
Interspecific Hybridization of Transgenic Brassica napus and Brassica rapa—An Overview
by
 , , , and
, , , and
Soo-In Sohn
1,* ,
,
Senthil Kumar Thamilarasan
1 ,
,
Subramani Pandian
1 ,
,
Young-Ju Oh
2, 
Tae-Hun Ryu
1,
Gang-Seob Lee
1 and
Eun-Kyoung Shin
1 1
Department of Agricultural Biotechnology, National Institute of Agricultural Sciences, Rural Development Administration, Jeonju 54874, Korea
2
Institute for Future Environment Ecology Co., Ltd., Jeonju 54883, Korea
*
Author to whom correspondence should be addressed.
Genes 2022, 13(8), 1442; https://doi.org/10.3390/genes13081442
Submission received: 9 July 2022
/
Revised: 12 August 2022
/
Accepted: 12 August 2022
/
Published: 13 August 2022
(This article belongs to the Special Issue Genetic Research and Plant Breeding)
Abstract
:In nature, interspecific hybridization occurs frequently and can contribute to the production of new species or the introgression of beneficial adaptive features between species. It has great potential in agricultural systems to boost the process of targeted crop improvement. In the advent of genetically modified (GM) crops, it has a disadvantage that it involves the transgene escaping to unintended plants, which could result in non-specific weedy crops. Several crop species in the Brassica genus have close kinship: canola (Brassica napus) is an ancestral hybrid of B. rapa and B. oleracea and mustard species such as B. juncea, B. carinata, and B. nigra share common genomes. Hence, intraspecific hybridization among the Brassica species is most common, especially between B. napus and B. rapa. In general, interspecific hybrids cause numerous genetic and phenotypic changes in the parental lines. Consequently, their fitness and reproductive ability are also highly varied. In this review, we discuss the interspecific hybridization and reciprocal hybridization studies of B. napus and B. rapa and their potential in the controlled environment. Further, we address the fate of transgenes (herbicide resistance) and their ability to transfer to their progenies or generations. This could help us to understand the environmental influence of interspecific hybrids and how to effectively manage their transgene escape in the future.
1. Introduction
Globally, the cropping area of genetically modified (GM) crops has constantly increased since 1996 [1]. GM crops cause huge nuisances to the environment, such as super weeds and introgressive hybridization. Concerns regarding the environmental consequences of the release of transgenic crops have led to considerable research to reduce the degree of ambiguity surrounding the risk of transgene escape via hybridization [2]. While reports of hybridization in natural environments are the most conclusive proof that transgenes can escape by hybridization, they are insufficient to evaluate the complete frequency of hybridization [3]. On the other hand, reports of hybridization between crops and their relatives through artificial hand-pollination are valuable sources of information because they allow for the assessment of reproductive compatibility between species and the detection of undesirable species combinations. This helps us to conduct a conservative analysis of species that should be considered for their potential as targets for transgene escape in the local environment [2,4].
In this review, we aimed to discuss the potential transgene escape via interspecific hybrids in the genus Brassica, one of the important genera in the Brassicaceae family, which comprises 39 species [5]. It is mainly cultivated for its edible roots, stems, leaves, buds, flowers, mustard, and oilseeds [6]. Oilseed rape (Brassica napus L.) is an allotetraploid species that arose through a spontaneous hybridization of Brassica rapa L. and Brassica oleracea L. It has the complete diploid chromosome sets of the highly homologous A and C genomes of B. rapa and B. oleracea, respectively. Among the various GM crops, B. napus L. is widely cultivated and has a high potential for hybridization with the closely related Brassica species through interspecific hybridization. It can spontaneously hybridize with B. rapa in both greenhouse and field experiments [7,8,9,10,11,12,13,14]. One of the main issues in the cultivation of transgenic B. napus is that the transgene may have been transferred through hand pollination and/or spontaneously to their wild relatives/cultivars, with undesired ecological consequences that can increase the fitness and invasiveness of weedy populations [1]. Aside from that, GM crops and their transgenes spread via seed spillage during transportation and pollen-mediated gene transfers, resulting in feral populations [1,15]. If this occurs, weeds with GM traits may provide new and substantial weed control challenges [16]. The risk of crop genes transferring to weedy relatives is determined by their genetic and structural similarities as well as the strength of the transgenic selection in the weedy relative. The transfer of transgenes through introgression also depends on the fitness of the first and successive generations of hybrids [16,17,18]. However, the level of hybridization and introgression among the Brassica species is highly varied. Therefore, in this review, we provide an overview of a different combination of interspecific hybridization between transgenic B. napus and close relative Brassica species in controlled greenhouse conditions. In addition, we have discussed GM traits fitness in interspecific hybridization, further highlighted the fate of transgenes, and addressed the risk factors for cross-combination effects.
2. Interspecific Hybridization of Transgenic B. napus and B. rapa
Interspecific hybridization is a common and important evolutionary mechanism in the Brassicaceae family. Genome polyploidization, genome duplication, and gene flow maintenance may occur several times during evolution [19,20,21]. Parental cross or reciprocal interspecific hybridization and backcrossing are important factors that can result in significant differences in male and female fitness. It strongly suggests that the likelihood of gene transfer is influenced by a number of factors, including the origin of wild plants; genome constitution; population structure; mating system of the hybridizing plants; field experiment designs; weed control measures; several growing seasons; co-existing species; and the possibility of introgression [7,22,23,24].
2.1. Hybrid Generation: Brassica rapa (♀) × GM Brassica napus (♂)
The crossability of weedy relatives or cultivated species of B. rapa (♀) and GM B. napus (♂) has been dependent on various environmental factors such as spatial distribution, maternal and paternal traits, and field or controlled conditions. Accordingly, various studies have reported that hybridization in controlled greenhouse circumstances will result in higher opportunities for genetically modified organism (GMO) crops and prevent controlled transgene flow in nature. An overview on interspecific hybridization between transgenic B. napus and B. rapa is represented in Figure 1 and Table 1. Initially, Vacher et al. [25] studied transgenic F1 hybrids with high fertility and backcrossing abilities under various ecological factors. It may enhance the fitness of transgenic hybrids and wild relatives. The high frequency (27%) occurred in areas with a higher plant density than usual (25%). However, the first-generation backcross and F2 hybrids might slow down the process of transgene spread with lower fitness. Likewise, F1 hybrids had higher fitness and silique per plant than backcross generations [26]. Pallett [27] has found a 5 to 100% hybridization rate between wild UK B. rapa and transgenic B. napus in optimum conditions. Consequently, the weedy population is highly variable when the hybridization is carried out, even in controlled conditions. Concerned with heterospecific pollination (removal of any co-flowering or likelihood that plants may interact via pollinators), they even observed matromorphs and apomicts (an asexual mode of reproduction; the ovule develops into seeds without involving meiosis and fertilization) due to parental combinations. In particular, the F1 generation had a higher percentage of C genome and transgene presence in all progenies than the backcross generation. Similarly, Vacher [28] found a predominantly out-crossing rate and added counter selection of the wild weedy phenotype population of UK B. rapa (♀) against transgenic B. napus.
The hybridization occurred due to flowering and for longer periods of time, which were likely to receive pollen from the transgenic trait. Naturally, the weedy plants had wider stems/stem diameters. Transgenes are involved in promoting flowering in nature. However, it produced a lower number of seeds. Also found, maternal weeds are less fit due to the longer period of flowering, which has a higher probability of hybridization with GM crops. Xiao et al. [31] extended different varieties of B. rapa to exhibit different levels of crossability index under controlled greenhouse conditions and compared them to spontaneous hybridization outcrossing. Due to pollen adhesion on the stigma and pollen tubes in the style, their numbers were reduced during self-pollination and were highly related to the genotypes of the parents. Likewise, broken or precocious germination was found in all the interspecific hybrids. Similarly, Sohn et al. [4] observed moderate crossability between B. rapa ssp. and transgenic GM B. napus through hand pollination under controlled conditions, with 100% crossability indices in F1 hybrids. Due to callus tissue formation during seed development and hormonal imbalance, the combination of the parental lines may cause precocious or cracked seeds, resulting in smaller sizes and affecting seed germination.
2.2. Backcross Generation: B. rapa (♀) × F1 (♂) (B. rapa × GM B. napus)
The transgenic F1 hybrids are likely to transfer the transgenes into backcrossing populations as seed parents of B. rapa. Initially, in field experiments, introgression of transgenic B. napus and weedy B. rapa were grown together. The weedy plants of B. rapa sp. have produced herbicide-tolerant BC1 generations with the same morphology and chromosomes as B. rapa, but they are more fertile and produced as early as the BC1 generation [42]. The phosphinothricin (PPT)-tolerant backcross generation had a lower ratio of monogenic segregation than the PPT-susceptible mendelian segregation ratio, indicating that the PPT-tolerant plants are homozygous or hemizygous. During the initial hybridization event with B. rapa as female parent and PPT-tolerant hybrid plants from backcrossed used as male parent transgene, the transgene transmission was reduced in the subsequent generations [35]. Furthermore, these results corroborate the transgene flow process and lower fitness level in the first generation or offspring or BC1 as a seed parent for B. rapa [26,43]. From another perspective, Halfhill et al. [11] used green fluorescent protein GFP/Bt as a tracking tool to assess the transgene flow and easy-to-use monitoring qualities of GFP with an agronomically significant transgenic. Though herbicide tolerance can be detected more effectively in a large number of plants than in GFP, this requires a visual assay of each individual for accurate screening [38]. In addition, the evidence of 12 transgenic at similar rates are the largest sample ever examined, in contrast to [35], two independent herbicide tolerant canola produced BC1 plants at dramatically different rates. After repeated backcrosses, Halfhill et al. [9] extended the work with other lines of weedy B. napus and B. rapa varieties to document gene flow and show that the resultant transgenic plants have fewer chromosomes and take on the morphological characteristics of their weedy B. rapa parent. Even though the hybridization rate was similar for both the weedy and cultivar of B. rapa when they were co-occurring, transgenic B. napus depends on density and spatial distribution [44]. Subsequently, he proposed to mitigate the gene flow and several factors in order to underestimate the actual frequency of gene flow from crop to weedy plants. Hence, the additive transgene has been used to locate the two copies of the transgene expression in homozygous individuals of canola as well as in hemizygous individuals (F1, BC1F1 and BC2F2). The F1 generation contained 95 to 97% of the genetic nature of B. napus, while subsequent backcross generations lost 15 to 29% of the genetic content in the BC2F2 bulk population [32]. This was followed by transgene mitigating B. napus containing the dwarfing gene, which demonstrated that the transgene mitigating (TM) strategy was effective in limiting seed production and thereby mitigating transgene flow from B. napus to B. rapa. In subsequent generations, the deleterious allele would only be expressed in homozygous individuals, which would strongly reduce its ability to decrease fitness [33].
In another study, transgene mitigating with additional genetic load and interspecific competition with wheat or more weed-like conditions was imposed, but still effective in limiting transgene persistence in weedy relatives [30]. Then, hybridization frequencies of 1 to 17% were observed with B. napus varieties and B. rapa [9], extended with multiple independent transformed lines, and several experimental conditions and locations were used to observe hybridization frequencies. The backcross frequencies are very low (0.074%) compared to expected (2.5%) but have a high level of potential to produce transgenic seeds [38]. Hence, backcrossing with a single transgenic event under a wide range of field conditions with competitive and non-competitive species, they found a lower vegetative growth rate and reduced from the triploid F1 generation to the diploid BC2F2 generation for the transgenic progenies of B. rapa. Subsequently, Vacher et al. [25] demonstrated that 1.4 times more seed in hybrids and backcross generations enhanced relative fitness under high herbivore pressure or selection pressure with more complex environments, using high fertilities and high backcrossing abilities of F1 hybrids. Sutherland et al. [45] agreed with the results, stimulating herbivore to their transgenic hybrids may increase the fitness and numbers of their progenies. However, the absence of herbivore pressure and continuous backcross generations maintain the physiological characteristics and decrease in fitness of transgenic hybrids in contrast to Halfhill et al. [9]. Another study showed remarkably similar growth and nitrogen utilization efficiency when compared to backcross generations of B. napus and B. rapa and transgenic B. napus F1 hybrids. These parameters, meanwhile, were less favorable than those of the wild relative, B. rapa, indicating that transgenic hybrids are less adapted to their natural environmental conditions [46]. In similar case, multiple transgenic (GT) lines were used with wild B. rapa the frequency of hybrids in BC1 progenies was higher than Halfhill et al. [11], but the segregation ratio was significantly deviated from BC2 to BC3 [10]. There are various factors affecting hybridization success irrespective of the presence or absence of transgenes that are less fit than the parental weed populations [40]. The transgene persistence was measured over six years under agro-environmental conditions, as claimed in the first report. In B. rapa, introgression may have a local gene pool which shows reduced fertility in progenies though the parents had normal fertility. However, continuous advanced backcross hybrid generation may reduce (6.2% lower) the fitness of hybrids over time [41]. On the other hand, herbicide drift CP4 EPSPS selectively neutralizes transgenes, which does not affect the relative fitness relationship between the parental and backcross generation. Due to synchronizing the flowering time as early as possible, it may desynchronize from B. napus flowering time to reduce the potential gene flow. F1 hybrids may affect gene flow rates by preferentially pollinating with transgenic pollen rather than B. rapa pollen. It may be that sub-lethal application may be sufficient to alter the fitness and gene flow dynamics of transgenes [37].
3. Ploidy Determination for Transgenic Hybrids and Backcross Generation
Ploidy levels maintain desirable hybrid combinations during sexual reproduction in interspecific hybridization. During the hybridization process of a transgene from B. napus (2n = 38, AACC) to B. rapa (2n = 20, AA), a hybrid with (2n = 29, AAC) herbicide-resistant or transgenic lines was produced. Many researchers have observed that F1 hybrids produced from the hybridization of B. rapa and B. napus were triploid (AAC; 2n = 29) [4,9,11,35,45,47]. However, rather than a gene, the chromosome number determines an individual’s fitness in the backcross progenies [48]. Moreover, it was hypothesized that the loss of a C-chromosome during meiosis in backcross generation accounted for the lower transmission rate of a C chromosome in the BC1 generation [35,49,50]. Previously, Metz et al. [35] proposed that B. rapa transgenic individuals with 2n = 21 to 2n = 23 with transgenic TP2 produce AACC with a lower frequency of BC2, BC3, and BC4 populations by using only 1–4 herbicide-resistant individual plants, and that the frequency of a gene transmitted through individuals with 2n = 21 to 23 ranged from 8.7 to 10.6% in the backcross generation [51]. Contrastingly, the triploid AAC hybrids can transmit higher rates of 2n = 20 to 24 and 34 to 38, depending on the female parent. The number of chromosomes transmitted in the hybrid was found to be incomplete because a C chromosome had been introgressed onto another A or C chromosome [52]. Meanwhile, assessed from nuclear DNA content, the ploidy of the BC1F1 generation changed towards that of B. rapa [9]. However, it differed from B. rapa, indicating a small portion of the C genome, possibly as few as one or two chromosomes were present in the first meiotic division that gave BC1F1 plants. In the case of continuous backcrosses for BC2F2, the ploidy is stable after an intermating generation of BC2F2 Bulk [32].
4. Genetically Modified Herbicide Resistance Traits
The effects of hybridization will vary by trait, with certain qualities being more likely to promote weediness or invasiveness, and leading to reduced fitness in hybridization and introgression than others, details provided in Table 1. The CP4 EPSPS gene (5-enolpyruvulshikimate-3 phosphate synthase) in transgenic B. napus c.v. GT73 decreases binding affinity for glyphosate, conferring increased tolerance to glyphosate herbicide, and the GOX gene, which carries glyphosate oxidase, confers tolerance to glyphosate herbicide by degrading glyphosate into aminomethylphosphonic acid (AMPA). The traits were used in many studies [31,36,37]. The bar gene is responsible for phosphinothricin (PPT) tolerance in B. napus cv. Drakkar. This gene encodes an acetyltransferase that acetylates the free NH2 group of PPT to inactivate it. PPT inhibits glutamine synthetase, resulting in rapid ammonia accumulation and cell death [26,34,35]. The two gene constructs in B. napus cv. Westar include the Bt cry1 Ac gene, which is highly resistant to common defoliating lepidopterans such as the diamond black moth [53], and a plasmid containing GFP fluorescence was used to detect visual assay transgenic lines of GFP/BT events (GT1-9) [9,11,25,30,32,46] and another variant of GFP mGFP5er [10,38,54]. On another perspective, shortening flowering time to add early flowering genes (BrAGL20) with herbicide-resistance (bar) and hygromycin-resistance gene (hpt) in B. napus cv. “Youngsan” [4,55]. Subsequently, transgene mitigating genes (Δgai as intact tandem genes) were inserted into B. napus cv. Westar herbicide resistance ahasR (acetohydroxy acid synthase; conferring resistance to imidazolinone herbicides) [33,56].
5. Fate of Transgenes in Interspecific Hybrid Plants
The potential risk of transgenic B. napus plants with B. rapa weedy or cultivars, the gene frequency or gene transmission rate is inevitable for the subsequent generations and is very complex due to their chromosome numbers and fitness of the progenies. Metz et al. [35] was evaluated in the backcross population from BC1 to BC4 under selection pressure. The BC4 generation maintains a 10% frequency of transgene plants, which indicates that a large resource of transgenic herbicide tolerance may unintentionally gene flow into weedy populations. The PPT-tolerant of BC1 and their subsequent generations of BC2 and BC3 yielded PPT-susceptible plants, which might be the transgene presence on one of the chromosomes of the C genome transmitted at a low frequency after a few generations [44]. Suggestively, transgenes can more safely integrate into the C-chromosome than into the A chromosome, which could reduce the risk of introgression in nature [57]. Subsequently, Zhu et al. [10] observed three types of genetic behavior for PPT tolerant B. napus when crossed with B. rapa. The first one is to inherit transgenes. Secondly, small portions of the seeds contained transgenes located on a non-homologous C chromosome. During successive backcrossing, the C chromosome could be lost and, thirdly, through the recombination between two genomes, the C chromosome might be incorporated into an A chromosome. Later on, Lu et al. [51] agreed with the results and proposed various statistical models with and without selection pressure. The frequency of the A chromosome transgene did not vary, and the transgene’s transmission rate in both selections was 50%. However, the C chromosome frequency varied from 9 to 40% until BC3, when herbicidal selection stabilized the transgene at around 6% in BC4 and BC5. Tomiuk et al. [50] have not admitted the statement of integration site without more detailed genetic information about the transgenic lines of B. napus. The frequency of homologous and homeologous recombination events, as well as the persistence time of transgenic A or C chromosomes in backcross populations, determines the safety of an integration site. However, the herbicide-resistant gene was transferred from B. napus to B. rapa. The transgene may integrate into the B. napus A-set of chromosome [41]. In the case that the transgene is found on the C genome, the transgene will be deleted or greater genomic incompatibility in the next generation, resulting in no transgenic backcrosses. This could be an important investigation option for decreasing introgression [11,35]. Another study found that a transgene carried by the C chromosome is less likely to be transferred in a B. rapa background than a gene carried by the A chromosome, and the chance of transfer varies across the C genome [52]. Even though it is a fact that the A and C genomes share a large degree of similarity, the safe-spot idea has been questioned by several authors, and contradicting evidence suggests that transgene insertion position may not lead to greater biosafety in terms of gene flow [9,11,50,58]. Subsequently, another piece of statistical evidence with the biased and unbiased model using a mixed population of different C chromosome numbers is the relative fitness of B. rapa BC1 and F1 hybrids. The possible way for introgression with the transgene in the C chromosome to have a positive effect on fitness is by making the plant herbicide-resistant. An extra chromosome leads to aneuploidy. Another way is by using homeologous recombination. The transgene on the C chromosome might be integrated into the A genome. This might happen during the F1 hybrid’s meiosis stage. At this stage, F1 hybrid introgression is substantially less likely for a transgene that is already on genome A [59].
Overall, B. napus is an economically important crop for improvement through the addition of commercially released transgenic traits (herbicide resistance, Bt, and TuMV). GMO traits could have negative effects on non-target species. Crops realize their environmental harm through gene flow and their effect on interspecific hybridization to B. rapa subspecies or viable seeds from transgenic hybrids. Gene flow can be widespread enough to pass genes into wild relatives even when those genes are carried on unshared non-homologous chromosomes. In some GM traits, using multiple transgenes may be difficult to detect in weed populations unless the weed populations have limiting factors such as abiotic stress or herbivores. In particular, novel genes have the potential to create weed issues by providing novel traits that enable weeds to compete better, produce more seeds, and grow widely. In the review, we mainly focus on likely crops for B. napus such as B. rapa being involved in stable introgression, F1 hybrids, and their selfing progenies having increased the ploidy level (enhance the plant’s evolution fitness), genetic diversity of the wild relatives, and hormonal imbalance of the seeds, such as vivipary or precocious germination effect on the seeds. This could be an environmental risk for a transgene trait to persist in nature through gene flow. To reduce the gene flow, transgenes may integrate into the C genome, the transgenic plants maintained in controlled greenhouse condition, designated experimental farm, spatial distribution, and variation between the pollen recipients.
6. Conclusions and Future Perspective
In conclusion, we updated the progress that has been made to date in the use of interspecific hybridization of transgenic B. napus and B. rapa wild, weed, and cultivars. The hybridization in controlled environments such as greenhouses, experimental fields, and cage setups allows the GM B. napus to successfully hybridize with several B. rapa subspecies. It can generate numerous fertile and viable generations and pass the herbicide-resistant transgene to their offspring. Artificial hand pollination with GM B. napus produced 100% outcrossing rate in a greenhouse environment. However, spontaneous hybridization has an outcrossing rate in the field that ranges from 0.02 to 2.78% [31,60]. In comparison to greenhouse settings, the outcrossing rate is significantly lower because of many external factors. All the way through, there is no control of the transgene spread, not even using a transgene mitigation system in crops, in nature, or in greenhouse conditions. Thus, what will be the future direction of transgenic research to continue to control or mitigate the transgene spread in wild or weed populations? Previous research indicates that greenhouse containment is the best strategy for preventing natural gene flow. Another possibility is that the transgene can be transferred into the C chromosome, which could be used to eliminate it in subsequent backcross generations. The recent technological advances in genome sequencing, genotyping-by-sequencing, transcriptomic, high throughput-phenomics platforms, machine learning algorithms using methods to discriminate the transgenic plants in fields and controlled conditions, and most recent conditionally accepted methods of genome editing are being used to develop improved crop plants with different flower or leaf colors to accumulate anthocyanin that could help to eliminate transgenic volunteers or weeds and control the gene flow.
Author Contributions
Conceptualization: S.-I.S. and S.K.T.; writing–original draft preparation: S.-I.S., S.K.T., S.P., Y.-J.O., T.-H.R., G.-S.L. and E.-K.S.; writing—review and editing: S.K.T., S.P. and Y.-J.O.; project administration: S.-I.S.; funding acquisition: S.-I.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Research Program for Agricultural Science and Technology Development (Project. No. PJ01672604) and supported by the 2022 Fellowship program (Project. No. PJ01672604 and PJ01494301) (S.K.T. and S.P.) of the National Institute of Agricultural Sciences, Rural Development Administration, Republic of Korea.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sohn, S.-I.; Pandian, S.; Oh, Y.-J.; Kang, H.-J.; Ryu, T.-H.; Cho, W.-S.; Shin, E.-K.; Shin, K.-S. A Review of the Unintentional Release of Feral Genetically Modified Rapeseed into the Environment. Biology 2021, 10, 1264. [Google Scholar] [CrossRef]
- FitzJohn, R.G.; Armstrong, T.T.; Newstrom-Lloyd, L.E.; Wilton, A.D.; Cochrane, M. Hybridisation within Brassica and allied genera: Evaluation of potential for transgene escape. Euphytica 2007, 158, 209–230. [Google Scholar] [CrossRef]
- Armstrong, T.T.; Fitzjohn, R.G.; Newstrom, L.E.; Wilton, A.D.; Lee, W.G. Transgene escape: What potential for crop–wild hybridization? Mol. Ecol. 2005, 14, 2111–2132. [Google Scholar] [CrossRef] [PubMed]
- Sohn, S.I.; Oh, Y.J.; Lee, K.R.; Ko, H.C.; Cho, H.S.; Lee, Y.H.; Chang, A. Characteristics analysis of F1 hybrids between genetically modified Brassica napus and B. rapa. PLoS ONE 2016, 11, 1–18. [Google Scholar] [CrossRef]
- Katche, E.; Quezada-Martinez, D.; Katche, E.I.; Vasquez-Teuber, P.; Mason, A.S. Interspecific hybridization for Brassica crop improvement. Crop Breed. Genet. Genom. 2019, 1–32. [Google Scholar]
- Rakow, G. Species origin and economic importance of Brassica. In Brassica; Springer: Berlin/Heidelberg, Germany, 2004; pp. 3–11. [Google Scholar]
- Jorgensen, R.B.; Andersen, B. Spontaneous Hybridization Between Oilseed Rape (Brassica napus) and Weedy B. campestris (Brassicaceae): A Risk of Growing Genetically Modified Oilseed Rape. Am. J. Bot. 1994, 81, 1620–1626. [Google Scholar] [CrossRef]
- Mikkelsen, T.R.; Jensen, J.; Jørgensen, R.B. Inheritance of oilseed rape (Brassica napus) RAPD markers in a backcross progeny with Brassica campestris. Theor. Appl. Genet. 1996, 92, 492–497. [Google Scholar] [CrossRef] [PubMed]
- Halfhill, M.D.; Millwood, R.J.; Raymer, P.L.; Stewart, C.N., Jr. undefined Bt-transgenic oilseed rape hybridization with its weedy relative, Brassica rapa. Environ. Biosafety Res. 2002, 1, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Lawrence, J.R.; Warwick, S.I.; Mason, P.; Braun, L.; Halfhill, M.D.; Stewart, C.N., Jr. Inheritance of GFP-Bt transgenes from Brassica napus in backcrosses with three wild B. rapa accessions. Environ. Biosafety Res. 2004, 3, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Warwick, S.I.; Simard, M.-J.; Légère, A.; Beckie, H.J.; Braun, L.; Zhu, B.; Mason, P.; Séguin-Swartz, G.; Stewart, C.N. Hybridization between transgenic Brassica napus L. and its wild relatives: Brassica rapa L., Raphanus raphanistrum L., Sinapis arvensis L., and Erucastrum gallicum (Willd.) O.E. Schulz. Theor. Appl. Genet. 2003, 107, 528–539. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, M.J.; Elliott, L.J.; Allainguillaume, J.; Shaw, M.W.; Norris, C.; Welters, R.; Alexander, M.; Sweet, J.; Mason, D.C. Hybridization between Brassica napus and B. rapa on a national scale in the United Kingdom. Science 2003, 302, 457–459. [Google Scholar] [CrossRef] [PubMed]
- Simard, M.-J.; Legere, A.; Warwick, S.I. Transgenic Brassica napus fields and Brassica rapa weeds in Quebec: Sympatry and weed-crop in situ hybridization. Botany 2006, 84, 1842–1851. [Google Scholar] [CrossRef]
- Landbo, L.; Jørgensen, R.B. Seed germination in weedy Brassica campestris and its hybrids with B. napus: Implications for risk assessment of transgenic oilseed rape. Euphytica 1997, 97, 209–216. [Google Scholar] [CrossRef]
- Clark, M.; Maselko, M. Transgene biocontainment strategies for molecular farming. Front. Plant Sci. 2020, 11, 210. [Google Scholar] [CrossRef]
- Song, X.; Yan, J.; Zhang, Y.; Li, H.; Zheng, A.; Zhang, Q.; Wang, J.; Bian, Q.; Shao, Z.; Wang, Y.; et al. Gene Flow Risks from Transgenic Herbicide-Tolerant Crops to Their Wild Relatives Can Be Mitigated by Utilizing Alien Chromosomes. Front. Plant Sci. 2021, 12, 1–20. [Google Scholar] [CrossRef]
- Jenczewski, E.; Eber, F.; Grimaud, A.; Huet, S.; Lucas, M.O.; Monod, H.; Chevre, A.M. PrBn, a major gene controlling homeologous pairing in oilseed rape (Brassica napus) haploids. Genetics 2003, 164, 645–653. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xu, A.; Liang, F.; Yao, X.; Wang, Y.; Liu, X.; Zhang, Y.; Dalelhan, J.; Zhang, B.; Qin, M. Screening of clubroot-resistant varieties and transfer of clubroot resistance genes to Brassica napus using distant hybridization. Breed. Sci. 2018, 68, 258–267. [Google Scholar] [CrossRef]
- Adams, K.L.; Wendel, J.F. Polyploidy and genome evolution in plants. Curr. Opin. Plant Biol. 2005, 8, 135–141. [Google Scholar] [CrossRef]
- Adams, K.L. Evolution of duplicate gene expression in polyploid and hybrid plants. J. Hered. 2007, 98, 136–141. [Google Scholar] [CrossRef] [PubMed]
- Schranz, M.E.; Lysak, M.A.; Mitchell-Olds, T. The ABC’s of comparative genomics in the Brassicaceae: Building blocks of crucifer genomes. Trends Plant Sci. 2006, 11, 535–542. [Google Scholar] [CrossRef] [PubMed]
- Hauser, T.P.; Damgaard, C.; Jørgensen, R.B. Frequency-dependent fitness of hybrids between oilseed rape (Brassica napus) and weedy B. rapa (Brassicaceae). Am. J. Bot. 2003, 90, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Heyn, F.W. Analysis of unreduced gametes in the Brassiceae by crosses between species and ploidy levels. Z. Pflanzenzuchtg 1977, 78, 13–30. [Google Scholar]
- Scheffler, J.A.; Dale, P.J. Opportunities for gene transfer from transgenic oilseed rape (Brassica napus) to related species. Transgenic. Res. 1994, 3, 263–278. [Google Scholar] [CrossRef]
- Vacher, C.; Weis, A.E.; Hermann, D.; Kossler, T.; Young, C.; Hochberg, M.E. Impact of ecological factors on the initial invasion of Bt transgenes into wild populations of birdseed rape (Brassica rapa). Theor. Appl. Genet. 2004, 109, 806–814. [Google Scholar] [CrossRef]
- Ammitzbøll, H.; Mikkelsen, T.N.; Jørgensen, R.B. Transgene expression and fitness of hybrids between GM oilseed rape and Brassica rapa. Environ. Biosafety Res. 2005, 4, 3–12. [Google Scholar] [CrossRef]
- Pallett, D.W.; Huang, L.; Cooper, J.I.; Wang, H. Within-population variation in hybridisation and transgene transfer between wild Brassica rapa and Brassica napus in the UK. Ann. Appl. Biol. 2006, 148, 147–155. [Google Scholar] [CrossRef]
- Vacher, C.; Kossler, T.M.; Hochberg, M.E.; Weis, A.E. Impact of interspecific hybridization between crops and weedy relatives on the evolution of flowering time in weedy phenotypes. PLoS ONE 2011, 6, 2–8. [Google Scholar] [CrossRef]
- Xiao, L.; Lu, C.; Zhang, B.; Bo, H.; Wu, Y.; Wu, G.; Cao, Y.; Yu, D. Gene transferability from transgenic Brassica napus L. to various subspecies and varieties of Brassica rapa. Transgenic. Res. 2009, 18, 733–746. [Google Scholar] [CrossRef]
- Mikkelsen, T.R.; Andersen, B.; Bagger Jørgensen, R. The risk of crop transgene spread. Nature 1996, 380, 31. [Google Scholar] [CrossRef]
- Metz, P.L.J.; Jacobsen, E.; Nap, J.P.; Pereira, A.; Stiekema, W.J. The impact on biosafety of the phosphinothricin-tolerance transgene in inter-specific B. rapa× B. napus hybrids and their successive backcrosses. Theor. Appl. Genet. 1997, 95, 442–450. [Google Scholar] [CrossRef]
- Hauser, T.P.; Jørgensen, R.B. Fitness of backcross and F 2 hybrids between weedy Brassica rapa and oilseed rape (B. napus). Heredity 1998, 81, 436–443. [Google Scholar] [CrossRef]
- Halfhill, M.D.; Richards, H.A.; Mabon, S.A.; Stewart, C.N. Expression of GFP and Bt transgenes in Brassica napus and hybridization with Brassica rapa. Theor. Appl. Genet. 2001, 103, 659–667. [Google Scholar] [CrossRef]
- Halfhill, M.D.; Zhu, B.; Warwick, S.I.; Raymer, P.L.; Millwood, R.J.; Weissinger, A.K.; Stewart, C.N., Jr. Hybridization and backcrossing between transgenic oilseed rape and two related weed species under field conditions. Environ. Biosafety Res. 2004, 3, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Halfhill, M.D.; Millwood, R.J.; Weissinger, A.K.; Warwick, S.I.; Stewart, C.N. Additive transgene expression and genetic introgression in multiple green-fluorescent protein transgenic crop × weed hybrid generations. Theor. Appl. Genet. 2003, 107, 1533–1540. [Google Scholar] [CrossRef]
- Al-Ahmad, H.; Gressel, J. Mitigation using a tandem construct containing a selectively unfit gene precludes establishment of Brassica napus transgenes in hybrids and backcrosses with weedy Brassica rapa. Plant Biotechnol. J. 2006, 4, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Rose, C.W.; Millwood, R.J.; Moon, H.S.; Rao, M.R.; Halfhill, M.D.; Raymer, P.L.; Warwick, S.I.; Al-Ahmad, H.; Gressel, J.; Stewart, C.N. Genetic load and transgenic mitigating genes in transgenic Brassica rapa (field mustard)× Brassica napus (oilseed rape) hybrid populations. BMC Biotechnol. 2009, 9, 1–14. [Google Scholar] [CrossRef]
- Sutherland, J.P.; Justinova, L.; Poppy, G.M. The responses of crop–wild Brassica hybrids to simulated herbivory and interspecific competition: Implications for transgene introgression. Environ. Biosafety Res. 2006, 5, 15–25. [Google Scholar] [CrossRef]
- Halfhill, M.D.; Sutherland, J.P.; Moon, H.S.; Poppy, G. u Y. m; Warwick, S.I.; Weissinger, A.K.; Rufty, T.W.; Raymer, P.L.; Stewart, C.N., Jr. Growth, productivity, and competitiveness of introgressed weedy Brassica rapa hybrids selected for the presence of Bt cry1Ac and gfp transgenes. Mol. Ecol. 2005, 14, 3177–3189. [Google Scholar] [CrossRef]
- Warwick, S.I. Gene flow between GM crops and related species in Canada. First Decad. Herbic. Resist. Crop. Canada. Top. Can. Weed Sci. 2007, 4, 101–113. [Google Scholar]
- Warwick, S.I.; Légère, A.; Simard, M.-J.; James, T. Do escaped transgenes persist in nature? The case of an herbicide resistance transgene in a weedy Brassica rapa population. Mol. Ecol. 2008, 17, 1387–1395. [Google Scholar] [CrossRef]
- Londo, J.P.; Bautista, N.S.; Sagers, C.L.; Lee, E.H.; Watrud, L.S. Glyphosate drift promotes changes in fitness and transgene gene flow in canola (Brassica napus) and hybrids. Ann. Bot. 2010, 106, 957–965. [Google Scholar] [CrossRef] [PubMed]
- Luijten, S.H.; Schidlo, N.S.; Meirmans, P.G.; de Jong, T.J. Hybridisation and introgression between Brassica napus and B. rapa in the Netherlands. Plant Biol. 2015, 17, 262–267. [Google Scholar] [CrossRef]
- Lu, C.; Kato, M. Fertilization fitness and relation to chromosome number in interspecific progeny between Brassica napus and B. rapa: A comparative study using natural and resynthesized B. napus. Breed. Sci. 2001, 51, 73–81. [Google Scholar] [CrossRef]
- McGrath, J.M.; Quiros, C.F. Generation of alien chromosome addition lines from synthetic Brassica napus: Morphology, cytology, fertility, and chromosome transmission. Genome 1990, 33, 374–383. [Google Scholar] [CrossRef]
- Tomiuk, J.; Hauser, T.P.; Bagger-Jørgensen, R. A- or C-chromosomes, does it matter for the transfer of transgenes from Brassica napus. Theor. Appl. Genet. 2000, 100, 750–754. [Google Scholar] [CrossRef]
- Lu, C.; Kato, M.; Kakihara, F. Destiny of a transgene escape from Brassica napus into Brassica rapa. Theor. Appl. Genet. 2002, 105, 78–84. [Google Scholar] [CrossRef]
- Leflon, M.; Eber, F.; Letanneur, J.C.; Chelysheva, L.; Coriton, O.; Huteau, V.; Ryder, C.D.; Barker, G.; Jenczewski, E.; Chèvre, A.M. Pairing and recombination at meiosis of Brassica rapa (AA) × Brassica napus (AACC) hybrids. Theor. Appl. Genet. 2006, 113, 1467–1480. [Google Scholar] [CrossRef] [PubMed]
- Londo, J.P.; Bollman, M.A.; Sagers, C.L.; Lee, E.H.; Watrud, L.S. Changes in fitness-associated traits due to the stacking of transgenic glyphosate resistance and insect resistance in Brassica napus L. Heredity 2011, 107, 328–337. [Google Scholar] [CrossRef]
- Snow, A.A.; Andersen, B.; Jørgensen, R.B. Costs of transgenic herbicide resistance introgressed from Brassica napus into weedy B. rapa. Mol. Ecol. 1999, 8, 605–615. [Google Scholar] [CrossRef]
- Stewart, C.N.; All, J.N.; Raymer, P.L.; Ramachandran, S. Increased fitness of transgenic insecticidal rapeseed under insect selection pressure. Mol. Ecol. 1997, 6, 773–779. [Google Scholar] [CrossRef]
- Moon, H.S.; Halfhill, M.D.; Good, L.L.; Raymer, P.L.; Neal Stewart, C. Characterization of directly transformed weedy Brassica rapa and introgressed B. rapa with Bt cry1Ac and gfp genes. Plant Cell Rep. 2007, 26, 1001–1010. [Google Scholar] [CrossRef]
- Hong, J.K.; Kim, S.-Y.; Kim, K.-S.; Kwon, S.-J.; Kim, J.S.; Kim, J.A.; Lee, S.I.; Lee, Y.-H. Overexpression of a Brassica rapa MADS-box gene, BrAGL20, induces early flowering time phenotypes in Brassica napus. Plant Biotechnol. Rep. 2013, 7, 231–237. [Google Scholar] [CrossRef]
- Al-Ahmad, H.; Dwyer, J.; Moloney, M.; Gressel, J. Mitigation of establishment of Brassica napus transgenes in volunteers using a tandem construct containing a selectively unfit gene. Plant Biotechnol. J. 2006, 4, 7–21. [Google Scholar] [CrossRef] [PubMed]
- Guan, Z.-J.; Shen, B.-C.; Wei, W.; Tang, Z.-X.; Stewart, C.; Neal, J. Loss of C-genome-specific markers during transgene introgression from Brassica napus to wild Brassica juncea. Nord. J. Bot. 2020, 38, 1–5. [Google Scholar] [CrossRef]
- Hansen, L.B.; Siegismund, H.R.; Jørgensen, R.B. Progressive introgression between Brassica napus (oilseed rape) and B. rapa. Heredity 2003, 91, 276–283. [Google Scholar] [CrossRef] [PubMed]
- De Jong, T.J.; Hesse, E. Selection against hybrids in mixed populations of Brassica rapa and Brassica napus: Model and synthesis. N. Phytol. 2012, 194, 1134–1142. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.-C.; Wang, P.-S.; Yang, J.-L.; Hong, H.; Lin, T.-K.; Tu, Y.-K.; Kuo, B.-J. Using a zero-inflated model to assess gene flow risk and coexistence of Brassica napus L. and Brassica rapa L. on a field scale in Taiwan. Bot. Stud. 2020, 61, 17. [Google Scholar] [CrossRef]
- Tu, Y.-K.; Chen, H.-W.; Tseng, K.-Y.; Lin, Y.-C.; Kuo, B.-J. Morphological and genetic characteristics of F(1) hybrids introgressed from Brassica napus to B. rapa in Taiwan. Bot. Stud. 2020, 61, 1. [Google Scholar] [CrossRef]
- Mikhaylova, E.V.; Kuluev, B.R.; Khaziakhmetov, R.M. Assessment of the propensity for hybridization between genetically modified oilseed rape and its nontransgenic relatives. Russ. J. Genet. Appl. Res. 2016, 6, 684–697. [Google Scholar] [CrossRef]
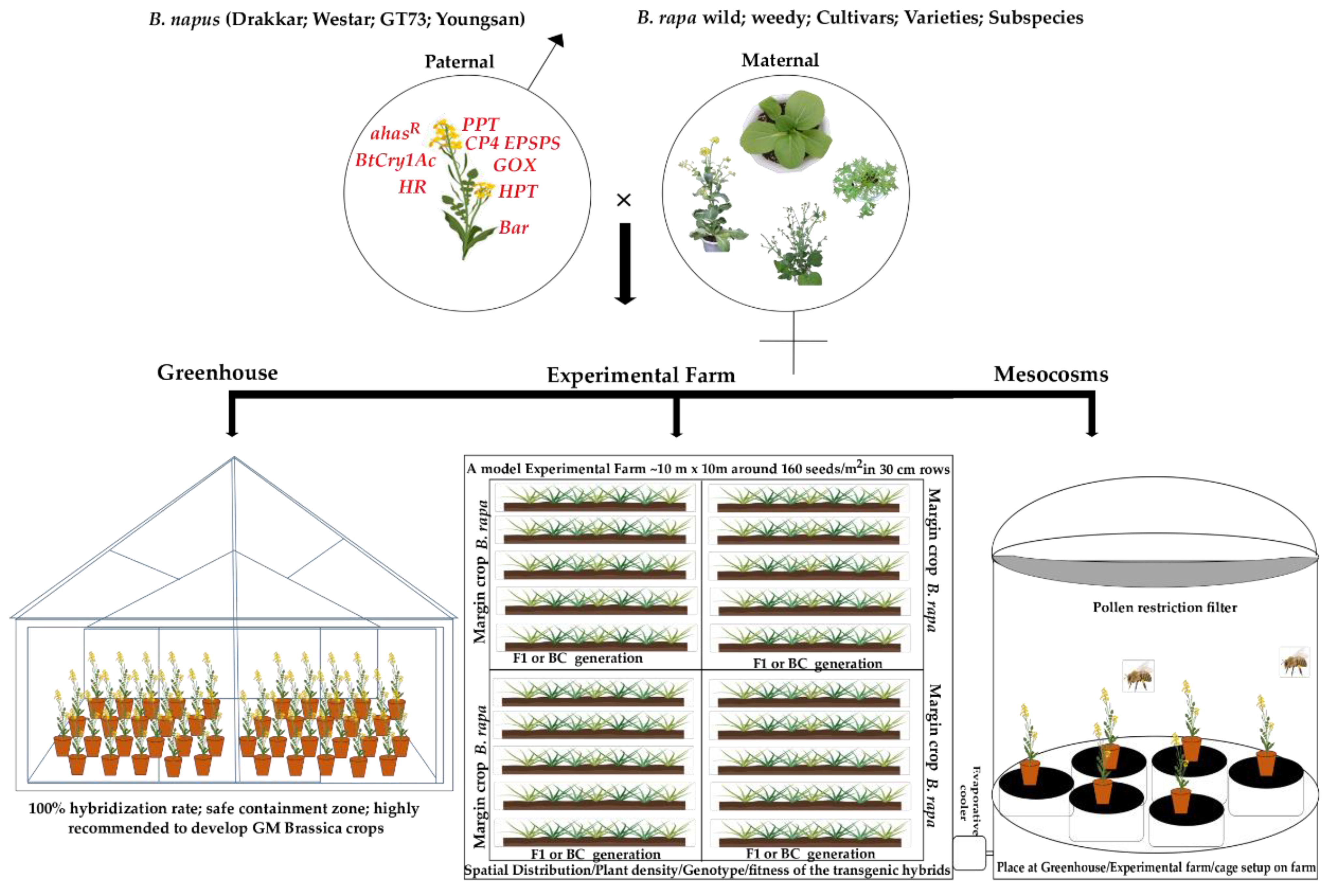
Figure 1.
An overview of interspecific hybridization between B. rapa × GM B. napus through various conditions.
Figure 1.
An overview of interspecific hybridization between B. rapa × GM B. napus through various conditions.


{kind=link}
Table 1.
List of studies on interspecific hybridization with B. rapa × GM B. napus.
| Countries | Hybridization | Variety/Cultivar | Transgenic Traits | Growing Conditions | Pollination/Mediated | References |
|---|---|---|---|---|---|---|
| France, USA | B. rapa × B. napus F1 hybrids, BC1 | B. napus (Westar); B. rapa (weedy; Back Bay, near Irvine, California) | Bt-transgenic (Btcry1AC, green fluroscent protein (GFP) mGFP5er gene) | Glasshouse | Hand pollination and bumble bee | [28] |
| Taiwan | B. rapa × B. napus F1 hybrids | B. rapa var. (Nongxing, Wansheng rape and Edible rape); B. napus (Var. Deza oil no. 18, Gueiza No. 4, Zhong oil No. 36, Wan oil No. 25, cultivar FTHEB1001) | Synthetic GM B. napus | Greenhouse | Manual pollination | [29] |
| USA | B. rapa × B. napus F1, BC1 and F2 individuals | B. napus (Westar GT1-9); NT: B. napus (cv. Westar); B. rapa acc.2974) | Btcry1Ac (GFP mGFP5er gene) and mitigation gene (pPZP212-ahasR-Δgai-1) confers ALS (acetolactate synthase) | Interspecific and intraspecific, competition conditions, greenhouse and shade house | Hand crossed | [30] |
| South Korea | B. rapa × B. napus F1 hybrids | B. napus L. (Youngsan), B. rapa L. ssp. pekinensis ‘Jangkang’ | Herbicide resistance and hygromycin resistance gene, pPBrAGL20 and pHBrAGL20 | GMO greenhouse | Artificial emasculation | [4] |
| China | B. rapa × B. napus c.v.GT73 F1 hybrids | B. napus c.v. GT73, B. napus c.v.Ms8x Rf3, B. napus c.v. Zhongyou 821 (CK, control); B. rapa L. ssp pekinensis Olsson (60), chinensis var. chinesis Kitam (33), chinensis var. purpurea Mao (4), chinensis var. parachinensis Tsen et Lee (10), chinensis var. rosularis Tsen et Lee (6), chinensis var. oleifera (3), raifera Matzg (2) | Glyphosate tolerant, Phosphinothricin tolerant | Greenhouse | Artificial emasculation and Spontaneous outcrossing | [31] |
| USA | B. rapa × B. napus F1 and BC1 F1, BC2 F2 | B. rapa wild accession, B. napus cv. Darkkar GT 1-9 | Btcry1Ac, GFP | Greenhouse | Houseflies | [32] |
| Israel | B. rapa × B. napus, F1 and F1BC1 | T1B.napus L. cv. Westar (16 transgene mitigating (TM) lines) and J7, J9 and J16 independent lines, B. rapa (#2974) | pPZP212-ahasR-Δgai-1 (TM 1), herbicide resistance | Glasshouse | Manual pollination | [33] |
| Canada | B. rapa × B. napus GT lines F1 and BC1-4 | B. napus c.v. Westar (GT1-9); B. rapa 2974 and 2975, CA | Bt-transgenic (Btcry1AC, GFP mGFP5er gene) | Growth chamber | Manually emasculated | [10] |
| Denmark | B. rapa × B. napus F1 hybrids and BC1-3 | B. napus Drakkar 93B1104, B. rapa BC25 (wild population, Denmark) | Glufosinate resistance, neo genes | Growth rooms | Bumblebees semi natural | [34] |
| USA | B. rapa × B. napus | seven T3 B. napus L. cv. Oscars 48,52,96,124, Westar:45,58,63; B. rapa weedy (CA), (MT) | BrCry1Ac | Growth chamber | Hand crossed | [9] |
| USA | B. rapa × B. napus F1 hybrids, BC1 | T1 B. napus cv. Westar (GT1-9) | BtCry1Ac, mGFP5er gene | Controlled condition | Agrobacterium | [11] |
| Netherlands | B. rapa × B. napus F1, BC1-4 | B. rapa chinensis, B. rapa pekinensis, B. napus cv. Drakkar | PPT, bar | Pollen cage at greenhouse | Emasculated | [35] |
| USA | B. rapa × B. napus F1 hybrids | B. napus RaideRR GT73, weedy B. rapa USDA-GRIN (PI 549154) | CP4 EPSPS, GOX and Cry1Ac | Glasshouse and outdoor mesocosms | Houseflies | [36] |
| USA | B. rapa × B. napus F1 and BC1 | B. napus RaideRR GT73, weedy B. rapa USDA-GRIN (PI 633155) | CP4 EPSPS, GOX | Outdoor mesocosms | manual pollination | [37] |
| Denmark | B. rapa × B. napus F1, BC1, | B. napus ssp. oleifera (DC) var. Darkkar, NMS1, NMS1 and RF1 | bar (barnase and barstar) encoding PAT resistant to PPT | Growth chamber to conviron growth cabinet to field | Random pollination | [26] |
| USA | B. rapa × B. napus F1, Bc1 | T0 B. napus cv. Westar (GT2-4, GT8-9 and GFP1-3), B. rapa (wild relatives Br CA, Br QC-2974, Br QC-2975) | BtCry1Ac, mGFP5er gene | Field to greenhouse | spontaneous | [38] |
| Russia | B. rapa × B. napus, BC | NT B. napus cv. Ratnik, Belinda, Heros | Hygromycin phosphotransferase (HPT) | Field to greenhouse | spontaneous | [39] |
| France, USA | B. rapa × F1 hybrids, | B. napus ssp. oleifera, B. rapa (weed) | Bt-transgenic (Btcry1AC, GFP mGFP5er gene) | Greenhouse with microcosms | spontaneous | [25] |
| Canada | B. rapa × B. napus F1 hybrids | B. napus (HR 45A51, 45A50 and Westar, GT-2,6,7,8 and 9 with GP); B. rapa wild | BtCry1Ac, mGFP5er gene, HR, | Field experimental farm | spontaneous | [11] |
| Canada | B. rapa × B. napus field border plants | B. rapa QC-9039, QC-9047; B. napus (Glyphosate resistant) | Glyphosate resistant | Field experimental farm with two different sites | spontaneous | [40] |
| Denmark | B. rapa × B. napus, BC1-2 | B. napus (Basta), B. rapa (weedy) | Basta herbicide tolerance | Field | spontaneous | [8] |
| Canada | B. rapa × B. napus F1 and BC1 | B. napus 45A51 HR glyphosate resistant (volunteer of CP4 EPSPS), B. rapa (QC-9039, QC-9047) | HR glyphosate | Commercial field | spontaneous | [41] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sohn, S.-I.; Thamilarasan, S.K.; Pandian, S.; Oh, Y.-J.; Ryu, T.-H.; Lee, G.-S.; Shin, E.-K. Interspecific Hybridization of Transgenic Brassica napus and Brassica rapa—An Overview. Genes 2022, 13, 1442. https://doi.org/10.3390/genes13081442
AMA Style
Sohn S-I, Thamilarasan SK, Pandian S, Oh Y-J, Ryu T-H, Lee G-S, Shin E-K. Interspecific Hybridization of Transgenic Brassica napus and Brassica rapa—An Overview. Genes. 2022; 13(8):1442. https://doi.org/10.3390/genes13081442
Chicago/Turabian StyleSohn, Soo-In, Senthil Kumar Thamilarasan, Subramani Pandian, Young-Ju Oh, Tae-Hun Ryu, Gang-Seob Lee, and Eun-Kyoung Shin. 2022. "Interspecific Hybridization of Transgenic Brassica napus and Brassica rapa—An Overview" Genes 13, no. 8: 1442. https://doi.org/10.3390/genes13081442
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.