Polyamine Oxidase Is Involved in Spermidine Reduction of Transglutaminase Type 2-Catalyzed βH-Crystallins Polymerization in Calcium-Induced Experimental Cataract

 ,
,  , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Lens Culturing
2.3. Separation of Rabbit Lens Crystallins by Gel Permeation and SDS-PAGE
2.4. Incorporation of [3H]-SPD into Lens Crystallins from Gel-Permeation by Purified GPL-TG2
2.5. RP-HPLC Chromatography of βH-Crystallins
2.6. N-terminal Amino Acids Sequencing of βH-Crystallins
2.7. Determination of Endogenous Free SPD and Its (γ-Glutamyl) Derivatives
2.8. FAD-PAO Activity Assay
2.9. Transglutaminase Activity Assay
2.10. Lens Transparency Evaluation
2.11. Statistical Analysis
3. Results
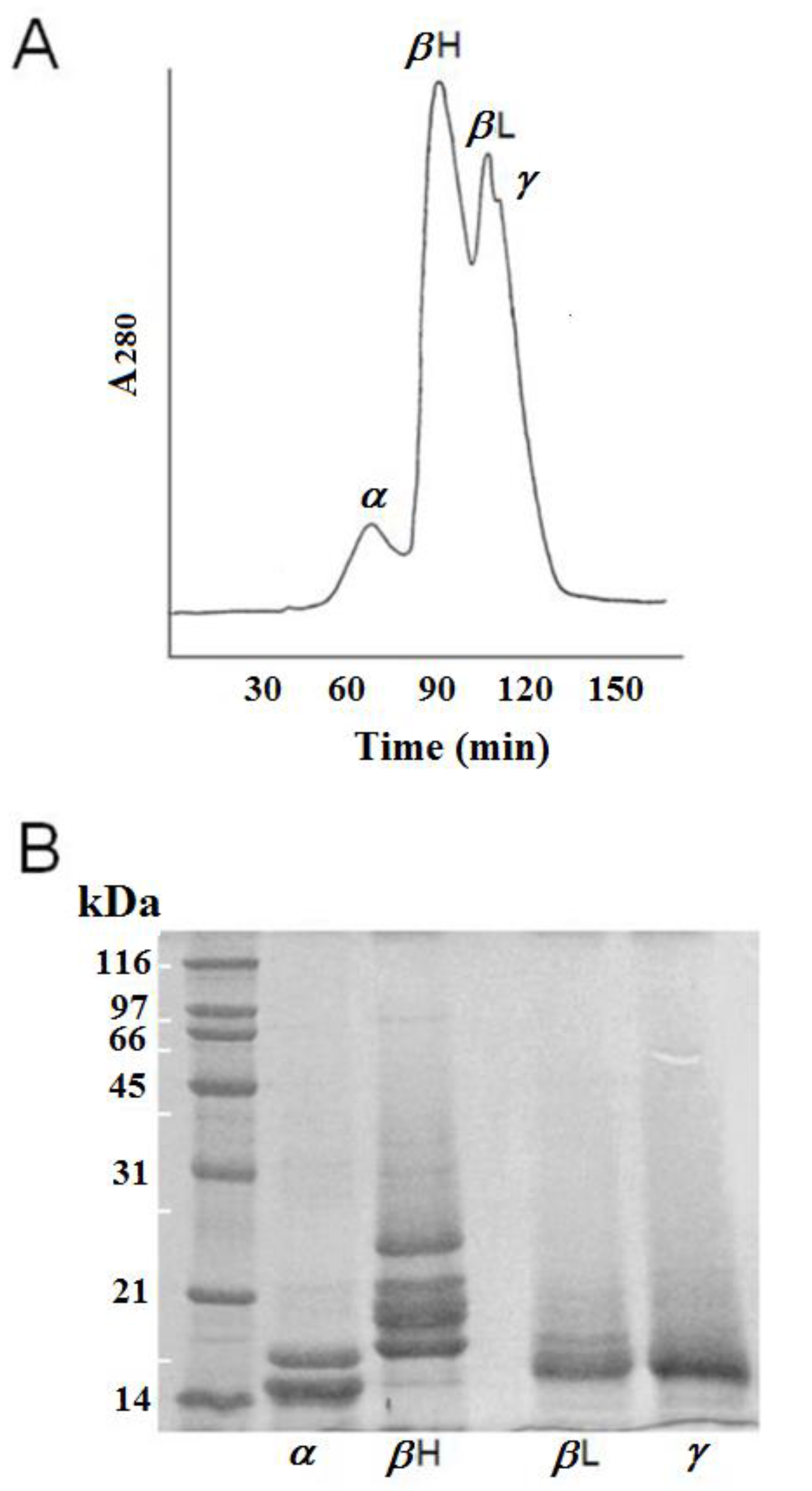
3.1. Separation of Crystallins by Gel-Filtration and Identification by SDS-PAGE Analysis
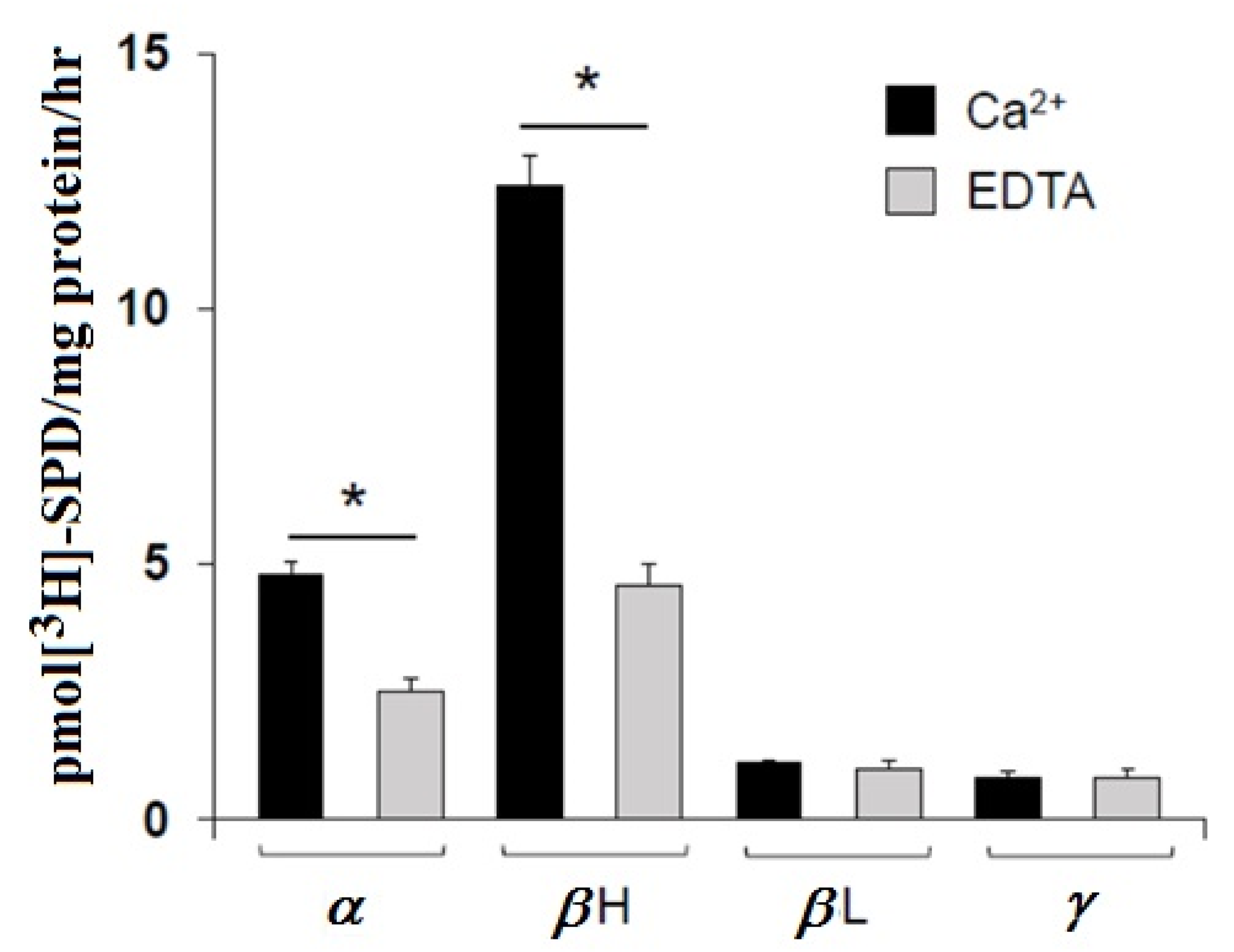
3.2. Transglutaminase-Catalyzed Incorporation of Polyamines into Crystallins
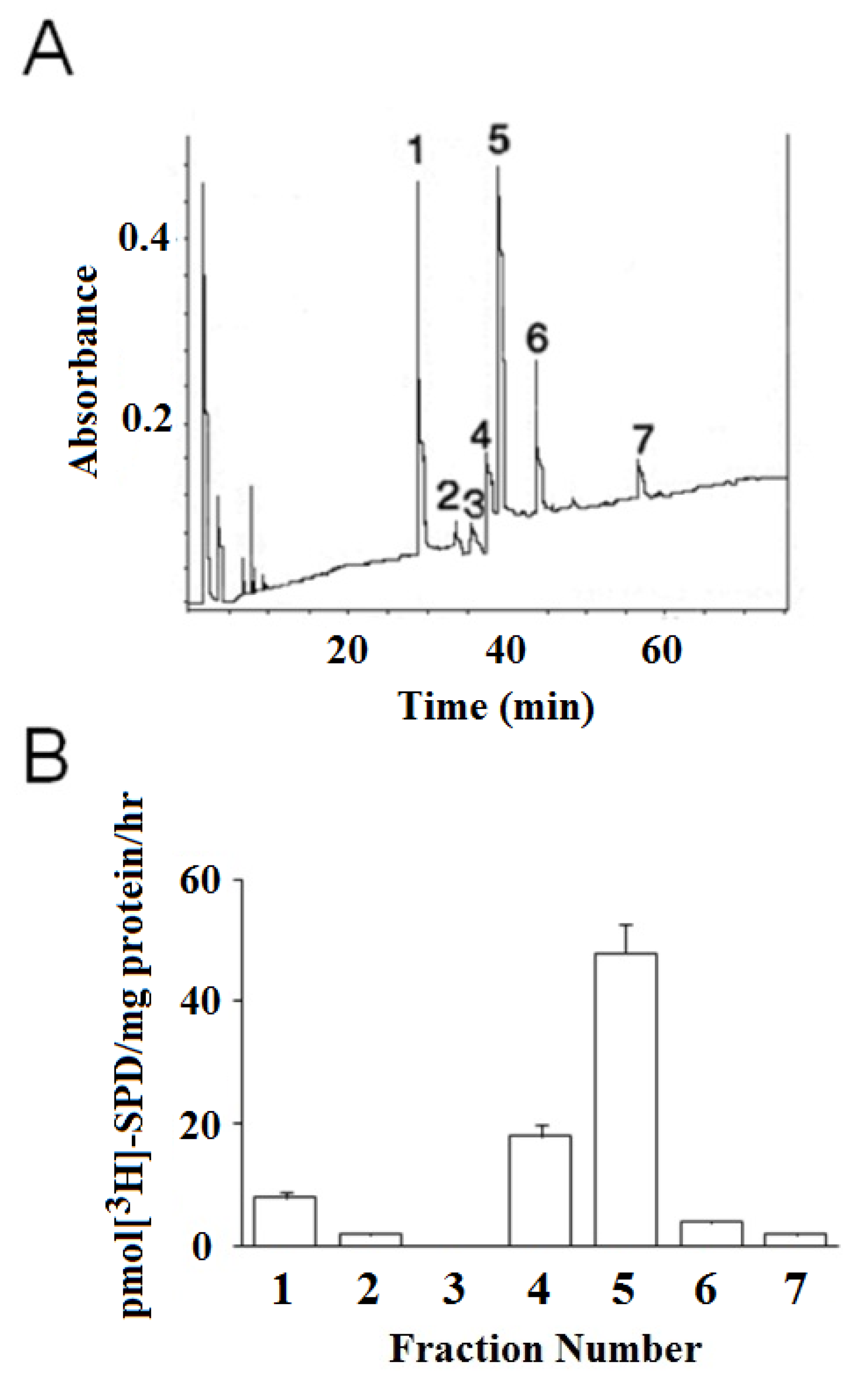
3.3. HPLC Separation of βH-Crystallins and Identification of TG2 Substrates
3.4. Levels of the TG2 Catalyzed, SPD Derivatives, Linked to the Crystallins of Cultured Rabbit Lens, in Presence of Exogenous SPD
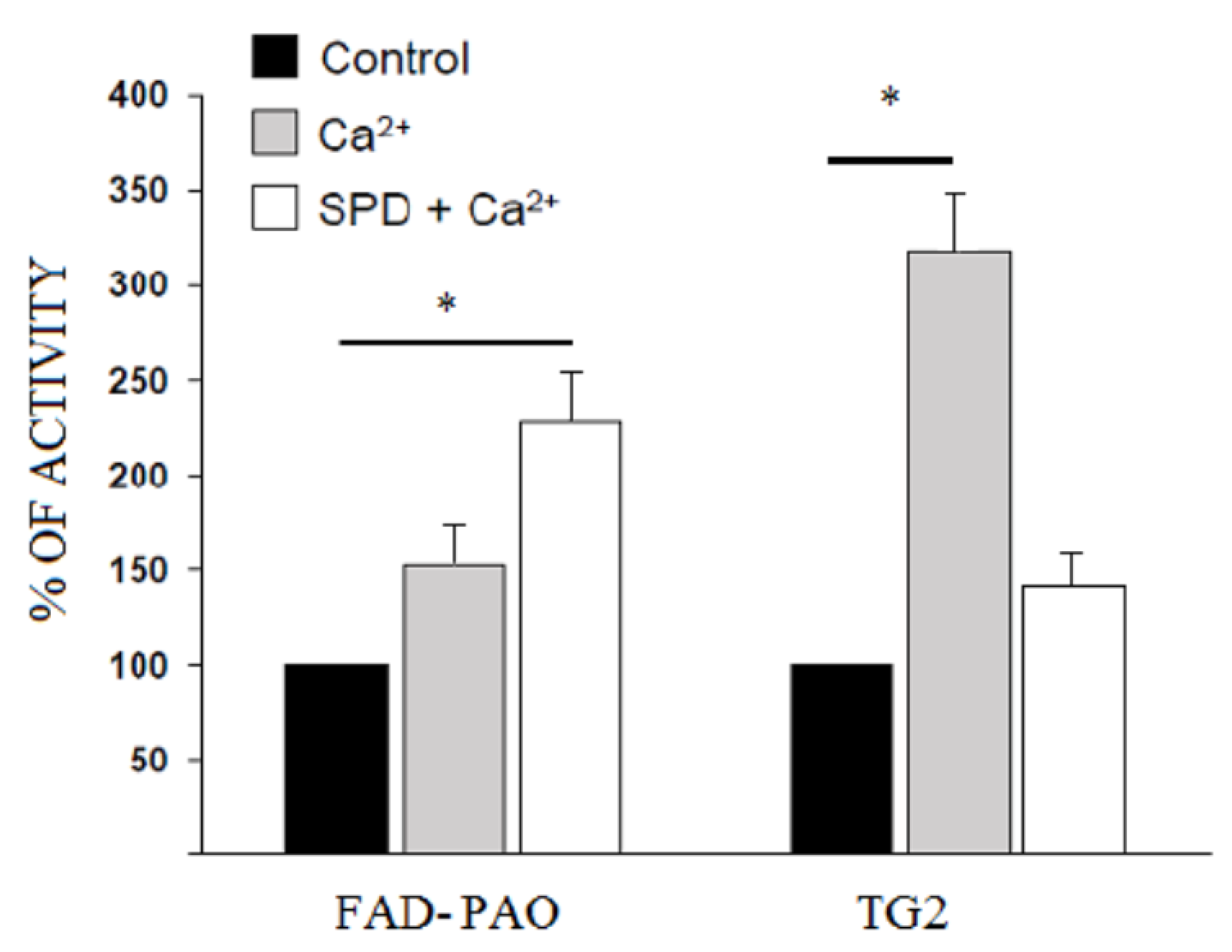
3.5. FAD-PAO and TG2 Activity in Rabbit Lens
3.6. Levels of the TG2-Catalyzed SPD Derivatives, in the Proteolytic Digest of Cultured Rabbit Lens upon PAO Activity Inhibition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wistow, G. The human crystallin gene families. Hum. Genom. 2012, 6, 26. [Google Scholar] [CrossRef] [Green Version]
- Harding, J.J.; Crabbe, M.J.C. The lens: Development, proteins, metabolism and cataract. Eye 1984, 1B, 207–492. [Google Scholar]
- Haslbeck, M.; Peschek, J.; Buchner, J.; Weinkauf, S. Structure and function of α-crystallins: Traversing from in vitro to in vivo. Biochim. Biophys. Acta 2016, 1860, 149–166. [Google Scholar] [CrossRef] [PubMed]
- Bloemendal, H.; de Jong, W.; Jaenicke, R.; Lubsen, N.H.; Slingsby, C.; Tardieu, A. Ageing and vision: Structure, stability and function of lens crystallins. Prog. Biophys. Mol. Biol. 2004, 86, 407–485. [Google Scholar] [CrossRef] [PubMed]
- Ray, N.J. Biophysical chemistry of the ageing eye lens. Biophys. Rev. 2015, 7, 353–368. [Google Scholar] [CrossRef]
- Zhang, K.; Zhu, X.; Lu, Y. The Proteome of Cataract Markers: Focus on Crystallins. Adv. Clin. Chem. 2018, 86, 179–210. [Google Scholar]
- Lorand, L.; Graham, R.M. Transglutaminases: Crosslinking enzymes with pleiotropic functions. Nat. Rev. Mol. Cell. Biol. 2003, 4, 140–156. [Google Scholar] [CrossRef]
- Boros, S.; Wilmarth, P.A.; Kamps, B.; de Jong, W.W.; Bloemendal, H.; Lampi, K.; Boelens, W.C. Tissue transglutaminase catalyzes the deamidation of glutamines in lens betaB(2)- and betaB(3)-crystallins. Exp. Eye Res. 2008, 86, 383–393. [Google Scholar] [CrossRef]
- Tabolacci, C.; Lentini, A.; Provenzano, B.; Beninati, S. Evidences for a role of protein cross-links in transglutaminase-related disease. Amino Acids 2012, 42, 975–986. [Google Scholar] [CrossRef]
- Tabolacci, C.; De Martino, A.; Mischiati, C.; Feriotto, G.; Beninati, S. The Role of Tissue Transglutaminase in Cancer Cell Initiation, Survival and Progression. Med. Sci. 2019, 7, 19. [Google Scholar] [CrossRef] [Green Version]
- Zhuang, R.; Khosla, C. Substrates, inhibitors, and probes of mammalian transglutaminase. Anal. Biochem. 2020, 591, 113560. [Google Scholar] [CrossRef] [PubMed]
- Lai, T.S.; Lin, C.J.; Greenberg, C.S. Role of tissue transglutaminase-2 (TG2)-mediated aminylation in biological processes. Amino Acids 2017, 49, 501–515. [Google Scholar] [CrossRef]
- Folk, J.E.; Park, M.H.; Chung, S.I.; Schrode, J.; Lester, E.P.; Cooper, H.L. Polyamines as physiological substrates for transglutaminases. J. Biol. Chem. 1980, 255, 3695–3700. [Google Scholar] [PubMed]
- Lentini, A.; Tabolacci, C.; Mattioli, P.; Provenzano, B.; Beninati, S. Spermidine delays eye lens opacification in vitro by suppressing transglutaminase-catalyzed crystallin cross-linking. Protein J. 2011, 30, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Lentini, A.; Mattioli, P.; Provenzano, B.; Abbruzzese, A.; Caraglia, M.; Beninati, S. Role of the FAD-dependent polyamine oxidase in the selective formation of N(1),N(8)-bis(gamma-glutamyl)spermidine protein cross-links. Biochem. Soc. Trans. 2007, 35, 396–400. [Google Scholar] [CrossRef] [PubMed]
- Provenzano, B.; Lentini, A.; Tatti, R.; De Martino, A.; Borromeo, I.; Mischiati, C.; Feriotto, G.; Forni, C.; Tabolacci, C.; Beninati, S. Evaluation of polyamines as marker of melanoma cell proliferation and differentiation by an improved high-performance liquid chromatographic method. Amino Acids 2019, 51, 1623–1631. [Google Scholar] [CrossRef]
- Beninati, S.; Martinet, N.; Folk, J.E. High-performance liquid chromatographic method for the determination of epsilon-(gamma-glutamyl)lysine and mono- and bis-gamma-glutamyl derivatives of putrescine and spermidine. J. Chromatogr. 1988, 443, 329–335. [Google Scholar] [CrossRef]
- Bachrach, U.; Reches, B. Enzymic assay for spermine and spermidine. Anal. Biochem. 1966, 17, 38–48. [Google Scholar] [CrossRef]
- Quash, G.; Keolouangkhot, T.; Gazzolo, L.; Ripoll, H.; Saez, S. Diamine oxidase and polyamine oxidase activities in normal and transformed cells. Biochem. J. 1979, 177, 275–282. [Google Scholar] [CrossRef] [Green Version]
- Jain, A.K.; Lim, G.; Langford, M.; Jain, S.K. Effect of high-glucose levels on protein oxidation in cultured lens cells, and in crystalline and albumin solution and its inhibition by vitamin B6 and N-acetylcysteine: Its possible relevance to cataract formation in diabetes. Free Radic. Biol. Med. 2002, 33, 1615–1621. [Google Scholar] [CrossRef]
- Chiou, S.H.; Chen, S.W.; Lo, T.B. The amino-terminal sequences of four major carp gamma-crystallin polypeptides and their homology with frog and calf gamma-crystallins. FEBS Lett. 1986, 209, 107–110. [Google Scholar] [CrossRef] [Green Version]
- Bey, P.; Bolkenius, F.N.; Seiler, N.; Casara, P. N-2,3-butadienyl-1,4-butanediamine derivatives: Potent irreversible inactivators of mammalian polyamine oxidase. J. Med. Chem. 1985, 28, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Cao, H.; Liu, G.; Heinzmann, D.; Chen, H.; Umbach, A.T.; Gawaz, M.; Lang, F. Effect of Lysosomotropic Polyamineoxidase Inhibitor MDL-72527 on Platelet Activation. Cell Physiol. Biochem. 2016, 38, 1695–1702. [Google Scholar] [CrossRef]
- Ghosh, K.S.; Chauhan, P. Crystallins and Their Complexes. Subcell. Biochem. 2019, 93, 439–460. [Google Scholar]
- Periyasamy, P.; Shinohara, T. Age-related cataracts: Role of unfolded protein response, Ca2+ mobilization, epigenetic DNA modifications, and loss of Nrf2/Keap1 dependent cytoprotection. Prog. Retin. Eye Res. 2017, 60, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Lorand, L.; Hsu, L.K.; Siefring, G.E., Jr.; Rafferty, N.S. Lens transglutaminase and cataract formation. Proc. Natl. Acad. Sci. USA 1981, 78, 1356–1360. [Google Scholar] [CrossRef] [Green Version]
- Velasco, P.T.; Lorand, L. Acceptor-donor relationships in the transglutaminase-mediated cross-linking of lens beta-crystallin subunits. Biochemistry 1987, 26, 4629–4634. [Google Scholar] [CrossRef]
- Beninati, S.; Piacentini, M.; Bergamini, C.M. Transglutaminase 2, a double face enzyme. Amino Acids 2017, 49, 415–423. [Google Scholar] [CrossRef]
- Igarashi, K.; Kashiwagi, K. The functional role of polyamines in eukaryotic cells. Int. J. Biochem. Cell Biol. 2019, 107, 104–115. [Google Scholar] [CrossRef]
- Folk, J.E. Transglutaminases. Ann. Rev. Biochem. 1980, 49, 517–531. [Google Scholar] [CrossRef]
- Sanderson, J.; Marcantonio, J.M.; Duncan, G.A. Human lens model of cortical cataract: Ca2+-induced protein loss, vimentin cleavage and opacification. Investig. Ophthalmol. Vis. Sci. 2000, 41, 2255–2261. [Google Scholar]
- Seiler, N.; Duranton, B.; Raul, F. The polyamine oxidase inactivator MDL. Prog. Drug. Res. 2002, 59, 1–40. [Google Scholar] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (γ-Glutamyl)SPD Derivatives (pmol/mg of Lens Proteins) | ||||||
|---|---|---|---|---|---|---|
| Time (h) | 72 | 96 | 120 | |||
| Protein bound derivative | −SPD | +SPD | −SPD | +SPD | −SPD | +SPD |
| N8-mono(γ-glutamyl)SPD | 12.3 ± 0.5 | 37.8 ± 2.5 | 10.2 ± 3.5 | 68.2 ± 4.3 | 6.7 ± 0.2 | 87.7 ± 5.7 |
| N1-mono(γ-glutamyl)SPD | 50.3 ± 3.5 | 28.7 ± 1.5 | 36.4 ± 8.7 | 30.7 ± 5.4 | 22.6 ± 9.2 | 15.3 ± 0.7 |
| N1,N8-mono(γ-glutamyl)SPD | 65.7 ± 8.4 | 50.5 ± 2.5 | 96.4 ± 13.0 | 37.2 ± 6.0 | 125.4 ± 14.9 | 17.4 ± 0.4 |
| Lens transparency % | 40 | 50 | 30 | 60 | 10 | 80 |
| (γ-Glutamyl)SPD Derivatives (pmol/mg of Lens Proteins) | ||||||
|---|---|---|---|---|---|---|
| Time (h) | 72 | 96 | 120 | |||
| Protein bound derivative | CTRL | MDL 72527 | CTRL | MDL 72527 | CTRL | MDL 72527 |
| N8-mono(γ-glutamyl)SPD | 38.3 ± 7.4 | 36.5 ± 8.3 | 78.2 ± 3.6 | 67.2 ± 3.5 | 98.7 ± 13.8 | 92.7 ± 14.8 |
| N1-mono(γ-glutamyl)SPD | 25.5 ± 4.5 | 65.3 ± 6.8 | 20.8 ± 3.6 | 78.2 ± 5.4 | 14.2 ± 0.4 | 88.7 ± 3.2 |
| N1,N8-mono(γ-glutamyl)SPD | 46.4 ± 6.8 | 76.0 ± 4.7 | 29.2 ± 2.8 | 85.3 ± 4.9 | 15.2 ± 0.6 | 7.6 ± 0.9 |
| Lens transparency % | 50 | 35 | 60 | 30 | 80 | 90 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mischiati, C.; Feriotto, G.; Tabolacci, C.; Domenici, F.; Melino, S.; Borromeo, I.; Forni, C.; De Martino, A.; Beninati, S. Polyamine Oxidase Is Involved in Spermidine Reduction of Transglutaminase Type 2-Catalyzed βH-Crystallins Polymerization in Calcium-Induced Experimental Cataract. Int. J. Mol. Sci. 2020, 21, 5427. https://doi.org/10.3390/ijms21155427
Mischiati C, Feriotto G, Tabolacci C, Domenici F, Melino S, Borromeo I, Forni C, De Martino A, Beninati S. Polyamine Oxidase Is Involved in Spermidine Reduction of Transglutaminase Type 2-Catalyzed βH-Crystallins Polymerization in Calcium-Induced Experimental Cataract. International Journal of Molecular Sciences. 2020; 21(15):5427. https://doi.org/10.3390/ijms21155427
Chicago/Turabian StyleMischiati, Carlo, Giordana Feriotto, Claudio Tabolacci, Fabio Domenici, Sonia Melino, Ilaria Borromeo, Cinzia Forni, Angelo De Martino, and Simone Beninati. 2020. "Polyamine Oxidase Is Involved in Spermidine Reduction of Transglutaminase Type 2-Catalyzed βH-Crystallins Polymerization in Calcium-Induced Experimental Cataract" International Journal of Molecular Sciences 21, no. 15: 5427. https://doi.org/10.3390/ijms21155427