; De May, E.; Paelink, H.; De Vust, L.; Leroy, F. Technology induced of selection towards the spoilage microbiota of artisan-type cooked ham packed udner modified atmosphere. Food Microbiol. 2010, 27, 77–84. [Google Scholar] [CrossRef]
- Casaburi, A.; De Filippis, F.; Villani, F.; Ercolini, D. Activities of strains of Brochothrix thermosphacta in vitro and in meat. Food Res. Int. 2014, 62, 366–374. [Google Scholar] [CrossRef]
- Ercolini, D.; Russo, F.; Nasi, A.; Ferranti, P.; Villani, F. Mesophilic and psychrotrophic bacteria from meat and their spoilage potential in vitro and in beef. Appl. Environ. Microbiol. 2009, 75, 1990–2001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ercolini, D.; Ferrocino, I.; La Storia, A.; Mauriello, G.; Gigli, S.; Masi, P.; Villani, F. Development of spoilage microbiota in beef stored in nisin activated packaging. Int. J. Food Microbiol. 2010, 27, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Nieminen, T.T.; Vihavainen, E.; Paloranta, A.; Lehto, J.; Paulin, L.; Auvinen, P.; Solisma, M.; Björkroth, K.J. Characterization of psychrotrophic bacterial communities in modified atmosphere-packed meat with terminal restriction fragment length polymorphism. Int. J. Food Microbiol. 2011, 144, 360–366. [Google Scholar] [CrossRef]
- Pothakos, V.; Devlieghere, F.; Villani, F.; Björkroth, J.; Ercolini, D. Lactic acid bacteria and their controversial role in fresh meat spoilage. Meat Sci. 2015, 109, 66–74. [Google Scholar] [CrossRef]
- Casaburi, A.; Nasi, A.; Ferrocino, I.; Di Monaco, R.; Mauriello, G.; Villani, F.; Ercolini, D. Spoilage-related activity of Carnobacterium maltaromaticum strains in air-stored and vacuum-packed meat. Appl. Environ. Microbiol. 2011, 77, 382–7393. [Google Scholar] [CrossRef] [Green Version]
- Tsigarida, E.; Nychas, G.-J.E. Ecophysiological attributes of a Lactobacillus sp. and a Pseudomonas sp. on sterile beef fillets in relation to storage temperature and film permeability. J. Appl. Microbiol. 2001, 90, 696–705. [Google Scholar] [CrossRef]
- Papadopoulou, O.; Panagou, E.Z.; Mohareb, F.R.; Nychas, G.-J.E. Sensory and microbiological quality assessment of beef fillets using a portable electronic nose in tandem with support vector machine analysis. Food Res. Int. 2013, 50, 241–249. [Google Scholar] [CrossRef]
- Argyri, A.A.; Mallouchos, A.; Panagou, E.Z.; Nychas, G.-J.E. The dynamics of the HS/SPME-GC/MS as a tool to assess the spoilage of beef stored under different packaging and temperature conditions. Int. J. Food Microbiol. 2015, 193, 51–58. [Google Scholar] [CrossRef]
- Baranyi, J.; Roberts, T.A. A dynamic approach to predicting bacterial growth in food. Int. J. Food Microbiol. 1994, 23, 277–294. [Google Scholar] [CrossRef]
- Pavlidis, D.E.; Mallouchos, A.; Ercolini, D.; Panagou, E.Z.; Nychas, G.-J.E. A volatilomics approach for off-line discrimination of minced beef and pork meat and their admixture using HS-SPME GC/MS in tandem with multivariate data analysis. Meat Sci. 2019, 151, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Sinelnikov, I.V.; Han, B.; Wishart, D.S. Metaboanalyst 3.0—Making metabolomics more meaningful. Nucleic Acids Res. 2015, 43, W251–W257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otto, M. Chemometrics: Statistics and Computer Application in Analytical Chemistry, 3rd ed.; Wiley-VCH: Weinheim, Germany, 2007; pp. 184–200. [Google Scholar]
- Mohareb, F.; Iriondo, M.; Doulgeraki, A.I.; Van Hoek, A.; Aarts, H.; Cauchia, M.; Nychas, G.-J.E. Identification of meat spoilage gene biomarkers in Pseudomonas putida using gene profiling. Food Control 2015, 57, 152–160. [Google Scholar] [CrossRef] [Green Version]
- Argyri, A.A.; Doulgeraki, A.I.; Blana, V.A.; Panagou, E.Z.; Nychas, G.-J.E. Potential of a simple HPLC-based approach for the identification of the spoilage status of minced beef stored at various temperatures and packaging systems. Int. J. Food Microbiol. 2011, 150, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Ferrocino, I.; La Storia, A.; Torrieri, E.; Musso, S.S.; Mauriello, G.; Villani, F.; Ercolini, D. Antimicrobial packaging to retard the growth of spoilage bacteria and to reduce the release of volatile metabolites in meat stored under vacuum at 1 °C. J. Food Prot. 2013, 76, 52–58. [Google Scholar] [CrossRef]
- Hernandez-Macedo, M.L.; Contreras-Castillo, C.J.; Tsai, S.M.; Da Cruz, S.H.; Sarantopoulos, C.I.G.L.; Padula, M.; Dias, C.T.S. Gases and volatile compounds associated with microorganisms in blown pack spoilage of Brazilian vacuum-packed beef. Lett. Appl. Microbiol. 2012, 55, 467–475. [Google Scholar] [CrossRef]
- Cubero-Leon, E.; Peñalver, R.; Maquet, A. Review on metabolomics for food authentication. Food Res. Int. 2014, 60, 95–107. [Google Scholar] [CrossRef]
- Xiao, Z.; Xu, P. Acetoin Metabolism in Bacteria. Crit. Rev. Microbiol. 2007, 33, 127–140. [Google Scholar]
- Huang, M.; Oppermann, F.B.; Steinbiichel, A. Molecular characterization of the Pseudomonas putida 2,3-butanediol catabolic pathway. FEMS Microbiol. Lett. 1994, 124, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Insausti, H.K.; Beriain, M.J.; Gorraiz, C.; Purroy, A. Volatile Compounds of Raw Beef from 5 Local Spanish Cattle Breeds Stored Under Modified Atmosphere. J. Food Sci. 2002, 67, 1580–1589. [Google Scholar] [CrossRef]
- Nurjuliana, M.; Che Man, Y.B.; Mat Hashim, D.; Mohamed, A.K.S. Rapid identification of pork for halal authentication using the electronic nose and gas chromatography mass spectrometer with headspace analyzer. Meat Sci. 2011, 88, 638–644. [Google Scholar] [CrossRef] [PubMed]
- Leroy, F.; Vasilopoulos, C.; Van Hemelryck, S.; Falony, G.; De Vuyst, L. Volatile analysis of spoiled, artisan-type, modified-atmosphere-packaged cooked ham stored under different temperatures. Int. J. Food Microbiol. 2009, 26, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Jääskeläinen, E.; Hultman, J.; Parshintsev, J.; Riekkola, M.L.; Björkroth, J. Development of spoilage bacterial community and volatile compounds in chilled beef under vacuum or high oxygen atmospheres. Int. J. Food Microbiol. 2016, 223, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Saraiva, C.; Oliveira, I.; Silva, J.A.; Martins, C.; Ventanas, J.; García, C. Implementation of multivariate techniques for the selection of volatile compounds as indicators of sensory quality of raw beef. J. Food Sci. Technol. 2015, 52, 3887–3898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Resconi, V.C.; Bueno, M.; Escudero, A.; Magalhaes, D.; Ferreira, V.; Mar Campo, M. Ageing and retail display time in raw beef odour according to the degree of lipid oxidation. Food Chem. 2018, 242, 288–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulz, S.; Dickschat, J.S. Bacterial volatiles: The smell of small organisms. Nat. Prod. Rep. 2007, 24, 814–842. [Google Scholar] [CrossRef]
- Montel, M.C.; Masson, F.; Talon, R. Bacterial role in flavor development. Meat Sci. 1998, 49, 111–123. [Google Scholar] [CrossRef]
- Dainty, R.H.; Edwards, R.A.; Hibbard, C.M.; Marnewick, J.J. Volatile compound associated with microbial growth on normal and high pH beef stored at chill temperatures. J. Appl. Bacteriol. 1989, 66, 281–289. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Inoculation | Lag Phase (h) 1 | y0 (log10CFU/g) 2 | yend (log10CFU/g) 3 | μmax (h−1) 4 | R2 |
|---|---|---|---|---|---|
| Monoculture | |||||
| Pseudomonas fragi (F) | 9.31 ± 10.04 | 1.95 ± 0.22 | 10.51 ± 2.38 | 0.04 ± 0.01 | 0.99 |
| Pseudomonas putida (P) | 148.23 ± 15.43 | 2.26 ± 0.21 | - 5 | 0.01 ± 0.01 | - |
| Leuconostoc mesenteroides (M) | 30.98 ± 14.01 | 1.75 ± 0.20 | - | 0.02 ± 0.01 | 0.86 |
| Lactobacillus sakei (S) | 50.67 ± 16.45 | 30.98 ± 14.01 | - | 0.02 ± 0.00 | 0.96 |
| Dual Culture | |||||
| F + P (CF) | 7.39 ± 18.55 | 2.26 ± 0.32 | 8.86 ± 0.28 | 0.05 ± 0.01 | 0.98 |
| M + S (MF) | 50.67 ± 16.45 | 2.03 ± 0.19 | - | 0.02 ± 0.00 | 0.91 |
| Cocktail | |||||
| CF (cocktail) | 68.31 ± 17.44 | 2.07 ± 0.55 | 9.00 ± 0.57 | 0.04 ± 0.01 | 0.94 |
| MF (cocktail) | 17.80 ± 33.98 | 2.19 ± 0.42 | 6.82 ± 10.46 | 0.02 ± 0.01 | 0.84 |
| Inoculation | Lag Phase (h) 1 | y0 (log10CFU/g) 2 | yend (log10CFU/g) 3 | μmax (h−1) 4 | R2 |
|---|---|---|---|---|---|
| Monoculture | |||||
| P. fragi (F) | 1.24 ± 3.01 | 1.99 ± 0.14 | 10.18 ± 0.17 | 0.09 ± 0.00 | 1.00 |
| P. putida (P) | 13.87 ± 9.63 | 2.80 ± 0.39 | 9.34 ± 0.54 | 0.07 ± 0.01 | 0.95 |
| Ln. mesenteroides (M) | 8.82 ± 7.92 | 1.86 ± 0.33 | 7.93 ± 0.37 | 0.08 ± 0.01 | 0.96 |
| Lb. sakei (S) | 24.72 ± 6.32 | 2.02 ± 0.19 | 7.49 ± 0.31 | 0.06 ± 0.01 | 0.97 |
| Dual Culture | |||||
| F + P (CF) | 2.22 ± 3.3 | 2.35 ± 0.15 | 9.52 ± 0.14 | 0.08 ± 0.00 | 0.99 |
| M + S (MF) | 26.77 ± 6.17 | 2.36 ± 0.26 | 7.91 ± 0.31 | 0.10 ± 0.01 | 0.95 |
| Cocktail | |||||
| CF (cocktail) | 17.80± 5.09 | 1.73± 0.23 | 9.27± 0.29 | 0.09± 0.01 | 0.99 |
| MF (cocktail) | 10.31± 6.24 | 2.04± 0.22 | 8.28± 0.28 | 0.06± 0.01 | 0.98 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Papadopoulou, O.S.; Iliopoulos, V.; Mallouchos, A.; Panagou, E.Z.; Chorianopoulos, N.; Tassou, C.C.; Nychas, G.-J.E. Spoilage Potential of Pseudomonas (P. fragi, P. putida) and LAB (Leuconostoc mesenteroides, Lactobacillus sakei) Strains and Their Volatilome Profile during Storage of Sterile Pork Meat Using GC/MS and Data Analytics. Foods 2020, 9, 633. https://doi.org/10.3390/foods9050633
Papadopoulou OS, Iliopoulos V, Mallouchos A, Panagou EZ, Chorianopoulos N, Tassou CC, Nychas G-JE. Spoilage Potential of Pseudomonas (P. fragi, P. putida) and LAB (Leuconostoc mesenteroides, Lactobacillus sakei) Strains and Their Volatilome Profile during Storage of Sterile Pork Meat Using GC/MS and Data Analytics. Foods. 2020; 9(5):633. https://doi.org/10.3390/foods9050633
Chicago/Turabian StylePapadopoulou, Olga S., Vasilis Iliopoulos, Athanasios Mallouchos, Efstathios Z. Panagou, Nikos Chorianopoulos, Chrysoula C. Tassou, and George-John E. Nychas. 2020. "Spoilage Potential of Pseudomonas (P. fragi, P. putida) and LAB (Leuconostoc mesenteroides, Lactobacillus sakei) Strains and Their Volatilome Profile during Storage of Sterile Pork Meat Using GC/MS and Data Analytics" Foods 9, no. 5: 633. https://doi.org/10.3390/foods9050633