
Fitness Costs of Two Maize Lepidopteran Pests Fed on Bacillus thuringiensis (Bt) Diets Enriched with Vitamins A and C

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Diets
2.3. Plant Material
2.4. Development Studies
2.5. Enzymatic Studies
2.6. Statistical Analysis
3. Results
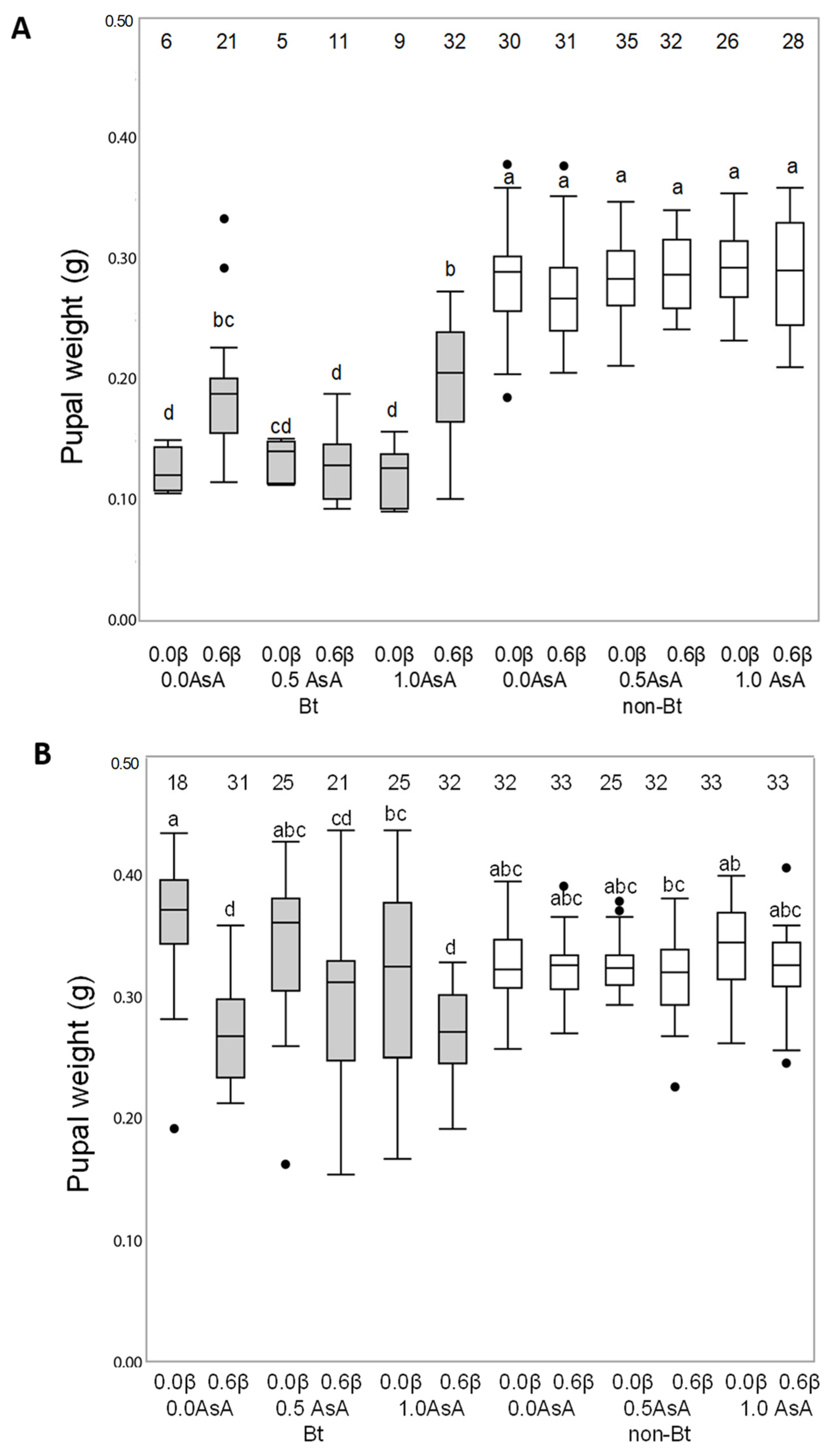
3.1. Effect of Bt Toxin, β-Carotene and Ascorbic Acid (Vitamins A and C) on Larval Development Duration and Pupal Weight
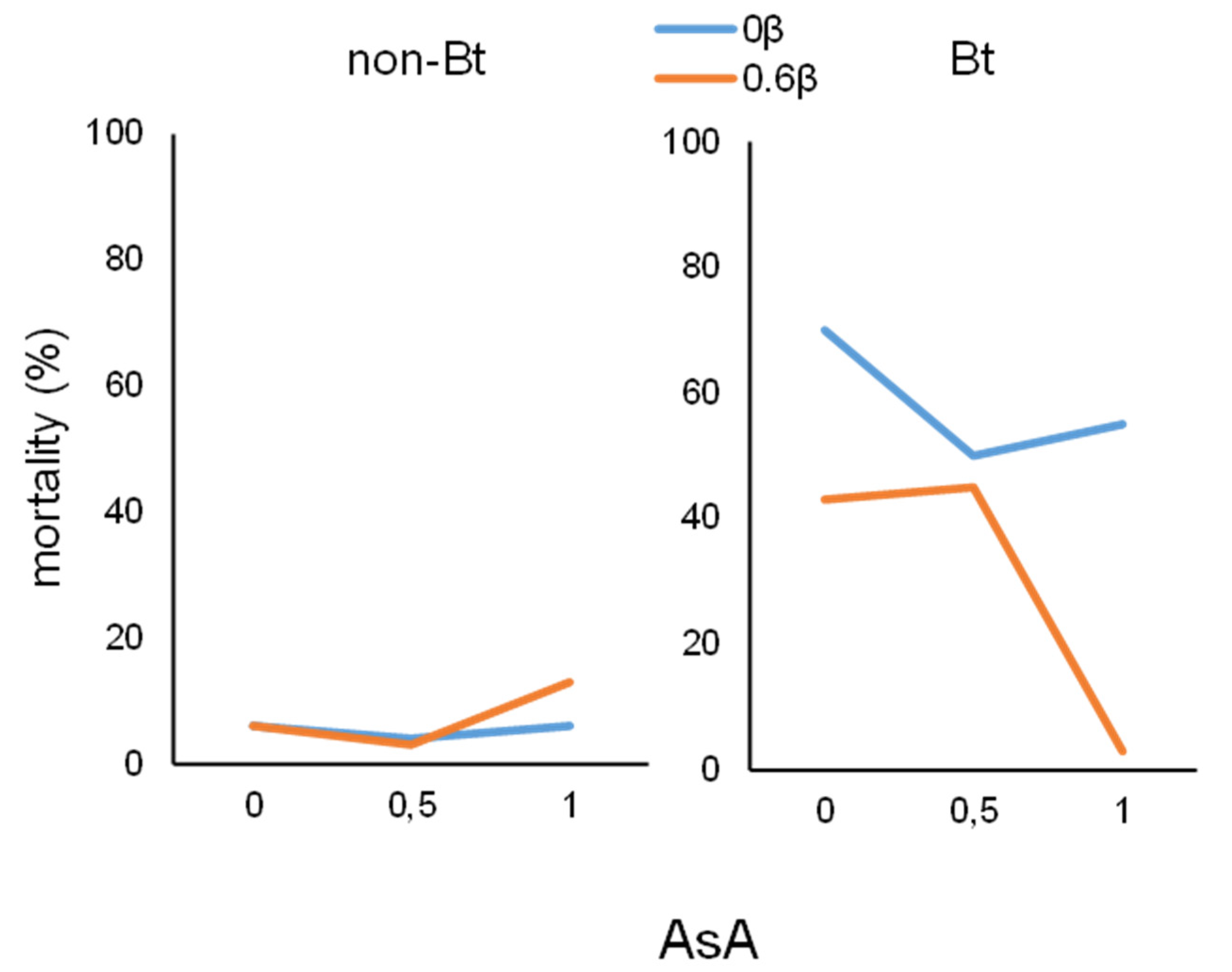
3.2. Larval Mortality
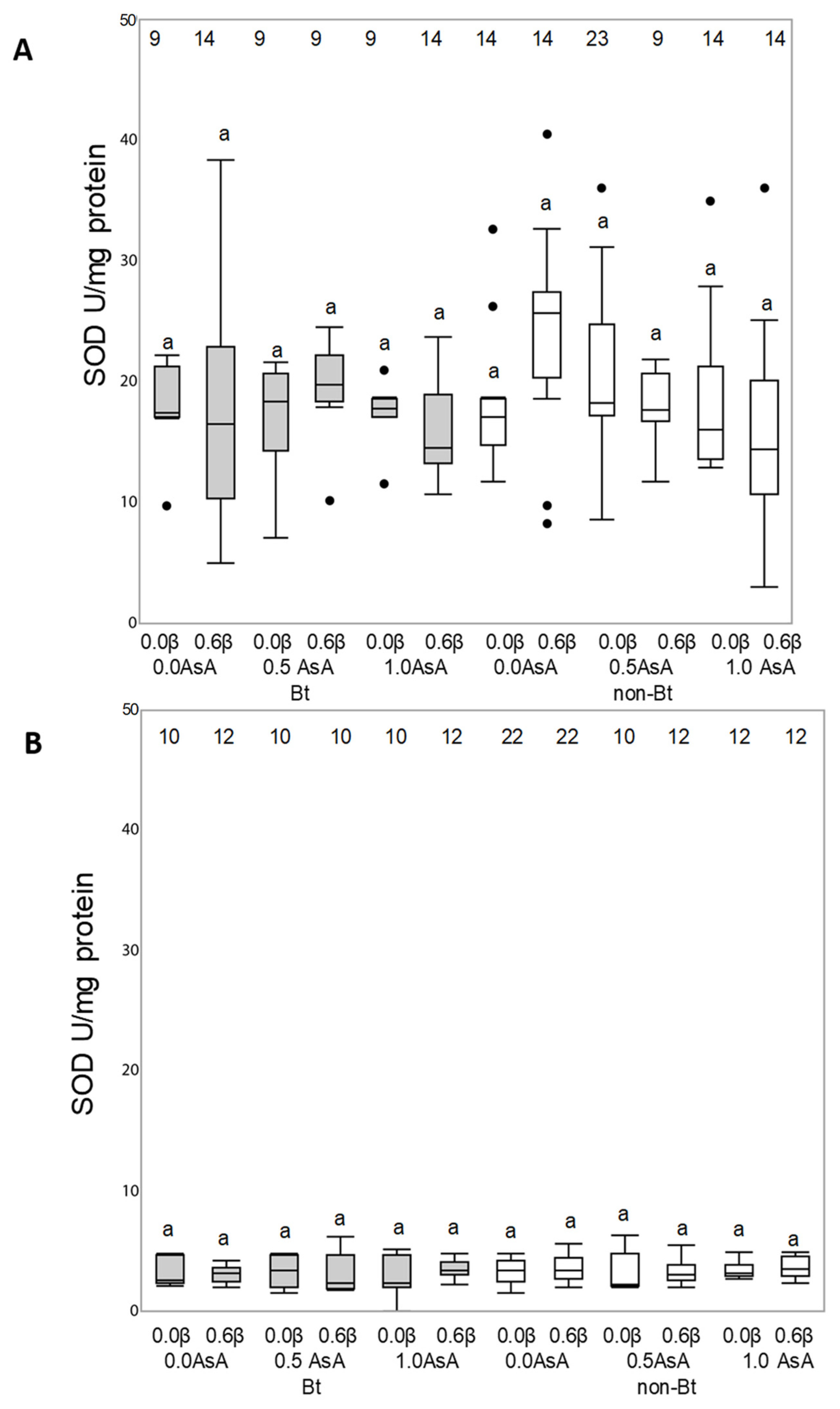
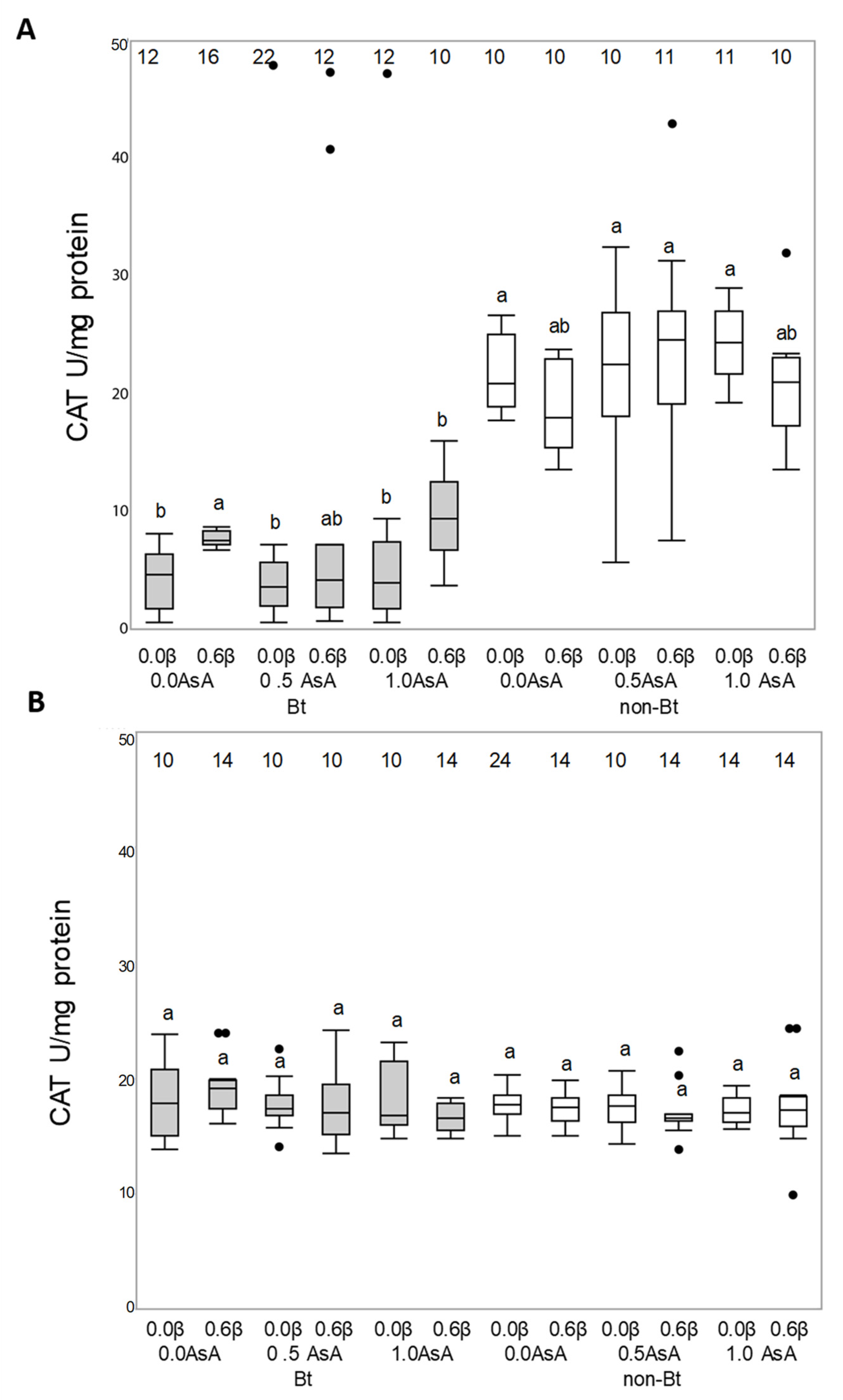
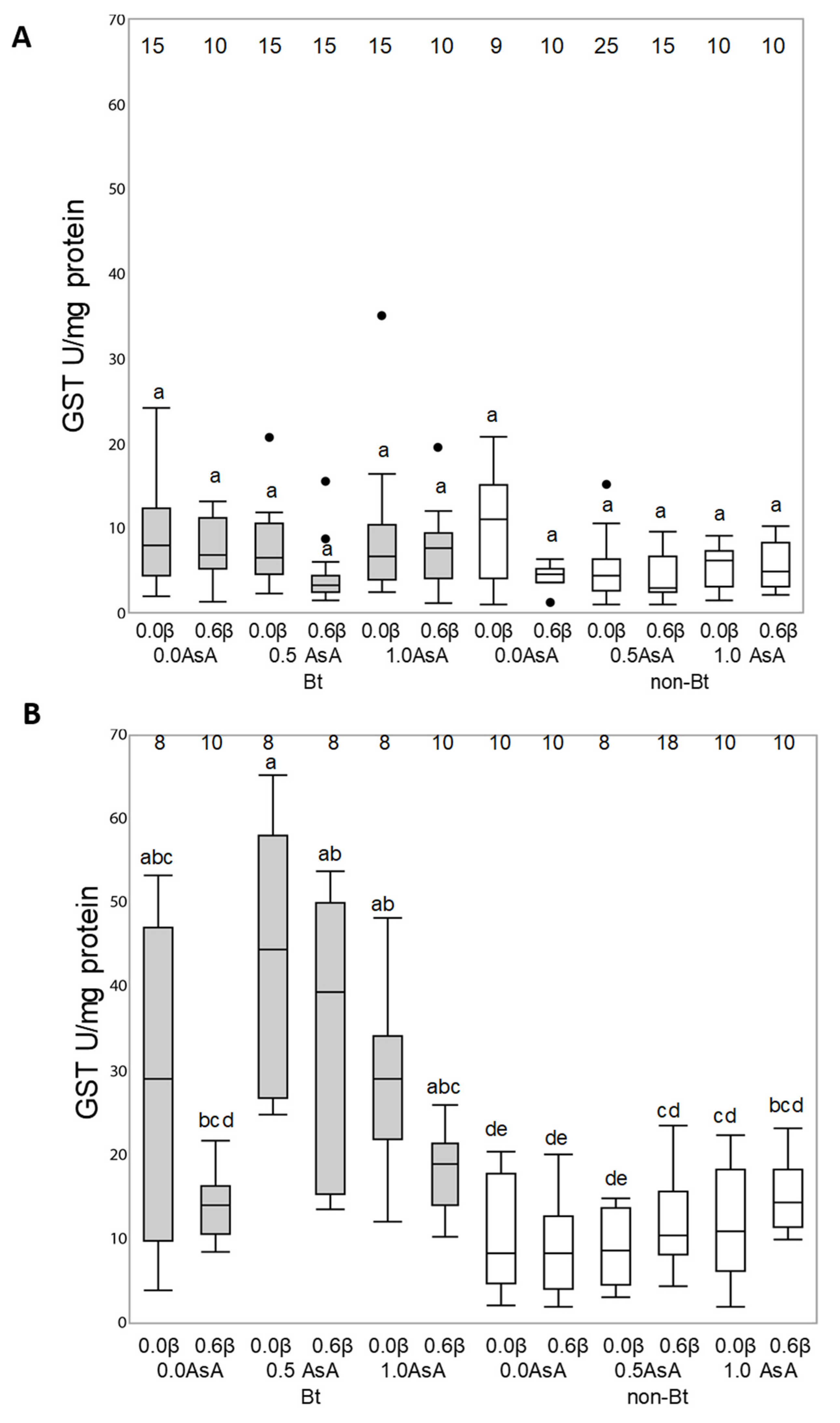
3.3. Antioxidant Enzymatic Activity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IFPRI International Food Policy Research Institute. Global Nutrition Report 2015: Actions and Accountability to Advance Nutrition and Sustainable Development. 2015. Available online: http://www.fao.org/fileadmin/user_upload/raf/uploads/files/129654.pdf (accessed on 20 April 2021).
- FSIN (Food Security International Network, World Food Programme). The 2017 Global Report on Food Crises. 2017. Available online: http://www.fao.org/3/br323e/br323e.pdf (accessed on 20 April 2021).
- Garg, M.; Sharma, N.; Sharma, S.; Kapoor, P.; Kumar, A.; Chunduri, V.; Arora, P. Biofortified Crops Generated by Breeding, Agronomy, and Transgenic Approaches Are Improving Lives of Millions of People around the World. Front Nutr. 2018, 5, 12. [Google Scholar] [CrossRef] [PubMed]
- ISAAA (International Service for the Acquisition of Agri-biotech Applications). Brief 54: 2018 Global Status of Commercialized Biotech/GM Crops: 2018. Available online: https://www.isaaa.org/resources/publications/briefs/54/ (accessed on 10 August 2021).
- Unnevehr, L.; Pray, C.; Paarlberg, R. Addressing micronutrient deficiencies: Alternative interventions and technologies. AgBioforum 2007, 10, 124–134. [Google Scholar]
- ICRISAT International Crops Research Institute for the Semi-Tropics. Annual Report. 2018. Available online: https://www.icrisat.org/wp-ontent/uploads/2019/11/ICRISAT_Annual_Report_2018.pdf (accessed on 20 April 2021).
- Chinedua, O.; Sanoub, E.; Tur-Cardona, J.; Bartolinia, F.; Gheysenc, G.; Speelmanb, S. Farmers’ valuation of transgenic biofortified sorghum for nutritional improvement in Burkina Faso: A latent class approach. Food Policy 2018, 79, 132–140. [Google Scholar] [CrossRef]
- IRRI Annual Report. Golden Rice Meets Food Safety Standards in Three Global Leading Regulatory Agencies. 2018. Available online: https://www.irri.org/news-and-events/news/golden-rice-meets-food-safety-standards-three-global-leading-regulatory-0 (accessed on 20 April 2021).
- FAO. Agricultural Biotechnologies: FAO Statement on Biotechnology. 2020. Available online: http://www.fao.org/biotech/fao-statement-on-biotechnology/en/ (accessed on 20 April 2021).
- Felton, G.W.; Summers, C.B. Antioxidant systems in Insects. Arch. Insect Biochem. Physiol. 1995, 29, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Aucoin, R.R.; Fields, P.; Lewis, M.A.; Philogene, B.J.R.; Arnason, J.T. The protective effect of antioxidants to a phototoxin-sensitive insect herbivore, Manduca sexta. J. Chem. Ecol. 1990, 16, 2913–2924. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S. Biochemical defense of prooxidant plant allelochemicals by herbivorous insects. Biochem. Syst. Ecol. 1992, 20, 269–296. [Google Scholar] [CrossRef]
- Zanga, D.; Sanahuja, G.; Eizaguirre, M.; Albajes, R.; Christou, P.; Capell, T.; Fraser, P.; Gerrisch, G.; López, C. Carotenoids moderate the effectiveness of a Bt gene against the European corn borer, Ostrinia nubilalis. PLoS ONE 2018, 13, 1–16. [Google Scholar] [CrossRef]
- Girón-Calva, P.S.; López, C.; Albacete, A.; Albajes, R.; Christou, P.; Eizaguirre, M. β-carotene and Bacillus thuringiensis insecticidal protein differentially modulate feeding behaviour, mortality and physiology of European corn borer (Ostrinia nubilalis). PLoS ONE 2021, 16, 2. [Google Scholar] [CrossRef]
- Dubovskiy, I.M.; Martemyanov, V.V.; Vorontsova, Y.L.; Rantala, M.J.; Gryzanova, E.V.; Glupovaet, V.V. Effect of bacterial infection on antioxidantactivity and lipidperoxidation in the midgut of Galleria mellonella L. larvae (Lepidoptera, Pyralidae). Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2008, 148, 1–5. [Google Scholar] [CrossRef]
- Ibargutxi, M.A.; Estela, A.; Ferré, J.; Primitivo Caballero, P. Use of Bacillus thuringiensis Toxins for Control of the Cotton Pest Earias insulana (Boisd.) (Lepidoptera: Noctuidae). Appl. Environ. Microbiol. 2006, 72, 437–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bird, L.; Akhurst, R.J. Variation in susceptibility of Helicoverpa armigera (Hubner) and Helicoverpa punctigera (Wallengren) (Lepidoptera: Noctuidae) in Australia to two Bacillus thuringiensis toxins. J. Invertebr. Pathol. 2007, 94, 84–94. [Google Scholar] [CrossRef]
- Hellmich, R.L.; Albajes, R.; Bergvinson, D.; Prasifka, J.R.; Wang, Z.Y.; Weiss, M.J. The Present and Future Role of Insect-Resistant Genetically Modified Maize in IPM. In Integration of Insect-Resistant Genetically Modified Crops within IPM Programs. Progress in Biological Control; Romeis, J., Shelton, A.M., Kennedy, G.G., Eds.; Springer: Dordrecht, The Netherlands, 2008; Volume 5, pp. 119–158. [Google Scholar] [CrossRef] [Green Version]
- Bues, R.; Poitout, S.; Anglade, P.; Robin, J.C. Life cycle and overwintering of Mythimna (syn. Pseudaletia) unipuncta Haw. (Lepidoptera, Noctuidae) in the south of France. Acta Oecol. (Oecol. Appl.) 1986, 7, 151–166. [Google Scholar]
- Mcneil, J.N. The true armyworm, Pseudaletia unipuncta a victim of the pied piper or a seasonal migrant. Insect Sci. Appl. 1987, 8, 591–597. [Google Scholar] [CrossRef]
- Fitt, G.P. The ecology of Heliothis species inrelation to agro-ecosystems. Annu. Rev. Entomol. 1989, 34, 17–52. [Google Scholar] [CrossRef]
- Torres-Vila, L.M.; Rodrı́guez-Molina, M.C.; Lacasa-Plasencia, A.; Bielza-Lino, P. Insecticide resistance of Helicoverpa armigera to endosulfan, carbamates and organophosphates: The Spanish case. Crop. Prot. 2002, 21, 1003–1013. [Google Scholar] [CrossRef]
- Downes, S.; Walsh, T.; Tay, W.T. Bt resistance in Australian insect pest species. Curr. Opin. Insect Sci. 2016, 15, 78–83. [Google Scholar] [CrossRef]
- Eizaguirre, M.; Madeira, F.; Lopez, C. Effects of Bt maize on non-target Lepidopteran pests. In Proceedings of the Fourth Meeting on Ecological Impact of Genetically Modified Organisms, Rostock, Germany, 14–16 May 2009. [Google Scholar]
- Eizaguirre, M.; Albajes, R. Diapause induction in the stem corn-borer, Sesamia nonagrioides (Lepidoptera, Noctuidae). Entomol. Gen. 1992, 17, 277–283. [Google Scholar] [CrossRef]
- Lorch, A.; Then, C. How Much Bt Toxin Do GE MON810 Maize Plants Actually Produce? Greenpeace-Report. 2007. Available online: www.greenpeace.de/fileadmin/gpd/user_upload/themen/gentechnik/greenpeace_bt_maize_engl.pdf (accessed on 20 April 2021).
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- JMP® Pro 15.2.0 Software. SAS Institute. 2019. Available online: http://www.jmp.com/en_us/software.html (accessed on 20 April 2021).
- Apirajkamol, N.; James, B.; Gordon, K.H.J.; Walsh, T.K.; McGaughran, A. Oxidative stress delays development and alters gene expression in the agricultural pest moth, Helicoverpa armigera. Ecol. Evol. 2020, 10, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Govender, L.; Pillay, K.; Siwela, M.; Modi, A.T.; Mabhaudhi, T. Improving the Dietary Vitami A Content of Rural Communities in South Africa by Replacing Non-Biofortified White Maize and Sweet Potato with Biofortified Maize and Sweet Potato in Traditional Dishes. Nutrients 2019, 11, 1198. [Google Scholar] [CrossRef] [Green Version]
- Engelmann, F. The Physiology of Insect Reproduction, 1st ed.; International Series of Monographs in Pure and Applied Biology: Zoology; Kerkut, G.A., Ed.; Pergamon Press: Oxford, UK, 1970; p. 320. ISBN 9781483186535. [Google Scholar]
- Leather, S.R. Size, reproductive potential and fecundity in insects: Things aren’t as simple as they seem. Oikos 1988, 51, 386–389. [Google Scholar] [CrossRef]
- Tammaru, T.; Kaitaniemi, P.; Ruohomäki, K. Realized fecundity in Epirrita automnata (Lepidoptera: Geometridae): Relation to body size and consequences to population dynamics. Oikos 1996, 77, 407–416. [Google Scholar] [CrossRef]
- Calvo, D.; Molina-Rodriguez, J.M. Fecundity–body size relationship and other reproductive aspects of Streblote panda (Lepidoptera: Lasiocampidae). Ann. Entomol. Soc. Am. 2005, 98, 191–196. [Google Scholar] [CrossRef] [Green Version]
- Enayati, A.A.; Ranson, H.; Hemingway, H.J. Insect glutathione transferases and insecticide. Mol. Biol. 2005, 14, 3–8. [Google Scholar] [CrossRef] [Green Version]
- Broderick, N.A.; Raffa, K.F.; Handelsman, J. Chemical modulators of the innate immune response alter gypsymoth larval susceptibility to Bacillus thuringiensis. BMC Microbiol. 2010, 10, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Cabrera, J.; García, M.; Hernández-Crespo, P.; Farinós, G.P.; Ortego, F.; Castañera, P. Resistance to Bt maize in Mythimna unipuncta (Lepidoptera: Noctuidae) is mediated by alteration in Cry1Ab protein activation. Insect Biochem. Mol. Biol. 2013, 43, 635–643. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diets | % Lyophilized Leaves | % AsA | % β-Carotene | |||
|---|---|---|---|---|---|---|
| Maize Leaves | Ascorbic Acid (AsA) | β-Carotene (β) | Non-Bt | Bt | ||
| non-Bt | 0AsA | 0β | 11.1 | 0.0 | 0.0 | 0.0 |
| non-Bt | 0AsA | 0.6β | 11.1 | 0.0 | 0.0 | 0.6 |
| non-Bt | 0.5AsA | 0β | 11.1 | 0.0 | 0.5 | 0.0 |
| non-Bt | 0.5AsA | 0.6β | 11.1 | 0.0 | 0.5 | 0.6 |
| non-Bt | 1AsA | 0β | 11.1 | 0.0 | 1.0 | 0.0 |
| non-Bt | 1AsA | 0.6β | 11.1 | 0.0 | 1.0 | 0.6 |
| Bt | 0AsA | 0β | 0.0 | 11.1 | 0.0 | 0.0 |
| Bt | 0AsA | 0.6β | 0.0 | 11.1 | 0.0 | 0.6 |
| Bt | 0.5AsA | 0β | 0.0 | 11.1 | 0.5 | 0.0 |
| Bt | 0.5AsA | 0.6β | 0.0 | 11.1 | 0.5 | 0.6 |
| Bt | 1AsA | 0β | 0.0 | 11.1 | 1.0 | 0.0 |
| Bt | 1AsA | 0.6β | 0.0 | 11.1 | 1.0 | 0.6 |
| Diet | Ascorbic Acid (AsA) | β-Carotene | Mortality (%) | |
|---|---|---|---|---|
| H. armigera | M. unipuncta | |||
| non-Bt | 0 | 0 | 6 | 3 |
| non-Bt | 0 | 0.6 | 6 | 0 |
| non-Bt | 0.5 | 0 | 4 | 0 |
| non-Bt | 0.5 | 0.6 | 3 | 3 |
| non-Bt | 1 | 0 | 6 | 0 |
| non-Bt | 1 | 0.6 | 13 | 0 |
| Bt | 0 | 0 | 70 | 14 |
| Bt | 0 | 0.6 | 43 | 6 |
| Bt | 0.5 | 0 | 50 | 0 |
| Bt | 0.5 | 0.6 | 45 | 9 |
| Bt | 1 | 0 | 55 | 0 |
| Bt | 1 | 0.6 | 3 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
López, C.; Muñoz, P.; Zanga, D.; Girón-Calva, P.S.; Eizaguirre, M. Fitness Costs of Two Maize Lepidopteran Pests Fed on Bacillus thuringiensis (Bt) Diets Enriched with Vitamins A and C. Insects 2021, 12, 718. https://doi.org/10.3390/insects12080718
López C, Muñoz P, Zanga D, Girón-Calva PS, Eizaguirre M. Fitness Costs of Two Maize Lepidopteran Pests Fed on Bacillus thuringiensis (Bt) Diets Enriched with Vitamins A and C. Insects. 2021; 12(8):718. https://doi.org/10.3390/insects12080718
Chicago/Turabian StyleLópez, Carmen, Pilar Muñoz, Daniela Zanga, Patricia Sarai Girón-Calva, and Matilde Eizaguirre. 2021. "Fitness Costs of Two Maize Lepidopteran Pests Fed on Bacillus thuringiensis (Bt) Diets Enriched with Vitamins A and C" Insects 12, no. 8: 718. https://doi.org/10.3390/insects12080718