Lipid Peroxidation in Hepatopancreas, Gill, and Hemolymph of Male and Female Crabs Platyxanthus orbignyi after Air Exposure



Abstract
:1. Introduction
2. Material and Methods
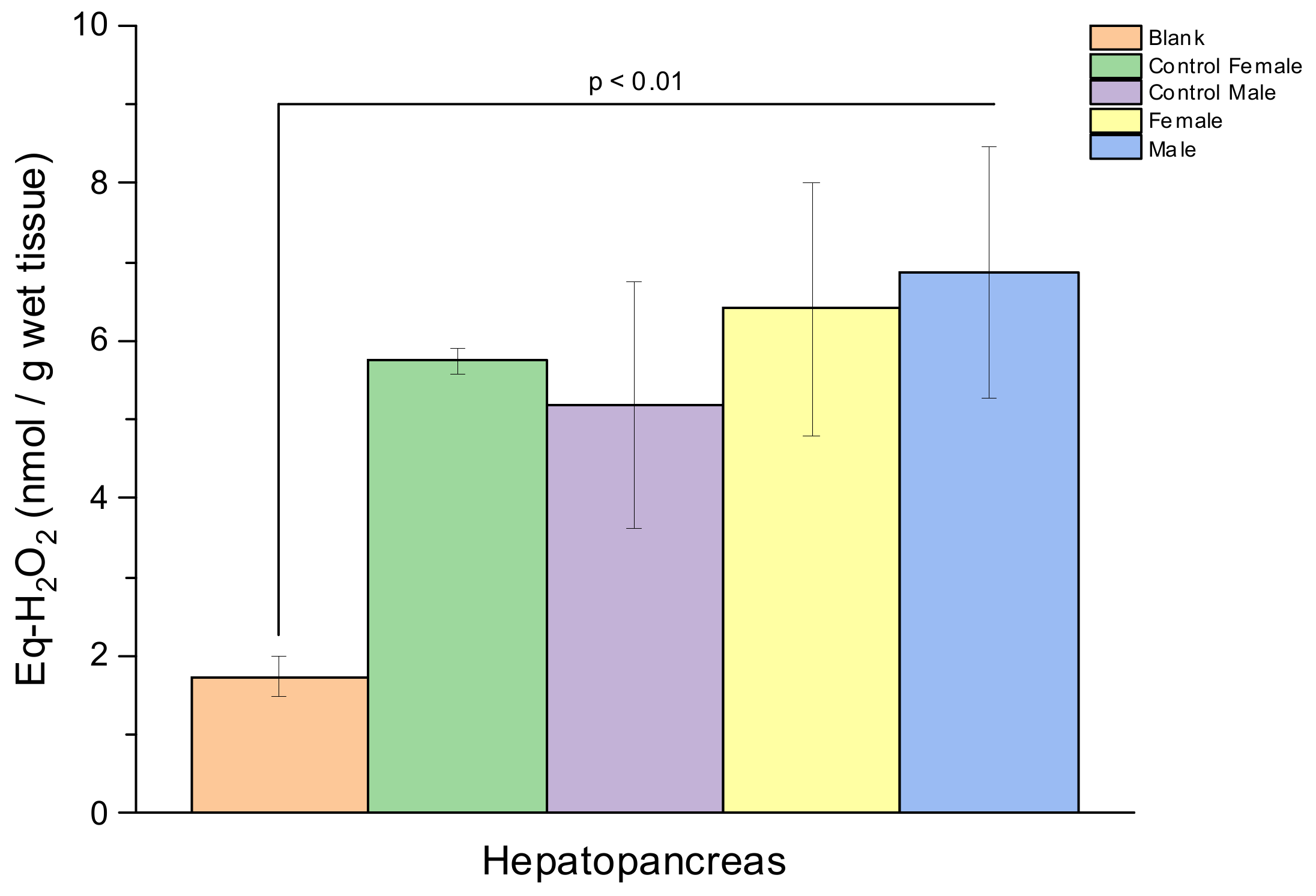
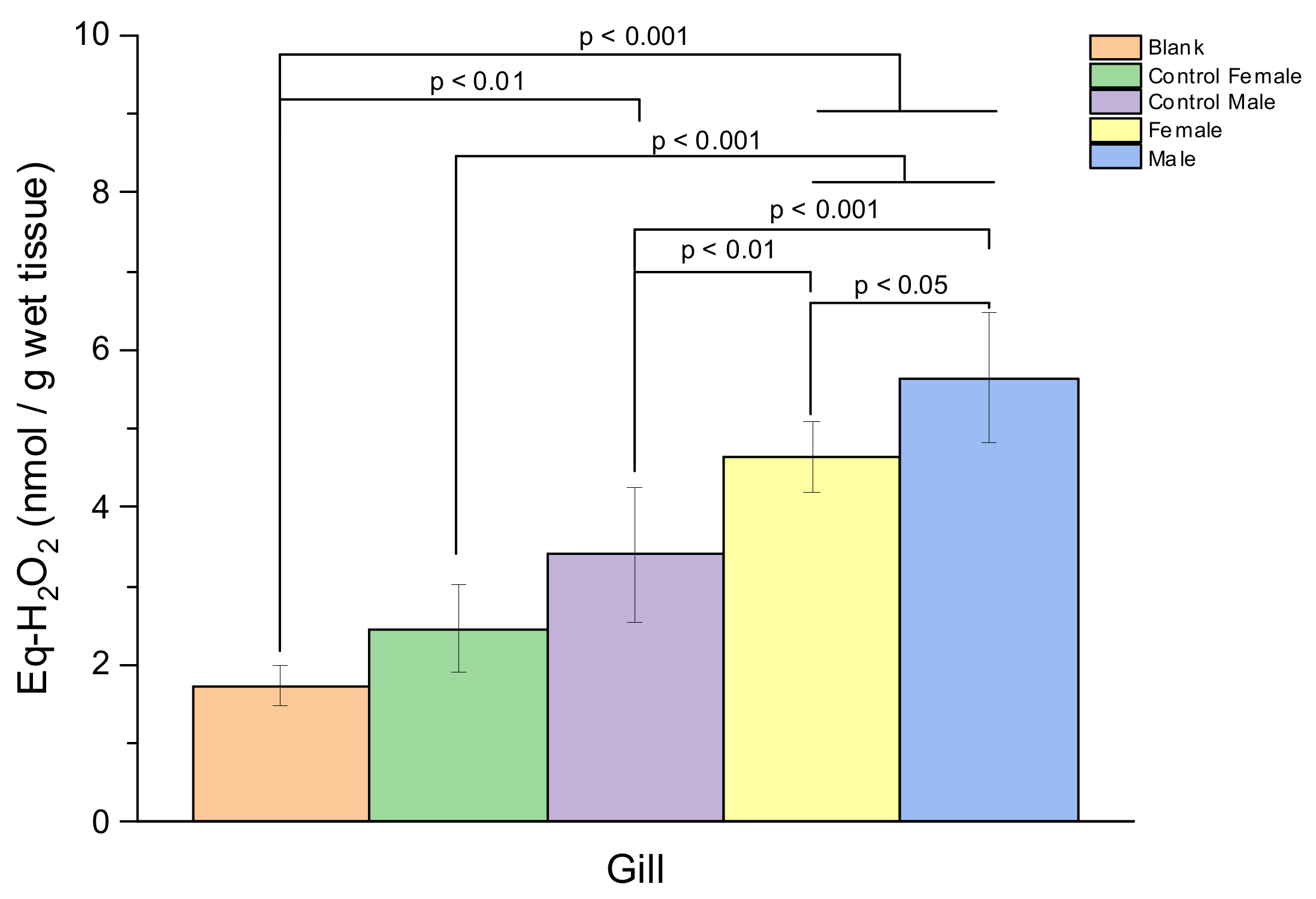
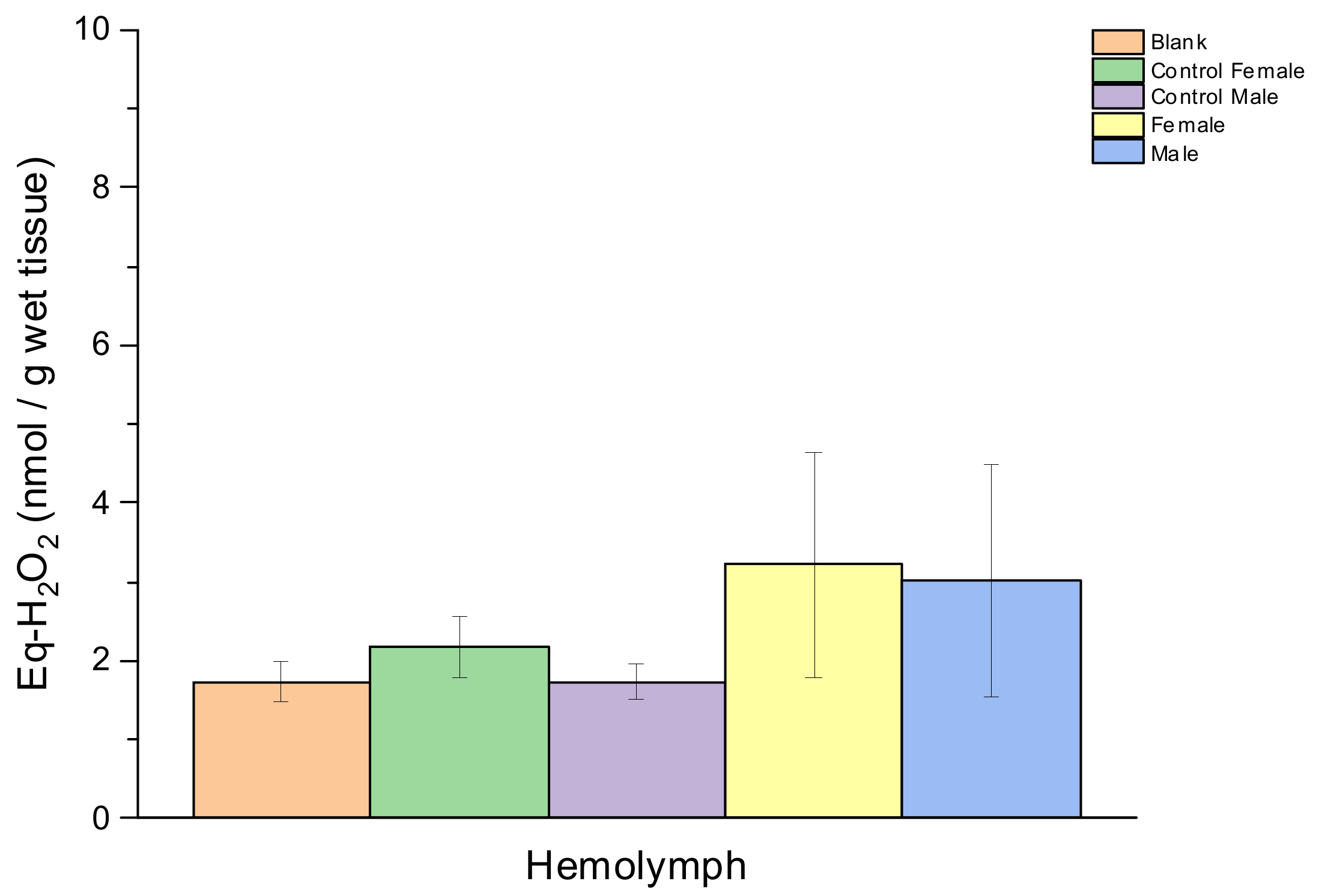
3. Result
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Milne Edwards, H.; Lucas, H. Voyage dans l’Amérique Méridionale: (le Brésil, la République Orientale de l’Uruguay, la République Argentine, la Patagonie, la République du Chili, la République de Bolivia, la Républiquedu Pérou), Exécuté Pendant les Années 1826, 1827, 1828, 1829, 1830; Bertrand, P., Ed.; Crustacés: Paris, France, 1843; Volume IV. [Google Scholar]
- Rathbun, M.J. The stalk-eyed Crustacea of Peru and the adjacent coast. Proc. US Natl. Mus. 1910, 38, 351–620. [Google Scholar] [CrossRef]
- Dillehay, T.D. Where the Land Meets the Sea: Fourteen Millennia of Human History at Huaca Prieta, Peru; University of Texas Press: Austin, TX, USA, 2018; Volume 29. [Google Scholar]
- Romero, M.C.; Ansaldo, M.; Lovrich, G.A. Effect of aerial exposure on the antioxidant status in the subantarctic stone crab Paralomis granulosa (Decapoda: Anomura). Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2007, 146, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Freire, C.A.; Togni, V.G.; Hermes-Lima, M. Responses of free radical metabolism to air exposure or salinity stress, in crabs (Callinectes danae and C. ornatus) with different estuarine distributions. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2011, 160, 291–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Lima, T.M.; Geihs, M.A.; Nery, L.E.M.; Maciel, F.E. Air exposure behavior of the semiterrestrial crab Neohelice granulata allows tolerance to severe hypoxia but not prevent oxidative damage due to hypoxia–reoxygenation cycle. Physiol. Behav. 2015, 151, 97–101. [Google Scholar] [CrossRef] [PubMed]
- De Fur, P.L.; Pease, A.; Siebelink, A.; Elfers, S. Respiratory responses of blue crabs, Callinectes sapidus, to emersion. Comp. Biochem. Physiol. Part A Physiol. 1988, 89, 97–101. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine, 4th ed.; Oxford University Press: Oxford, UK, 2007. [Google Scholar]
- Hermes-Lima, M. Quantification of lipid peroxidation in tissue extracts based on Fe(III)xylenol orange complex formation. Free Radic. Biol. Med. 1995, 19, 271–280. [Google Scholar] [CrossRef]
- Xu, J.; Hwang, J.C.; Lees, H.A.; Wohlgemuth, S.E.; Knutson, M.D.; Judge, A.R.; Dupont-Versteegden, E.E.; Marzetti, E.; Leeuwenburgh, C. Long-term perturbation of muscle iron homeostasis following hindlimb suspension in old rats is associated with high levels of oxidative stress and impaired recovery from atrophy. Exp. Gerontol. 2012, 47, 100–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmitt, A.; Santos, E.A. Behaviour and haemolymphatic ionic composition of the intertidal crab Chasmagnathus granulata dana, 1851 (Crustacea: Decapoda) during emersion. Comp. Biochem. Physiol. Part A Physiol. 1993, 106, 337–342. [Google Scholar] [CrossRef]
- Stillman, J.; Somero, G. Adaptation to temperature stress and aerial exposure in congeneric species of intertidal porcelain crabs (genus Petrolisthes): Correlation of physiology, biochemistry and morphology with vertical distribution. J. Exp. Biol. 1996, 199, 1845–1855. [Google Scholar]
- Jerome, F.C.; Hassan, A.A.; Chukwuka, A.V. Sex-specific affinity for redox-active metals influences antioxidant responses of Callinectes amnicola (blue crab) populations in littoral and open water habitats of a tropical coastal lagoon. Mar. Ecol. 2017, 38, e12437. [Google Scholar] [CrossRef]
- Ministerio de la Producción. Establecen Talla Mínima de Captura (TMC) del Recurso Cangrejo Violáceo, y Prohíben la Extracción, Recepción, Transporte, Almacenamiento, Procesamiento y Comercialización de Hembras Ovígeras; Perú: Art. 2 Ley General de Pesca. D.L. 25977; Resolución Ministerial: El Peruano, Peru, 2016; pp. 544300–584299.
- Ortega, P.; Vitorino, H.A.; Moreira, R.G.; Pinheiro, M.A.; Almeida, A.A.; Custódio, M.R.; Zanotto, F.P. Physiological differences in the crab Ucides cordatus from two populations inhabiting mangroves with different levels of cadmium contamination. Environ. Toxicol. Chem. 2016, 36, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.-Y.; Hunt, J.V.; Wolff, S.P. Ferrous ion oxidation in the presence of xylenol orange for detection of lipid hydroperoxide in low density lipoprotein. Anal. Biochem. 1992, 202, 384–389. [Google Scholar] [CrossRef]
- Vitorino, H.A.; Alta, R.Y.P.; Alta, E.C.P.; Ortega, P. Hepatopancreatic cells of a stone crab Menippe frontalis from Perú: Separation, viability study, and evaluation of lipoperoxidation against cadmium contamination. Vitr. Cell. Dev. Biol. 2017, 53, 778–781. [Google Scholar] [CrossRef] [PubMed]
- Sokal, R.; Rohlf, F. Biometry: The Principles and Practice of Statistics in Biological Research, 3rd ed.; W.H. Freeman and Company: New York, NY, USA, 1995. [Google Scholar]
- Laitano, M.V.; Farías, N.E.; Cledón, M. Prey preference of the stone crab Platyxanthus crenulatus (Decapoda: Platyxanthidae) in laboratory conditions. Nauplius 2013, 21, 17–23. [Google Scholar] [CrossRef] [Green Version]
- Barnwell, F.H. The Role of Rhythmic Systems in the Adaptation of Fiddler Crabs to the Intertidal Zone; American Zoologist; Oxford University Press: Oxford, UK, 1986; Volume 8, pp. 569–583. [Google Scholar]
- Styrishave, B.; Andersen, O. Seasonal variations in hepatopancreas fatty acid profiles of two colour forms of shore crabs, Carcinus maenas. Mar. Biol. 2000, 137, 415–422. [Google Scholar] [CrossRef]
- Monserrat, J.M.; Geracitano, L.A.; Pinho, G.L.L.; Vinagre, T.M.; Faleiros, M.; Alciati, J.C.; Bianchini, A. Determination of Lipid Peroxides in Invertebrates Tissues Using the Fe(III) Xylenol Orange Complex Formation. Arch. Environ. Contam. Toxicol. 2003, 45, 177–183. [Google Scholar] [CrossRef]
- Pinheiro, M.A.A.; e Silva, P.P.G.; de Almeida Duarte, L.F.; Almeida, A.A.; Zanotto, F.P. Accumulation of six metals in the mangrove crab Ucides cordatus (Crustacea: Ucididae) and its food source, the red mangrove Rhizophora mangle (Angiosperma: Rhizophoraceae). Ecotoxicol. Environ. Saf. 2012, 81, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Liu, D.; Jing, W.; Dahms, H.-U.; Wang, L. Effects of Cadmium on Lipid Storage and Metabolism in the Freshwater Crab Sinopotamon henanense. PLoS ONE 2013, 8, e77569. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, P.; Shen, Q.; Wang, Q.; Liu, D.; Li, J.; Wang, L. The Effects of Cadmium Exposure on the Oxidative State and Cell Death in the Gill of Freshwater Crab Sinopotamon henanense. PLoS ONE 2013, 8, e64020. [Google Scholar] [CrossRef]
- Ortega, P.; Custódio, M.R.; Zanotto, F.P. Characterization of cadmium transport in hepatopancreatic cells of a mangrove crab Ucides cordatus: The role of calcium. Aquat. Toxicol. 2017, 188, 92–99. [Google Scholar] [CrossRef]
- Dansa-Petretski, M.; Ribeiro, J.M.C.; Atella, G.C.; Masuda, H.; Oliveira, P.L. Antioxidant Role of Rhodnius prolixus Heme-binding Protein. J. Biol. Chem. 1995, 270, 10893–10896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maya-Monteiro, C.M.; Daffre, S.; Logullo, C.; Lara, F.A.; Alves, E.W.; Capurro, M.L.; Zingali, R.; Almeida, I.C.; Oliveira, P.L. HeLp, a heme lipoprotein from the hemolymph of the cattle tick, Boophilus microplus. J. Biol. Chem. 2000, 275, 36584–36589. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Lipid Peroxidation (Eq-H2O2 (nmol/g Wet Tissue)) | ||||
|---|---|---|---|---|
| Sex | Description | Hepatopancreas | Gill | Hemolymph |
| Female | Control | 4.62 ± 0.13 | 0.72 ± 0.56 | 0.46 ± 0.39 |
| Sample | 4.68 ± 1.60 | 2.90 ± 0.45 | 1.48 ± 1.42 | |
| Male | Control | 3.44 ± 1.27 | 1.66 ± 0.84 | 0.19 ± 0.10 |
| Sample | 5.12 ± 1.59 | 3.75 ± 0.83 | 1.28 ± 1.06 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aguilar Vitorino, H.; Pastrana Alta, R.Y.; Ortega, P. Lipid Peroxidation in Hepatopancreas, Gill, and Hemolymph of Male and Female Crabs Platyxanthus orbignyi after Air Exposure. J. Mar. Sci. Eng. 2019, 7, 347. https://doi.org/10.3390/jmse7100347
Aguilar Vitorino H, Pastrana Alta RY, Ortega P. Lipid Peroxidation in Hepatopancreas, Gill, and Hemolymph of Male and Female Crabs Platyxanthus orbignyi after Air Exposure. Journal of Marine Science and Engineering. 2019; 7(10):347. https://doi.org/10.3390/jmse7100347
Chicago/Turabian StyleAguilar Vitorino, Hector, Roxana Yesenia Pastrana Alta, and Priscila Ortega. 2019. "Lipid Peroxidation in Hepatopancreas, Gill, and Hemolymph of Male and Female Crabs Platyxanthus orbignyi after Air Exposure" Journal of Marine Science and Engineering 7, no. 10: 347. https://doi.org/10.3390/jmse7100347