Blocking IL-17: A Promising Strategy in the Treatment of Systemic Rheumatic Diseases

 , and
, and
Abstract
:
1. Introduction
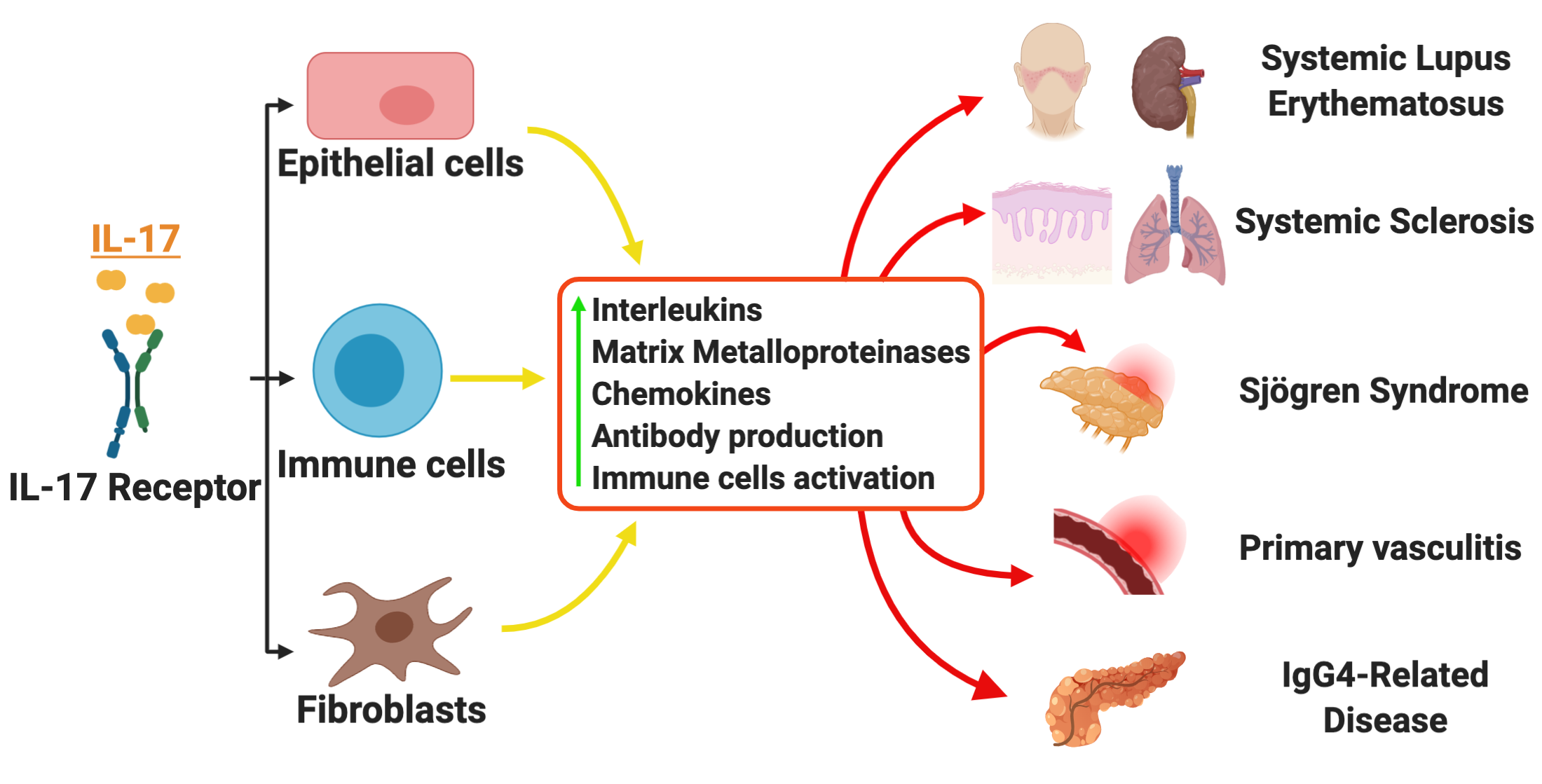
2. IL-17 Expression and Function
3. IL-17 in Systemic Rheumatic Diseases
3.1. Systemic Lupus Erythematosus
3.2. Systemic Sclerosis
3.3. Sjögren Syndrome
3.4. Other Systemic Rheumatic Diseases
4. IL-17 Inhibitors in Systemic Rheumatic Diseases
- An active and recruiting phase III trial of Secukinumab for lupus nephritis (Clinicaltrials.gov identifier: NCT04181762) a 2-year, phase III randomized, double-blind, parallel-group, placebo-controlled trial to evaluate the efficacy and safety of secukinumab in combination with the standard of care therapy in patients with active lupus nephritis. The primary endpoint is the proportion of subjects achieving complete renal response (CRR) [125];
- An active but not recruiting phase II trial of secukinumab for giant cell arteritis (Clinicaltrials.gov identifier: NCT03765788) randomized, parallel-group, double-blind, placebo-controlled, multicenter, Phase II study to evaluate the efficacy and safety of secukinumab to maintain disease remission up to 28 weeks including corticosteroid tapering, in patients with newly diagnosed or relapsing giant cell arteritis (GCA) who are naïve to biological therapy [125].
- An active but not recruiting phase III clinical trial (Clinicaltrials.gov identifier: NCT03957681) of brodalumab for moderate to severe systemic sclerosis: placebo-controlled, double-blind comparative study of brodalumab with an open-label extension period in subjects with systemic sclerosis who have moderate to severe skin thickening. The primary outcome measures the change in modified Rodnan skin score (mRSS) from baseline at week 24 [125];
- An active but not recruiting phase I clinical trial (Clinicaltrials.gov identifier: NCT04368403), open-label, multiple-dose study of brodalumab in subjects with systemic sclerosis. The primary outcome measures the serum concentration of brodalumab [125].
5. Concluding Remarks and Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Quintero-Ronderos, P.; Montoya-Ortiz, G. Epigenetics and Autoimmune Diseases. Autoimmune Dis. 2012, 2012, 593720. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, F.-S.; Gershwin, M.E. Human autoimmune diseases: A comprehensive update. J. Intern. Med. 2015, 278, 369–395. [Google Scholar] [CrossRef] [PubMed]
- Richmond, J.M.; Harris, J. The Pathobiologic Basis of Autoimmunity; Elsevier Inc.: Amsterdam, The Netherlands, 2014; pp. 39–48. [Google Scholar]
- Dinarello, C.A. Historical Review of Cytokines. Eur. J. Immunol. 2007, 37 (Suppl. 1), S34–S45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabarkiewicz, J.; Pogoda, K.; Karczmarczyk, A.; Pozarowski, P.; Giannopoulos, K. The Role of IL-17 and Th17 Lymphocytes in Autoimmune Diseases. Arch. Immunol. Ther. Exp. 2015, 63, 435–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, D.D.; Kuchroo, V.K. Th17 Cell Pathway in Human Immunity: Lessons from Genetics and Therapeutic Interventions. Immunity 2015, 43, 1040–1051. [Google Scholar] [CrossRef] [Green Version]
- Murdaca, G.; Colombo, B.M.; Puppo, F. The role of Th17 lymphocytes in the autoimmune and chronic inflammatory diseases. Intern. Emerg. Med. 2011, 6, 487–495. [Google Scholar] [CrossRef]
- Sieper, J.; Poddubnyy, D.; Miossec, P. The IL-23–IL-17 pathway as a therapeutic target in axial spondyloarthritis. Nat. Rev. Rheumatol. 2019, 15, 747–757. [Google Scholar] [CrossRef]
- Blauvelt, A.; Chiricozzi, A. The Immunologic Role of IL-17 in Psoriasis and Psoriatic Arthritis Pathogenesis. Clin. Rev. Allergy Immunol. 2018, 55, 379–390. [Google Scholar] [CrossRef] [Green Version]
- Naik, G.S.; Ming, W.-K.; Magodoro, I.M.; Akinwunmi, B.; Dar, S.; Poulsen, H.E.; Kristensen, L.E.; Ellervik, C. Th17 Inhibitors in Active Psoriatic Arthritis: A Systematic Review and Meta-Analysis of Randomized Controlled Clinical Trials. Dermatology 2017, 233, 366–377. [Google Scholar] [CrossRef]
- Burkett, P.R.; Kuchroo, V.K. IL-17 Blockade in Psoriasis. Cell 2016, 167, 1669. [Google Scholar] [CrossRef]
- Lubberts, E. The IL-23–IL-17 axis in inflammatory arthritis. Nat. Rev. Rheumatol. 2015, 11, 415–429. [Google Scholar] [CrossRef] [PubMed]
- Robert, M.; Miossec, P. IL-17 in Rheumatoid Arthritis and Precision Medicine: From Synovitis Expression to Circulating Bioactive Levels. Front. Med. 2019, 5, 364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rouvier, E.; Luciani, M.F.; Mattéi, M.G.; Denizot, F.; Golstein, P. CTLA-8, cloned from an activated T cell, bearing AU-rich messenger RNA instability sequences, and homologous to a herpesvirus saimiri gene. J. Immunol. 1993, 150, 5445–5456. [Google Scholar] [PubMed]
- Monin, L.; Gaffen, S. Interleukin 17 Family Cytokines: Signaling Mechanisms, Biological Activities, and Therapeutic Implications. Cold Spring Harb. Perspect. Boil. 2017, 10, a028522. [Google Scholar] [CrossRef] [PubMed]
- Cua, D.J.; Tato, C.M. Innate IL-17-producing cells: The sentinels of the immune system. Nat. Rev. Immunol. 2010, 10, 479–489. [Google Scholar] [CrossRef] [PubMed]
- McGeachy, M.J.; Cua, D.J.; Gaffen, S. The IL-17 Family of Cytokines in Health and Disease. Immunity 2019, 50, 892–906. [Google Scholar] [CrossRef] [PubMed]
- Harrington, L.E.; Hatton, R.D.; Mangan, P.R.; Turner, H.; Murphy, T.L.; Murphy, K.M.; Weaver, C.T. Interleukin 17-producing CD4+ effector T cells develop via a lineage distinct from the T helper type 1 and 2 lineages. Nat. Immunol. 2005, 6, 1123–1132. [Google Scholar] [CrossRef]
- Park, H.; Li, Z.; Yang, X.O.; Chang, S.H.; Nurieva, R.; Wang, Y.-H.; Wang, Y.; Hood, L.; Zhu, Z.; Tian, Q.; et al. A distinct lineage of CD4 T cells regulates tissue inflammation by producing interleukin 17. Nat. Immunol. 2005, 6, 1133–1141. [Google Scholar] [CrossRef]
- Tzartos, J.S.; Friese, M.A.; Craner, M.J.; Palace, J.; Newcombe, J.; Esiri, M.M.; Fugger, L. Interleukin-17 Production in Central Nervous System-Infiltrating T Cells and Glial Cells is Associated with Active Disease in Multiple Sclerosis. Am. J. Pathol. 2008, 172, 146–155. [Google Scholar] [CrossRef] [Green Version]
- Komiyama, Y.; Nakae, S.; Matsuki, T.; Nambu, A.; Ishigame, H.; Kakuta, S.; Sudo, K.; Iwakura, Y. IL-17 Plays an Important Role in the Development of Experimental Autoimmune Encephalomyelitis. J. Immunol. 2006, 177, 566–573. [Google Scholar] [CrossRef] [Green Version]
- Fujino, S.; Andoh, A.; Bamba, S.; Ogawa, A.; Hata, K.; Araki, Y.; Bamba, T.; Fujiyama, Y. Increased expression of interleukin 17 in inflammatory bowel disease. Gut 2003, 52, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Yen, D.; Cheung, J.; Scheerens, H.; Poulet, F.; McClanahan, T.; McKenzie, B.; Kleinschek, M.A.; Owyang, A.; Mattson, J.; Blumenschein, W.; et al. IL-23 is essential for T cell–mediated colitis and promotes inflammation via IL-17 and IL-6. J. Clin. Investig. 2006, 116, 1310–1316. [Google Scholar] [CrossRef] [PubMed]
- Radstake, T.R.; Van Bon, L.; Broen, J.; Hussiani, A.; Hesselstrand, R.; Wuttge, D.M.; Deng, Y.; Simms, R.; Lubberts, E.; Lafyatis, R. The Pronounced Th17 Profile in Systemic Sclerosis (SSc) Together with Intracellular Expression of TGFβ and IFNγ Distinguishes SSc Phenotypes. PLoS ONE 2009, 4, e5903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Yang, J.; Xing, X.; Wan, L.; Li, M. Increased frequency of Th17 cells in systemic sclerosis is related to disease activity and collagen overproduction. Arthritis Res. Ther. 2014, 16, R4. [Google Scholar] [CrossRef] [Green Version]
- Zaba, L.C.; Cardinale, I.; Gilleaudeau, P.; Sullivan-Whalen, M.; Suárez-Fariñas, M.; Fuentes-Duculan, J.; Novitskaya, I.; Khatcherian, A.; Bluth, M.J.; Lowes, M.A.; et al. Amelioration of epidermal hyperplasia by TNF inhibition is associated with reduced Th17 responses. J. Exp. Med. 2007, 204, 3183–3194. [Google Scholar] [CrossRef]
- Kyttaris, V.C.; Wang, Y.; Juang, Y.-T.; Weinstein, A.; Tsokos, G.C. Increased levels of NF-ATc2 differentially regulate CD154 and IL-2 genes in T cells from patients with systemic lupus erythematosus. J. Immunol. 2007, 178, 1960–1966. [Google Scholar] [CrossRef] [Green Version]
- Kotake, S.; Udagawa, N.; Takahashi, N.; Matsuzaki, K.; Itoh, K.; Ishiyama, S.; Saito, S.; Inoue, K.; Kamatani, N.; Gillespie, M.T.; et al. IL-17 in synovial fluids from patients with rheumatoid arthritis is a potent stimulator of osteoclastogenesis. J. Clin. Investig. 1999, 103, 1345–1352. [Google Scholar] [CrossRef]
- Zhang, L.; Li, Y.-G.; Li, Y.-H.; Qi, L.; Liu, X.; Yuan, C.-Z.; Hu, N.-W.; Ma, D.; Li, Z.-F.; Yang, Q.; et al. Increased Frequencies of Th22 Cells as well as Th17 Cells in the Peripheral Blood of Patients with Ankylosing Spondylitis and Rheumatoid Arthritis. PLoS ONE 2012, 7, e31000. [Google Scholar] [CrossRef]
- Shen, H.; Goodall, J.C.; Gaston, H. Frequency and phenotype of peripheral blood Th17 cells in ankylosing spondylitis and rheumatoid arthritis. Arthritis Rheum. 2009, 60, 1647–1656. [Google Scholar] [CrossRef]
- Jin, W.; Dong, C. IL-17 cytokines in immunity and inflammation. Emerg. Microbes Infect. 2013, 2, 1–5. [Google Scholar] [CrossRef]
- Strzępa, A.; Szczepanik, M. IL-17-expressing cells as a potential therapeutic target for treatment of immunological disorders. Pharmacol. Rep. 2011, 63, 30–44. [Google Scholar] [CrossRef]
- Zuniga, L.A.; Jain, R.; Haines, C.; Cua, D.J. Th17 cell development: From the cradle to the grave. Immunol. Rev. 2013, 252, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Korn, T.; Bettelli, E.; Gao, W.; Awasthi, A.; Jäger, A.; Strom, T.B.; Oukka, M.; Kuchroo, V.K. IL-21 initiates an alternative pathway to induce proinflammatory T(H)17 cells. Nature 2007, 448, 484–487. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Laurence, A.; Elias, K.M.; O’Shea, J.J. IL-21 is Produced by Th17 Cells and Drives IL-17 Production in a STAT3-dependent Manner. J. Boil. Chem. 2007, 282, 34605–34610. [Google Scholar] [CrossRef] [Green Version]
- Zielinski, C.E.; Mele, F.; Aschenbrenner, D.; Jarrossay, D.; Ronchi, F.; Gattorno, M.; Monticelli, S.; Lanzavecchia, A.; Sallusto, F. Pathogen-induced human TH17 cells produce IFN-γ or IL-10 and are regulated by IL-1β. Nature 2012, 484, 514–518. [Google Scholar] [CrossRef] [PubMed]
- Veldhoen, M.; Hocking, R.J.; Atkins, C.J.; Locksley, R.M.; Stockinger, B. TGFβ in the Context of an Inflammatory Cytokine Milieu Supports De Novo Differentiation of IL-17-Producing T Cells. Immunity 2006, 24, 179–189. [Google Scholar] [CrossRef] [Green Version]
- Bettelli, E.; Carrier, Y.; Gao, W.; Korn, T.; Strom, T.B.; Oukka, M.; Weiner, H.L.; Kuchroo, V.K. Reciprocal developmental pathways for the generation of pathogenic effector TH17 and regulatory T cells. Nature 2006, 441, 235–238. [Google Scholar] [CrossRef]
- Gaffen, S. Structure and signalling in the IL-17 receptor family. Nat. Rev. Immunol. 2009, 9, 556–567. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Bechara, R.; Zhao, J.; McGeachy, M.J.; Gaffen, S. IL-17 receptor–based signaling and implications for disease. Nat. Immunol. 2019, 20, 1594–1602. [Google Scholar] [CrossRef]
- Onishi, R.M.; Park, S.J.; Hanel, W.; Ho, A.W.; Maitra, A.; Gaffen, S.L. SEF/IL-17R (SEFIR) Is Not Enough. J. Boil. Chem. 2010, 285, 32751–32759. [Google Scholar] [CrossRef] [Green Version]
- Amatya, N.; Garg, A.V.; Gaffen, S. IL-17 Signaling: The Yin and the Yang. Trends Immunol. 2017, 38, 310–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zenobia, C.; Hajishengallis, G. Basic biology and role of interleukin-17 in immunity and inflammation. Periodontol. 2000 2015, 69, 142–159. [Google Scholar] [CrossRef] [PubMed]
- Herjan, T.; Hong, L.; Bubenik, J.; Bulek, K.; Qian, W.; Liu, C.; Li, X.; Chen, X.; Yang, H.; Ouyang, S.; et al. IL-17-receptor-associated adaptor Act1 directly stabilizes mRNAs to mediate IL-17 inflammatory signaling. Nat. Immunol. 2018, 19, 354–365. [Google Scholar] [CrossRef]
- Bulek, K.; Liu, C.; Swaidani, S.; Wang, L.; Page, R.C.; Gulen, M.F.; Herjan, T.; Abbadi, A.; Qian, W.; Sun, D.; et al. The inducible kinase IKKi is required for IL-17-dependent signaling associated with neutrophilia and pulmonary inflammation. Nat. Immunol. 2011, 12, 844–852. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Cao, X. Interleukin-17 and its expanding biological functions. Cell. Mol. Immunol. 2010, 7, 164–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korn, T.; Bettelli, E.; Oukka, M.; Kuchroo, V.K. IL-17 and Th17 Cells. Annu. Rev. Immunol. 2009, 27, 485–517. [Google Scholar] [CrossRef]
- Crispín, J.C.; Tsokos, G.C. IL-17 in Systemic Lupus Erythematosus. J. Biomed. Biotechnol. 2010, 2010, 943254. [Google Scholar] [CrossRef] [Green Version]
- Tsokos, G.C.; Lo, M.S.; Reis, P.C.; Sullivan, K.E. New insights into the immunopathogenesis of systemic lupus erythematosus. Nat. Rev. Rheumatol. 2016, 12, 716–730. [Google Scholar] [CrossRef]
- Ramos, P.S.; Brown, E.E.; Kimberly, R.P.; Langefeld, C.D. Genetic Factors Predisposing to Systemic Lupus Erythematosus and Lupus Nephritis. Semin. Nephrol. 2010, 30, 164–176. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Kyttaris, V.C.; Tsokos, G.C. The Role of IL-23/IL-17 Axis in Lupus Nephritis. J. Immunol. 2009, 183, 3160–3169. [Google Scholar] [CrossRef]
- Lee, S.-Y.; Lee, S.H.; Seo, H.-B.; Ryu, J.-G.; Jung, K.; Choi, J.W.; Jhun, J.; Park, J.-S.; Kwon, J.Y.; Kwok, S.-K.; et al. Inhibition of IL-17 ameliorates systemic lupus erythematosus in Roquinsan/san mice through regulating the balance of TFH cells, GC B cells, Treg and Breg. Sci. Rep. 2019, 9, 5227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henriques, A.; Inês, L.; Couto, M.; Pedreiro, S.; Santos, C.; Magalhaes, M.; Rodrigues-Santos, P.; Velada, I.; Almeida, A.; Carvalheiro, T.; et al. Frequency and functional activity of Th17, Tc17 and other T-cell subsets in Systemic Lupus Erythematosus. Cell. Immunol. 2010, 264, 97–103. [Google Scholar] [CrossRef]
- López, P.; Rodríguez-Carrio, J.; Caminal-Montero, L.; Mozo, L.; Suárez, A. A pathogenic IFNα, BLyS and IL-17 axis in Systemic Lupus Erythematosus patients. Sci. Rep. 2016, 6, 20651. [Google Scholar] [CrossRef] [PubMed]
- Saber, N.Z.; Maroof, S.H.; Soliman, D.; Fathi, M.S. Expression of T helper 17 cells and interleukin 17 in lupus nephritis patients. Egypt. Rheumatol. 2017, 39, 151–157. [Google Scholar] [CrossRef]
- Crispín, J.C.; Oukka, M.; Bayliss, G.; Cohen, R.A.; Van Beek, C.A.; Stillman, I.E.; Kyttaris, V.C.; Juang, Y.-T.; Tsokos, G.C.; Oukka, M. Expanded Double Negative T Cells in Patients with Systemic Lupus Erythematosus Produce IL-17 and Infiltrate the Kidneys1. J. Immunol. 2008, 181, 8761–8766. [Google Scholar] [CrossRef]
- Chen, X.Q.; Yu, Y.C.; Deng, H.; Sun, J.Z.; Dai, Z.; Wu, Y.W.; Yang, M. Plasma IL-17A is Increased in New-Onset SLE Patients and Associated with Disease Activity. J. Clin. Immunol. 2010, 30, 221–225. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.K.; Lit, L.C.W.; Tam, L.-S.; Li, E.K.M.; Wong, P.T.Y.; Lam, C.W.K. Hyperproduction of IL-23 and IL-17 in patients with systemic lupus erythematosus: Implications for Th17-mediated inflammation in auto-immunity. Clin. Immunol. 2008, 127, 385–393. [Google Scholar] [CrossRef]
- Galil, S.M.A.; Ezzeldin, N.; El-Boshy, M.E. The role of serum IL-17 and IL-6 as biomarkers of disease activity and predictors of remission in patients with lupus nephritis. Cytokine 2015, 76, 280–287. [Google Scholar] [CrossRef]
- Vincent, F.B.; Northcott, M.; Hoi, A.; Mackay, F.; Morand, E.F. Clinical associations of serum interleukin-17 in systemic lupus erythematosus. Arthritis Res. Ther. 2013, 15, R97. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Zhao, P.; Ma, L.; Shan, Y.; Jiang, Z.; Wang, J.; Jiang, Y. Increased Interleukin 21 and Follicular Helper T-like Cells and Reduced Interleukin 10+ B cells in Patients with New-onset Systemic Lupus Erythematosus. J. Rheumatol. 2014, 41, 1781–1792. [Google Scholar] [CrossRef]
- Lan, Y.; Luo, B.; Wang, J.-L.; Jiang, Y.-W.; Wei, Y.-S. The association of interleukin-21 polymorphisms with interleukin-21 serum levels and risk of systemic lupus erythematosus. Gene 2014, 538, 94–98. [Google Scholar] [CrossRef]
- Fuchs, T.A.; Abed, U.; Goosmann, C.; Hurwitz, R.; Schulze, I.; Wahn, V.; Weinrauch, Y.; Brinkmann, V.; Zychlinsky, A. Novel cell death program leads to neutrophil extracellular traps. J. Cell Boil. 2007, 176, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Villanueva, E.; Yalavarthi, S.; Berthier, C.C.; Hodgin, J.B.; Khandpur, R.; Lin, A.M.; Rubin, C.J.; Zhao, W.; Olsen, S.H.; Klinker, M.; et al. Netting neutrophils induce endothelial damage, infiltrate tissues, and expose immunostimulatory molecules in systemic lupus erythematosus. J. Immunol. 2011, 187, 538–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pisitkun, P.; Ha, H.-L.; Wang, H.; Claudio, E.; Tivy, C.C.; Zhou, H.; Mayadas, T.N.; Illei, G.G.; Siebenlist, U. Interleukin-17 cytokines are critical in development of fatal lupus glomerulonephritis. Immunity 2012, 37, 1104–1115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogura, H.; Murakami, M.; Okuyama, Y.; Tsuruoka, M.; Kitabayashi, C.; Kanamoto, M.; Nishihara, M.; Iwakura, Y.; Hirano, T. Interleukin-17 Promotes Autoimmunity by Triggering a Positive-Feedback Loop via Interleukin-6 Induction. Immunity 2008, 29, 628–636. [Google Scholar] [CrossRef] [Green Version]
- Doreau, A.; Belot, A.; Bastid, J.; Riche, B.; Trescol-Biémont, M.-C.; Ranchin, B.; Fabien, N.; Cochat, P.; Pouteil-Noble, C.; Trolliet, P.; et al. Interleukin 17 acts in synergy with B cell–activating factor to influence B cell biology and the pathophysiology of systemic lupus erythematosus. Nat. Immunol. 2009, 10, 778–785. [Google Scholar] [CrossRef] [PubMed]
- Van Caam, A.; Vonk, M.; Hoogen, F.V.D.; Van Lent, P.; Van Der Kraan, P. Unraveling SSc Pathophysiology; The Myofibroblast. Front. Immunol. 2018, 9, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Pattanaik, D.; Brown, M.; Postlethwaite, B.C.; Postlethwaite, A.E. Pathogenesis of Systemic Sclerosis. Front. Immunol. 2015, 6, 272. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Bao, J.; Yang, J. Genistein-triggered anticancer activity against liver cancer cell line HepG2 involves ROS generation, mitochondrial apoptosis, G2/M cell cycle arrest and inhibition of cell migration. Arch. Med. Sci. 2018, 15, 1001–1009. [Google Scholar] [CrossRef]
- Kurasawa, K.; Hirose, K.; Sano, H.; Endo, H.; Shinkai, H.; Nawata, Y.; Takabayashi, K.; Iwamoto, I. Increased interleukin-17 production in patients with systemic sclerosis. Arthritis Rheum. 2000, 43, 2455–2463. [Google Scholar] [CrossRef]
- Fenoglio, D.; Battaglia, F.; Parodi, A.; Stringara, S.; Negrini, S.; Panico, N.; Rizzi, M.; Kalli, F.; Conteduca, G.; Ghio, M.; et al. Alteration of Th17 and Treg cell subpopulations co-exist in patients affected with systemic sclerosis. Clin. Immunol. 2011, 139, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Reyna, T.S.; Furuzawa-Carballeda, J.; Cabiedes, J.; Fajardo-Hermosillo, L.D.; Martinez-Reyes, C.; Díaz-Zamudio, M.; Llorente, L. Th17 peripheral cells are increased in diffuse cutaneous systemic sclerosis compared with limited illness: A cross-sectional study. Rheumatol. Int. 2011, 32, 2653–2660. [Google Scholar] [CrossRef]
- Zhou, Y.; Hou, W.; Xu, K.; Han, D.; Jiang, C.; Mou, K.; Li, Y.; Meng, L.; Lu, S. The elevated expression of Th17-related cytokines and receptors is associated with skin lesion severity in early systemic sclerosis. Hum. Immunol. 2015, 76, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Truchetet, M.-E.; Brembilla, N.C.; Montanari, E.; Lonati, P.; Raschi, E.; Zeni, S.; Fontao, L.; Meroni, P.L.; Chizzolini, C. Interleukin-17A+ Cell Counts Are Increased in Systemic Sclerosis Skin and Their Number is Inversely Correlated With the Extent of Skin Involvement. Arthritis Rheum. 2013, 65, 1347–1356. [Google Scholar] [CrossRef] [PubMed]
- Chizzolini, C.; Dufour, A.M.; Brembilla, N.C. Is there a role for IL-17 in the pathogenesis of systemic sclerosis? Immunol. Lett. 2018, 195, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Park, M.-J.; Moon, S.-J.; Lee, E.-J.; Jung, K.-A.; Kim, E.-K.; Kim, D.-S.; Lee, J.-H.; Kwok, S.-K.; Min, J.-K.; Park, S.-H.; et al. IL-1-IL-17 Signaling Axis Contributes to Fibrosis and Inflammation in Two Different Murine Models of Systemic Sclerosis. Front. Immunol. 2018, 9, 1611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okamoto, Y.; Hasegawa, M.; Matsushita, T.; Hamaguchi, Y.; Le Huu, D.; Iwakura, Y.; Fujimoto, M.; Takehara, K. Potential roles of interleukin-17A in the development of skin fibrosis in mice. Arthritis Rheum. 2012, 64, 3726–3735. [Google Scholar] [CrossRef] [PubMed]
- Lei, L.; Zhao, C.; Qin, F.; He, Z.; Wang, X.; Zhong, X.-N. Th17 cells and IL-17 promote the skin and lung inflammation and fibrosis process in a bleomycin-induced murine model of systemic sclerosis. Clin. Exp. Rheumatol. 2016, 34, 14–22. [Google Scholar]
- Brembilla, N.C.; Montanari, E.; Truchetet, M.-E.; Raschi, E.; Meroni, P.L.; Chizzolini, C. Th17 cells favor inflammatory responses while inhibiting type I collagen deposition by dermal fibroblasts: Differential effects in healthy and systemic sclerosis fibroblasts. Arthritis Res. Ther. 2013, 15, R151. [Google Scholar] [CrossRef] [Green Version]
- Carvalheiro, T.; Affandi, A.J.; Malvar-Fernández, B.; Dullemond, I.; Cossu, M.; Ottria, A.; Mertens, J.S.; Giovannone, B.; Bonte-Mineur, F.; Kok, M.R.; et al. Induction of Inflammation and Fibrosis by Semaphorin 4A in Systemic Sclerosis. Arthritis Rheumatol. 2019, 71, 1711–1722. [Google Scholar] [CrossRef] [Green Version]
- Dufour, A.M.; Borowczyk, J.; Alvarez, M.; Truchetet, M.-E.; Modarressi, A.; Brembilla, N.C.; Chizzolini, C. IL-17A Dissociates Inflammation from Fibrogenesis in Systemic Sclerosis. J. Investig. Dermatol. 2019, 140, 103–112. [Google Scholar] [CrossRef] [Green Version]
- Xing, X.; Yang, J.; Yang, X.; Wei, Y.; Zhu, L.; Gao, D.; Li, M. IL-17A Induces Endothelial Inflammation in Systemic Sclerosis via the ERK Signaling Pathway. PLoS ONE 2013, 8, e85032. [Google Scholar] [CrossRef] [PubMed]
- Flament, T.; Bigot, A.; Chaigne, B.; Henique, H.; Diot, E.; Marchand-Adam, S. Pulmonary manifestations of Sjögren’s syndrome. Eur. Respir. Rev. 2016, 25, 110–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brito-Zerón, P.; Baldini, C.; Bootsma, H.; Bowman, S.J.; Jonsson, R.; Mariette, X.; Sivils, K.; Theander, E.; Tzioufas, A.; Ramos-Casals, M. Sjögren syndrome. Nat. Rev. Dis. Prim. 2016, 2, 16047. [Google Scholar] [CrossRef] [PubMed]
- Baimpa, E.; Dahabreh, I.J.; Voulgarelis, M.; Moutsopoulos, H.M. Hematologic Manifestations and Predictors of Lymphoma Development in Primary Sjögren Syndrome. Medicine 2009, 88, 284–293. [Google Scholar] [CrossRef]
- Sakai, A.; Sugawara, Y.; Kuroishi, T.; Sasano, T.; Sugawara, S. Identification of IL-18 and Th17 cells in salivary glands of patients with Sjögren’s syndrome, and amplification of IL-17-mediated secretion of inflammatory cytokines from salivary gland cells by IL-18. J. Immunol. 2008, 181, 2898–2906. [Google Scholar] [CrossRef]
- Alunno, A.; Bistoni, O.; Bartoloni, E.; Caterbi, S.; Bigerna, B.; Tabarrini, A.; Mannucci, R.; Falini, B.; Gerli, R. IL-17-producing CD4-CD8- T cells are expanded in the peripheral blood, infiltrate salivary glands and are resistant to corticosteroids in patients with primary Sjogren’s syndrome. Ann. Rheum. Dis. 2012, 72, 286–292. [Google Scholar] [CrossRef]
- Maehara, T.; Moriyama, M.; Hayashida, J.; Tanaka, A.; Shinozaki, S.; Kubo, Y.; Matsumura, K.; Nakamura, S. Selective localization of T helper subsets in labial salivary glands from primary Sjögren’s syndrome patients. Clin. Exp. Immunol. 2012, 169, 89–99. [Google Scholar] [CrossRef] [Green Version]
- Katsifis, G.E.; Rekka, S.; Moutsopoulos, N.M.; Pillemer, S.; Wahl, S.M. Systemic and Local Interleukin-17 and Linked Cytokines Associated with Sjögren’s Syndrome Immunopathogenesis. Am. J. Pathol. 2009, 175, 1167–1177. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.; Gao, C.; Chen, H.; Li, Y.; Jin, Y.; Qi, H. Analysis of Th17-associated cytokines and clinical correlations in patients with dry eye disease. PLoS ONE 2017, 12, e0173301. [Google Scholar] [CrossRef] [Green Version]
- Kang, M.H.; Kim, M.K.; Lee, H.J.; Lee, H.I.; Wee, W.R.; Lee, J.H. Interleukin-17 in Various Ocular Surface Inflammatory Diseases. J. Korean Med. Sci. 2011, 26, 938–944. [Google Scholar] [CrossRef] [PubMed]
- Miletić, M.; Stojanović, R.; Pajic, O.; Bugarski, D.; Mojsilović, S.; Čokić, V.; Milenković, P. Serum interleukin-17 & nitric oxide levels in patients with primary Sjögren’s syndrome. Indian J. Med. Res. 2012, 135, 513–519. [Google Scholar] [PubMed]
- Pollard, R.P.E.; Abdulahad, W.H.; Bootsma, H.; Meiners, P.M.; Spijkervet, F.K.; Huitema, M.G.; Burgerhof, J.G.M.; Vissink, A.; Kroese, F.G.M. Predominantly proinflammatory cytokines decrease after B cell depletion therapy in patients with primary Sjögren’s syndrome. Ann. Rheum. Dis. 2013, 72, 2048–2050. [Google Scholar] [CrossRef] [PubMed]
- Verstappen, G.M.; Meiners, P.M.; Corneth, O.B.J.; Visser, A.; Arends, S.; Abdulahad, W.H.; Hendriks, R.W.; Vissink, A.; Kroese, F.G.M.; Bootsma, H. Attenuation of Follicular Helper T Cell-Dependent B Cell Hyperactivity by Abatacept Treatment in Primary Sjögren’s Syndrome. Arthritis Rheumatol. 2017, 69, 1850–1861. [Google Scholar] [CrossRef]
- Lee, S.Y.; Han, S.J.; Nam, S.M.; Yoon, S.; Ahn, J.M.; Kim, T.-I.; Kim, E.K.; Seo, K.Y. Analysis of Tear Cytokines and Clinical Correlations in Sjögren Syndrome Dry Eye Patients and Non–Sjögren Syndrome Dry Eye Patients. Am. J. Ophthalmol. 2013, 156, 247–253. [Google Scholar] [CrossRef]
- Tan, X.; Sun, S.; Liu, Y.; Zhu, T.; Wang, K.; Ren, T.; Wu, Z.; Xu, H.; Zhu, L. Analysis of Th17-associated cytokines in tears of patients with dry eye syndrome. Eye 2014, 28, 608–613. [Google Scholar] [CrossRef]
- Mieliauskaite, D.; Dumalakienė, I.; Rugiene, R.; Mackiewicz, Z. Expression of IL-17, IL-23 and Their Receptors in Minor Salivary Glands of Patients with Primary Sjögren’s Syndrome. Clin. Dev. Immunol. 2012, 2012, 187258. [Google Scholar] [CrossRef]
- Turpie, B.; Yoshimura, T.; Gulati, A.; Rios, J.D.; Dartt, D.A.; Masli, S. Sjögren’s Syndrome-Like Ocular Surface Disease in Thrombospondin-1 Deficient Mice. Am. J. Pathol. 2009, 175, 1136–1147. [Google Scholar] [CrossRef] [Green Version]
- Voigt, A.; Esfandiary, L.; Wanchoo, A.; Glenton, P.; Donate, A.; Craft, W.F.; Craft, S.L.M.; Nguyen, C.Q. Sexual dimorphic function of IL-17 in salivary gland dysfunction of the C57BL/6.NOD-Aec1Aec2 model of Sjögren’s syndrome. Sci. Rep. 2016, 6, 38717. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, C.Q.; Hu, M.H.; Li, Y.; Stewart, C.; Peck, A.B. Salivary gland tissue expression of interleukin-23 and interleukin-17 in Sjögren’s syndrome: Findings in humans and mice. Arthritis Rheum. 2008, 58, 734–743. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Rui, K.; Deng, J.; Tian, J.; Wang, X.; Wang, S.; Ko, K.-H.; Jiao, Z.; Chan, V.S.-F.; Lau, C.S.; et al. Th17 cells play a critical role in the development of experimental Sjögren’s syndrome. Ann. Rheum. Dis. 2014, 74, 1302–1310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subbarayal, B.; Chauhan, S.K.; Di Zazzo, A.; Dana, R. IL-17 Augments B Cell Activation in Ocular Surface Autoimmunity. J. Immunol. 2016, 197, 3464–3470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verstappen, G.M.; Corneth, O.B.; Bootsma, H.; Kroese, F.G. Th17 cells in primary Sjögren’s syndrome: Pathogenicity and plasticity. J. Autoimmun. 2018, 87, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Hulkkonen, J.; Pertovaara, M.; Antonen, J.; Pasternack, A.; Hurme, M.; Pöllänen, P.; Lehtimäki, T. Matrix metalloproteinase 9 (MMP-9) gene polymorphism and MMP-9 plasma levels in primary Sjogren’s syndrome. Rheumatology 2004, 43, 1476–1479. [Google Scholar] [CrossRef] [Green Version]
- Pérez, P.; Kwon, Y.-J.; Alliende, C.; Leyton, L.; Aguilera, S.; Molina, C.; Labra, C.; Julio, M.; Leyton, C.; González, M.-J. Increased acinar damage of salivary glands of patients with Sjögren’s syndrome is paralleled by simultaneous imbalance of matrix metalloproteinase 3/tissue inhibitor of metalloproteinases 1 and matrix metalloproteinase 9/tissue inhibitor of metalloproteinases 1 ratios. Arthritis Rheum. 2005, 52, 2751–2760. [Google Scholar] [CrossRef]
- De Paiva, C.S.; Chotikavanich, S.; Pangelinan, S.B.; Pitcher, J.D.; Fang, B.; Zheng, X.; Ma, P.; Farley, W.J.; Siemasko, K.F.; Niederkorn, J.Y.; et al. IL-17 disrupts corneal barrier following desiccating stress. Mucosal Immunol. 2009, 2, 243–253. [Google Scholar] [CrossRef] [Green Version]
- Okazaki, T.; Shinagawa, S.; Mikage, H. Vasculitis syndrome—Diagnosis and therapy. J. Gen. Fam. Med. 2017, 18, 72–78. [Google Scholar] [CrossRef]
- Espígol-Frigolé, G.; Corbera-Bellalta, M.; Planas-Rigol, E.; Lozano, E.; Segarra, M.; García-Martínez, A.; Prieto-González, S.; Hernández-Rodríguez, J.; Grau, J.M.; Rahman, M.U.; et al. Increased IL-17A expression in temporal artery lesions is a predictor of sustained response to glucocorticoid treatment in patients with giant-cell arteritis. Ann. Rheum. Dis. 2012, 72, 1481–1487. [Google Scholar] [CrossRef]
- Deng, J.; Younge, B.R.; Olshen, R.A.; Goronzy, J.J.; Weyand, C.M. Th17 and Th1 T-cell responses in giant cell arteritis. Circulation 2010, 121, 906–915. [Google Scholar] [CrossRef] [Green Version]
- Visvanathan, S.; Rahman, M.U.; Hoffman, G.S.; Xu, S.; García-Martínez, A.; Segarra, M.; Lozano, E.; Espígol-Frigolé, G.; Hernández-Rodríguez, J.; Cid, M.C. Tissue and serum markers of inflammation during the follow-up of patients with giant-cell arteritis—A prospective longitudinal study. Rheumatology 2011, 50, 2061–2070. [Google Scholar] [CrossRef] [Green Version]
- Thom, V.; Schmid, S.; Gelderblom, M.; Hackbusch, R.; Kolster, M.; Schuster, S.; Thomalla, G.; Keminer, O.; Pless, O.; Bernreuther, C.; et al. IL-17 production by CSF lymphocytes as a biomarker for cerebral vasculitis. Neurol. Neuroimmunol. Neuroinflamm. 2016, 3, e214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, M.M.-H.; Tseng, W.-N.; Ko, C.-H.; Pan, H.-M.; Hsieh, K.-S.; Kuo, H.-C. Th17- and Treg-related cytokine and mRNA expression are associated with acute and resolving Kawasaki disease. Allergy 2015, 70, 310–318. [Google Scholar] [CrossRef] [PubMed]
- Chi, W.; Yang, P.; Zhu, X.; Wang, Y.; Chen, L.; Huang, X.; Liu, X. Production of interleukin-17 in Behcet’s disease is inhibited by cyclosporin A. Mol. Vis. 2010, 16, 880–886. [Google Scholar]
- Von Vietinghoff, S.; Ley, K. Interleukin 17 in vascular inflammation. Cytokine Growth Factor Rev. 2010, 21, 463–469. [Google Scholar] [CrossRef] [Green Version]
- Della-Torre, E.; Lanzillotta, M.; Doglioni, C. Immunology of IgG4-related disease. Clin. Exp. Immunol. 2015, 181, 191–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haldar, D.; Cockwell, P.; Richter, A.G.; Roberts, K.J.; Hirschfield, G.M. An overview of the diagnosis and management of immunoglobulin G4-related disease. Can. Med. Assoc. J. 2016, 188, 953–961. [Google Scholar] [CrossRef] [Green Version]
- Trampert, D.C.; Hubers, L.M.; Van De Graaf, S.F.; Beuers, U. On the role of IgG4 in inflammatory conditions: Lessons for IgG4-related disease. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2018, 1864, 1401–1409. [Google Scholar] [CrossRef]
- Grados, A.; Ebbo, M.; Piperoglou, C.; Groh, M.; Régent, A.; Samson, M.; Terrier, B.; Loundou, A.; Morel, N.; Audia, S.; et al. T Cell Polarization toward TH2/TFH2 and TH17/TFH17 in Patients with IgG4-Related Disease. Front. Immunol. 2017, 8, 235. [Google Scholar] [CrossRef]
- Akiyama, M.; Yasuoka, H.; Yamaoka, K.; Suzuki, K.; Kaneko, Y.; Kondo, H.; Kassai, Y.; Koga, K.; Miyazaki, T.; Morita, R.; et al. Enhanced IgG4 production by follicular helper 2 T cells and the involvement of follicular helper 1 T cells in the pathogenesis of IgG4-related disease. Arthritis Res. 2016, 18, 167. [Google Scholar] [CrossRef] [Green Version]
- Ohta, N.; Makihara, S.; Okano, M.; Kurakami, K.; Ishida, A.; Furukawa, T.; Suzuki, Y.; Watanabe, T.; Kakehata, S.; Aoyagi, M. Roles of IL-17, Th1, and Tc1 cells in patients with IgG4-related sclerosing sialadenitis. Laryngoscope 2012, 122, 2169–2174. [Google Scholar] [CrossRef]
- Gaffen, S. Recent advances in the IL-17 cytokine family. Curr. Opin. Immunol. 2011, 23, 613–619. [Google Scholar] [CrossRef] [Green Version]
- Accessdata.fda.gov. 2020. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/label/2018/125504s013lbl.pdf (accessed on 30 August 2020).
- Cosentyx Summary of Product Characteristics. Novartis Europharm Limited. Available online: http://www.ema.europa.eu/ema/index.jsp?curl=pages/medicines/human/medicines/003729/human_med_001832.jsp&mid=WC0b01ac058001d124 (accessed on 7 May 2020).
- Clinicaltrials.gov. 2020. Home-Clinicaltrials.Gov. Available online: https://clinicaltrials.gov/ (accessed on 19 August 2020).
- Lebwohl, M.; Strober, B.; Menter, A.; Gordon, K.; Weglowska, J.; Puig, L.; Papp, K.; Spelman, L.; Toth, D.; Kerdel, F.; et al. Phase 3 Studies Comparing Brodalumab with Ustekinumab in Psoriasis. N. Engl. J. Med. 2015, 373, 1318–1328. [Google Scholar] [CrossRef] [PubMed]
- Papp, K.; Reich, K.; Paul, C.; Blauvelt, A.; Baran, W.; Bolduc, C.; Tóth, D.; Langley, R.; Cather, J.; Gottlieb, A.; et al. A prospective phase 3, randomised, double-blind, placebo-controlled study of brodalumab in patients with moderate-to-severe plaque psoriasis. Br. J. Dermatol. 2016, 175, 273–286. [Google Scholar] [CrossRef] [PubMed]
- Gordon, K.B.; Blauvelt, A.; Papp, K.; Langley, R.; Luger, T.; Ohtsuki, M.; Reich, K.; Amato, D.; Ball, S.G.; Braun, D.K.; et al. Phase 3 Trials of Ixekizumab in Moderate-to-Severe Plaque Psoriasis. N. Engl. J. Med. 2016, 375, 345–356. [Google Scholar] [CrossRef] [PubMed]
- Accessdata.fda.gov. 2020. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/label/2017/125521s004lbl.pdf (accessed on 30 August 2020).
- Van Rhijn-Brouwer, F.C.C.; Gremmels, H.; Fledderus, J.O.; Radstake, T.R.D.; Verhaar, M.C.; Van Laar, J.M. Cellular Therapies in Systemic Sclerosis: Recent Progress. Curr. Rheumatol. Rep. 2016, 18, 12. [Google Scholar] [CrossRef] [Green Version]
- Hardy, R.; Raza, K.; Cooper, M.S. Therapeutic glucocorticoids: Mechanisms of actions in rheumatic diseases. Nat. Rev. Rheumatol. 2020, 16, 133–144. [Google Scholar] [CrossRef] [Green Version]
- Tsokos, G.C. Systemic Lupus Erythematosus. N. Engl. J. Med. 2011, 365, 2110–2121. [Google Scholar] [CrossRef] [Green Version]
- Ramos-Casals, M.; Brito-Zerón, P.; Bombardieri, S.; Bootsma, H.; De Vita, S.; Dörner, T.; A Fisher, B.; Gottenberg, J.-E.; Hernández-Molina, G.; Kocher, A.; et al. EULAR recommendations for the management of Sjögren’s syndrome with topical and systemic therapies. Ann. Rheum. Dis. 2019, 79, 3–18. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
| Drug | Mechanism of Action | Description | Current Trials in Systemic Rheumatic Diseases | Trial Identifier |
|---|---|---|---|---|
| Secukinumab | Anti-IL-17A | Human Monoclonal antibody | Phase II trial in Giant cell arteritis | NCT03765788 |
| Phase III trial in Lupus nephritis | NCT04181762 | |||
| Brodalumab | Anti-IL-17RA | Human Monoclonal antibody | Phase III trial in Systemic sclerosis | NCT03957681 |
| Phase I trial in Systemic sclerosis | NCT04368403 | |||
| Ixekizumab | Anti-IL-17A and IL-17A/F | Humanized Monoclonal antibody | - | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rafael-Vidal, C.; Pérez, N.; Altabás, I.; Garcia, S.; Pego-Reigosa, J.M. Blocking IL-17: A Promising Strategy in the Treatment of Systemic Rheumatic Diseases. Int. J. Mol. Sci. 2020, 21, 7100. https://doi.org/10.3390/ijms21197100
Rafael-Vidal C, Pérez N, Altabás I, Garcia S, Pego-Reigosa JM. Blocking IL-17: A Promising Strategy in the Treatment of Systemic Rheumatic Diseases. International Journal of Molecular Sciences. 2020; 21(19):7100. https://doi.org/10.3390/ijms21197100
Chicago/Turabian StyleRafael-Vidal, Carlos, Nair Pérez, Irene Altabás, Samuel Garcia, and Jose M. Pego-Reigosa. 2020. "Blocking IL-17: A Promising Strategy in the Treatment of Systemic Rheumatic Diseases" International Journal of Molecular Sciences 21, no. 19: 7100. https://doi.org/10.3390/ijms21197100