
Cholinergic Modulation of Locomotor Circuits in Vertebrates

1
Institut des Neurosciences Cognitives et Intégratives d’Aquitaine (INCIA), UMR 5287, Université de Bordeaux-CNRS, F-33076 Bordeaux, France
2
Department of Neurosciences, Université de Montréal, Montréal, QC H3C 3J7, Canada
3
Department of Physical Activity Sciences and Research Group in Adapted Physical Activity, Université du Québec à Montréal, Montréal, QC H3C 3P8, Canada
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Int. J. Mol. Sci. 2022, 23(18), 10738; https://doi.org/10.3390/ijms231810738
Submission received: 25 July 2022
/
Revised: 8 September 2022
/
Accepted: 9 September 2022
/
Published: 14 September 2022
(This article belongs to the Special Issue Neuronal Control of Locomotion)
{kind=link}
{kind=link}
Abstract
:Locomotion is a basic motor act essential for survival. Amongst other things, it allows animals to move in their environment to seek food, escape predators, or seek mates for reproduction. The neural mechanisms involved in the control of locomotion have been examined in many vertebrate species and a clearer picture is progressively emerging. The basic muscle synergies responsible for propulsion are generated by neural networks located in the spinal cord. In turn, descending supraspinal inputs are responsible for starting, maintaining, and stopping locomotion as well as for steering and controlling speed. Several neurotransmitter systems play a crucial role in modulating the neural activity during locomotion. For instance, cholinergic inputs act both at the spinal and supraspinal levels and the underlying mechanisms are the focus of the present review. Much information gained on supraspinal cholinergic modulation of locomotion was obtained from the lamprey model. Nicotinic cholinergic inputs increase the level of excitation of brainstem descending command neurons, the reticulospinal neurons (RSNs), whereas muscarinic inputs activate a select group of hindbrain neurons that project to the RSNs to boost their level of excitation. Muscarinic inputs also reduce the transmission of sensory inputs in the brainstem, a phenomenon that could help in sustaining goal directed locomotion. In the spinal cord, intrinsic cholinergic inputs strongly modulate the activity of interneurons and motoneurons to control the locomotor output. Altogether, the present review underlines the importance of the cholinergic inputs in the modulation of locomotor activity in vertebrates.
1. Introduction
The organization of the neuronal structures responsible for generating and controlling locomotor activity among vertebrate species is conserved remarkably well. Comparable neuronal mechanisms operate from agnathans to humans [1,2]. The basic muscle synergies responsible for body propulsion are generated by spinal cord neurons, collectively known as “central pattern generators (CPGs)” for locomotion (for review see [1,3,4,5,6,7,8]). In turn, the spinal CPGs are activated and controlled by supraspinal structures that play a crucial role in starting, maintaining, and stopping locomotion [9,10] as well as controlling speed and direction (reviewed in [11]).
Sensory inputs play a crucial role in adapting locomotor activity to prevailing external and internal conditions [12,13]. Significant advances have been made in characterizing how sensory inputs act on spinal cord neurons and supraspinal structures to control locomotion and in defining the neural mechanisms involved (reviewed in [1,5,13,14,15]).
The supraspinal structures controlling locomotion are in large part organized in a serial fashion. Reticulospinal neurons (RSNs) give rise to descending pathways, comprising axons that make direct and indirect connections with interneurons and motoneurons in the spinal cord [16,17]. Several populations of RSNs have been identified and found to play a crucial role in starting, maintaining, and stopping locomotion [9,10,18,19] (for review see [20]).
The RSNs receive inputs from brainstem and forebrain regions, some of which are specifically dedicated to the control of locomotion. Locomotor regions include the mesencephalic (MLR) and diencephalic (DLR) locomotor regions [21,22,23,24,25]. The DLR corresponds to the zona incerta in the diencephalon [26], and its detailed connections with other forebrain and brainstem regions are not fully understood. In lampreys, DLR neurons have been shown to make monosynaptic glutamatergic connections with downstream RSNs [27]. The MLR was first discovered in the cat in the mid 1960s by the group of G. Orlovsky [28]. Since then, the MLR has been found to be present in all of the vertebrate species examined to date. It controls locomotion through connections to the RSNs [23,29,30,31] (for a recent review, see [32]). The MLR is in turn under the influence of the basal ganglia [33] (for reviews see [1,34,35,36,37]).
As indicated above, sensory inputs play a crucial role in adapting locomotion to the external environment into which the animal is moving. To this end, RSNs receive both direct and relayed sensory information from multiple sensory modalities, and this allows them to generate an adapted descending motor command to the spinal locomotor networks [38,39]. As of now, sensory inputs to RSNs have been shown in the lamprey to originate from skin mechanoreceptors, and from vestibular, visual, and olfactory receptors [15,40,41,42,43,44].
Several neurotransmitter systems modulate the neural circuitry controlling locomotion. Cholinergic neurons intervene along the locomotor control chain by acting on supraspinal and spinal neurons. In vertebrates, acetylcholine (ACh) is also involved in the primary developmental processes such as cell proliferation, migration, growth, and differentiation (for a review see [45]). Cholinergic neurons and ACh receptors appear in the developing neuronal networks during embryonic life. In the rat for example, the two families of ACh receptors, namely the nicotinic and muscarinic ACh receptors (respectively nAChRs and mAChRs), are detected early in the brainstem and in the spinal cord. The nAChRs appear at the embryonic day 12 (E12), whereas the mAChRs at E16. Thereafter, both nAChRs and mAChRs progressively emerge in the more anterior brain regions of the mesencephalon/diencephalon and neocortex during the E14-18 and E18-22 prenatal periods, respectively [46,47].
Amongst a large variety of roles that ACh may have during the animal’s life, it is involved in general motor functions, from the generation of supraspinal motor commands to the activation of muscles in the periphery. ACh plays also an important role in cortical arousal and, consequently, it modulates attentional [48,49,50] and motivational processes (nAChRs: [51]; mAChRs: [52]). The latter effects were proposed to impact motor planning in humans [53]. There are several examples of ACh modulation in the CNS that are not limited to motor function. In this review, we will focus on the mechanisms directly linked to the control of movement, and more specifically, locomotion.
2. Brainstem Cholinergic Mechanisms Controlling Motor Activity
2.1. The MLR Contains ACh Neurons
Large populations of cholinergic neurons are present in the caudal mesencephalon and rostral pons, in regions traditionally associated with the location of the MLR. The initial studies describing the MLR in decerebrate cats [28], revealed that the cuneiform nucleus (CuN) was part of this locomotor region (for reviews see [21,54]). The most striking observation at the time was the tight coupling between the MLR stimulation strength and the intensity of the locomotor output that ensued: as the MLR stimulation intensity increased, the generated locomotor speed also increased proportionally, as if locomotion was controlled by a rheostat. The initial discovery of the MLR ignited significant new interest in the supraspinal control of locomotion. Several animal species became research subjects in the field, and the MLR was functionally identified in all of the vertebrate species in which it was investigated (reviewed in [35,54]). Perhaps the most noticeable observation was the similar location of the MLR in those vertebrate species. Moreover, it was found that the MLR projections were very similar (reviewed in [55]). In the cat, the comparison between the distribution of the choline acetyltransferase-labeled cells (according to Kimura et al. [56]) and the regions that efficiently induced locomotion upon their electrical stimulation [57,58] showed a striking similarity [59]. It is now generally accepted that the MLR contains a mixture of glutamatergic, GABAergic, and cholinergic neurons distributed over several adjacent brainstem nuclei.
The pedunculopontine nucleus (PPN) was also shown in rats to be part of the MLR thanks to the elegant work of E. Garcia-Rill [60,61] (for reviews see [62,63]). The CuN and PPN occupy a large part of the mesencephalon, and, therefore, it is likely that the different parts of these two nuclei could be associated with different aspects of locomotor control. Additionally, it was shown in both lampreys and salamanders, that the laterodorsal tegmental nucleus (LDT) also controlled locomotion efficiently. The LDT is located medially near the caudal pole of the PPN, and it contains a large proportion of cholinergic neurons. In lampreys and in salamanders, the LDT was shown to be the most efficient region from which locomotion could be elicited and controlled [24,64] (for reviews see [37,54]).
It appears therefore that the PPN and CuN, and the LDT, at least in basal vertebrates, constitute the main brainstem nuclei responsible for the locomotor effects of the MLR stimulation (e.g., [24,60,64,65]). In addition, recent work carried out on subjects with Parkinson’s disease [66] showed that the mesencephalic deep brain stimulation of the PPN and CuN areas efficiently treated the freezing of gait, suggesting that the PPN and CuN may also be constitutive of the MLR in humans. These nuclei project to a variety of brain areas [67], and contain different proportions of glutamatergic and cholinergic projection neurons [68,69,70,71].
2.2. MLR Implications in Locomotor Control: Targets, Pathways, and Pharmacology
Imaging experiments in humans revealed that both the CuN and the dorsal PPN are active during imaginary fast walking [72]. Cholinergic neurons of the PPN have also been proposed to be involved in several motor functions including REM sleep, cervical tone, startle responses and locomotion (reviewed in [73]). In primates including humans, PPN cholinergic neurons play a role in controlling gait and posture [72]. Similarly, activating the ventral PPN, which mostly contains cholinergic neurons in cats, suppresses tonic muscle tone, subsequently allowing locomotion [50], whereas the genetic suppression of the vesicular ACh transporter in cholinergic neurons of both the PPN and LDT generates dramatic motor deficits in mice [74]. In contrast, it was shown that a complete excitotoxic lesion of the PPN alone failed to affect gait significantly in rats [75].
The MLR sends cholinergic axons to many key forebrain and brainstem structures controlling locomotion [67]. Therefore, it is likely that the MLR constitutes the major source of ACh modulation for locomotion. The basal ganglia regulate goal-directed behaviors by exerting an inhibitory control on the MLR, as shown by experiments in which a MLR disinhibition allowed for the initiation of locomotion (see [1]). In turn, MLR neurons project to the basal forebrain structures involved in locomotor regulation such as the basal ganglia [76,77] and the ventral tegmental area (VTA; e.g., [78]). Indeed, in mammals, the PPN and LDT cholinergic neurons projecting to the VTA were proposed to modulate locomotion related to drug- and novelty-seeking behaviors, specifically (reviewed in [79]) via both mAChRs [52] and nAChRs [51]. In rats, the optogenetic activation of PPN cholinergic neuron terminals in the VTA increases locomotion, whereas the activation of cholinergic terminals from the LDT tends to reduce locomotion [78] (but see also [80]). These effects presumably result from modulatory effects exerted by the VTA on the striatum.
Stimulation of the PPN in the rat triggers a dopamine release in the basal ganglia through the activation of both ionotropic glutamate receptors and nAChRs, and through the activation of mAChRs, all found on substantia nigra neurons. On the other hand, the M2-type muscarinic autoreceptors located on PPN neurons decrease the nigrostriatal dopamine [76]. In addition, ACh interneurons are present in the striatum where they exert a counterbalancing effect to dopamine inputs, thus regulating the striatum activity (reviewed in [81]). Modeling experiments suggested that this interaction is necessary to induce locomotion [82]. In contrast, the cholinergic innervation originating in the PPN was proposed to allow the basal ganglia to modulate/adapt ongoing locomotion to behavioral constraints (reviewed in [83]). Nevertheless, whatever the source of ACh (local interneurons within basal ganglia or PPN neurons), interactions between ACh and dopamine in the striatum appear essential for motor control, and motor dysfunctions in Parkinson’s disease have been correlated with the dysregulation of such interactions (e.g., [72,84]; for a review, see [81]). Although an increasing amount of data illustrate the impact of the MLR stimulation on forebrain motor structures, the control exerted by the MLR on locomotion is largely associated with the regulation of downstream motor nuclei of the brainstem (reviewed in [35,54]).
The cholinergic neurons seem to represent only about one quarter of the PPN neurons projecting to the reticular nuclei in mice [19]. In contrast, various studies in another rodent species, the rat, showed that most descending PPN neurons [85,86] and a substantial part of the reticular cell-connecting CuN neurons [87] are cholinergic. It is noteworthy that rat CuN cholinergic neurons are involved most likely in sensory modulation and cardiovascular regulation [87], which suggests a secondary role for these cholinergic neurons in locomotor control in this species. Therefore, recent optogenetic studies in rodents have attempted to establish the specific contribution of the glutamatergic and cholinergic neuronal populations in the MLR to the control of locomotion.
Glutamatergic cells in the CuN were shown to play a role in generating the locomotion at different speeds [88,89,90,91], whereas glutamatergic neurons of the PPN rather control locomotion at lower speeds [89,90]. In some cases, locomotion was halted by activating PPN glutamatergic neurons [90,92,93], and the induction or termination of locomotion was shown to depend on the respective activation of the glutamatergic and GABAergic reticulospinal systems of the lower brainstem [19]. Such a MLR-controlled start/stop system seems remarkably well conserved in vertebrates since comparable results were found in the lamprey [10]. The same approach has been used to examine the role of the PPN cholinergic neurons in the control of locomotion. As of now, the results are contradictory as increases and decreases in locomotor output were observed [88,89,90].
All these results strongly suggest that glutamatergic MLR neurons play a predominant role in triggering locomotion through the activation of downstream RSNs compared to cholinergic MLR neurons [94]. This is supported by the observation in cats that cholinergic antagonists seem to produce only little and temporary effects on spontaneous [95] or MLR stimulation-induced locomotion [96]. In striking contrast however, the direct injection of ACh agonists in the reticular formation is sufficient to trigger locomotor bouts in rats, birds, and lampreys [64,97,98], while ACh antagonists block MLR-induced RSN depolarization and locomotion in rats [97,99]. The latter results indicate that ACh may also play a significant role in MLR-Induced locomotion. Therefore, activating either the glutamatergic or cholinergic system independently may yield an incomplete picture of the specific role of these two neurotransmitter systems in controlling locomotor behavior. In addition, because slight variations in the experimental approaches (stimulation site, intensity/frequency parameters) may trigger different motor performances (e.g., [100]) a degree of caution is required in the interpretation of the behavioral observations resulting from localized brain stimulation.
Recently, the group of Bretzner [90] has shown in mice that the joint activation of glutamatergic and cholinergic neurons in the PPN modulated CuN-evoked locomotion, converting running into walking, thereby suggesting that both the glutamatergic and cholinergic systems participate in the MLR command. Interestingly, neurotransmitter interactions in the context of the MLR-controlled locomotion has already been reported in the lamprey, and it may be conserved across vertebrates [36], where forebrain-originating dopamine enhances both the MLR command onto RSNs [33] and RSNs directly [101]. Both the glutamatergic and cholinergic MLR inputs were shown electrophysiologically in the lamprey to converge onto RSNs, and each transmitter system can trigger a sustained RSN depolarization associated with locomotion in a semi-intact preparation (reviewed in [35]). Therefore, analyzing the RSN responses to both the glutamatergic and cholinergic MLR inputs is necessary to eventually understand the exact contribution of each neurotransmitter in the MLR-evoked locomotion in mammals.
2.3. Cholinergic Effects on Reticulospinal Neurons
Whereas the cholinergic effects of the MLR on other supraspinal structures is still not understood at the cellular level, the lamprey model has allowed researchers to gain a better understanding about the MLR-related ACh effects on downstream RSNs in the context of goal-directed locomotion (Figure 1).
The local application of ACh or nicotine on pontine reticulospinal nuclei (MRRN) consistently produced excitatory postsynaptic potentials (EPSPs) in a dose-dependent fashion in the RSNs. Large doses triggered locomotor activity in either isolated brainstem/spinal cord (fictive locomotion in spinal ventral roots) or in semi-intact preparations (active locomotion) [64]. In addition, the ACh-evoked EPSPs showed summation properties coherent with the known graded effect of the MLR stimulation. Moreover, during spontaneous swimming in a semi-intact preparation, both nicotinic agonists significantly accelerated the ongoing rhythm [64]. As the first direct demonstration of ACh inputs on RSNs, this indicates that the ACh inputs from the MLR are likely to play a substantial role in the initiation and the control of locomotion in lampreys, by allowing the pontine RSNs to generate sustained depolarization and firing under cholinergic effects. In lampreys, the pontine RSNs and their activation by ACh are crucial for MLR-induced locomotion, while the bulbar RSNs (PRRN) seem to produce less powerful effects, since only the glutamatergic antagonists prevented the acceleration of the locomotor rhythm by a stronger MLR stimulation [102]. The latter finding underlines the major contribution of the glutamatergic MLR inputs onto the bulbar neurons of the lateral paragigantocellular nucleus reported in mammals [94]. These similarities between lampreys and mammals certainly warrant a much needed, redirected attention to the pontine RSNs in mammals.
2.4. A Parallel Muscarinic Hyperdrive to Boost the Locomotor Output
As indicated above, the MLR projections to RSNs are essential to control locomotion. In lampreys, the MLR projects directly to RSNs via monosynaptic glutamatergic and cholinergic connections, and it also projects to RSNs indirectly via cholinergic connections (Figure 1). The direct cholinergic excitation is nicotinic in nature [64], while the indirect projection is muscarinic [103]. A bath application of muscarine elicits sustained and recurring depolarizations in RSNs. Calcium imaging revealed oscillations in calcium levels that occurred synchronously within the entire RSN population that was imaged. These oscillations were abolished by the bath application of TTX indicating that they were not intrinsic but driven by other neurons. Subsequent experiments revealed that such RSN oscillations were driven by a group of cells located at the junction between the pontine and bulbar reticular formation. The driving cells were named “muscarinoceptive”, and anatomical studies revealed that these neurons project directly to the reticular formation. The resulting effect of activating this group of cells is a marked amplification of the activity in the RSNs and an increase in the duration of locomotor output. The muscarinoceptive cells were shown themselves to respond to muscarine with long-lasting bouts of activity, to receive a direct muscarinic excitation from the MLR, and to send a glutamatergic excitation to the RSNs. Blocking the mAChRs on these neurons dramatically reduced the MLR-induced excitation of RSNs and slowed locomotion. These results further explain the previous observations in the lamprey that muscarine elicited a sustained depolarization in RSNs [104].
The presence of such a boosting mechanism has yet to be demonstrated in other vertebrate species. However, findings from older studies revealed that the activation of muscarinic receptors in the same region of the reticular formation in birds and mammals can elicit bouts of locomotion. Indeed, the locomotor behavior in birds can result from brainstem injections of carbachol, a nonspecific cholinergic agonist, and these effects are blocked by the muscarine receptor antagonist, atropine [98]. Cholinergic inputs are also believed to activate brainstem neurons in mammals [97,99], and a group of muscarine-sensitive neurons receiving cholinergic inputs from the PPN was described in rats in the ventromedial medulla close to the pontine border [105], at a location similar to that of the muscarinoceptive cells in lampreys. The role of these neurons was not described in relation to locomotion in mammals, but the similarities in their properties and location support the idea that they could amplify the reticulospinal descending signals to boost the locomotor output, just as has been observed in lampreys.
The presence of muscarinic effects in birds and mammals, as in lampreys, suggests that the muscarinic amplifying mechanism is conserved in evolution. It further suggests that the supraspinal control of locomotion in vertebrates is not exclusively composed of linear projections down from the forebrain to the spinal cord. Rather, there exists an additional feedforward cholinergic “hyperdrive” component, originating in the MLR and allowing for a supplementary activation of the locomotor system.
2.5. Muscarinic Control of Brainstem Sensory Inputs
In lampreys [40,41] as in mammals (e.g., [106]) the reticular neurons, and especially the RSNs, integrate information from various sensory modalities in order to generate adaptive motor commands. It is also well established that cat RSNs that are rhythmically active during locomotion, in addition receive sensory inputs from a large portion of the body [107]. Interestingly, the MLR has been shown to modulate sensory inputs that reach the RSNs in the lamprey [108] (Figure 1). Indeed, a short duration electrical stimulation of the MLR depresses, for several tens of minutes, the EPSPs evoked in RSNs by trigeminal nerve stimulation. These effects were prevented by perfusing the brainstem with the muscarinic antagonist atropine. Moreover, mAChRs were immunohistochemically identified on the cells that relay trigeminal sensory signals to the RSNs [109], as well as on the RSNs themselves. Similar muscarinic effects were observed after ejecting ACh or the muscarinic agonist pilocarpine directly onto intracellularly recorded RSNs while electrically stimulating the trigeminal nerve [110]. This muscarinic modulation was not related to any changes in the RSN membrane properties, but rather depended on a presynaptic control of the synaptic transmission between the trigeminal relay cells and RSNs. In contrast, the perfusion of atropine strongly potentiated the RSN responses to trigeminal nerve stimulation, suggesting the presence of a tonic inhibitory mAChR-mediated regulation of RSNs and/or trigeminal relay cells. These observations in lampreys suggest that predictable sensory inputs would be gated by MLR inputs, allowing for a smooth behavioral output to occur during goal-directed locomotion. Such a filtering has been proposed in numerous species for phasic sensory inputs impinging on the locomotor CPGs in the spinal cord where there is a strong presynaptic modulation of the primary afferent terminal inputs during fictive locomotion (see below and [111]), but was not envisioned yet in the brainstem. In addition, when the MLR is inactive the muscarinic modulation would be strongly decreased, and this would open the gates for sensory inputs and thus facilitate sensory-evoked locomotion (e.g., escape swimming [40,41]). Moreover, blocking the muscarinic inputs unmasked persistent RSN membrane potential oscillations after applying NMDA onto the RSNs [110] that could further support escape swimming.
3. Cholinergic Mechanisms in the Spinal Cord
Cholinergic neuromodulation plays a crucial role in spinal cord function. The spinal cholinergic system (Figure 2) is exclusively intrinsic to the spinal cord (propriospinal) and the cholinergic modulation results from the activity of a relatively small number of spinal neuronal populations [112]. Among them, the somatic motoneurons (MNs) play a key role during spinal cord early development by releasing ACh onto their neighboring cells, which triggers spontaneous motor bursts that spread over the entire spinal cord (reviewed in [113]). This bursting activity allows the motor networks and neuronal phenotypes to develop properly [114,115,116,117]. Indeed, in the spinal cord, most of the cholinergic cells are MNs that innervate skeletal muscles, with their cell bodies distributed within the ventral gray matter throughout the spinal cord. In mammals, the preganglionic cells (e.g., [118]), a population of “motoneuron-like” neurons with axons in the ventral roots are located in the thoracic and lumbar segments and also use ACh as their neurotransmitter. In addition, various populations of cholinergic interneurons (INs) have also been described, such as small neurons distributed within the dorsal horn and around the central canal, as well as the large partition cells at the border between the dorsal and ventral gray matter [118,119,120]. Although the roles of these INs are less understood, it was suggested that they may constitute a major source of cholinergic neuromodulation in the spinal cord [121,122], notably involved in the control of spinally-generated locomotor activity [123]. Both nicotinic and muscarinic effects were described in the spinal cord.
3.1. Nicotinic Modulation of Spinal Motor Circuits
The nicotinic effects on locomotor networks were mainly attributed to the modulation of the activity of premotor INs [119,124,125,126] or direct effects on the excitability of MNs [119,126,127,128] in various vertebrate species, and lamprey experiments showed no evidence of a modulation of descending commands [129] or long distance-projecting INs in the spinal cord [130]. As observed in many species, MNs are depolarized in response to the nAChR activation [119,124,127,131], the origin of which remains unclear. MNs are known to also release ACh centrally, through axon collaterals terminating notably on the other synergistic MNs, as first demonstrated in adult cats [132,133] and later in other species (e.g., [125,128]). Some spinal cholinergic INs also project directly onto MNs [121,122] (Figure 2). Whereas in mammals, the cholinergic modulation of MN activity seems to be mostly linked with the activation of mAChRs (see below), the nicotinic effects on MNs are widespread in lower vertebrates such as the lamprey [119,126] and the amphibian Xenopus laevis embryo [124,134].
In addition to the local activation of MNs, the nAChR activation from MNs also plays a major role in the homeostatic modulation of the MNs’ own activity, through the activation of the Renshaw recurrent inhibition: when a MN is active, an axon collateral releases ACh, together with glutamate [135], to activate low affinity nAChRs on Renshaw cells [136,137]. The latter cells in turn inhibit MNs via glycinergic/GABAergic neurotransmission [138].
In rodents nAChRs are also present on primary afferent terminals in the dorsal horn [139,140], and a nicotinic modulation of somatosensory information has been found both in the periphery [141] and centrally on the primary afferent terminals [142,143]. The level of polarization of the terminals is now known to be continuously modulated and different mechanisms are involved. A depolarization of the afferent terminals has been classically associated with presynaptic inhibition [144,145,146] and ACh could play a role. In addition to membrane potential changes in primary afferent terminals, action potentials leaving the spinal cord via the dorsal roots (centrifugal sensory discharges) have been known to occur in vertebrates [147]. Such antidromically conducted discharges occur spontaneously or may result from the activation of other sensory afferents, or from the central neural networks [111,148]. Antidromic discharges can also be elicited by the MN activation itself [149]. However, although a direct modulation of sensory afferents by the MNs was extensively described in an invertebrate [150,151] it is unlikely in vertebrates that such discharges result from direct connections from MN axons onto the primary afferent terminals, but more likely from the involvement of spinal inhibitory interneurons co-releasing GABA and ACh [112,143]. Because the primary afferents are responsible for the fast adaptation of the locomotor activity to various behavioral constraints, a nicotinic modulation of local sensory inputs could obviously impact the function of the spinal motor circuits.
3.2. Muscarinic Modulation of Spinal Motor Circuits
In mammals, the cholinergic neuromodulation of the spinal networks is mainly achieved through muscarinic effects with a contribution of several of the mAChRs described so far (for a recent review, see [152]). Originating from the V0c INs, most likely part of the medially-located partition cells, the well characterized C-bouton synapses constitute one source of muscarinic modulation onto MNs (structure and function reviewed in [152,153]; Figure 2). In the in vitro spinal cord preparation of neonatal rats, for example, enhancing the endogenous ACh activity by perfusing the cholinesterase inhibitor edrophonium, triggers episodes of organized fictive locomotion, the pattern of which is comparable to that of active locomotion [96]. An exhaustive analysis of the cholinergic pharmacology demonstrated the lack of nAChR involvement and characterized the mAChRs responsible for such an endogenous neuromodulation and their actions on both MNs and local INs [96]. Interestingly, previous studies in rat neonates also showed some kind of motor activation in lumbar spinal segments in response to the application of exogenous ACh [154], although the extensor/flexor alternation characteristic of locomotion was not observed. These effects were insensitive to large spectrum nAChR antagonists, but were prevented by the classical mAChR blocker, atropine.
Muscarinic agonists exert various, and sometimes opposite effects on the spinal networks. For instance in mice, the endogenous activation of type-3 receptors enhances the amplitude of glutamatergic miniature potentials recorded from MNs and stabilizes the locomotor-related bursting, whereas the type-2 receptors mediate the ACh-induced decrease of glutamatergic miniature potentials while increasing the fictive locomotor frequency and burst amplitude [155]. The link between the changes in miniature potential amplitude and the effects on locomotion remains to be determined. The source of the glutamatergic miniature potentials was not investigated in detail but they could originate, at least partly, in the commissural interneuronal system, which has been demonstrated in rat neonates to be lastingly facilitated following the activation of motoneuronal mAChRs by local cholinergic INs in an atropine-sensitive manner [156]. Although type-2 and type-3 mAChRs exert distinct effects in the mouse spinal cord, these receptors do not seem to be present on the same pools of MNs [155], and a comparable MN subtype-specific muscarinic neuromodulation was also reported in zebrafish [157].
In many vertebrate species, MNs are the main targets of muscarinic modulation in the spinal cord, as mAChR activation globally results in an increase in MN excitability (see however [119,124]) via a large variety of cellular alterations. In urodeles, turtles and rodents, the mAChR activity tends to reduce several potassium currents in spinal MNs, such as the delayed rectifier and the M-current, as well as the hyperpolarization-induced cation current IH and the calcium-activated potassium current responsible for the after-hyperpolarization IAHP [155,156,158,159,160], all of which are responsible for bringing back or maintaining neurons in a hyperpolarized state. In contrast, voltage-gated calcium currents in turtle MNs are enhanced under muscarine, allowing for the generation and maintenance of longer plateau potentials [158,161,162].
Direct motoneuronal effects in combination with additional excitatory effects on spinal network INs [122,123] and local sensory afferents [140,141], result in a global excitation of motor networks under ACh neuromodulation. In the absence of a supplementary excitation provided by other neurotransmitters or neuromodulators, such as glutamate agonists or serotonin, the exogenous ACh (or muscarinic agonists) alone triggers slow non rhythmic ventral root motor bursts in newborn rodents [154] and adult turtles [161]. Those ventral root bursts were recently proposed in rodents to participate in the coupling between somatic and autonomic functions, as an intraspinal mechanism of coordination between the two neuronal functions [163]. In the presence of an additional excitatory drive, the cholinergic agonists rather modulate the ongoing locomotor rhythm in lampreys [126], Xenopus embryos [125], urodeles [164], and mammals (e.g., [155,156,160]).
4. Conclusions
Historically, ACh has been mostly associated with the synaptic transmission from motoneurons to muscles in vertebrates (see however [165]). It progressively became clear that a variety of neuronal populations widely distributed in the brain and spinal cord are cholinergic. The latter neurons are involved in numerous sensorimotor and cognitive functions conferring to the cholinergic system a broader role in the fundamental neuromodulation of neural activities, thus constituting a possible target of therapeutic approaches for a variety of neuronal and non-neuronal, motor and non-motor pathologies (e.g., [66,84,166,167,168]). For instance, the MLR, known to contain abundant populations of cholinergic neurons, is at present the target of new therapeutic approaches for Parkinson’s disease using deep brain stimulation. Currently, mixed effects have been found in patients [169,170], possibly due to the degeneration of MLR neurons in these patients as the disease progresses [171,172]. It has been proposed that deep brain stimulation of the MLR could also be used to ameliorate locomotor function after a partial spinal cord injury. At present, this approach appears to have excellent potential [173,174,175]. Further research is needed to improve our understanding of the cholinergic modulation in the brainstem and spinal cord.
Author Contributions
D.L.R., S.S.B. and R.D. were involved in the conception of this review article, the writing and revision of the text, and the conception of the figures. The three authors approved the submitted version. All authors have read and agreed to the published version of the manuscript.
Funding
Canadian Institutes of Health Research, Grant/Award Number: 15129; Fonds de Recherche du Québec-Santé, Grant/Award Number: 5249; Natural Sciences and Engineering Research Council of Canada, Grant/Award Numbers: 217435-0; Great Lakes Fishery Commission, Grant/Award Numbers: 54021, 54035, 54067. Centre National de la Recherche Scientifique (France); Fondation pour la Recherche Médicale “Equipe FRM 2017-M. Thoby-Brisson” (France) Number: DEQ20170336764.
Acknowledgments
We thank Danielle Veilleux for her expert technical assistance as well as François Auclair for his input on the manuscript and the graphic creation of the figures.
Conflicts of Interest
The authors declare that this review article was written in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Grillner, S.; El Manira, A. Current Principles of Motor Control, with Special Reference to Vertebrate Locomotion. Physiol. Rev. 2020, 100, 271–320. [Google Scholar] [CrossRef] [PubMed]
- Grillner, S. Evolution: Vertebrate Limb Control over 420 Million Years. Curr. Biol. 2018, 28, R162–R164. [Google Scholar] [CrossRef] [PubMed]
- Grillner, S. Control of Locomotion in Bipeds, Tetrapods and Fish. In Handbook of Physiology. The Nervous System II. Motor Control; American Physiological Society: Rockville, MD, USA; Waverly Press: Baltimore, MD, USA, 1981; pp. 1179–1236. [Google Scholar]
- Grillner, S. Neurobiological Bases of Rhythmic Motor Acts in Vertebrates. Science 1985, 228, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Kiehn, O. Decoding the Organization of Spinal Circuits That Control Locomotion. Nat. Rev. Neurosci. 2016, 17, 224–238. [Google Scholar] [CrossRef] [PubMed]
- Wyart, C. Taking a Big Step towards Understanding Locomotion. Trends Neurosci. 2018, 41, 869–870. [Google Scholar] [CrossRef]
- Grillner, S.; Kozlov, A. The CPGs for Limbed Locomotion—Facts and Fiction. IJMS 2021, 22, 5882. [Google Scholar] [CrossRef] [PubMed]
- Grillner, S. Biological Pattern Generation: The Cellular and Computational Logic of Networks in Motion. Neuron 2006, 52, 751–766. [Google Scholar] [CrossRef]
- Juvin, L.; Grätsch, S.; Trillaud-Doppia, E.; Gariépy, J.-F.; Büschges, A.; Dubuc, R. A Specific Population of Reticulospinal Neurons Controls the Termination of Locomotion. Cell Rep. 2016, 15, 2377–2386. [Google Scholar] [CrossRef]
- Grätsch, S.; Auclair, F.; Demers, O.; Auguste, E.; Hanna, A.; Büschges, A.; Dubuc, R. A Brainstem Neural Substrate for Stopping Locomotion. J. Neurosci. 2019, 39, 1044–1057. [Google Scholar] [CrossRef]
- Deliagina, T.G.; Musienko, P.E.; Zelenin, P.V. Nervous Mechanisms of Locomotion in Different Directions. Curr. Opin. Physiol. 2019, 8, 7–13. [Google Scholar] [CrossRef]
- Clarac, F. How Do Sensory and Motor Signals Interact during Locomotion? In Motor Control: Concepts and Issues; Wiley: New York, NY, USA, 1991; pp. 199–221. [Google Scholar]
- Rossignol, S.; Dubuc, R.; Gossard, J.-P. Dynamic Sensorimotor Interactions in Locomotion. Physiol. Rev. 2006, 86, 89–154. [Google Scholar] [CrossRef] [PubMed]
- Rossignol, S.; Bouyer, L. Adaptive Mechanisms of Spinal Locomotion in Cats. Integr. Comp. Biol. 2004, 44, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Daghfous, G.; Green, W.W.; Alford, S.T.; Zielinski, B.S.; Dubuc, R. Sensory Activation of Command Cells for Locomotion and Modulatory Mechanisms: Lessons from Lampreys. Front. Neural Circuits 2016, 10, 18. [Google Scholar] [CrossRef] [PubMed]
- Buchanan, J.T. Spinal Locomotor Inputs to Individually Identified Reticulospinal Neurons in the Lamprey. J. Neurophysiol. 2011, 106, 2346–2357. [Google Scholar] [CrossRef] [PubMed]
- Buchanan, J.T.; Cohen, A.H. Activities of Identified Interneurons, Motoneurons, and Muscle Fibers during Fictive Swimming in the Lamprey and Effects of Reticulospinal and Dorsal Cell Stimulation. J. Neurophysiol. 1982, 47, 948–960. [Google Scholar] [CrossRef]
- Bouvier, J.; Caggiano, V.; Leiras, R.; Caldeira, V.; Bellardita, C.; Balueva, K.; Fuchs, A.; Kiehn, O. Descending Command Neurons in the Brainstem That Halt Locomotion. Cell 2015, 163, 1191–1203. [Google Scholar] [CrossRef]
- Capelli, P.; Pivetta, C.; Soledad Esposito, M.; Arber, S. Locomotor Speed Control Circuits in the Caudal Brainstem. Nature 2017, 551, 373–377. [Google Scholar] [CrossRef]
- Grätsch, S.; Büschges, A.; Dubuc, R. Descending Control of Locomotor Circuits. Curr. Opin. Physiol. 2019, 8, 94–98. [Google Scholar] [CrossRef]
- Shik, M.L.; Orlovsky, G.N. Neurophysiology of Locomotor Automatism. Physiol. Rev. 1976, 56, 465–501. [Google Scholar] [CrossRef]
- McClellan, A.D. Command Systems for Initiating Locomotion in Fish and Amphibians: Parallels to Initiation Systems in Mammals. In Neurobiology of Vertebrate Locomotion: Proceedings of an International Symposium Held at The Wenner-Gren Center, Stockholm, 17–19 June 1985; Grillner, S., Stein, P.S.G., Stuart, D.G., Forssberg, H., Herman, R.M., Eds.; Wenner-Gren Center International Symposium Series; Palgrave Macmillan: London, UK, 1986; pp. 3–20. ISBN 978-1-349-09148-5. [Google Scholar]
- Sirota, M.G.; Di Prisco, G.V.; Dubuc, R. Stimulation of the Mesencephalic Locomotor Region Elicits Controlled Swimming in Semi-Intact Lampreys: Brainstem-Induced Swimming. Eur. J. Neurosci. 2000, 12, 4081–4092. [Google Scholar] [CrossRef]
- Cabelguen, J.-M.; Bourcier-Lucas, C.; Dubuc, R. Bimodal Locomotion Elicited by Electrical Stimulation of the Midbrain in the Salamander Notophthalmus Viridescens. J. Neurosci. 2003, 23, 2434–2439. [Google Scholar] [CrossRef] [PubMed]
- Ryczko, D.; Auclair, F.; Cabelguen, J.-M.; Dubuc, R. The Mesencephalic Locomotor Region Sends a Bilateral Glutamatergic Drive to Hindbrain Reticulospinal Neurons in a Tetrapod: MLR Downstream Connectivity in Salamanders. J. Comp. Neurol. 2016, 524, 1361–1383. [Google Scholar] [CrossRef] [PubMed]
- Parker, S.; Sinnamon, H. Forward Locomotion Elicited by Electrical Stimulation in the Diencephalon and Mesencephalon of the Awake Rat. Physiol. Behav. 1983, 31, 581–587. [Google Scholar] [PubMed]
- El Manira, A.; Pombal, M.A.; Grillner, S. Diencephalic Projection to Reticulospinal Neurons Involved in the Initiation of Locomotion in Adult Lampreys Lampetra Fluviatilis. J. Comp. Neurol. 1997, 389, 603–616. [Google Scholar] [CrossRef]
- Shik, M.L.; Severin, F.V.; Orlovsky, G.N. Control of Walking and Running by Means of Electric Stimulation of the Midbrain. Biofizika 1966, 11, 659–666. [Google Scholar]
- Brocard, F.; Ryczko, D.; Fenelon, K.; Hatem, R.; Gonzales, D.; Auclair, F.; Dubuc, R. The Transformation of a Unilateral Locomotor Command into a Symmetrical Bilateral Activation in the Brainstem. J. Neurosci. 2010, 30, 523–533. [Google Scholar] [CrossRef]
- Garcia-Rill, E.; Skinner, R.D. The Mesencephalic Locomotor Region. II. Projections to Reticulospinal Neurons. Brain Res. 1987, 411, 13–20. [Google Scholar] [CrossRef]
- Skinner, R.D.; Kinjo, N.; Ishikawa, Y.; Biedermann, J.A.; Garcia-Rill, E. Locomotor Projections from the Pedunculopontine Nucleus to the Medioventral Medulla. Neuroreport 1990, 1, 207–210. [Google Scholar] [CrossRef]
- Brownstone, R.M.; Chopek, J.W. Reticulospinal Systems for Tuning Motor Commands. Front. Neural Circuits 2018, 12, 30. [Google Scholar] [CrossRef]
- Ryczko, D.; Grätsch, S.; Auclair, F.; Dubé, C.; Bergeron, S.; Alpert, M.H.; Cone, J.J.; Roitman, M.F.; Alford, S.; Dubuc, R. Forebrain Dopamine Neurons Project down to a Brainstem Region Controlling Locomotion. Proc. Natl. Acad. Sci. USA 2013, 110, E3235–E3242. [Google Scholar] [CrossRef]
- Grillner, S.; Wallén, P.; Saitoh, K.; Kozlov, A.; Robertson, B. Neural Bases of Goal-Directed Locomotion in Vertebrates—An Overview. Brain Res. Rev. 2008, 57, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Le Ray, D.; Juvin, L.; Ryczko, D.; Dubuc, R. Supraspinal Control of Locomotion. In Progress in Brain Research; Elsevier: Amsterdam, The Netherlands, 2011; Volume 188, pp. 51–70. ISBN 978-0-444-53825-3. [Google Scholar]
- Ryczko, D.; Cone, J.J.; Alpert, M.H.; Goetz, L.; Auclair, F.; Dubé, C.; Parent, M.; Roitman, M.F.; Alford, S.; Dubuc, R. A Descending Dopamine Pathway Conserved from Basal Vertebrates to Mammals. Proc. Natl. Acad. Sci. USA 2016, 113, E2440–E2449. [Google Scholar] [CrossRef] [PubMed]
- Ryczko, D.; Dubuc, R. Dopamine and the Brainstem Locomotor Networks: From Lamprey to Human. Front. Neurosci. 2017, 11, 295. [Google Scholar] [CrossRef] [PubMed]
- Dubuc, R.; Brocard, F.; Antri, M.; Fénelon, K.; Gariépy, J.-F.; Smetana, R.; Ménard, A.; Le Ray, D.; Viana Di Prisco, G.; Pearlstein, É.; et al. Initiation of Locomotion in Lampreys. Brain Res. Rev. 2008, 57, 172–182. [Google Scholar] [CrossRef] [PubMed]
- Yeomans, J.S.; Li, L.; Scott, B.W.; Frankland, P.W. Tactile, Acoustic and Vestibular Systems Sum to Elicit the Startle Reflex. Neurosci. Biobehav. Rev. 2002, 26, 1–11. [Google Scholar] [CrossRef]
- Viana Di Prisco, G.; Pearlstein, E.; Robitaille, R.; Dubuc, R. Role of Sensory-Evoked NMDA Plateau Potentials in the Initiation of Locomotion. Science 1997, 278, 1122–1125. [Google Scholar] [CrossRef]
- Viana Di Prisco, G.; Pearlstein, E.; Le Ray, D.; Robitaille, R.; Dubuc, R. A Cellular Mechanism for the Transformation of a Sensory Input into a Motor Command. J. Neurosci. 2000, 20, 8169–8176. [Google Scholar] [CrossRef]
- Karayannidou, A.; Zelenin, P.V.; Orlovsky, G.N.; Deliagina, T.G. Responses of Reticulospinal Neurons in the Lamprey to Lateral Turns. J. Neurophysiol. 2007, 97, 512–521. [Google Scholar] [CrossRef]
- Derjean, D.; Moussaddy, A.; Atallah, E.; St-Pierre, M.; Auclair, F.; Chang, S.; Ren, X.; Zielinski, B.; Dubuc, R. A Novel Neural Substrate for the Transformation of Olfactory Inputs into Motor Output. PLoS Biol. 2010, 8, e1000567. [Google Scholar] [CrossRef]
- Suzuki, D.; Perez-Fernandez, J.; Wibble, T.; Kardamakis, A.; Grillner, S. The Role of the Optic Tectum for Visually Evoked Orienting and Evasive Movements. Proc. Natl. Acad. Sci. USA 2019, 116, 15272–15281. [Google Scholar] [CrossRef]
- Lauder, J.M.; Schambra, U.B. Morphogenetic Roles of Acetylcholine. Environ. Health Perspect. 1999, 107, 5. [Google Scholar]
- Lichtensteiger, W.; Ribary, U.; Schlumpf, M.; Odermatt, B.; Widmer, H.R. Prenatal Adverse Effects of Nicotine on the Developing Brain. Prog. Brain Res. 1988, 73, 137–157. [Google Scholar] [CrossRef] [PubMed]
- Schlumpf, M.; Palacios, J.M.; Cortes, R.; Lichtensteiger, W. Regional Development of Muscarinic Cholinergic Binding Sites in the Prenatal Rat Brain. Neuroscience 1991, 45, 347–357. [Google Scholar] [CrossRef]
- Hasselmo, M.E.; Sarter, M. Modes and Models of Forebrain Cholinergic Neuromodulation of Cognition. Neuropsychopharmacology 2011, 36, 52–73. [Google Scholar] [CrossRef]
- Mena-Segovia, J.; Winn, P.; Bolam, J.P. Cholinergic Modulation of Midbrain Dopaminergic Systems. Brain Res. Rev. 2008, 58, 265–271. [Google Scholar] [CrossRef]
- Takakusaki, K.; Chiba, R.; Nozu, T.; Okumura, T. Brainstem Control of Locomotion and Muscle Tone with Special Reference to the Role of the Mesopontine Tegmentum and Medullary Reticulospinal Systems. J. Neural Transm. 2016, 123, 695–729. [Google Scholar] [CrossRef]
- Morel, C.; Fattore, L.; Pons, S.; Hay, Y.A.; Marti, F.; Lambolez, B.; De Biasi, M.; Lathrop, M.; Fratta, W.; Maskos, U.; et al. Nicotine Consumption Is Regulated by a Human Polymorphism in Dopamine Neurons. Mol. Psychiatry 2014, 19, 930–936. [Google Scholar] [CrossRef]
- Steidl, S. Muscarinic Cholinergic Receptor Antagonists in the VTA and RMTg Have Opposite Effects on Morphine-Induced Locomotion in Mice. Behav. Brain Res. 2017, 323, 111–116. [Google Scholar] [CrossRef]
- Tattersall, T.L.; Stratton, P.G.; Coyne, T.J.; Cook, R.; Silberstein, P.; Silburn, P.A.; Windels, F.; Sah, P. Imagined Gait Modulates Neuronal Network Dynamics in the Human Pedunculopontine Nucleus. Nat. Neurosci. 2014, 17, 449–454. [Google Scholar] [CrossRef]
- Ryczko, D.; Dubuc, R. The Multifunctional Mesencephalic Locomotor Region. CPD 2013, 19, 4448–4470. [Google Scholar] [CrossRef]
- Garcia-Rill, E.; Skinner, R.D.; Conrad, C.; Mosley, D.; Campbell, C. Projections of the Mesencephalic Locomotor Region in the Rat. Brain Res. Bull. 1986, 17, 33–40. [Google Scholar] [CrossRef]
- Kimura, H.; McGeer, P.L.; Peng, J.H.; McGeer, E.G. The Central Cholinergic System Studied by Choline Acetyltransferase Immunohistochemistry in the Cat. J. Comp. Neurol. 1981, 200, 151–201. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Rill, E.; Skinner, R.D.; Gilmore, S.A. Pallidal Projections to the Mesencephalic Locomotor Region (MLR) in the Cat. Am. J. Anat. 1981, 161, 311–321. [Google Scholar] [CrossRef]
- Garcia-Rill, E.; Skinner, R.D.; Jackson, M.B.; Smith, M.M. Connections of the Mesencephalic Locomotor Region (MLR) I. Substantia Nigra Afferents. Brain Res. Bull. 1983, 10, 57–62. [Google Scholar] [CrossRef]
- Garcia-Rill, E. The Basal Ganglia and the Locomotor Regions. Brain Res. 1986, 396, 47–63. [Google Scholar] [CrossRef]
- Garcia-Rill, E.; Houser, C.R.; Skinner, R.D.; Smith, W.; Woodward, D.J. Locomotion-Inducing Sites in the Vicinity of the Pedunculopontine Nucleus. Brain Res. Bull. 1987, 18, 731–738. [Google Scholar] [CrossRef]
- Garcia-Rill, E. The Pedunculopontine Nucleus. Prog. Neurobiol. 1991, 36, 363–389. [Google Scholar] [CrossRef]
- Garcia-Rill, E.; Mahaffey, S.; Hyde, J.R.; Urbano, F.J. Bottom-up Gamma Maintenance in Various Disorders. Neurobiol. Dis. 2019, 128, 31–39. [Google Scholar] [CrossRef]
- Virmani, T.; Urbano, F.J.; Bisagno, V.; Garcia-Rill, E. The Pedunculopontine Nucleus: From Posture and Locomotion to Neuroepigenetics. AIMS Neurosci. 2019, 6, 219–230. [Google Scholar] [CrossRef]
- Le Ray, D.; Brocard, F.; Bourcier-Lucas, C.; Auclair, F.; Lafaille, P.; Dubuc, R. Nicotinic Activation of Reticulospinal Cells Involved in the Control of Swimming in Lampreys: Nicotinic Inputs to Reticulospinal Cells. Eur. J. Neurosci. 2003, 17, 137–148. [Google Scholar] [CrossRef]
- Coles, S.K.; Iles, J.F.; Nicolopoulos-Stournaras, S. The Mesencephalic Centre Controlling Locomotion in the Rat. Neuroscience 1989, 28, 149–157. [Google Scholar] [CrossRef]
- Goetz, L.; Bhattacharjee, M.; Ferraye, M.U.; Fraix, V.; Maineri, C.; Nosko, D.; Fenoy, A.J.; Piallat, B.; Torres, N.; Krainik, A.; et al. Deep Brain Stimulation of the Pedunculopontine Nucleus Area in Parkinson Disease: MRI-Based Anatomoclinical Correlations and Optimal Target. Neurosurgery 2019, 84, 506–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, B.E.; Webster, H.H. Neurotoxic Lesions of the Dorsolateral Pontomesencephalic Tegmentum-Cholinergic Cell Area in the Cat. I. Effects upon the Cholinergic Innervation of the Brain. Brain Res. 1988, 451, 13–32. [Google Scholar] [CrossRef]
- Lai, Y.Y.; Clements, J.R.; Wu, X.Y.; Shalita, T.; Kuo, J.S.; Siegel, J.M. Brainstem Projections to the Ventromedial Medulla in Cat: Retrograde Transport Horseradish Peroxidase and Immunohistochemical Studies. J. Comp. Neurol. 1999, 408, 419–436. [Google Scholar] [CrossRef]
- Mesulam, M.M.; Mufson, E.J.; Wainer, B.H.; Levey, A.I. Central Cholinergic Pathways in the Rat: An Overview Based on an Alternative Nomenclature (Ch1-Ch6). Neuroscience 1983, 10, 1185–1201. [Google Scholar] [CrossRef]
- Pombal, M.A.; Marin, O.; Lez, A.G. Distribution of Choline Acetyltransferase-Immunoreactive Structures in the Lamprey Brain. J. Comp. Neurol. 2001, 431, 105–126. [Google Scholar] [CrossRef]
- Wang, H.-L.; Morales, M. Pedunculopontine and Laterodorsal Tegmental Nuclei Contain Distinct Populations of Cholinergic, Glutamatergic and GABAergic Neurons in the Rat. Eur. J. Neurosci. 2009, 29, 340–358. [Google Scholar] [CrossRef]
- Karachi, C.; Grabli, D.; Bernard, F.A.; Tandé, D.; Wattiez, N.; Belaid, H.; Bardinet, E.; Prigent, A.; Nothacker, H.-P.; Hunot, S.; et al. Cholinergic Mesencephalic Neurons Are Involved in Gait and Postural Disorders in Parkinson Disease. J. Clin. Investig. 2010, 120, 2745–2754. [Google Scholar] [CrossRef]
- Lee, M.S.; Rinne, J.O.; Marsden, C.D. The Pedunculopontine Nucleus: Its Role in the Genesis of Movement Disorders. Yonsei Med. J. 2000, 41, 167–184. [Google Scholar] [CrossRef]
- Janickova, H.; Rosborough, K.; Al-Onaizi, M.; Kljakic, O.; Guzman, M.S.; Gros, R.; Prado, M.A.M.; Prado, V.F. Deletion of the Vesicular Acetylcholine Transporter from Pedunculopontine/Laterodorsal Tegmental Neurons Modifies Gait. J. Neurochem. 2017, 140, 787–798. [Google Scholar] [CrossRef]
- Gut, N.K.; Winn, P. Deep Brain Stimulation of Different Pedunculopontine Targets in a Novel Rodent Model of Parkinsonism. J. Neurosci. 2015, 35, 4792–4803. [Google Scholar] [CrossRef] [PubMed]
- Forster, G.L.; Blaha, C.D. Pedunculopontine Tegmental Stimulation Evokes Striatal Dopamine Efflux by Activation of Acetylcholine and Glutamate Receptors in the Midbrain and Pons of the Rat. Eur. J. Neurosci. 2003, 17, 751–762. [Google Scholar] [CrossRef] [PubMed]
- Woolf, N.J.; Hernit, M.C.; Butcher, L.L. Cholinergic and Non-Cholinergic Projections from the Rat Basal Forebrain Revealed by Combined Choline Acetyltransferase and Phaseolus Vulgaris Leucoagglutinin Immunohistochemistry. Neurosci. Lett. 1986, 66, 281–286. [Google Scholar] [CrossRef]
- Dautan, D.; Souza, A.S.; Huerta-Ocampo, I.; Valencia, M.; Assous, M.; Witten, I.B.; Deisseroth, K.; Tepper, J.M.; Bolam, J.P.; Gerdjikov, T.V.; et al. Segregated Cholinergic Transmission Modulates Dopamine Neurons Integrated in Distinct Functional Circuits. Nat. Neurosci. 2016, 19, 1025–1033. [Google Scholar] [CrossRef]
- Zhang, C.; Liu, X.; Zhou, P.; Zhang, J.; He, W.; Yuan, T.-F. Cholinergic Tone in Ventral Tegmental Area: Functional Organization and Behavioral Implications. Neurochem. Int. 2018, 114, 127–133. [Google Scholar] [CrossRef]
- Xiang, H.-B.; Zhu, W.-Z.; Guan, X.-H.; Ye, D.-W. The Cuneiform Nucleus May Be Involved in the Regulation of Skeletal Muscle Tone by Motor Pathway: A Virally Mediated Trans-Synaptic Tracing Study in Surgically Sympathectomized Mice. Brain 2013, 136, e251. [Google Scholar] [CrossRef]
- Ztaou, S.; Amalric, M. Contribution of Cholinergic Interneurons to Striatal Pathophysiology in Parkinson’s Disease. Neurochem Int 2019, 126, 1–10. [Google Scholar] [CrossRef]
- Lindroos, R.; Kotaleski, J.H. Predicting Complex Spikes in Striatal Projection Neurons of the Direct Pathway Following Neuromodulation by Acetylcholine and Dopamine. Eur. J. Neurosci. 2021, 53, 2117–2134. [Google Scholar] [CrossRef]
- Mena-Segovia, J.; Bolam, J.P. Rethinking the Pedunculopontine Nucleus: From Cellular Organization to Function. Neuron 2017, 94, 7–18. [Google Scholar] [CrossRef]
- Morris, R.; Martini, D.N.; Madhyastha, T.; Kelly, V.E.; Grabowski, T.J.; Nutt, J.; Horak, F. Overview of the Cholinergic Contribution to Gait, Balance and Falls in Parkinson’s Disease. Parkinsonism Relat. Disord. 2019, 63, 20–30. [Google Scholar] [CrossRef]
- Martinez-Gonzalez, C.; van Andel, J.; Bolam, J.P.; Mena-Segovia, J. Divergent Motor Projections from the Pedunculopontine Nucleus Are Differentially Regulated in Parkinsonism. Brain Struct. Funct. 2014, 219, 1451–1462. [Google Scholar] [CrossRef] [PubMed]
- Rye, D.B.; Lee, H.J.; Saper, C.B.; Wainer, B.H. Medullary and Spinal Efferents of the Pedunculopontine Tegmental Nucleus and Adjacent Mesopontine Tegmentum in the Rat. J. Comp. Neurol. 1988, 269, 315–341. [Google Scholar] [CrossRef] [PubMed]
- Shafei, M.N.; Niazmand, S.; Enayatfard, L.; Hosseini, M.; Daloee, M.H. Pharmacological Study of Cholinergic System on Cardiovascular Regulation in the Cuneiform Nucleus of Rat. Neurosci. Lett. 2013, 549, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Roseberry, T.K.; Lee, A.M.; Lalive, A.L.; Wilbrecht, L.; Bonci, A.; Kreitzer, A.C. Cell-Type-Specific Control of Brainstem Locomotor Circuits by Basal Ganglia. Cell 2016, 164, 526–537. [Google Scholar] [CrossRef]
- Caggiano, V.; Leiras, R.; Goñi-Erro, H.; Masini, D.; Bellardita, C.; Bouvier, J.; Caldeira, V.; Fisone, G.; Kiehn, O. Midbrain Circuits That Set Locomotor Speed and Gait Selection. Nature 2018, 553, 455–460. [Google Scholar] [CrossRef] [PubMed]
- Josset, N.; Roussel, M.; Lemieux, M.; Lafrance-Zoubga, D.; Rastqar, A.; Bretzner, F. Distinct Contributions of Mesencephalic Locomotor Region Nuclei to Locomotor Control in the Freely Behaving Mouse. Curr. Biol. 2018, 28, 884–901.e3. [Google Scholar] [CrossRef]
- van der Zouwen, C.I.; Boutin, J.; Fougère, M.; Flaive, A.; Vivancos, M.; Santuz, A.; Akay, T.; Sarret, P.; Ryczko, D. Freely Behaving Mice Can Brake and Turn During Optogenetic Stimulation of the Mesencephalic Locomotor Region. Front. Neural Circuits 2021, 15, 18. [Google Scholar] [CrossRef]
- Dautan, D.; Kovács, A.; Bayasgalan, T.; Diaz-Acevedo, M.A.; Pal, B.; Mena-Segovia, J. Modulation of Motor Behavior by the Mesencephalic Locomotor Region. Cell Rep. 2021, 36, 109594. [Google Scholar] [CrossRef]
- Carvalho, M.M.; Tanke, N.; Kropff, E.; Witter, M.P.; Moser, M.-B.; Moser, E.I. A Brainstem Locomotor Circuit Drives the Activity of Speed Cells in the Medial Entorhinal Cortex. Cell Rep. 2020, 32, 108123. [Google Scholar] [CrossRef]
- Leiras, R.; Cregg, J.M.; Kiehn, O. Brainstem Circuits for Locomotion. Annu. Rev. Neurosci. 2022, 45, 63–85. [Google Scholar] [CrossRef]
- Webster, H.H.; Jones, B.E. Neurotoxic Lesions of the Dorsolateral Pontomesencephalic Tegmentum-Cholinergic Cell Area in the Cat. II. Effects upon Sleep-Waking States. Brain Res. 1988, 458, 285–302. [Google Scholar] [CrossRef]
- Jordan, L.M.; McVagh, J.R.; Noga, B.R.; Cabaj, A.M.; Majczyński, H.; Sławińska, U.; Provencher, J.; Leblond, H.; Rossignol, S. Cholinergic Mechanisms in Spinal Locomotion—Potential Target for Rehabilitation Approaches. Front. Neural Circuits 2014, 8, 132. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Rill, E.; Skinner, R.D. The Mesencephalic Locomotor Region. I. Activation of a Medullary Projection Site. Brain Res. 1987, 411, 1–12. [Google Scholar] [CrossRef]
- Sholomenko, G.N.; Funk, G.D.; Steeves, J.D. Avian Locomotion Activated by Brainstem Infusion of Neurotransmitter Agonists and Antagonists. Exp. Brain Res. 1991, 85, 659–673. [Google Scholar] [CrossRef] [PubMed]
- Homma, Y.; Skinner, R.D.; Garcia-Rill, E. Effects of Pedunculopontine Nucleus (PPN) Stimulation on Caudal Pontine Reticular Formation (PnC) Neurons In Vitro. J. Neurophysiol. 2002, 87, 3033–3047. [Google Scholar] [CrossRef]
- Chang, S.J.; Santamaria, A.J.; Sanchez, F.J.; Villamil, L.M.; Saraiva, P.P.; Benavides, F.; Nunez-Gomez, Y.; Solano, J.P.; Opris, I.; Guest, J.D.; et al. Deep Brain Stimulation of Midbrain Locomotor Circuits in the Freely Moving Pig. Brain Stimul. 2021, 14, 467–476. [Google Scholar] [CrossRef]
- Ryczko, D.; Grätsch, S.; Alpert, M.H.; Cone, J.J.; Kasemir, J.; Ruthe, A.; Beauséjour, P.-A.; Auclair, F.; Roitman, M.F.; Alford, S.; et al. Descending Dopaminergic Inputs to Reticulospinal Neurons Promote Locomotor Movements. J. Neurosci. 2020, 40, 8478–8490. [Google Scholar] [CrossRef]
- Brocard, F.; Dubuc, R. Differential Contribution of Reticulospinal Cells to the Control of Locomotion Induced By the Mesencephalic Locomotor Region. J. Neurophysiol. 2003, 90, 1714–1727. [Google Scholar] [CrossRef]
- Smetana, R.; Juvin, L.; Dubuc, R.; Alford, S. A Parallel Cholinergic Brainstem Pathway for Enhancing Locomotor Drive. Nat. Neurosci. 2010, 13, 731–738. [Google Scholar] [CrossRef]
- Smetana, R.W.; Alford, S.; Dubuc, R. Muscarinic Receptor Activation Elicits Sustained, Recurring Depolarizations in Reticulospinal Neurons. J. Neurophysiol. 2007, 97, 3181–3192. [Google Scholar] [CrossRef]
- Mamiya, K.; Bay, K.; Skinner, R.D.; Garcia-Rill, E. Induction of Long-Lasting Depolarization in Medioventral Medulla Neurons by Cholinergic Input from the Pedunculopontine Nucleus. J. Appl. Physiol. 2005, 99, 1127–1137. [Google Scholar] [CrossRef] [PubMed]
- Kow, L.; Pfaff, D.W. Responses of Medullary Reticulospinal and Other Reticular Neurons to Somatosensory and Brainstem Stimulation in Anesthetized or Freely-Moving Ovariectomized Rats with or without Estrogen Treatment. Exp. Brain Res. 1982, 47, 191–202. [Google Scholar] [CrossRef] [PubMed]
- Drew, T.; Dubuc, R.; Rossignol, S. Discharge Patterns of Reticulospinal and Other Reticular Neurons in Chronic, Unrestrained Cats Walking on a Treadmill. J. Neurophysiol. 1986, 55, 375–401. [Google Scholar] [CrossRef]
- Le Ray, D.; Juvin, L.; Boutin, T.; Auclair, F.; Dubuc, R. A Neuronal Substrate for a State-Dependent Modulation of Sensory Inputs in the Brainstem: MLR-Induced Depression of Sensory Inputs. Eur. J. Neurosci. 2010, 32, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Viana Di Prisco, G.; Boutin, T.; Petropoulos, D.; Brocard, F.; Dubuc, R. The Trigeminal Sensory Relay to Reticulospinal Neurones in Lampreys. Neuroscience 2005, 131, 535–546. [Google Scholar] [CrossRef]
- Le Ray, D.; Brocard, F.; Dubuc, R. Muscarinic Modulation of the Trigemino-Reticular Pathway in Lampreys. J. Neurophysiol. 2004, 92, 926–938. [Google Scholar] [CrossRef]
- Dubuc, R.; Cabelguen, J.M.; Rossignol, S. Rhythmic Fluctuations of Dorsal Root Potentials and Antidromic Discharges of Primary Afferents during Fictive Locomotion in the Cat. J. Neurophysiol. 1988, 60, 2014–2036. [Google Scholar] [CrossRef]
- Barber, R.P.; Phelps, P.E.; Houser, C.R.; Crawford, G.D.; Salvaterra, P.M.; Vaughn, J.E. The Morphology and Distribution of Neurons Containing Choline Acetyltransferase in the Adult Rat Spinal Cord: An Immunocytochemical Study. J. Comp. Neurol. 1984, 229, 329–346. [Google Scholar] [CrossRef]
- Falgairolle, M.; O’Donovan, M.J. Feedback Regulation of Locomotion by Motoneurons in the Vertebrate Spinal Cord. Curr. Opin. Physiol. 2019, 8, 50–55. [Google Scholar] [CrossRef]
- Hanson, M.G.; Landmesser, L.T. Increasing the Frequency of Spontaneous Rhythmic Activity Disrupts Pool-Specific Axon Fasciculation and Pathfinding of Embryonic Spinal Motoneurons. J. Neurosci. 2006, 26, 12769–12780. [Google Scholar] [CrossRef]
- Hanson, M.G.; Landmesser, L.T. Normal Patterns of Spontaneous Activity Are Required for Correct Motor Axon Guidance and the Expression of Specific Guidance Molecules. Neuron 2004, 43, 687–701. [Google Scholar] [CrossRef] [PubMed]
- Myers, C.P.; Lewcock, J.W.; Hanson, M.G.; Gosgnach, S.; Aimone, J.B.; Gage, F.H.; Lee, K.-F.; Landmesser, L.T.; Pfaff, S.L. Cholinergic Input Is Required during Embryonic Development to Mediate Proper Assembly of Spinal Locomotor Circuits. Neuron 2005, 46, 37–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rima, M.; Lattouf, Y.; Abi Younes, M.; Bullier, E.; Legendre, P.; Mangin, J.M.; Hong, E. Dynamic Regulation of the Cholinergic System in the Spinal Central Nervous System. Sci. Rep. 2020, 10, 15338. [Google Scholar] [CrossRef] [PubMed]
- Alkaslasi, M.R.; Piccus, Z.E.; Hareendran, S.; Silberberg, H.; Chen, L.; Zhang, Y.; Petros, T.J.; Le Pichon, C.E. Single Nucleus RNA-Sequencing Defines Unexpected Diversity of Cholinergic Neuron Types in the Adult Mouse Spinal Cord. Nat. Commun. 2021, 12, 2471. [Google Scholar] [CrossRef]
- Quinlan, K.A.; Buchanan, J.T. Cellular and Synaptic Actions of Acetylcholine in the Lamprey Spinal Cord. J. Neurophysiol. 2008, 100, 1020–1031. [Google Scholar] [CrossRef]
- Roberts, A.; Li, W.-C.; Soffe, S.R.; Wolf, E. Origin of Excitatory Drive to a Spinal Locomotor Network. Brain Res. Rev. 2008, 57, 22–28. [Google Scholar] [CrossRef]
- Stepien, A.E.; Tripodi, M.; Arber, S. Monosynaptic Rabies Virus Reveals Premotor Network Organization and Synaptic Specificity of Cholinergic Partition Cells. Neuron 2010, 68, 456–472. [Google Scholar] [CrossRef]
- Zagoraiou, L.; Akay, T.; Martin, J.F.; Brownstone, R.M.; Jessell, T.M.; Miles, G.B. A Cluster of Cholinergic Premotor Interneurons Modulates Mouse Locomotor Activity. Neuron 2009, 64, 645–662. [Google Scholar] [CrossRef]
- Matzner, H.; Zelinger, M.; Cherniak, M.; Anglister, L.; Lev-Tov, A. Rhythmogenic Networks Are Potently Modulated by Activation of Muscarinic Acetylcholine Receptors in the Rodent Spinal Cord. J. Neurochem. 2021, 158, 1263–1273. [Google Scholar] [CrossRef]
- Perrins, R.; Roberts, A. Nicotinic and Muscarinic ACh Receptors in Rhythmically Active Spinal Neurones in the Xenopus Laevis Embryo. J. Physiol. 1994, 478, 221–228. [Google Scholar] [CrossRef]
- Perrins, R.; Roberts, A. Cholinergic Contribution to Excitation in a Spinal Locomotor Central Pattern Generator in Xenopus Embryos. J. Neurophysiol. 1995, 73, 1013–1019. [Google Scholar] [CrossRef] [PubMed]
- Quinlan, K.A.; Placas, P.G.; Buchanan, J.T. Cholinergic Modulation of the Locomotor Network in the Lamprey Spinal Cord. J. Neurophysiol. 2004, 92, 1536–1548. [Google Scholar] [CrossRef] [PubMed]
- Blake, J.F.; Evans, R.H.; Smith, D.A. The Effect of Nicotine on Motoneurones of the Immature Rat Spinal Cord in Vitro. Br. J. Pharmacol. 1987, 90, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Perrins, R.; Roberts, A. Cholinergic and Electrical Motoneuron-to-Motoneuron Synapses Contribute to on-Cycle Excitation during Swimming in Xenopus Embryos. J. Neurophysiol. 1995, 73, 1005–1012. [Google Scholar] [CrossRef]
- Homma, S. Physiology and Pharmacology of Putative Transmitters in Lamprey Central Nervous System. Prog. Neurobiol. 1983, 20, 287–311. [Google Scholar] [CrossRef]
- Martin, A.R.; Wickelgren, W.O.; Berànek, R. Effects of Iontophoretically Applied Drugs on Spinal Interneurones of the Lamprey. J. Physiol. 1970, 207, 653–665. [Google Scholar] [CrossRef]
- Ogier, R.; Liu, X.; Tribollet, E.; Bertrand, D.; Raggenbass, M. Identified Spinal Motoneurons of Young Rats Possess Nicotinic Acetylcholine Receptors of the Heteromeric Family. Eur. J. Neurosci. 2004, 20, 2591–2597. [Google Scholar] [CrossRef]
- Cullheim, S.; Lipsenthal, L.; Burke, R.E. Direct Monosynaptic Contacts between Type-Identified a-Motoneurons in the Cat. Brain Res. 1984, 308, 196–199. [Google Scholar] [CrossRef]
- Cullheim, S.; Kellerth, J.-O.; Conradi, S. Evidence for Direct Synaptic Interconnections between Cat Spinal A-Motoneurones via the Recurrent Axon Collaterals: A Morphological Study Using Intracellular Injection of Horseradish Peroxidase. Brain Res. 1977, 132, 1–10. [Google Scholar] [CrossRef]
- Perrins, R.; Roberts, A. Cholinergic and Electrical Synapses between Synergistic Spinal Motoneurones in the Xenopus Laevis Embryo. J. Physiol. 1995, 485, 135–144. [Google Scholar] [CrossRef]
- Mentis, G.Z.; Alvarez, F.J.; Bonnot, A.; Richards, D.S.; Gonzalez-Forero, D.; Zerda, R.; O’Donovan, M.J. Noncholinergic Excitatory Actions of Motoneurons in the Neonatal Mammalian Spinal Cord. Proc. Natl. Acad. Sci. USA 2005, 102, 7344–7349. [Google Scholar] [CrossRef] [PubMed]
- Lamotte d’Incamps, B.; Ascher, P. High Affinity and Low Affinity Heteromeric Nicotinic Acetylcholine Receptors at Central Synapses. J. Physiol. 2014, 592, 4131–4136. [Google Scholar] [CrossRef] [PubMed]
- Lamotte d’Incamps, B.; Ascher, P. Four Excitatory Postsynaptic Ionotropic Receptors Coactivated at the Motoneuron-Renshaw Cell Synapse. J. Neurosci. 2008, 28, 14121–14131. [Google Scholar] [CrossRef] [PubMed]
- Szczupak, L. Recurrent Inhibition in Motor Systems, a Comparative Analysis. J. Physiol. Paris 2014, 108, 148–154. [Google Scholar] [CrossRef]
- Naser, P.V.; Kuner, R. Molecular, Cellular and Circuit Basis of Cholinergic Modulation of Pain. Neuroscience 2018, 387, 135–148. [Google Scholar] [CrossRef]
- Ribeiro-da-Silva, A.; Cuello, A.C. Choline Acetyltransferase-Immunoreactive Profiles Are Presynaptic to Primary Sensory Fibers in the Rat Superficial Dorsal Horn. J. Comp. Neurol. 1990, 295, 370–384. [Google Scholar] [CrossRef]
- Steen, K.H.; Reeh, P.W. Actions of Cholinergic Agonists and Antagonists on Sensory Nerve Endings in Rat Skin, in Vitro. J. Neurophysiol. 1993, 70, 397–405. [Google Scholar] [CrossRef]
- Genzen, J.R.; Van Cleve, W.; McGehee, D.S. Dorsal Root Ganglion Neurons Express Multiple Nicotinic Acetylcholine Receptor Subtypes. J. Neurophysiol. 2001, 86, 1773–1782. [Google Scholar] [CrossRef]
- Shreckengost, J.; Halder, M.; Mena-Avila, E.; Garcia-Ramirez, D.L.; Quevedo, J.; Hochman, S. Nicotinic Receptor Modulation of Primary Afferent Excitability with Selective Regulation of Aδ-Mediated Spinal Actions. J. Neurophysiol. 2021, 125, 568–585. [Google Scholar] [CrossRef]
- Nicoll, R.A.; Alger, B. Presynaptic Inhibition: Transmitter and Ionic Mechanisms—PubMed. Int. Rev. Neurobiol. 1979, 21, 217–258. [Google Scholar]
- Rudomin, P.; Schmidt, R.F. Presynaptic Inhibition in the Vertebrate Spinal Cord Revisited. Exp. Brain Res. 1999, 129, 1–37. [Google Scholar] [CrossRef] [PubMed]
- Comitato, A.; Bardoni, R. Presynaptic Inhibition of Pain and Touch in the Spinal Cord: From Receptors to Circuits. Int. J. Mol. Sci. 2021, 22, 414. [Google Scholar] [CrossRef]
- Barron, D.H.; Matthews, B.H.C. Intermittent Conduction in the Spinal Cord. J. Physiol. 1935, 85, 73–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubuc, R.; Cabelguen, J.-M.; Rossignol, S. Rhythmic Antidromic Discharges of Single Primary Afferents Recorded in Cut Dorsal Root Filaments during Locomotion in the Cat. Brain Res. 1985, 359, 375–378. [Google Scholar] [CrossRef]
- Decima, E.E.; Goldberg, L.J. Centrifugal Dorsal Root Discharges Induced by Motoneurone Activation. J. Physiol. 1970, 207, 103–118. [Google Scholar] [CrossRef]
- Cattaert, D.; Le Ray, D. Direct Glutamate-Mediated Presynaptic Inhibition of Sensory Afferents by the Postsynaptic Motor Neurons: Glutamate-Mediated Presynaptic Inhibition. Eur. J. Neurosci. 1998, 10, 3737–3746. [Google Scholar] [CrossRef] [PubMed]
- Le Ray, D.; Cattaert, D. Active Motor Neurons Potentiate Their Own Sensory Inputs via Glutamate-Induced Long-Term Potentiation. J. Neurosci. 1999, 19, 1473–1483. [Google Scholar] [CrossRef]
- Mille, T.; Quilgars, C.; Cazalets, J.-R.; Bertrand, S.S. Acetylcholine and Spinal Locomotor Networks: The Insider. Physiol. Rep. 2021, 9, e14736. [Google Scholar] [CrossRef]
- Witts, E.C.; Zagoraiou, L.; Miles, G.B. Anatomy and Function of Cholinergic C Bouton Inputs to Motor Neurons. J. Anat. 2014, 224, 52–60. [Google Scholar] [CrossRef]
- Cowley, K.C.; Schmidt, B.J. A Comparison of Motor Patterns Induced by N-Methyl-D-Aspartate, Acetylcholine and Serotonin in the in Vitro Neonatal Rat Spinal Cord. Neurosci. Lett. 1994, 171, 147–150. [Google Scholar] [CrossRef]
- Nascimento, F.; Spindler, L.R.; Miles, G.B. Balanced Cholinergic Modulation of Spinal Locomotor Circuits via M2 and M3 Muscarinic Receptors. Sci. Rep. 2019, 9, 14051. [Google Scholar] [CrossRef] [PubMed]
- Bertrand, S.S.; Cazalets, J.R. Cholinergic Partition Cells and Lamina X Neurons Induce a Muscarinic-Dependent Short-Term Potentiation of Commissural Glutamatergic Inputs in Lumbar Motoneurons. Front. Neural Circuits 2011, 5, 15. [Google Scholar] [CrossRef] [PubMed]
- Bertuzzi, M.; Ampatzis, K. Spinal Cholinergic Interneurons Differentially Control Motoneuron Excitability and Alter the Locomotor Network Operational Range. Sci. Rep. 2018, 8, 1988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alaburda, A.; Perrier, J.-F.; Hounsgaard, J. An M-like Outward Current Regulates the Excitability of Spinal Motoneurones in the Adult Turtle. J. Physiol. 2002, 540, 875–881. [Google Scholar] [CrossRef] [PubMed]
- Chevallier, S.; Nagy, F.; Cabelguen, J.-M. Cholinergic Control of Excitability of Spinal Motoneurones in the Salamander: Cholinergic Control of Motoneurone Excitability in Salamander. J. Physiol. 2006, 570, 525–540. [Google Scholar] [CrossRef] [PubMed]
- Miles, G.B.; Hartley, R.; Todd, A.J.; Brownstone, R.M. Spinal Cholinergic Interneurons Regulate the Excitability of Motoneurons during Locomotion. Proc. Natl. Acad. Sci. USA 2007, 104, 2448–2453. [Google Scholar] [CrossRef]
- Guertin, P.A.; Hounsgaard, J. L-Type Calcium Channels but Not N-Methyl-d-Aspartate Receptor Channels Mediate Rhythmic Activity Induced by Cholinergic Agonist in Motoneurons from Turtle Spinal Cord Slices. Neurosci. Lett. 1999, 261, 81–84. [Google Scholar] [CrossRef]
- Svirskis, G.; Hounsgaard, J. Transmitter Regulation of Plateau Properties in Turtle Motoneurons. J. Neurophysiol. 1998, 79, 45–50. [Google Scholar] [CrossRef]
- Sourioux, M.; Bertrand, S.S.; Cazalets, J.-R. Cholinergic-Mediated Coordination of Rhythmic Sympathetic and Motor Activities in the Newborn Rat Spinal Cord. PLOS Biol. 2018, 16, e2005460. [Google Scholar] [CrossRef]
- Fok, M.; Stein, R.B. Effects of Cholinergic and Noradrenergic Agents on Locomotion in the Mudpuppy (Necturus Maculatus). Exp. Brain Res. 2002, 145, 498–504. [Google Scholar] [CrossRef]
- Lambert, F.M.; Cardoit, L.; Courty, E.; Bougerol, M.; Thoby-Brisson, M.; Simmers, J.; Tostivint, H.; Le Ray, D. Functional Limb Muscle Innervation Prior to Cholinergic Transmitter Specification during Early Metamorphosis in Xenopus. eLife 2018, 7, e30693. [Google Scholar] [CrossRef] [PubMed]
- Arias, H.R.; Targowska-Duda, K.M.; García-Colunga, J.; Ortells, M.O. Is the Antidepressant Activity of Selective Serotonin Reuptake Inhibitors Mediated by Nicotinic Acetylcholine Receptors? Molecules 2021, 26, 2149. [Google Scholar] [CrossRef] [PubMed]
- Cox, M.A.; Bassi, C.; Saunders, M.E.; Nechanitzky, R.; Morgado-Palacin, I.; Zheng, C.; Mak, T.W. Beyond Neurotransmission: Acetylcholine in Immunity and Inflammation. J. Intern. Med. 2020, 287, 120–133. [Google Scholar] [CrossRef] [PubMed]
- Ohkawara, B.; Ito, M.; Ohno, K. Secreted Signaling Molecules at the Neuromuscular Junction in Physiology and Pathology. IJMS 2021, 22, 2455. [Google Scholar] [CrossRef]
- Hamani, C.; Aziz, T.; Bloem, B.R.; Brown, P.; Chabardes, S.; Coyne, T.; Foote, K.; Garcia-Rill, E.; Hirsch, E.C.; Lozano, A.M.; et al. Pedunculopontine Nucleus Region Deep Brain Stimulation in Parkinson Disease: Surgical Anatomy and Terminology. Stereotact Funct. Neurosurg. 2016, 94, 298–306. [Google Scholar] [CrossRef]
- Hamani, C.; Lozano, A.M.; Mazzone, P.A.M.; Moro, E.; Hutchison, W.; Silburn, P.A.; Zrinzo, L.; Alam, M.; Goetz, L.; Pereira, E.; et al. Pedunculopontine Nucleus Region Deep Brain Stimulation in Parkinson Disease: Surgical Techniques, Side Effects, and Postoperative Imaging. Stereotact Funct. Neurosurg. 2016, 94, 307–319. [Google Scholar] [CrossRef]
- Grabli, D.; Karachi, C.; Folgoas, E.; Monfort, M.; Tande, D.; Clark, S.; Civelli, O.; Hirsch, E.C.; Francois, C. Gait Disorders in Parkinsonian Monkeys with Pedunculopontine Nucleus Lesions: A Tale of Two Systems. J. Neurosci. 2013, 33, 11986–11993. [Google Scholar] [CrossRef]
- Müller, M.L.T.M.; Bohnen, N.I. Cholinergic Dysfunction in Parkinson’s Disease. Curr. Neurol. Neurosci. Rep. 2013, 13, 377. [Google Scholar] [CrossRef]
- Stieglitz, L.H.; Hofer, S.; Bolliger, M.; Oertel, M.F.; Filli, L.; Willi, R.; Cathomen, A.; Meyer, C.; Schubert, M.; Hubli, M.; et al. Deep Brain Stimulation for Locomotion in Incomplete Human Spinal Cord Injury (DBS—SCI): Protocol of a Prospective One—Armed Multi—Centre Study. BJM Open 2021, 11, e047670. [Google Scholar] [CrossRef]
- Bachmann, L.C.; Matis, A.; Lindau, N.T.; Felder, P.; Gullo, M.; Schwab, M.E. Deep Brain Stimulation of the Midbrain Locomotor Region Improves Paretic Hindlimb Function After Spinal Cord Injury in Rats. Sci. Transl. Med. 2013, 5, 208ra146. [Google Scholar] [CrossRef]
- Hofer, A.-S.; Scheuber, M.; Sartori, A.; Good, N.; Stalder, S.; Hammer, N.; Fricke, K.; Schalbetter, S.; Engmann, A.; Weber, R.; et al. Stimulation of the Cuneiform Nucleus Enables Training and Boosts Recovery after Spinal Cord Injury. Brain 2022, awac184. [Google Scholar] [CrossRef] [PubMed]
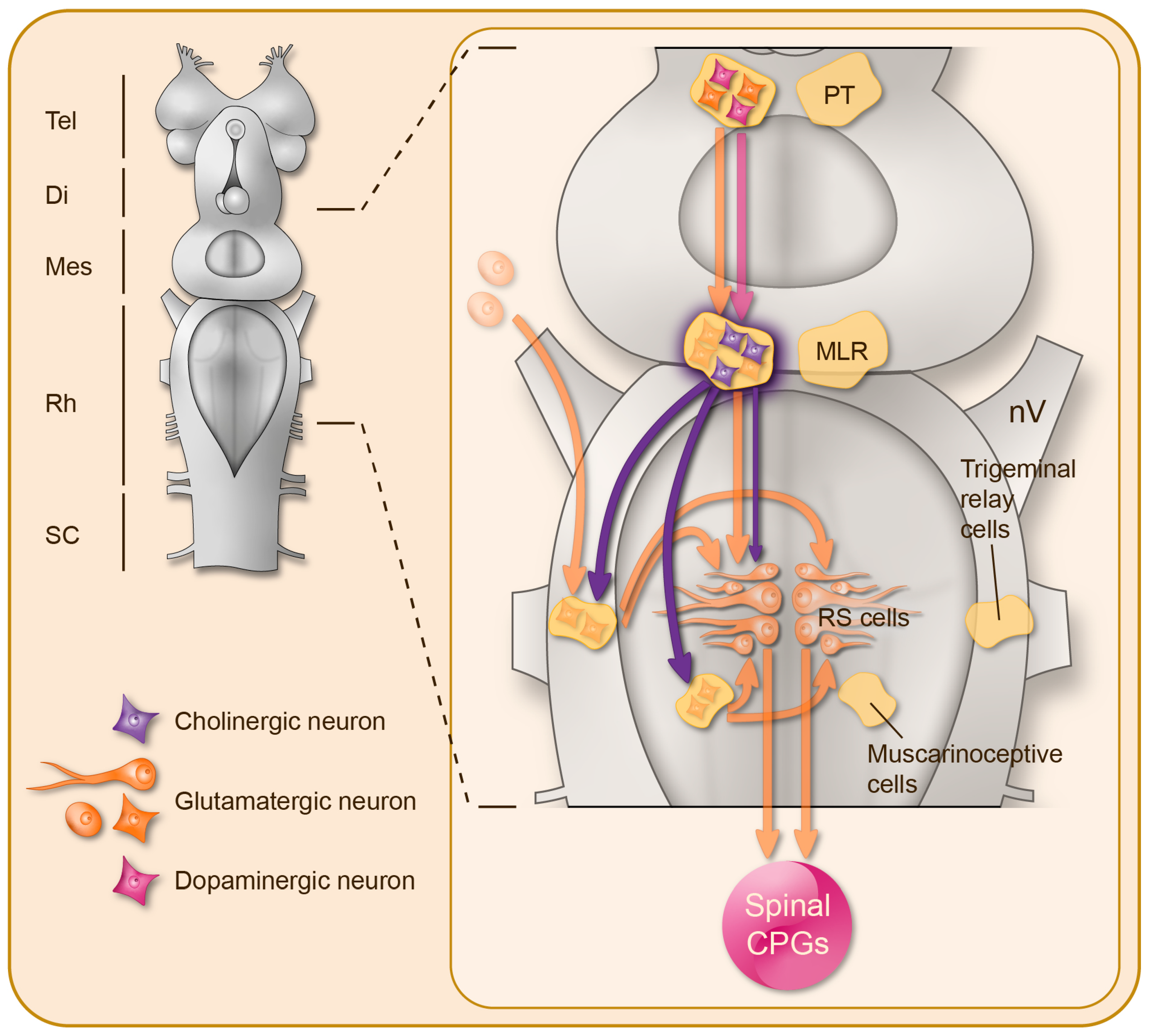
Figure 1.
Cholinergic control of the reticulospinal neurons in the lamprey brainstem. Right: Schematic representation of the lamprey CNS, illustrating from top to bottom: the forebrain (Tel: telencephalon; Di: diencephalon), the brainstem (Mes: mesencephalon; Rh: rhombencephalon), and the spinal cord (SC). Left: Magnification of the brainstem illustrating the distribution of the cholinergic neurons (purple) and their projections. Under the control of glutamatergic (orange) and dopaminergic (pink) descending inputs from the posterior tuberculum, the mesencephalic locomotor region exerts glutamatergic and nicotinic cholinergic excitation on the reticulospinal (RS) neurons that in turn activate the downstream spinal central pattern generators for locomotion (CPGs). There is also a muscarinic cholinergic neuromodulation onto a population of excitatory ponto-medullary muscarinoceptive neurons, which in turn boost the reticulospinal neuron activation, and onto the trigeminal relay cells in order to filter the sensory inputs conveyed by the trigeminal nerves (nV) to the reticulospinal neurons.
Figure 1.
Cholinergic control of the reticulospinal neurons in the lamprey brainstem. Right: Schematic representation of the lamprey CNS, illustrating from top to bottom: the forebrain (Tel: telencephalon; Di: diencephalon), the brainstem (Mes: mesencephalon; Rh: rhombencephalon), and the spinal cord (SC). Left: Magnification of the brainstem illustrating the distribution of the cholinergic neurons (purple) and their projections. Under the control of glutamatergic (orange) and dopaminergic (pink) descending inputs from the posterior tuberculum, the mesencephalic locomotor region exerts glutamatergic and nicotinic cholinergic excitation on the reticulospinal (RS) neurons that in turn activate the downstream spinal central pattern generators for locomotion (CPGs). There is also a muscarinic cholinergic neuromodulation onto a population of excitatory ponto-medullary muscarinoceptive neurons, which in turn boost the reticulospinal neuron activation, and onto the trigeminal relay cells in order to filter the sensory inputs conveyed by the trigeminal nerves (nV) to the reticulospinal neurons.

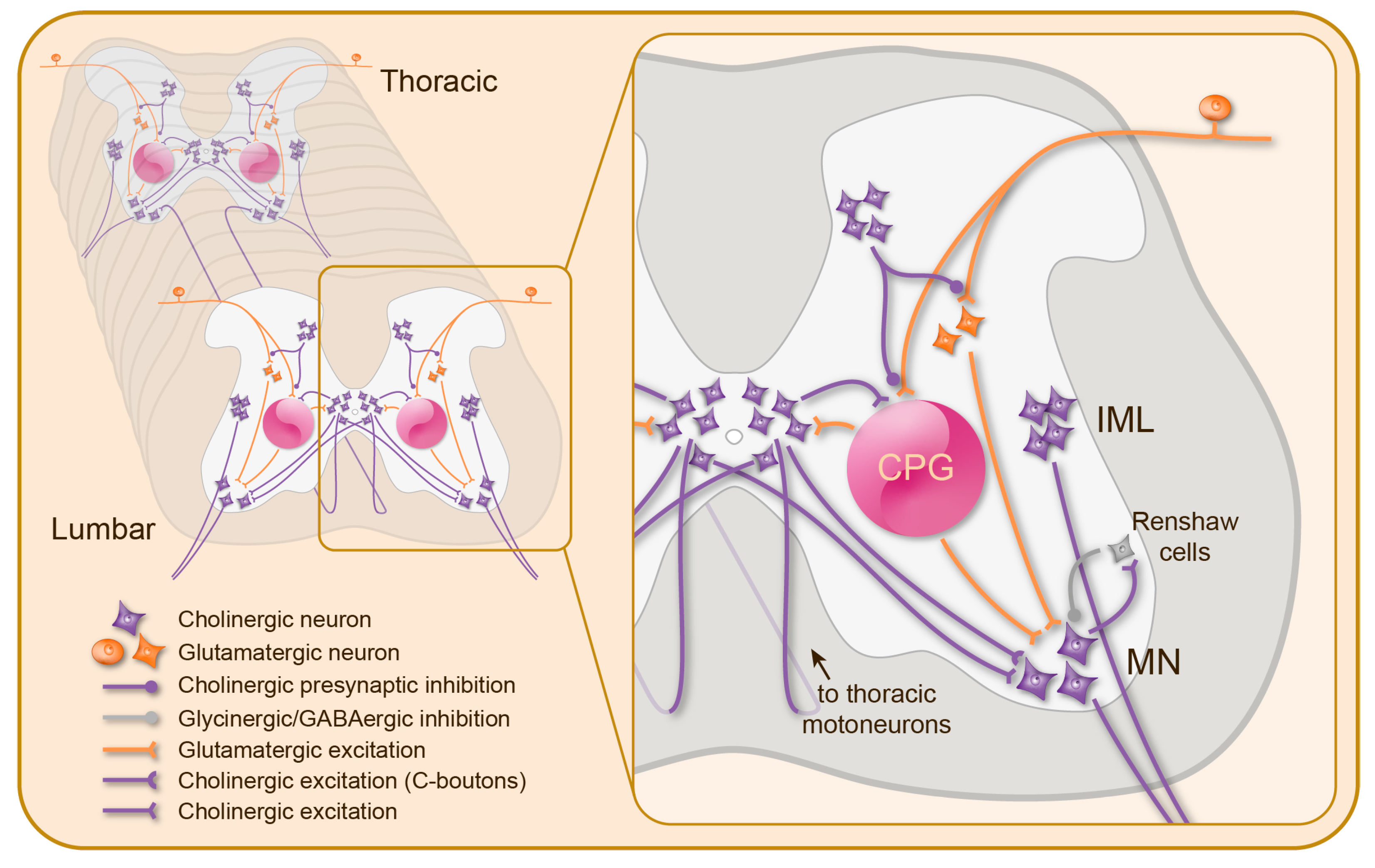
Figure 2.
Cholinergic neuromodulation of spinal motor networks. Motoneurons (MN), intermedio-lateral neurons (IML), and propriospinal interneurons are the only sources of acetylcholine (purple) in the spinal cord, while local glutamatergic excitation (orange) is provided by segmental sensory afferents and interneurons, some of which are part of the central pattern generator (CPG) for locomotion. Except for a group of dorsal interneurons that perform a presynaptic inhibition (filled circle) onto the primary afferent terminals, all propriospinal cholinergic interneurons exert excitation on a variety of cellular targets, including motoneurons (notably via the well described C-boutons) and CPG interneurons.
Figure 2.
Cholinergic neuromodulation of spinal motor networks. Motoneurons (MN), intermedio-lateral neurons (IML), and propriospinal interneurons are the only sources of acetylcholine (purple) in the spinal cord, while local glutamatergic excitation (orange) is provided by segmental sensory afferents and interneurons, some of which are part of the central pattern generator (CPG) for locomotion. Except for a group of dorsal interneurons that perform a presynaptic inhibition (filled circle) onto the primary afferent terminals, all propriospinal cholinergic interneurons exert excitation on a variety of cellular targets, including motoneurons (notably via the well described C-boutons) and CPG interneurons.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Le Ray, D.; Bertrand, S.S.; Dubuc, R. Cholinergic Modulation of Locomotor Circuits in Vertebrates. Int. J. Mol. Sci. 2022, 23, 10738. https://doi.org/10.3390/ijms231810738
AMA Style
Le Ray D, Bertrand SS, Dubuc R. Cholinergic Modulation of Locomotor Circuits in Vertebrates. International Journal of Molecular Sciences. 2022; 23(18):10738. https://doi.org/10.3390/ijms231810738
Chicago/Turabian StyleLe Ray, Didier, Sandrine S. Bertrand, and Réjean Dubuc. 2022. "Cholinergic Modulation of Locomotor Circuits in Vertebrates" International Journal of Molecular Sciences 23, no. 18: 10738. https://doi.org/10.3390/ijms231810738
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.