Mass Spectrometry: A Rosetta Stone to Learn How Fungi Interact and Talk
by
 , , and
, , and
Erika Calla-Quispe
1 ,
,
Hammerly Lino Fuentes-Rivera
1,2,
Pablo Ramírez
2,
Carlos Martel
1,3 and
Alfredo J. Ibañez
1,*
1
Instituto de Ciencias Ómicas y Biotecnología Aplicada (ICOBA), Pontificia Universidad Católica del Perú (PUCP), Av. Universitaria 1801, San Miguel 15088, Lima, Peru
2
Laboratory of Molecular Microbiology and Biotechnology, Faculty of Biological Sciences, Universidad Nacional Mayor de San Marcos (UNMSM), Av. Germán Amézaga 375, Lima 15081, Peru
3
Museo de Historia Natural, Universidad Nacional Mayor de San Marcos (UNMSM), Av. Arenales 1256, Jesús María 15072, Lima, Peru
*
Author to whom correspondence should be addressed.
Life 2020, 10(6), 89; https://doi.org/10.3390/life10060089
Submission received: 30 May 2020
/
Revised: 16 June 2020
/
Accepted: 18 June 2020
/
Published: 20 June 2020
(This article belongs to the Special Issue Advances in Fungal -Omics)
Abstract
:Fungi are a highly diverse group of heterotrophic organisms that play an important role in diverse ecological interactions, many of which are chemically mediated. Fungi have a very versatile metabolism, which allows them to synthesize a large number of still little-known chemical compounds, such as soluble compounds that are secreted into the medium and volatile compounds that are chemical mediators over short and long distances. Mass spectrometry (MS) is currently playing a dominant role in mycological studies, mainly due to its inherent sensitivity and rapid identification capabilities of different metabolites. Furthermore, MS has also been used as a reliable and accurate tool for fungi identification (i.e., biotyping). Here, we introduce the readers about fungal specialized metabolites, their role in ecological interactions and provide an overview on the MS-based techniques used in fungal studies. We particularly present the importance of sampling techniques, strategies to reduce false-positive identification and new MS-based analytical strategies that can be used in mycological studies, further expanding the use of MS in broader applications. Therefore, we foresee a bright future for mass spectrometry-based research in the field of mycology.
1. Introduction
Fungi comprise a monophyletic yet unresolved kingdom. At the moment, it is still not possible to diagnose fungi based on specific characters and they should be regarded as a group without synapomorphies [1]. Nevertheless, it is clear that the evolution of both osmotrophic and rigid cells, which are widespread among fungi, are among the evolutionary developments that allow their diversification and ecological success [1]. It is estimated that 1.5 to 3.8 million fungal species occur on Earth, despite only around 100,000 species being known to science [2,3,4,5]. This indicates that fungi are far more diverse than other kingdoms, and a perspective research area. Fungi have been very successful in inhabiting the world, and they can be found in terrestrial and aquatic environments, as symbiotic associations as well as saprophytic or parasitic organisms. Fungal ecological success is also a consequence of their huge metabolic machinery that allows them to produce diverse chemical compounds, which are important for their ecological interactions and survival [6,7,8,9].
Fungi produce a large diversity of secondary metabolites (hereafter referred as specialized metabolites), from which more than 15,600 are unique to them [10]. The production of specialized metabolites by fungi is an evolutionary advantage, as they use metabolites to defend themselves, takedown defenses, as well as to takedown defenses of their potential food sources, and for intra-/interspecific chemical communication [9,11,12,13]. For example, fungi communicate and interact within and with other species either via volatile organic compounds (VOCs) or non-volatile secreted compounds [14,15]. Most of these compounds have been identified using mass spectrometry (hereafter MS). Therefore, MS is a powerful tool in understanding fungi metabolism. Fungal metabolites are being continuously discovered and the rate of these discoveries has been accelerated significantly during the last two decades. This is thanks to the development of MS, which has been used in mycological studies such as biotyping, ecology and monitoring the production of predominantly specialized metabolites. Indeed, in a search within the Web of Science portal, we found that the number and pattern of scientific publications per year involving fungal metabolites are almost identical to the number and pattern of mycological studies involving MS techniques (Figure 1). The development of MS instruments and techniques during the last three decades has also stimulated the increasing number of publications related to fungal metabolites, which heavily increased from around 750 released publications in 1985 up to around 16,000 in 2019 (Figure 1).
The inherent sensitivity and rapid data acquisition capabilities of MS instruments, in addition to new MS-based analytical techniques, have thus prompted the further development of metabolomic studies on fungi. However, the use of cutting-edge MS instruments and techniques alone is not enough, if there are not clear analytical protocols that allow the researches to correctly address and achieve their study goals. Thereby, we believe that it is necessary and of current interest for a combination of sampling and analytical techniques used in MS-based fungal metabolomic studies. Our goal in this review is to show the versatility and potential use of MS-based techniques in mycological studies by providing information on the diverse metabolites known from fungi, their ecological role and the MS-based techniques used.
2. Fungi and Their Specialized Metabolites
2.1. Taxonomical Fungal Groups and Their Specialized Metabolites
2.1.1. Lower Fungi
Lower fungi are an unnatural group that comprises diverse phyla of basal fungi [16]. Two unnatural groups are found here: zoosporic and zygomycetous fungi. Zoosporic fungi are the most basal ones and are characterized by producing zoospores that bear a single flagellum, so they are able to move, whereas zygomycetous fungi are characterized by developing a zygospore by fusion of specialized hyphae during sexual reproduction and the absence of septate hyphae [17]. Zoosporic fungi embrace diverse phyla such as Chytridiomycota, Blastocladiomycota, Neocallimastigomycota and the Opisthosporidia group, whereas zygomycetous fungi embrace the phyla Glomeromycota, Mucoromycota and Zoopagomycota [16,18]. Lower fungi are highly diverse in regard to their nutrition and lifestyle from heterotrophs and saprobes to symbiotes and parasites, and they can be found in terrestrial and aquatic environments [16,19,20]. Members of lower fungi are not well represented in mycological studies compared with those of higher fungi, and they have been believed to be incapable of producing specialized metabolites [21] because some previously reported compounds have been found out to be actually produced by endosymbiotic bacteria (e.g., rhizoxins and rhizonins; [22,23]) and some screened lower fungi did not have enzyme-encoding genes, which are key in specialized metabolites production (e.g., polyketide synthases, nonribosomal peptides synthetases, tryptophan synthases, and dimethylallyl tryptophan synthase; [24]). However, recent studies involving clusters of orthologous groups of proteins would indicate that zoosporic and zygomycetous fungi produce specialized metabolites, but their capacity is underdeveloped compared with higher fungi [10]. In fact, sterol compounds [25,26] have been recorded for them and zygomycetous fungi are known for their diversity of carotenoids [21,27]. Moreover, the identification of VOCs in some Mucor species (phylum Mucoromycota) also confirmed that they release volatile compounds [28,29,30]. Therefore, it is likely that many other lower fungi produce some specialized metabolites, and in particular VOCs, but they have not been examined or the appropriate sampling technique has not been used (e.g., extracts are not optimal for volatile collection, but instead headspace sampling; see below).
2.1.2. Higher Fungi
Higher fungi or Dikarya contain the largest number of fungi described to date [16,31]. They were historically divided into two phyla (i.e., Ascomycota and Basidiomycota), although a third phylum (i.e., the Enthorrizomycota) was recently introduced [32]. Dikarya are mainly characterized by the absence of flagella on their gametes and the fact that their cells bear two genetically distinct haploid nuclei [33]. Members of Dikarya have evolved and developed very diverse lifestyles: some fungi evolved symbiotic associations with cyanobacteria (i.e., lichens) and with insects [34,35], others closely interact with plants, insects and bacteria [36,37,38]. Some species of this group biosynthesize compounds with biological activity against other fungi, bacteria and harmful insects and nematodes [39]. Due to well-developed machinery to synthesize specialized metabolites, higher fungi produce and store a variety of chemical compounds in them, e.g., mycelia and fruit bodies, to be used in diverse ecological interactions [40]. In addition, it is necessary to add that most of the specialized metabolites known from fungal origin belong to Ascomycota and Basidiomycota [9,10,11,41].
2.2. Classes and Ecological Role of Fungal Specialized Metabolites
2.2.1. Classes of the Specialized Metabolites of Fungi
Fungi produce an array of specialized metabolites. To this date, more than 15,600 specialized metabolites are reported [10]; however, although new specialized metabolites are being constantly described, their number seems to be still far from the real fungal chemical diversity. Less attention has been directed to fungal VOCs and, only during the last two decades, the number of studies on fungal VOCs has been greatly increased [42,43,44] and over 500 newly discovered volatile compounds from 340 fungi species have been described [45]. The increasing knowledge related to fungal specialized metabolites is driven by their importance for industrial and non-industrial activities such as pharmacy and agriculture, among others (see [46]). Fungal specialized metabolites mainly contain C, H, O and N atoms, but may also include S, P, Cl, Br and F [11]; their chemical structure commonly contains polar functional groups such as hydroxyl, carboxyl, carbonyl and amino, among others. [11,41]. A variety of enzymes are involved in the synthesis of specialized metabolites in fungi, among others, such as nonribosomal peptide synthetases, polyketide synthases and terpene synthases [10,12,47,48]. Here, we provide information on the main fungal specialized metabolites such as alkaloids, nonribosomal peptides, polyketides, shikimic acid derived compounds and terpenoids.
Nonribosomal Peptides
These specialized metabolites are synthesized by the group of enzymes known as nonribosomal peptide synthetases. Nonribosomal peptides are structurally diverse as they are formed by proteinogenic and non-proteinogenic amino acids, which can be connected in a linear or cyclic fashion, and be further modified by tailoring enzymes [49,50,51]. Nevertheless, they have mostly macrocyclic structures, which can be carried out by cyclization of the N-terminal amino group and the carboxy terminus of the peptide chain [49]. Their diverse chemical features address the broad spectrum of biological activities in which they are involved [49,51]. Functions of nonribosomal peptides reach from immunosuppressive and cytostatic to antibiotic and toxic [49], and therefore some are used as pharmaceutical products such as the well-known penicillin, cephalosporin and cyclosporin.
Polyketides
Polyketides are the largest and most diverse group of fungal specialized metabolites [47]. They are derived of short-chain carboxylic acids, such as acetate and malonate, and are biosynthesized by large iterative multifunctional polyketide synthases [10,52,53]. The diversity found in polyketide structures is achieved by the action of these enzymes that can modify chain lengths, reduce methylate in different position and modify the polyketide backbone, in addition to tailoring enzymes that can further rearrange and modify poliketide structures [53]. Thus, polyketides can be as simple as monocyclic to complex polycyclic aromatic compounds [52]. Polyketides play an important role in promoting survival advantages of the host organisms and often have antibiotic and biocidal properties [52,53]. Especially toxic are the powerful mycotoxins such as citrinin, ochratoxins and the well-known aflatoxin, which are produced by Aspergillus species and can lead to immunotoxicity in humans.
Shikimic Acid Derived Compounds
These specialized metabolites, as its name suggest, are derived from the shikimic acid pathway. This pathway started with the condensation of phosphoenolpyruvate and erythrose 4-phosphate to give shikimic acid, and its final product is chorismate, which is the precursor of the synthesis for the amino acids: tryptophan, L-phenylalanine and L-tyrosine [10,54]. Chorismate is also the precursor of diverse fungal specialized metabolites such as pulvinones, terphenyl quinones, macrolides and strobilurins [8]. Although the shikimic acid pathway occurs in bacteria, plant and fungi, the distinct enzymatic processes in fungi make it different from that in plants and bacteria, and therefore structures of many shikimic acid-derived metabolites are particular to fungi [10]. Shikimic acid-derived compounds can function as pigments and biocides [8].
Terpenoids
Although plants are well-known because of their terpenoid diversity, higher fungi (Ascomycota and Basidiomycota) also produce a large variety of terpenoids. Cyclic terpenoids in fungi are synthetized by enzymes of the terpene cyclase family [10,30,47]. In fungi, terpenoids can be linear and cyclic, saturated and unsaturated compounds, with skeletons of two (i.e., monoterpenes), three (i.e., sesquiterpenes), four (i.e., diterpenes) and several isoprenes (e.g., carotenoids) [10,30,41,47]. In addition, specialized metabolites from other classes (e.g., polyketides, shikimic acid-derived compounds) are sometimes incorporated to terpenoids, resulting in compounds with a mixed biosynthetic origin [10,11]. Sesquiterpenoids and diterpenoids are dominant terpenoids among fungi [10]. Terpenoids can act as attractants, repellents, toxins and pigments and they are widely used in industry as flavors, fragrances, pharmaceuticals, food additives and cosmetics.
Alkaloids
These are specialized metabolites that bear at least one N atom in a heterocyclic ring structure [11,55]. Indole alkaloids are particularly abundant in fungi, especially among Ascomycota. Most indole alkaloids are derived from L-tryptophan, which is the indole donor [56]. Among fungal indole alkaloids, ergot alkaloids synthetized by Claviceps purpurea are the best understood as its biosynthesis pathway is known [47,55]. Indole alkaloids are particularly interesting due to their diverse biological roles such as pigments and toxins [41]. For instance, ergot alkaloids are highly toxic and are used to produce lysergic acid diethylamide (LSD), but also against different degenerative diseases [55]. Therefore, fungal alkaloids have been mostly studied in search of further drugs that can be used in the pharmaceutic industry.
2.2.2. Ecological Role of Specialized Metabolites
Deterrents and Self-Protective Compounds
Fungi have evolved a variety of defense strategies in order to protect themselves against adverse abiotic (e.g., UV radiation) and biotic environmental conditions (e.g., feeding animals, infections by bacteria, other fungi and viruses) [7,13]. For instance, melanin, which is an indolic polymer, and other pigments, which are typically found in spores and hyphae, protect fungi against UV damage and antioxidant chemicals [13,57]. Fungi have developed compounds with bitter or pungent properties (e.g., some indole alkaloids) that can be also in some occasions linked to toxicity [6,40], and can deter potential predators and pathogens, or overtake the defenses of potential preys and host organisms. Indeed, it has been shown that fungivorous animals can learn to avoid fungi with pungent properties, such as the case of the opossum Didelphis virginiana, who learned to avoid fruiting bodies containing the toxin muscimol [58,59,60]. Furthermore, fungal volatile compounds can act as repellents, e.g., 1-octen-3-ol is a deterrent of the banana slug Arioliomax columbianus from consuming the mushroom Clitopilus prunulus [61]. Many fungal metabolites act as toxins, therefore they are known as mycotoxins, and further classify as aflatoxins, ochratoxins, zearalenone, trichothecenes, cytochalasins and fumonisins [40]. Although their ecological role in nature has been rarely tested, it is reasonable to think that they evolved as a mean to protect fungi against potential predators and overcome the defenses of potential preys.
Chemical Communication in Ecological Associations, and Mutualistic and Symbiotic Interactions
Fungi evolved diverse associations with fungal and non-fungal organisms. Fungi communicate and interact either via VOCs or secreted substances [14,15]. Examples of metabolic exchange include cell surface recognition, quorum-sensing, biofilm formation and secretion of antibiotics, among others [62,63,64]. Each of these types of interaction plays a vital part in the fungi metabolic exchange and provides the basis for their survival. VOCs also play a major role in intraspecific interactions as they trigger certain behaviors such as attraction between gametes for mating in sexual reproduction, sporulation and induction of morphological changes. For instance, in ascomycetes and basidiomycetes, there are metabolites that have a function of mating pheromones, which can allow the discrimination among fungi genotypes [65,66]. Some fungi attract animals to facilitate the dispersion of their spores; this attraction is usually achieved through individual VOCs such as dimethyl sulfide that is released by, e.g., truffles (i.e., Tuber spp.) to attract mammals and insects [67,68], or mimicking complete blends of VOCs such as the parasitic Puccinia monoica, that mimics the scent of co-occurring plants [69]. Both VOCs and non-VOCs also participate in the communication within strong mutualistic associations such as plants and mycorrhizal fungi. Metabolites of mycorrhizal fungi are able to trigger plant defense responses [40,70]. Fungi also seem to have a predominant role in the ecological interactions of lichens, symbiotic associations of fungi and algae organisms, as most of the lichen specialized metabolites are from fungal origin. These lichen metabolites (e.g., atranorin, usnic acid and vulpinic acid) may act as allelopathic compounds, repellents of potential herbivores and protective against radiation, among others [71,72].
3. Common Mass Spectrometry-Based Strategies Used in Mycology Studies
Mass spectrometry tools enable the analysis of multiple molecules from a complex sample; those identified chemical compounds can be then correlated to a relevant phenomenon or biological question. Thus, applications of MS in mycological studies are not restricted to the identification of chemical compounds, but it is also used to perform fungi identification and classification, especially of micro-fungi. In the following two sections, we show the most common uses of MS-based studies in mycology (see also Table 1).
3.1. Biotyping: Microbial Classification and Identification
Rapid and reliable biological identification, especially for pathogens, is crucial in clinical diagnosis for prescribing the most efficient treatment. In fact, biotyping is a type of chemotaxonomy. Chemotaxonomy is a biological classification based upon the analytical measurement of chemical constituents, either as unique biomarkers or as contributors to a fingerprint profile (e.g., distinctive ratios of chemical components). Hence, biotyping is a potential alternative to genetic-based taxonomy, when there are no genome information nor genetic sequencing facilities available [64,151,152]. Mass spectrometry has been routinely used in chemotaxonomic studies by utilizing MS data from known microbes as reference data (e.g., using internal standards or reliable databases) [78,153,154]. In particular, matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF MS) has emerged as a tool for the accurate, cost-effective, sensitive and robust biotyping of microorganisms in daily routine clinical microbiology [73,74,75,76,77,78,79,80,85,86,87,88,89,90,91,92].
The analysis of protein profiles via MALDI-TOF MS can help to differentiate and/or identify unknown microorganisms at the genus and even species level by matching microbial mass spectra against spectral libraries collected from known organisms. Taxonomical identification based on protein profiles has been reliably achieved at the species level in some fungi, such as Aspergillus, Candida, Cryptococcus, Galactomyces, Microsporum, Penicillium, Rhodotorula, Saccharomyces and Trichosporon, among others [73,74,75,76,77,78,79,80,81,82,83,85,86,87,88,89,90,91,92,155]. Biotyping has also been found useful in describing new fungal species [156]. However, whether MALDI-TOF MS data are appropriate for identification and discrimination below the species level (e.g., strains) is still controversial [157]. Nevertheless, new approaches and rapidly increasing data bases are in constant development to further increase the resolution of biotyping. The Biotyper (Bruker, Bremen, Germany) and SARAMIS (AnagnosTec, Potsdam-Golm, Germany) databases, which are associated to their respective Bruker Daltonics (Bruker, Bremen, Germany) and Shimadzu (Shimadzu-Biotech Corp., Kyoto, Japan) MALDI-TOF MS systems, have been widely applied in fungal identification [75,76,84,88,157,158]. Samples of undigested cell extracts are most commonly used for MALDI-TOF MS biotyping [152,153,156,159]. Independently of the sample preparation used, the MS measurement is performed in the range of 2 and 20 kDa for the identification of unique ribosomal protein profiles, which are later matched to the reference database. A high advantage of protein fingerprints is that they are constantly expressed; furthermore, protein biotyping is highly reproducible and mostly independent of the culture medium, incubation temperature and growth state [153,159].
Another strategy for MS biotyping is based on VOCs emitted by the fungi. Exploratory analysis of VOCs emitted by fungi typically involves a headspace analysis of in vitro cultures. VOCs-based biotyping has been used to identify species of Candida [93,94,98], Penicillium [123], Fusarium [124] and Rhizoctonia [125]. This type of biotyping can employ headspace absorptive extraction techniques associated with gas chromatography-time flight mass spectrometry (GC-TOF-MS) or liquid–liquid extraction methods associated with gas chromatography-mass spectrometry electron ionization (GC-MS-EI). Unlike biotyping using protein profiles, the comparison of VOCs profiles should be carried out under the same conditions to be reliable: this means that they should be sampled in similar conditions (e.g., similar growth conditions) and using the same sampling method because specialized metabolite production is sensitive to diverse factors such as interaction with other microorganisms, environmental contaminants or stress and sampling techniques [14,128,160,161].
3.2. Identification of Microbial Chemical Compounds
Sampling strategies in metabolomics and the minimization of unwanted sources of variation are highly important. The source of variation can be broadly summarized as: (i) sample collection (including environmental factors) and culture conditions, (ii) metabolite extraction or sample pre-treatment prior to analysis, (iii) parameters of the MS-based instrument and (iv) data processing and compound identification [162,163,164]. The omission or minimization of such unwanted variation can have negative impacts on the development of the study, resulting in, for example, ambiguous identification or the identification of fewer biomarkers if we use non-hyphenated MS, i.e., MS that is not coupled to an orthogonal separation system [162,165,166]. In Figure 2, we summarize the steps involved in metabolomic studies from the preparation of the fungi material through to sampling and MS analysis to data processing and interpretation.
3.2.1. Identification of VOCs of Fungal Origin
At the moment, there are many publications related to fungal volatilomics, and to the best of our knowledge, there are two public VOC databases that include volatile information of more than 10,000 fungal species [45,160,167]. In order to monitor VOCs produced by micro-fungi, it is relevant to remember that VOCs are partitioned between gas and liquid phases under optimal growth conditions. Due to the large diversity of these compounds, there is no “one” analytical protocol to study them all. Gas chromatography coupled to mass spectrometry (GC-MS) is an extremely reliable and affordable method for the identification and quantification of VOCs. In general, VOCs are collected from a headspace, trapped on an adsorptive surface or solvent extracts, followed by thermal desorption or direct injection into the GC for separation, and then the individual VOCs from complex mixtures are identified by comparisons of the mass spectra with databases, internal standards and/or chromatographic retention indices [14,99,115,128,129,168,169]. Other mass spectrometry methods that have been used for the identification of VOCs include real-time measurements, which omit the use of a chromatography technique, such as proton transfer reaction-mass spectrometry (PTR-MS) that, for example, has been used in measuring volatile emissions of Ascocoryne sacroides (100), Hypoxylon sp. [99], Muscodor albus [101] and Nodulisporium sp. [102]. Another direct MS technique is selected ion flow tube-mass spectrometry (SIFT-MS) that has been adapted for the detection of volatile traces from medically important fungi such as species of Aspergillus, Candida, Cryptococcus, Fusarium and Mucor [103,104,105]. Secondary electrospray ionization coupled to high-resolution mass spectrometry (SESI-MS) has also been used to analyze the VOCs produced during fermentation by the ale yeast Saccharomyces cerevisiae [106].
An exquisite advantage of the PTR-MS, SIFT-MS and SESI-MS online approaches is their relative simplicity [104,170,171], where small molecules can be directly detected from the volatile emissions of the samples of interest. Hence, the VOCs can be easily monitored in real-time and can continuously collect data about specific compounds [100]. However, online approaches have two major disadvantages when compared with offline methods (e.g., GC-MS; HPLC-MS): (i) a lack of resolution of isobaric compounds (i.e., compounds with identical mass-to-charge ratios), and ii) sensitivity to matrix effects [106,172]. In general, MSs that are not coupled to an orthogonal separation system may provide ambiguous identification of VOCs [162,165].
Though gas chromatography remains an excellent method for the separation of isobaric VOCs, this method is time-consuming. An alternative separation method of isobaric VOCs is ion mobility coupled to mass spectrometry (IMS). Ion mobility separation has a different principle [173,174] than the one used in GC or LC-MS; hence it is able to resolve isobaric species and provide the rapid, sensitive and broad spectrum detection of trace organic components in moderately complex gas mixtures such as those found in Aspergillus species [130].
The collection of volatile specialized metabolites can be performed by direct measurement, as in the case of PTR-MS, SIFT-MS and SESI-MS, or by using a direct collection system such as headspace techniques. Unfortunately, these collection strategies can be influenced by the volatiles that are also emitted by the cultivation media or by other sources in the environment during the analysis. These additional volatiles can distort the signal profiles that are detected on the mass spectrometer [103,162,164]. Therefore, the use of a suitable sample extraction strategy is recommended such as solid phase micro extraction (SPME), which involves a metallic fiber that has been coated with different adsorbent materials. Coating materials include divinylbenzene (DVB), polydimethylsiloxane (PDMS), carboxen (CAR), polyacrylate (PA) and carbon wide range (CWR). These coatings have been used solely or in combination (Table 1) [113,130,163]. Hence, SPME-based sampling consists of the preconcentration of the VOCs on a fiber via absorption, and the later thermal desorption (delivered) of the captured volatiles to the GC column or directly to the mass spectrometer instrument (see Figure 2) [114,128].
Volatiles can also be collected by solvent extraction or liquid–liquid extraction, when they have not yet been released to the environment and therefore not yet in the gas condition. The selection of the solvent depends mainly on the chemical class of VOCs [165,169]. For example, pentane and hexane can be used, although ethyl acetate and dichloromethane are also commonly used, as solvents for the extraction of volatile compounds of fungal species. Independently of the method used for volatile collection, it is important to consider that crucial differences in the VOC profile will emerge from biological factors, such as the different emission of VOCs due to fungi cultivation on solid or in liquid medium [15,160,163,175]. These differences in VOC profiles arise most likely due to the different cell numbers and varying diffusion of oxygen into the medium [176]. Other factors that can play a role in VOC profiling are the cultivation temperature, time points at which samples are collected and humidity, among others [177]. Furthermore, VOC profiles will depend on the air sampling collection technique used [99,115,116,117,129,165,169,178]. Air sampling collection can be classified into two categories: active and passive. Active air sampling (AAS) techniques have been considered the most accurate way to measure VOC concentrations in air. These methods exploit a pump or vacuum to force analytes present in the air to flow through a sampler. The sampler can be an SPME or another trapping surface. Passive air sampling (PAS) techniques do not use a driving force, they work based on permeation or diffusion [14,175].
3.2.2. Identification of Non-Volatile Compounds of Fungal Origin
Due to the chemical diversity of non-volatile compounds that are secreted/produced by fungi, it is practically impossible to extract quantitatively all specialized metabolites using a single sample preparation method. Consequently, sample preparation is an important aspect of specialized metabolite profiling and will, independently of the method, lead to a bias toward certain types of compounds. For example, for the extraction of non-polar components, such as lipids, polyketides and terpenoids, one may use water-immiscible solvents such as ethyl acetate or dichloromethane. Other water-immiscible solvents such as ethers, chloroform and carbon tetrachloride are less common due to their toxicity and negative environmental effects [134,179]. Additionally, pH is crucial for the extraction, due to ionizable moieties in the chemical structure of the metabolites being extracted into the organic phase to a much higher degree in their neutral form than in a charged state. In most of the cases is a low pH extraction necessary, but it can be replaced in some cases with a neutral extraction (e.g., for stability reasons) [118,165].
To extract polar components, one may use acetone, methanol and ethanol [143,144,150]. The most common solvent used to extract polar intracellular secreted compounds is methanol (see Figure 2). Nevertheless, it has been reported that methanol and ethanol extraction can yield in the extraction of non-polar waxes, sterols, and triglycerides, as well. Thus, it is necessary to perform a second extraction step, such as extraction with ethyl acetate [118,165].
A major obstacle in any sample preparation is the possibility of unwanted chemical reactions taking place, such as precipitation, complex formation and degradation, among others. To reduce this risk, fast extraction procedures are preferred. This means extraction procedures of 10 or fewer minutes [118,165,166]. One of the most common methods used to achieve the best separation of secreted specialized metabolites produced by fungi is reverse phase (C-18) chromatography since it is more than ideal to interact with polar and some non-polar metabolites [150,180,181,182]. In some cases, before injecting samples on LC, samples are needed to be purified on small solid phase extraction (SPE) columns to remove chromatography-impairing lipids and phospholipids [147,148,183].
Interestingly, performing liquid separations at low pH (by addition of formic or acetic acid) is statistically better since sharper peaks can be obtained on the chromatogram. Formic acid and acetic acid are acid additives that are compatible with mass spectrometry detection, meanwhile, at the same time, they keep acidic moieties in specialized metabolites protonated. Nevertheless, some metabolites can be affected by the addition of acidifiers or if the mass spectrometry measurement is done in the negative mode [149,184,185].
Other chromatographic techniques use two consecutive separation columns (2-dimensional liquid chromatography) or columns of different sorbent chemistry, such as ionic exchangers (see Table 1) [14,113,129,134,143]. The most common ionization techniques used in combination with 2D liquid chromatography are the ambient pressure ionization techniques, such as electrospray ionization (ESI) [134,135,137,138,140,141,142,143,144,145,150] and atmospheric pressure chemical ionization (APCI) [139,146]. These are all soft ionization techniques (i.e., they produce limited chemical fragmentation). Hence, mass spectrometry signals are extremely clean and easily interpretable.
The possibility of adducts formations as well as multiple charge metabolites can make researchers disagree with the “easily interpretable” statement [131,140,146]. However, new high-resolution mass spectrometers, which can perform tandem MS fragmentation and analysis, can improve the correct identification of the specialized metabolites by decreasing the incorrect chemical composition assignment of the ion signals. Furthermore, specialized data handling software can further decrease the risk of misassignments, since it is capable of analyzing tandem MS fragmentation signals to determine the “chemical” relation among the measured signals, or to search an unknown MS signal in a database by comparing the fragmentation patterns in the tandem MS spectra and the structural information of fungal specialized metabolites [45,139,141,186,187,188]. Today the high-resolution mass spectrometry market is mainly divided by time of flight (TOF), Orbitrap and Fourier transform ion cyclotron (FT-ICR).
Non-volatile compounds produced by fungi can be also monitored and measured in situ. This is particularly helpful, if it is not possible to gain enough fungi biomass or if it is interesting to analyze the spatial distribution of the metabolites in a culture. In this case, direct ionization sources compatible with mass spectrometry imaging (MSI), such as MALDI [107], laser electrospray ionization (LAESI) [131,189,190,191], desorption electrospray ionization (DESI) [108,109,110,111] and nano desorption electrospray ionization (nanoDESI) [110,131,192], can be directly used for examining fungal cultures such as for species of Clohesyomyces, Fusarium, Moniliophthora, Penicillium and Trichoderma.
Nevertheless, all the above-mentioned methods have limitations [110,131,193]. For example, typical surfaces observed in fungal cultures can be detrimental to DESI, since viscous biofilms can clog the spraying systems. Furthermore, MALDI- or LAESI-based MSI systems may have trouble achieving optimum ablation/ionization signals due to the lack of a constant laser focal distance to the sample surface, since fungal surfaces often possess complex topography because of the presence of aerial hyphae [109]. All MSI techniques may also suffer from misidentification in the case of isobaric species [161,173]. For several years, ion mobility instruments have been used to improve metabolite identification, in particular for MSI experiments. However, up today, we have not yet found a hybrid IMS-MS imaging experiment performed on fungi. We still believe that in the following years, better MSI instrumental setups would overcome the above-mentioned challenges.
4. Future Directions
The applications of MS in mycology are very promising. The recent developments in mass spectrometry imaging have dramatically facilitated and improved the identification of fungi species. However, to elucidate the ecological role of fungi in communities, as well as understand their ecological interactions with other group of organisms such as algae or bacteria, it is necessary to identify the metabolites they produce. Mass spectrometry has also improved the identification of fungal specialized metabolites; however, the identification reliability still needs to improve. The main challenge is still the limited number of dedicated fungi libraries and repositories containing MS data of specialized metabolites, where all chemical and taxonomic information have been merged together. As reported here, newer sample handling protocols as well as ionization sources that have been successfully applied to human, plants and animal studies have yet to migrate to mycology studies. One challenge is the wide fungi diversity as well as the lack of universal sample handling/MS protocols. Therefore, collaborative efforts may be needed to increase or enrich the MS libraries and consequently improve the reliability of the identification of specialized metabolites, as well as specific identification. Better repositories of specialized metabolites produced by fungi will then shed light on the molecular adaptations of the fungi to their environment and on their interactions with other organisms of the community. The idea is to treat the fungi and the community as a whole, integrating the fundamental biological knowledge, to obtain an understanding of every particular ecosystem for the purpose of biotechnological applications.
Author Contributions
Conceptualization, A.J.I.; methodology, E.C.-Q., C.M. and A.J.I.; writing—original draft preparation, E.C.-Q, H.L.F.-R., P.R., C.M. and A.J.I.; writing—review and editing, E.C.-Q, H.L.F.-R., P.R., C.M. and A.I.; supervision, C.M. and A.J.I.; project administration, A.J.I.; funding acquisition, A.J.I. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by “Proyecto de Mejoramiento y Ampliación de los Servicios del Sistema Nacional de Ciencia Tecnología e Innovación Tecnológica (8682-PE)”, CONCYTEC, and “The Max Planck Partner Group” (Max Planck Institute for Chemical Ecology–Jena).
Acknowledgments
We are grateful to the editors for the invitation to contribute to this special issue. E.C.-Q. was supported by a doctoral scholarship granted by “Proyecto de Mejoramiento y Ampliación de los Servicios del Sistema Nacional de Ciencia Tecnología e Innovación Tecnológica (8682-PE)” funded by the World Bank, CONCYTEC, and FONDECYT, through their Doctoral Program (010-20180). A.I., C.M., H.L.F-R. and E.C.-Q. also thank to “The Max Planck Partner Group” for their financial support. Madina Mansurova kindly reviewed and gave comments on an earlier version of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Richards, T.A.; Leonard, G.; Wideman, J.G. What defines the “kingdom” Fungi? Microbiol. Spectr. 2017, 5, FUNK-0044-2017. [Google Scholar] [CrossRef]
- Goyal, S.; Ramawat, K.G.; Mérillon, J.-M. Different shades of fungal metabolites: An overview. In Fungal Metabolites. Reference Series in Phytochemistry; Mérillon, J.M., Ramawat, K., Eds.; Springer: Cham, Switzerland, 2017; pp. 1–29. [Google Scholar] [CrossRef]
- Mueller, G.M.; Schmit, J.P. Fungal biodiversity: What do we know? What can we predict? Biodivers. Conserv. 2007, 16, 1–5. [Google Scholar] [CrossRef]
- Chethana, K.W.T.; Jayawardena, R.S.; Hyde, K.D. Hurdles in fungal taxonomy: Effectiveness of recent methods in discriminating taxa. Megataxa 2020, 1, 114–122. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, R.H.; Larsson, K.-H.; Taylor, A.F.S.; Bengtsson-Palme, J.; Jeppesen, T.S.; Schigel, D.; Kennedy, P.; Picard, K.; Glöckner, F.O.; Tedersoo, L.; et al. The UNITE database for molecular identification of fungi: Handling dark taxa and parallel taxonomic classifications. Nucleic Acids Res. 2018, 47, D259–D264. [Google Scholar] [CrossRef] [PubMed]
- Spiteller, P. Chemical defence strategies of higher fungi. Chem. A Eur. J. 2008, 14, 9100–9110. [Google Scholar] [CrossRef]
- Braga, R.M.; Dourado, M.N.; Araújo, W.L. Microbial interactions: Ecology in a molecular perspective. Braz. J. Microbiol. 2016, 47, 86–98. [Google Scholar] [CrossRef] [Green Version]
- Reck, B.; Spiteller, P. Synthesis of secondary metabolites from higher fungi. Synthesis 2015, 47, 2885–2911. [Google Scholar] [CrossRef]
- Chen, H.P.; Liu, J.K. Secondary metabolites from higher fungi. In Progress in the Chemistry of Organic Natural Products; Springer: Cham, Switzerland, 2017; Volume 106, pp. 1–201. [Google Scholar] [CrossRef]
- Bills, G.F.; Gloer, J.B. Biologically active secondary metabolites from the fungi. Microbiol. Spectr. 2016, 4, FUNK-0009-2016. [Google Scholar] [CrossRef]
- Zhong, J.J.; Xiao, J.H. Secondary metabolites from higher fungi: Discovery, bioactivity, and bioproduction. Adv. Biochem. Eng. Biotechnol. 2009, 113, 79–150. [Google Scholar] [CrossRef]
- Pusztahelyi, T.; Holb, I.J.; Pocsi, I. Secondary metabolites in fungus-plant interactions. Front. Plant Sci. 2015, 6, 573. [Google Scholar] [CrossRef] [Green Version]
- Keller, N.P. Fungal secondary metabolism: Regulation, function and drug discovery. Nat. Rev. Microbiol. 2019, 17, 167–180. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, W.M.; Geranios, P.; White, I.R.; Lawal, O.; Nijsen, T.M.; Bromley, M.J.; Goodacre, R.; Read, N.D.; Fowler, S.J. Development of an adaptable headspace sampling method for metabolic profiling of the fungal volatome. Analyst 2018, 143, 4155–4162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulz-Bohm, K.; Martín-Sánchez, L.; Garbeva, P. Microbial volatiles: Small molecules with an important role in intra- and inter-kingdom interactions. Front. Microbiol. 2017, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Naranjo-Ortiz, M.A.; Gabaldón, T. Fungal evolution: Diversity, taxonomy and phylogeny of the fungi. Biol. Rev. 2019, 94, 2101–2137. [Google Scholar] [CrossRef] [PubMed]
- Naranjo-Ortiz, M.A.; Gabaldón, T. Fungal evolution: Major ecological adaptations and evolutionary transitions. Biol. Rev. 2019, 94, 1443–1476. [Google Scholar] [CrossRef] [Green Version]
- Wijayawardene, N.N.; Pawłowska, J.; Letcher, P.M.; Kirk, P.M.; Humber, R.A.; Schüßler, A.; Wrzosek, M.; Muszewska, A.; Okrasińska, A.; Istel, Ł.; et al. Notes for genera: Basal clades of fungi (including Aphelidiomycota, Basidiobolomycota, Blastocladiomycota, Calcarisporiellomycota, Caulochytriomycota, Chytridiomycota, Entomophthoromycota, Glomeromycota, Kickxellomycota, Monoblepharomycota, Mortierellomycota, Mucoromycota, Neocallimastigomycota, Olpidiomycota, Rozellomycota and Zoopagomycota). Fungal Divers. 2018, 92, 43–129. [Google Scholar] [CrossRef]
- Karpov, S.A.; Mamkaeva, M.A.; Aleoshin, V.V.; Nassonova, E.; Lilje, O.; Gleason, F.H. Morphology, phylogeny, and ecology of the aphelids (Aphelidea, Opisthokonta) and proposal for the new superphylum Opisthosporidia. Front. Microbiol. 2014, 5, 112. [Google Scholar] [CrossRef] [Green Version]
- Powell, M.J. Chytridiomycota. In Handbook of the Protists; Archibald, J., Simpson, A., Slamovits, C., Eds.; Springer: Cham, Switzerland, 2016; pp. 1–36. [Google Scholar] [CrossRef]
- Voigt, K.; Wolf, T.; Ochsenreiter, K.; Nagy, G.; Kaerger, K.; Shelest, E.; Papp, T. Genetic and metabolic aspects of primary and secondary metabolism of the zygomycetes. In Biochemistry and Molecular Biology. The Mycota (A Comprehensive Treatise on Fungi as Experimental Systems for Basic and Applied Research); Hoffmeister, D., Ed.; Springer: Cham, Switzerland, 2016; pp. 361–385. [Google Scholar] [CrossRef]
- Partida-Martinez, L.P.; Hertweck, C. Pathogenic fungus harbours endosymbiotic bacteria for toxin production. Nature 2005, 437, 884–888. [Google Scholar] [CrossRef]
- Partida-Martinez, L.P.; de Looss, C.F.; Ishida, K.; Ishida, M.; Roth, M.; Buder, K.; Hertweck, C. Rhizonin, the first mycotoxin isolated from the Zygomycota, is not a fungal metabolite but is produced by bacterial endosymbionts. Appl. Environ. Microbiol. 2007, 73, 793–797. [Google Scholar] [CrossRef] [Green Version]
- Collemare, J.; Billard, A.; Bohnert, H.U.; Lebrun, M.H. Biosynthesis of secondary metabolites in the rice blast fungus Magnaporthe grisea: The role of hybrid PKS-NRPS in pathogenicity. Mycol. Res. 2008, 112, 207–215. [Google Scholar] [CrossRef]
- Kerwin, J.L. Lipid-mediated morphogenesis of oomycetous fungi. In Advances in Zoosporic Fungi; Dayal, R., Ed.; M. D. Publications Pvt. Ltd.: New Delhi, India, 1986; pp. 201–242. [Google Scholar]
- Weete, J.D.; Gandhi, S.R. Sterols of the phylum Zygomycota: Phylogenetic implications. Lipids 1997, 32, 309–1316. [Google Scholar] [CrossRef] [PubMed]
- Echavarri-Erasun, C.; Johnson, E.A. Fungal carotenoids. Appl. Mycol. Biotechnol. 2002, 2, 45–85. [Google Scholar] [CrossRef]
- Stotzky, G.; Schenck, S.; Papavizas, G.C. Volatile organic compounds and microorganisms. Crit. Rev. Microbiol. 1976, 4, 333–382. [Google Scholar] [CrossRef] [PubMed]
- Nieminen, T.; Neubauer, P.; Sivelā, S.; Vatamo, S.; Silfverberg, P.; Salkinoja-Salonen, M. Volatile compounds produced by fungi grown in strawberry jam. LWT Food Sci. Technol. 2008, 41, 2051–2056. [Google Scholar] [CrossRef]
- Schmidt-Dannert, C. Biosynthesis of terpenoid natural products in fungi. Adv. Biochem. Eng. Biotechnol. 2015, 148, 19–61. [Google Scholar] [CrossRef] [PubMed]
- Levetin, E.; Horner, W.E.; Scott, J.A.; Barnes, C.; Baxi, S.; Chew, G.L.; Grimes, C.; Kennedy, K.; Larenas-Linnemann, D.; Miller, J.D.; et al. Taxonomy of allergenic fungi. J. Allergy Clin. Immunol. Pract. 2016, 4, 375–385. [Google Scholar] [CrossRef]
- Riess, K.; Schön, M.E.; Ziegler, R.; Lutz, M.; Shivas, R.G.; Piątek, M.; Garnica, S. The origin and diversification of the Entorrhizales: Deep evolutionary roots but recent speciation with a phylogenetic and phenotypic split between associates of the Cyperaceae and Juncaceae. Org. Divers. Evol. 2019, 19, 13–30. [Google Scholar] [CrossRef] [Green Version]
- Wellehan, J.; Divers, S. Mycology. In Mader’s Reptile and Amphibian Medicine and Surgery; Stahl, S.J., Divers, S.J., Eds.; W.B. Saunders: St. Louis, MO, USA, 2019; pp. 270–280. [Google Scholar] [CrossRef]
- Li, Y.; Simmons, D.R.; Bateman, C.C.; Short, D.P.; Kasson, M.T.; Rabaglia, R.J.; Hulcr, J. New fungus-insect symbiosis: Culturing, molecular, and histological methods determine saprophytic polyporales mutualists of Ambrosiodmus ambrosia beetles. PLoS ONE 2015, 10, e0137689. [Google Scholar] [CrossRef] [Green Version]
- Klepzig, K.; Six, D. Bark beetle-fungal symbiosis: Context dependency in complex associations. Symbiosis 2004, 37, 189–205. [Google Scholar]
- Bauer, R.; Garnica, S.; Oberwinkler, F.; Riess, K.; Weiss, M.; Begerow, D. Entorrhizomycota: A new fungal phylum reveals new perspectives on the evolution of fungi. PLoS ONE 2015, 10, e0128183. [Google Scholar] [CrossRef] [Green Version]
- Jofré, N.; Pildain, M.B.; Cirigliano, A.M.; Cabrera, G.M.; Corley, J.C.; Martínez, A.S. Host selection by Ibalia leucospoides based on temporal variations of volatiles from the hosts’ fungal symbiont. J. Appl. Entomol. 2016, 140, 736–743. [Google Scholar] [CrossRef]
- Frey-Klett, P.; Burlinson, P.; Deveau, A.; Barret, M.; Tarkka, M.; Sarniguet, A. Bacterial-fungal interactions: Hyphens between agricultural, clinical, environmental, and food microbiologists. Microbiol. Mol. Biol. Rev. 2011, 75, 583–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subramaniyan, S.; Khusro, A.; Gabriel Paulraj, M.; Ignacimuthu, S.; Al-Dhabi, N. Biocontrol properties of basidiomycetes: An overview. J. Fungi 2017, 3, 2. [Google Scholar] [CrossRef] [Green Version]
- Spiteller, P. Chemical ecology of fungi. Nat Prod Rep. 2015, 32, 971–993. [Google Scholar] [CrossRef]
- Hanson, J. The Chemistry of Fungi; The Royal Society of Chemistry: Cambridge, UK, 2008. [Google Scholar] [CrossRef] [Green Version]
- Effmert, U.; Kalderas, J.; Warnke, R.; Piechulla, B. Volatile mediated interactions between bacteria and fungi in the soil. J. Chem. Ecol. 2012, 38, 665–703. [Google Scholar] [CrossRef]
- Morath, S.U.; Hung, R.; Bennett, J.W. Fungal volatile organic compounds: A review with emphasis on their biotechnological potential. Fungal Biol. Rev. 2012, 26, 73–83. [Google Scholar] [CrossRef]
- Lemfack, M.C.; Nickel, J.; Dunkel, M.; Preissner, R.; Piechulla, B. mVOC: A database of microbial volatiles. Nucleic Acids Res. 2014, 42, D744–D748. [Google Scholar] [CrossRef]
- Lemfack, M.C.; Gohlke, B.O.; Toguem, S.M.T.; Preissner, S.; Piechulla, B.; Preissner, R. mVOC 2.0: A database of microbial volatiles. Nucleic Acids Res. 2017, 46, D1261–D1265. [Google Scholar] [CrossRef] [Green Version]
- Hyde, K.D.; Xu, J.; Rapior, S.; Rapior, S.; Jeewon, R.; Lumyong, S.; Niego, A.G.T.; Abeywickrama, P.D.; Aluthmuhandiram, J.V.S.; Brahamanage, R.S.; et al. The amazing potential of fungi: 50 ways we can exploit fungi industrially. Fungal Divers. 2019, 97, 1–136. [Google Scholar] [CrossRef] [Green Version]
- Keller, N.P.; Turner, G.; Bennett, J.W. Fungal secondary metabolism - from biochemistry to genomics. Nat. Rev. Microbiol. 2005, 3, 937–947. [Google Scholar] [CrossRef]
- Hoffmeister, D.; Keller, N.P. Natural products of filamentous fungi: Enzymes, genes, and their regulation. Nat. Prod. Rep. 2007, 24, 393–416. [Google Scholar] [CrossRef] [PubMed]
- Schwarzer, D.; Finking, R.; Marahiel, M.A. Nonribosomal peptides: From genes to products. Nat. Prod. Rep. 2003, 20, 275–287. [Google Scholar] [CrossRef] [PubMed]
- Walsh, C.T.; Chen, H.; Keating, T.A.; Hubbard, B.K.; Losey, H.C.; Luo, L.; Marshall, C.G.; Miller, D.A.; Patel, H.M. Tailoring enzymes that modify nonribosomal peptides during and after chain elongation on NRPS assembly lines. Curr. Opin. Chem. Biol. 2001, 5, 525–534. [Google Scholar] [CrossRef]
- Soukup, A.A.; Keller, N.P.; Wiemann, P. Enhancing nonribosomal peptide biosynthesis in filamentous fungi. Methods Mol. Biol. 2016, 1401, 149–160. [Google Scholar] [CrossRef] [Green Version]
- Simpson, T.J.; Cox, R.J. Polyketides in fungi. In Natural Products in Chemical Biology; Civjan, N., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2012; pp. 143–161. [Google Scholar] [CrossRef]
- Cox, R.J.; Skellam, E.; Williams, K. Biosynthesis of fungal polyketides. In Physiology and Genetics. The Mycota (A Comprehensive Treatise on Fungi as Experimental Systems for Basic and Applied Research); Anke, T., Schüffler, A., Eds.; Springer: Cham, Switzerland, 2018; pp. 385–412. [Google Scholar] [CrossRef]
- Tzin, V.; Galili, G.; Aharoni, A. Shikimate pathway and aromatic amino acid biosynthesis. In eLS; John Wiley & Sons, Ltd.: Chichester, UK, 2012. [Google Scholar] [CrossRef]
- Aniszewski, T. Alkaloids—Secrets of Life; Elsevier: Amsterdam, The Netherlands, 2007. [Google Scholar] [CrossRef]
- Xu, W.; Gavia, D.J.; Tang, Y. Biosynthesis of fungal indole alkaloids. Nat. Prod. Rep. 2014, 31, 1474–1487. [Google Scholar] [CrossRef] [Green Version]
- Eisenman, H.C.; Casadevall, A. Synthesis and assembly of fungal melanin. Appl. Microbiol. Biotechnol. 2012, 93, 931–940. [Google Scholar] [CrossRef] [Green Version]
- Camazine, S. Olfactory aposematism: Association of food toxicity with naturally occurring odor. J. Chem. Ecol. 1985, 11, 1289–1295. [Google Scholar] [CrossRef]
- Camazine, S. Mushroom chemical defense: Food aversion learning induced by hallucinogenic toxin, muscimol. J. Chem. Ecol. 1983, 9, 1473–1481. [Google Scholar] [CrossRef]
- Camazine, S.M.; Resch, J.F.; Eisner, T.; Meinwald, J. Mushroom chemical defense: Pungent sesquiterpenoid dialdehyde antifeedant to opossum. J. Chem. Ecol. 1983, 9, 1439–1447. [Google Scholar] [CrossRef]
- Wood, W.F.; Archer, C.L.; Largent, D.L. 1-Octen-3-ol, a banana slug antifeedant from mushrooms. Biochem. Syst. Ecol. 2001, 29, 531–533. [Google Scholar] [CrossRef]
- Flemming, H.C.; Wingender, J. The biofilm matrix. Nat. Ver. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Phelan, V.V.; Liu, W.-T.; Pogliano, K.; Dorrestein, P.C. Microbial metabolic exchange—The chemotype-to-phenotype link. Nat. Chem. Biol. 2012, 8, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Luzzatto-Knaan, T.; Melnik, A.V.; Dorrestein, P.C. Mass spectrometry tools and workflows for revealing microbial chemistry. Analyst 2015, 140, 4949–4966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michaelis, S.; Barrowman, J. Biogenesis of the Saccharomyces cerevisiae pheromone a-factor, from yeast mating to human disease. Microbiol. Mol. Biol. Rev. 2012, 76, 626–651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coelho, M.A.; Bakkeren, G.; Sun, S.; Hood, M.E.; Giraud, T. Fungal sex: The Basidiomycota. Microbiol. Spectr. 2017, 5, FUNK-0046-2016. [Google Scholar] [CrossRef]
- Talou, T.; Gaset, A.; Delmas, M.; Kulifaj, M.; Montant, C. Dimethyl sulphide: The secret for black truffle hunting by animals? Mycol. Res. 1990, 94, 277–278. [Google Scholar] [CrossRef]
- Vahdatzadeh, M.; Deveau, A.; Splivallo, R. The role of the microbiome of truffles in aroma formation: A meta-analysis approach. Appl. Environ. Microbiol. 2015, 81, 6946–6952. [Google Scholar] [CrossRef] [Green Version]
- Roy, B.A.; Raguso, R.A. Olfactory versus visual cues in a floral mimicry system. Oecologia 1997, 109, 414–426. [Google Scholar] [CrossRef]
- Jung, S.C.; Martinez-Medina, A.; Lopez-Raez, J.A.; Pozo, M.J. Mycorrhiza-induced resistance and priming of plant defenses. J. Chem. Ecol. 2012, 38, 651–664. [Google Scholar] [CrossRef]
- Calcott, M.J.; Ackerley, D.F.; Knight, A.; Keyzers, R.A.; Owen, J.G. Secondary metabolism in the lichen symbiosis. Chem. Soc. Rev. 2018, 47, 1730–1760. [Google Scholar] [CrossRef]
- Solhaug, K.A.; Gauslaa, Y. Secondary lichen compounds as protection against excess solar radiation and herbivores. In Progress in Botany 73; Lüttge, U., Beyschlag, W., Büdel, B., Francis, D., Eds.; Springer: Berlin, Germany, 2012; pp. 283–304. [Google Scholar] [CrossRef]
- Marklein, G.; Josten, M.; Klanke, U.; Muller, E.; Horre, R.; Maier, T.; Wenzel, T.; Kostrzewa, M.; Bierbaum, G.; Hoerauf, A.; et al. Matrix-assisted laser desorption ionization-time of flight mass spectrometry for fast and reliable identification of clinical yeast isolates. J. Clin. Microbiol. 2009, 47, 2912–2917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stevenson, L.G.; Drake, S.K.; Shea, Y.R.; Zelazny, A.M.; Murray, P.R. Evaluation of matrix-assisted laser desorption ionization-time-of-flight mass spectrometry for identification of clinically important yeast species. J. Clin. Microbiol. 2010, 48, 3482–3486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenvinge, F.S.; Dzajic, E.; Knudsen, E.; Malig, S.; Andersen, L.B.; Løvig, A.; Arendrup, M.C.; Jensen, T.G.; Gahrn-Hansen, B.; Kemp, M. Performance of matrix-assisted laser desorption-time of flight mass spectrometry for identification of clinical yeast isolates. Mycoses 2013, 56, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Lohmann, C.; Sabou, M.; Moussaoui, W.; Lohmann, C.; Sabou, M.; Moussaoui, W.; Prévost, G.; Delarbre, J.-M.; Candolfi, E.; Gravet, A.; et al. Comparison between the biflex III-biotyper and the Axima-SARAMIS systems for yeast identification by matrix-assisted laser desorption ionization-time of flight mass spectrometry. J. Clin. Microbiol. 2013, 51, 1231–1236. [Google Scholar] [CrossRef] [Green Version]
- Reich, M.; Bosshard, P.P.; Stark, M.; Beyser, K.; Borgmann, S. Species identification of bacteria and fungi from solid and liquid culture media by MALDI-TOF mass spectrometry. J. Bacteriol. Parasitol. 2013, S5, 2. [Google Scholar] [CrossRef] [Green Version]
- Lau, A.F.; Drake, S.K.; Calhoun, L.B.; Henderson, C.M.; Zelazny, A.M. Development of a clinically comprehensive database and a simple procedure for identification of molds from solid media by matrix-assisted laser desorption ionization-time of flight mass spectrometry. J. Clin. Microbiol. 2013, 51, 828–834. [Google Scholar] [CrossRef] [Green Version]
- Karabıçak, N.; Karatuna, O.; İlkit, M.; Akyar, I. Evaluation of the Bruker matrix-assisted laser desorption–ionization time-of-flight mass spectrometry (MALDI-TOF MS) system for the identification of clinically important Dermatophyte species. Mycopathologia 2015, 180, 165–171. [Google Scholar] [CrossRef]
- L’Ollivier, C.; Ranque, S. MALDI-TOF-based dermatophyte identification. Mycopathologia 2017, 182, 183–192. [Google Scholar] [CrossRef] [Green Version]
- Moothoo-Padayachie, A.; Kandappa, H.R.; Krishna, S.B.N.; Maier, T.; Govender, P. Biotyping Saccharomyces cerevisiae strains using matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF MS). Eur. Food Res. Technol. 2013, 236, 351–364. [Google Scholar] [CrossRef]
- Normand, A.C.; Becker, P.; Gabriel, F.; Cassagne, C.; Accoceberry, I.; Gari-Toussaint, M.; Hasseine, L.; De Geyter, D.; Pierard, D.; Surmont, I.; et al. Validation of a new web application for identification of fungi by use of matrix-assisted laser desorption ionization–time of flight mass spectrometry. J. Clin. Microbiol. 2017, 55, 2661–2670. [Google Scholar] [CrossRef] [Green Version]
- Chalupová, J.; Raus, M.; Sedlářová, M.; Šebela, M. Identification of fungal microorganisms by MALDI-TOF mass spectrometry. Biotechnol. Adv. 2014, 32, 230–241. [Google Scholar] [CrossRef] [PubMed]
- Stein, M.; Tran, V.; Nichol, K.A.; Lagacé-Wiens, P.; Pieroni, P.; Adam, H.J.; Turenne, C.; Walkty, A.J.; Normand, A.-C.; Hendrickx, M.; et al. Evaluation of three MALDI-TOF mass spectrometry libraries for the identification of filamentous fungi in three clinical microbiology laboratories in Manitoba, Canada. Mycoses 2018, 61, 743–753. [Google Scholar] [CrossRef] [PubMed]
- Van Veen, S.Q.; Claas, E.C.J.; Kuijper, E.J. High-throughput identification of bacteria and yeast by matrix-assisted laser desorption ionization-time of flight mass spectrometry in conventional medical microbiology laboratories. J. Clin. Microbiol. 2010, 48, 900–907. [Google Scholar] [CrossRef] [Green Version]
- Dhiman, N.; Hall, L.; Wohlfiel, S.L.; Buckwalter, S.P.; Wengenack, N.L. Performance and cost analysis of matrix-assisted laser desorption ionization-time of flight mass spectrometry for routine identification of yeast. J. Clin. Microbiol. 2011, 49, 1614–1616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, C.; Lima, N.; Sampaio, P.; Pais, C. Matrix-assisted laser desorption/ionization time-of-flight intact cell mass spectrometry to detect emerging pathogenic Candida species. Diagn. Microbiol. Infect. Dis. 2011, 71, 304–308. [Google Scholar] [CrossRef] [Green Version]
- Bader, O.; Weig, M.; Taverne-Ghadwal, L.; Lugert, R.; Groß, U.; Kuhns, M. Improved clinical laboratory identification of human pathogenic yeasts by matrix-assisted laser desorption ionization time-of-flight mass spectrometry. Clin. Microbiol. Infect. 2011, 17, 1359–1365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinto, A.; Halliday, C.; Zahra, M.; Van Hal, S.; Olma, T.; Maszewska, K.; Iredell, J.R.; Meyer, W.; Chen, S.C.-A. Matrix-assisted laser desorption ionization-time of flight mass spectrometry identification of yeasts is contingent on robust reference spectra. PLoS ONE 2011, 6, e25712. [Google Scholar] [CrossRef]
- Pan, Y.L.; Chow, N.H.; Chang, T.C.; Chang, H.C. Identification of lethal Aspergillus at early growth stages based on matrix-assisted laser desorption/ionization time-of-flight mass spectrometry. Diagn. Microbiol. Infect. Dis. 2011, 70, 344–354. [Google Scholar] [CrossRef]
- Firacative, C.; Trilles, L.; Meyer, W. MALDI-TOF MS enables the rapid identification of the major molecular types within the Cryptococcus neoformans/C. gattii species complex. PLoS ONE 2012, 7, e37566. [Google Scholar] [CrossRef] [Green Version]
- Goyer, M.; Lucchi, G.; Ducoroy, P.; Vagner, O.; Bonnin, A.; Dalle, F. Optimization of the preanalytical steps of matrix-assisted laser desorption ionization-time of flight mass spectrometry identification provides a flexible and efficient tool for identification of clinical yeast isolates in medical laboratories. J. Clin. Microbiol. 2012, 50, 3066–3068. [Google Scholar] [CrossRef] [Green Version]
- Cameron, S.J.S.; Bolt, F.; Perdones-Montero, A.; Rickards, T.; Hardiman, K.; Abdolrasouli, A.; Burke, A.; Bodai, Z.; Karancsi, T.; Simon, D.; et al. Rapid evaporative ionisation mass spectrometry (REIMS) provides accurate direct from culture species identification within the genus Candida. Sci. Rep. 2016, 6, 36788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolt, F.; Cameron, S.J.S.; Karancsi, T.; Simon, D.; Schaffer, R.; Rickards, T.; Hardiman, K.; Burke, A.; Bodai, Z.; Perdones-Montero, A.; et al. Automated high-throughput identification and characterization of clinically important bacteria and fungi using rapid evaporative ionization mass spectrometry. Anal. Chem. 2016, 88, 9419–9426. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.J.; Chen, Y.P.; Wang, H.C. Identification of fungal pathogens from clinical specimens using multi-locus PCR coupled with electrospray ionization mass spectrometry. Diagn. Microbiol. Infect. Dis. 2014, 78, 141–143. [Google Scholar] [CrossRef]
- Jia, B.; Lovari, R.; Miller, H.; Metzgar, D.; Massire, C.; Carolan, H.; Toleno, D.; D’Alessio, F.; Rothman, R.; Blyn, L.B.; et al. Evaluation of a PCR-electrospray ionization mass spectrometry platform for detection and identification of fungal pathogens directly from prospectively collected bronchoalveolar lavage specimens. Diagn. Microbiol. Infect. Dis. 2020, 97, 114988. [Google Scholar] [CrossRef] [PubMed]
- Simner, P.J.; Uhl, J.R.; Hall, L.; Weber, M.M.; Walchak, R.C.; Buckwalter, S.; Wengenacke, N.L. Broad-range direct detection and identification of fungi by use of the PLEX-ID PCR-electrospray ionization mass spectrometry (ESI-MS) system. J. Clin. Microbiol. 2013, 51, 1699–1706. [Google Scholar] [CrossRef] [Green Version]
- Hamid, A.M.; Wei, P.; Jarmusch, A.K.; Pirro, V.; Cooks, R.G. Discrimination of Candida species by paper spray mass spectrometry. Int. J. Mass Spectrom. 2015, 378, 288–293. [Google Scholar] [CrossRef]
- Booth, E.; Strobel, G.; Knighton, B.; Sears, J.; Geary, B.; Avci, R. A rapid column technique for trapping and collecting of volatile fungal hydrocarbons and hydrocarbon derivatives. Biotechnol. Lett. 2011, 33, 1963–1972. [Google Scholar] [CrossRef]
- Mallette, N.D.; Knighton, W.B.; Strobel, G.A.; Carlson, R.P.; Peyton, B.M. Resolution of volatile fuel compound profiles from Ascocoryne sarcoides: A comparison by proton transfer reaction-mass spectrometry and solid phase microextraction gas chromatography mass spectrometry. AMB Expr. 2012, 2, 23. [Google Scholar] [CrossRef] [Green Version]
- Ezra, D.; Jasper, J.; Rogers, T.; Knighton, B.; Grimsrud, E.; Strobel, G. Proton transfer reaction-mass spectrometry as a technique to measure volatile emissions of Muscodor albus. Plant Sci. 2004, 166, 1471–1477. [Google Scholar] [CrossRef]
- Schoen, H.R.; Knighton, W.B.; Peyton, B.M. Endophytic fungal production rates of volatile organic compounds are highest under microaerophilic conditions. Microbiology 2017, 163, 1767–1777. [Google Scholar] [CrossRef]
- Scotter, J.M.; Langford, V.S.; Wilson, P.F.; McEwan, M.J.; Chambers, S.T. Real-time detection of common microbial volatile organic compounds from medically important fungi by selected ion flow tube-mass spectrometry (SIFT-MS). J. Microbiol. Methods 2005, 63, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.; Španěl, P. SIFT-MS and FA-MS methods for ambient gas phase analysis: Developments and applications in the UK. Analyst 2015, 140, 2573–2591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chippendale, T.W.E.; Gilchrist, F.J.; Španěl, P.; Alcock, A.; Lenney, W.; Smith, D. Quantification by SIFT-MS of volatile compounds emitted by Aspergillus fumigatus cultures and in co-culture with Pseudomonas aeruginosa, Staphylococcus aureus and Streptococcus pneumoniae. Anal. Methods 2014, 6, 8154–8164. [Google Scholar] [CrossRef]
- Tejero Rioseras, A.; Garcia Gomez, D.; Ebert, B.E.; Blank, L.M.; Ibáñez, A.J.; Sinues, P.M.L. Comprehensive real-time analysis of the yeast volatilome. Sci. Rep. 2017, 7, 14236. [Google Scholar] [CrossRef]
- Le Pogam, P.; Schinkovitz, A.; Legouin, B.; Le Lamer, A.C.; Boustie, J.; Richomme, P. Matrix-free UV-laser desorption ionization mass spectrometry as a versatile approach for accelerating dereplication studies on lichens. Anal. Chem. 2015, 87, 10421–10428. [Google Scholar] [CrossRef] [Green Version]
- Sica, V.P.; Raja, H.A.; El-Elimat, T.; Oberlies, N.H. Mass spectrometry imaging of secondary metabolites directly on fungal cultures. RSC Adv. 2014, 4, 63221–63227. [Google Scholar] [CrossRef] [Green Version]
- Tata, A.; Perez, C.J.; Ore, M.O.; Lostun, D.; Passas, A.; Morin, S.; Ifa, D.R. Evaluation of imprint DESI-MS substrates for the analysis of fungal metabolites. RSC Adv. 2015, 5, 75458–75464. [Google Scholar] [CrossRef]
- Parrot, D.; Papazian, S.; Foil, D.; Tasdemir, D. Imaging the unimaginable: Desorption electrospray ionization—Imaging mass spectrometry (DESI-IMS) in natural product research. Planta Med. 2018, 84, 584–593. [Google Scholar] [CrossRef] [Green Version]
- Figueroa, M.; Jarmusch, A.K.; Raja, H.A.; El-Elimat, T.; Kavanaugh, J.S.; Horswill, A.R.; Cooks, R.G.; Cech, N.B.; Oberlies, N.H. Polyhydroxyanthraquinones as quorum sensing inhibitors from the guttates of Penicillium restrictum and their analysis by desorption electrospray ionization mass spectrometry. J. Nat. Prod. 2014, 77, 1351–1358. [Google Scholar] [CrossRef] [Green Version]
- Larsen, T.; Taylor, D.L.; Leigh, M.B.; O’Brien, D.M. Stable isotope fingerprinting: A novel method for identifying plant, fungal, or bacterial origins of amino acids. Ecology 2009, 90, 3526–3535. [Google Scholar] [CrossRef] [Green Version]
- Erler, A.; Riebe, D.; Beitz, T.; Löhmannsröben, H.-G.; Grothusheitkamp, D.; Kunz, T.; Methner, F.-J. Characterization of volatile metabolites formed by molds on barley by mass and ion mobility spectrometry. J. Mass Spectrom. 2020, 55, e4501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kluger, B.; Zeilinger, S.; Wiesenberger, G.; Schöfbeck, D.; Schuhmacher, R. Detection and identification of fungal microbial volatile organic compounds by HS-SPME-GC–MS. In Laboratory Protocols in Fungal Biology: Current Methods in Fungal Biology; Gupta, V.K., Tuohy, M.G., Ayyachamy, M., Turner, K.M., O’Donovan, A., Eds.; Springer: New York, NY, USA, 2013; pp. 455–465. [Google Scholar] [CrossRef]
- Ewen, R.J.; Jones, P.R.H.; Ratcliffe, N.M.; Spencer-Phillips, P.T.N. Identification by gas chromatography-mass spectrometry of the volatile organic compounds emitted from the wood-rotting fungi Serpula lacrymans and Coniophora puteana, and from Pinus sylvestris timber. Mycol. Res. 2004, 108, 806–814. [Google Scholar] [CrossRef]
- Vu, T.H.; Catheline, D.; Delmail, D.; Boustie, J.; Legrand, P.; Lohézic-Le Dévéhat, F. Gas chromatographic analysis to compare the fatty acid composition of fifteen lichen species, with a focus on Stereocaulon. Lichenologist 2016, 48, 323–337. [Google Scholar] [CrossRef]
- Mends, M.T.; Yu, E.; Strobel, G.A.; Hassan, S.; Booth, E.; Geary, B.; Sears, J.; Taatjes, C.; Hadi, M. An endophytic Nodulisporium sp. producing volatile organic compounds having bioactivity and fuel potential. J. Pet. Environ. Biotechnol. 2012, 3, 117. [Google Scholar] [CrossRef] [Green Version]
- Wariishi, H.; Madla, S.; Miura, D. Optimization of extraction method for GC-MS based metabolomics for filamentous fungi. J. Microbiol. Biochem. Technol. 2012, 4, 5–9. [Google Scholar] [CrossRef] [Green Version]
- Smart, K.F.; Aggio, R.B.M.; Van Houtte, J.R.; Villas-Bôas, S.G. Analytical platform for metabolome analysis of microbial cells using methyl chloroformate derivatization followed by gas chromatography-mass spectrometry. Nat. Protoc. 2010, 5, 1709–1729. [Google Scholar] [CrossRef]
- Heddergott, C.; Calvo, A.M.; Latgé, J.P. The volatome of Aspergillus fumigatus. Eukaryot. Cell 2014, 13, 1014–1025. [Google Scholar] [CrossRef] [Green Version]
- Briard, B.; Heddergott, C.; Latgé, J.P. Volatile compounds emitted by Pseudomonas aeruginosa stimulate growth of the fungal pathogen Aspergillus fumigatus. MBio 2016, 7, e00219-16. [Google Scholar] [CrossRef] [Green Version]
- Romano, P.; Pietrafesa, R.; Romaniello, R.; Zambuto, M.; Calabretti, A.; Capece, A. Impact of yeast starter formulations on the production of volatile compounds during wine fermentation. Yeast 2015, 32, 245–256. [Google Scholar] [CrossRef]
- Wihlborg, R.; Pippitt, D.; Marsili, R. Headspace sorptive extraction and GC-TOFMS for the identification of volatile fungal metabolites. J. Microbiol. Methods 2008, 75, 244–250. [Google Scholar] [CrossRef]
- Senes, C.E.R.; Saldan, N.C.; Costa, W.F.; Svidzinski, T.I.E.; Oliveira, C.C. Identification of Fusarium oxysporum fungus in wheat based on chemical markers and qualitative GC-MS test. J. Braz. Chem. Soc. 2018, 29, 2626–2635. [Google Scholar] [CrossRef]
- Aliferis, K.A.; Cubeta, M.A.; Jabaji, S. Chemotaxonomy of fungi in the Rhizoctonia solani species complex performing GC/MS metabolite profiling. Metabolomics 2013, 9, 159–169. [Google Scholar] [CrossRef]
- Suchorab, Z.; Frąc, M.; Guz, Ł.; Oszust, K.; Łagód, G.; Gryta, A.; Bilińska-Wielgus, N.; Czerwińskiid, J. A method for early detection and identification of fungal contamination of building materials using e-nose. PLoS ONE 2019, 14, e0215179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Portalo-Calero, F.; Arroyo, P.; Suárez, J.I.; Lozano, J. Triangular test of Amanita mushrooms by using electronic nose and sensory panel. Foods 2019, 8, 414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roze, L.V.; Beaudry, R.M.; Linz, J.E. Analysis of volatile compounds emitted by filamentous fungi using solid-phase microextraction-gas chromatography/mass spectrometry. In Fungal Secondary Metabolism: Methods and Protocols; Keller, N.P., Turner, G., Eds.; Humana Press: Totowa, NJ, USA, 2012; Volume 944, pp. 133–142. [Google Scholar] [CrossRef]
- Maxwell, T.; Blair, R.G.; Wang, Y.; Kettring, A.H.; Moore, S.D.; Rex, M.; Harper, J.K. A solvent-free approach for converting cellulose waste into volatile organic compounds with endophytic fungi. J. Fungi 2018, 4, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perl, T.; Jünger, M.; Vautz, W.; Nolte, J.; Kuhns, M.; Borg von Zepelin, M.; Quintel, M. Detection of characteristic metabolites of Aspergillus fumigatus and Candida species using ion mobility spectrometry-metabolic profiling by volatile organic compounds. Mycoses 2011, 54, e828–e837. [Google Scholar] [CrossRef] [PubMed]
- Sica, V.P.; Raja, H.A.; El-Elimat, T.; Kertesz, V.; van Berkel, G.J.; Pearce, C.J.; Oberlies, N.H. Dereplicating and spatial mapping of secondary metabolites from fungal cultures in situ. J. Nat. Prod. 2015, 78, 1926–1936. [Google Scholar] [CrossRef]
- Oberlies, N.H.; Knowles, S.L.; Amrine, C.S.M.; Kao, D.; Kertesz, V.; Raja, H.A. Droplet probe: Coupling chromatography to the in situ evaluation of the chemistry of nature. Nat. Prod. Rep. 2019, 36, 944–959. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Kaushik, N.; Proksch, P. Identification of antifungal principle in the solvent extract of an endophytic fungus Chaetomium globosum from Withania somnifera. Springerplus 2013, 2, 37. [Google Scholar] [CrossRef] [Green Version]
- Calla-Quispe, E.; Robles, J.; Areche, C.; Sepulveda, B. Are ionic liquids better extracting agents than toxic volatile organic solvents? A combination of ionic liquids, microwave and LC/MS/MS, applied to the lichen Stereocaulon glareosum. Front. Chem. 2020, 8, 450. [Google Scholar] [CrossRef]
- Cornejo, A.; Salgado, F.; Caballero, J.; Vargas, R.; Simirgiotis, M.; Areche, C. Secondary metabolites in Ramalina terebrata detected by UHPLC/ESI/MS/MS and identification of parietin as tau protein inhibitor. Int. J. Mol. Sci. 2016, 17, 1303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, J.; Elias, J.E.; Thoreen, C.C.; Licklider, L.J.; Gygi, S.P. Evaluation of multidimensional chromatography coupled with tandem mass spectrometry (LC/LC-MS/MS) for large-scale protein analysis: The yeast proteome. J. Proteome Res. 2003, 2, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Musharraf, S.G.; Kanwal, N.; Thadhani, V.M.; Choudhary, M.I. Rapid identification of lichen compounds based on the structure–fragmentation relationship using ESI-MS/MS analysis. Anal. Methods 2015, 7, 6066–6076. [Google Scholar] [CrossRef]
- Smedsgaard, J.; Nielsen, J. Metabolite profiling of fungi and yeast: From phenotype to metabolome by MS and informatics. J. Exp. Bot. 2005, 56, 273–286. [Google Scholar] [CrossRef] [Green Version]
- Olivier-Jimenez, D.; Chollet-Krugler, M.; Rondeau, D.; Beniddir, M.A.; Ferron, S.; Delhaye, T.; Allard, P.-M.; Wolfender, J.-L.; Sipman, H.J.M.; Lücking, R.; et al. A database of high-resolution MS/MS spectra for lichen metabolites. Sci. Data 2019, 6, 294. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, K.F.; Månsson, M.; Rank, C.; Frisvad, J.C.; Larsen, T.O. Dereplication of microbial natural products by LC-DAD-TOFMS. J. Nat. Prod. 2011, 74, 2338–2348. [Google Scholar] [CrossRef]
- El-Elimat, T.; Figueroa, M.; Ehrmann, B.M.; Cech, N.B.; Pearce, C.J.; Oberlies, N.H. High-resolution MS, MS/MS, and UV database of fungal secondary metabolites as a dereplication protocol for bioactive natural products. J. Nat. Prod. 2013, 76, 1709–1716. [Google Scholar] [CrossRef] [Green Version]
- Carpentier, C.; Queiroz, E.F.; Marcourt, L.; Wolfender, J.-L.; Azelmat, J.; Grenier, D.; Boudreau, S.; Voyer, N. Dibenzofurans and pseudodepsidones from the lichen Stereocaulon paschale Collected in Northern Quebec. J. Nat. Prod. 2017, 80, 210–214. [Google Scholar] [CrossRef]
- Salgado, F.; Albornoz, L.; Cortez, C.; Stashenko, E.; Urrea-Vallejo, K.; Nagles, E.; Galicia-Virviescas, C.; Cornejo, A.; Ardiles, A.; Simirgiotis, M.; et al. Secondary metabolite profiling of species of the genus Usnea by UHPLC-ESI-OT-MS-MS. Molecules 2017, 23, 54. [Google Scholar] [CrossRef] [Green Version]
- Torres-Benitez, A.; Rivera-Montalvo, M.; Sepulveda, B.; Castro, O.N.; Nagles, E.; Simirgiotis, M.J.; Garcia-Beltran, O.; Areche, C. Metabolomic analysis of two Parmotrema lichens: P. robustum (Degel.) Hale and P. andinum (Mull. Arg.) Hale using UHPLC-ESI-OT-MS-MS. Molecules 2017, 22, 1861. [Google Scholar] [CrossRef] [Green Version]
- Castro, O.N.; Benites, J.; Rodilla, J.; Santiago, J.C.; Simirgiotis, M.; Sepulveda, B.; Areche, C. Metabolomic analysis of the lichen Everniopsis trulla using ultra high performance liquid chromatography-quadrupole-orbitrap mass spectrometry (UHPLC-Q-OT-MS). Chromatographia 2017, 80, 967–973. [Google Scholar] [CrossRef]
- Klitgaard, A.; Iversen, A.; Andersen, M.R.; Larsen, T.O.; Frisvad, J.C.; Nielsen, K.F. Aggressive dereplication using UHPLC-DAD-QTOF: Screening extracts for up to 3000 fungal secondary metabolites. Anal. Bioanal. Chem. 2014, 406, 1933–1943. [Google Scholar] [CrossRef] [Green Version]
- Ióca, L.P.; Romminger, S.; Santos, M.F.C.; Bandeira, K.F.; Rodrigues, F.T.; Kossuga, M.H.; Nicacio, K.J.; Ferreira, E.L.F.; Morais-Urano, R.P.; Passos, M.S.; et al. A strategy for the rapid identification of fungal metabolites and the discovery of the antiviral activity of pyrenocine a and harzianopyridone. Quím. Nova 2016, 39, 720–731. [Google Scholar] [CrossRef]
- Degenkolb, T.; Grafenhan, T.; Nirenberg, H.I.; Gams, W.; Bruckner, H. Trichoderma brevicompactum complex: Rich source of novel and recurrent plant-protective polypeptide antibiotics (peptaibiotics). J. Agric. Food Chem. 2006, 54, 7047–7061. [Google Scholar] [CrossRef]
- Stodůlková, E.; Man, P.; Kolarík, M.; Flieger, M. High-performance liquid chromatography-off line mass spectrometry analysis of anthraquinones produced by Geosmithia lavendula. J. Chromatogr. A 2010, 1217, 6296–6302. [Google Scholar] [CrossRef] [PubMed]
- Parrot, D.; Jan, S.; Baert, N.; Guyot, S.; Tomasi, S. Comparative metabolite profiling and chemical study of Ramalina siliquosa complex using LC-ESI-MS/MS approach. Phytochemistry 2013, 89, 114–124. [Google Scholar] [CrossRef]
- Melero, I.Q.; Peláez, T.; Rezusta Lopez, A.; Garcia-Rodríguez, J. Application of mass spectrometry in mycology. Enferm. Infecc. Microbiol. Clin. 2016, 34, 26–30. [Google Scholar] [CrossRef]
- Freiwald, A.; Sauer, S. Phylogenetic classification and identification of bacteria by mass spectrometry. Nat. Protoc. 2009, 4, 732–742. [Google Scholar] [CrossRef]
- Singhal, N.; Kumar, M.; Kanaujia, P.K.; Virdi, J.S. MALDI-TOF mass spectrometry: An emerging technology for microbial identification and diagnosis. Front. Microbiol. 2015, 6, 791. [Google Scholar] [CrossRef] [Green Version]
- Khot, P.D.; Fisher, M.A. Novel approach for differentiating Shigella species and Escherichia coli by matrix-assisted laser desorption ionization-time of flight mass spectrometry. J. Clin. Microbiol. 2013, 51, 3711–3716. [Google Scholar] [CrossRef] [Green Version]
- Santos, C.; Paterson, R.R.; Venancio, A.; Lima, N. Filamentous fungal characterizations by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry. J. Appl. Microbiol. 2010, 108, 375–385. [Google Scholar] [CrossRef] [Green Version]
- Rahi, P.; Prakash, O.; Shouche, Y.S. Matrix-assisted laser desorption/ionization time-of-flight mass-spectrometry (MALDI-TOF MS) based microbial identifications: Challenges and scopes for microbial ecologists. Front. Microbiol. 2016, 7, 1359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timperio, A.M.; Gorrasi, S.; Zolla, L.; Fenice, M. Evaluation of MALDI-TOF mass spectrometry and MALDI BioTyper in comparison to 16S rDNA sequencing for the identification of bacteria isolated from Arctic sea water. PLoS ONE 2017, 12, e0181860. [Google Scholar] [CrossRef] [PubMed]
- Grenfell, R.C.; Da Silva Junior, A.R.; Del Negro, G.M.B.; Munhoz, R.B.; Gimenes, V.M.F.; Assis, D.M.; Rockstroh, A.C.; Motta, A.L.; Rossi, F.; Juliano, L.; et al. Identification of Candida haemulonii complex species: Use of ClinProToolsTM to overcome limitations of the Bruker BiotyperTM, VITEK MSTM IVD, and VITEK MSTM RUO databases. Front. Microbiol. 2016, 7, 940. [Google Scholar] [CrossRef] [PubMed]
- Gekenidis, M.T.; Studer, P.; Wuthrich, S.; Brunisholz, R.; Drissner, D. Beyond the matrix-assisted laser desorption ionization (MALDI) biotyping workflow: In search of microorganism-specific tryptic peptides enabling discrimination of subspecies. Appl. Environ. Microbiol. 2014, 80, 4234–4241. [Google Scholar] [CrossRef] [Green Version]
- Hung, R.; Lee, S.; Bennett, J.W. Fungal volatile organic compounds and their role in ecosystems. Appl. Microbiol. Biotechnol. 2015, 99, 3395–3405. [Google Scholar] [CrossRef]
- Mansurova, M.; Ebert, B.E.; Blank, L.M.; Ibanez, A.J. A breath of information: The volatilome. Curr. Genet. 2018, 64, 959–964. [Google Scholar] [CrossRef]
- Scalbert, A.; Brennan, L.; Fiehn, O.; Hankemeier, T.; Kristal, B.S.; van Ommen, B.; Pujos-Guillot, E.; Verheij, E.; Wishart, D.; Wopereis, S. Mass-spectrometry-based metabolomics: Limitations and recommendations for future progress with particular focus on nutrition research. Metabolomics 2009, 5, 435–458. [Google Scholar] [CrossRef] [Green Version]
- Kai, M. Diversity and distribution of volatile secondary metabolites throughout Bacillus subtilis isolates. Front. Microbiol. 2020, 11, 559. [Google Scholar] [CrossRef]
- Wang, J.H.; Byun, J.; Pennathur, S. Analytical approaches to metabolomics and applications to systems biology. Semin. Nephrol. 2010, 30, 500–511. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, K.F.; Larsen, T.O. The importance of mass spectrometric dereplication in fungal secondary metabolite analysis. Front. Microbiol. 2015, 6, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mashego, M.R.; Rumbold, K.; Mey, M.D.; Vandamme, E.; Soetaert, W.; Heijnen, J.J. Microbial metabolomics: Past, present and future methodologies. Biotechnol. Lett. 2007, 29, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, A.A.; Altaf-Ul-Amin, M.; Ono, N.; Sato, T.; Sugiura, T.; Morita, A.H.; Katsuragi, T.; Muto, A.; Nishioka, T.; Kanaya, S. Development and mining of a volatile organic compound database. Biomed. Res. Int. 2015, 2015, 139254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neumann, S.; Böcker, S. Computational mass spectrometry for metabolomics: Identification of metabolites and small molecules. Anal. Bioanal. Chem. 2010, 398, 2779–2788. [Google Scholar] [CrossRef] [Green Version]
- Fiehn, O. Metabolomics by gas chromatography-mass spectrometry: Combined targeted and untargeted profiling. Curr. Protoc. Mol. Biol. 2016, 114, 21–33. [Google Scholar] [CrossRef]
- Singh, K.D.; Del Miguel, G.V.; Gaugg, M.T.; Ibanez, A.J.; Zenobi, R.; Kohler, M.; Frey, U.; Sinues, P.M. Translating secondary electrospray ionization-high-resolution mass spectrometry to the clinical environment. J. Breath Res. 2018, 12, 027113. [Google Scholar] [CrossRef]
- Rioseras, A.T.; Gaugg, M.T.; Martinez-Lozano Sinues, P. Secondary electrospray ionization proceeds via gas-phase chemical ionization. Anal. Methods 2017, 9, 5052–5057. [Google Scholar] [CrossRef] [Green Version]
- Lawal, O.; Ahmed, W.M.; Nijsen, T.M.E.; Goodacre, R.; Fowler, S.J. Exhaled breath analysis: A review of ‘breath-taking’ methods for off-line analysis. Metabolomics 2017, 13, 110. [Google Scholar] [CrossRef]
- Cumeras, R.; Figueras, E.; Davis, C.E.; Baumbach, J.I.; Gracia, I. Review on ion mobility spectrometry. Part 1: Current instrumentation. Analyst 2015, 140, 1376–1390. [Google Scholar] [CrossRef] [Green Version]
- Purkhart, R.; Becher, G.; Reinhold, P.; Köhler, H.U. Detection of mycobacteria by volatile organic compound analysis of invitro cultures using differential ion mobility spectrometry. J. Med. Microbiol. 2017, 66, 276–285. [Google Scholar] [CrossRef]
- Lan, H.; Hartonen, K.; Riekkola, M.-L. Miniaturised air sampling techniques for analysis of volatile organic compounds in air. TrAC Trends Anal. Chem. 2020, 126, 115873. [Google Scholar] [CrossRef]
- Somerville, G.A.; Proctor, R.A. Cultivation conditions and the diffusion of oxygen into culture media: The rationale for the flask-to-medium ratio in microbiology. BMC Microbiol. 2013, 13, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kai, M.; Crespo, E.; Cristescu, S.M.; Harren, F.J.; Francke, W.; Piechulla, B. Serratia odorifera: Analysis of volatile emission and biological impact of volatile compounds on Arabidopsis thaliana. Appl. Microbiol. Biotechnol. 2010, 88, 965–976. [Google Scholar] [CrossRef] [PubMed]
- Synytsya, A.; Monkai, J.; Bleha, R.; Macurkova, A.; Ruml, T.; Ahn, J.; Chukeatirote, E. Antimicrobial activity of crude extracts prepared from fungal mycelia. Asian Pac. J. Trop. Biomed. 2016, 7, 257–261. [Google Scholar] [CrossRef]
- Joshi, D.; Adhikari, N. An overview on common organic solvents and their toxicity. J. Pharm. Res. Int. 2019, 28, 1–18. [Google Scholar] [CrossRef]
- Kildgaard, S.; Mansson, M.; Dosen, I.; Klitgaard, A.; Frisvad, J.C.; Larsen, T.O.; Nielsen, K.F. Accurate dereplication of bioactive secondary metabolites from marine-derived fungi by UHPLC-DAD-QTOFMS and a MS/HRMS library. Mar. Drugs. 2014, 12, 3681–3705. [Google Scholar] [CrossRef] [Green Version]
- Wolfender, J.-L.; Marti, G.; Thomas, A.; Bertrand, S. Current approaches and challenges for the metabolite profiling of complex natural extracts. J. Chromatogr. A 2015, 1382, 136–164. [Google Scholar] [CrossRef]
- Euerby, M.R.; Petersson, P. Chromatographic classification and comparison of commercially available reversed-phase liquid chromatographic columns containing polar embedded groups/amino endcappings using principal component analysis. J. Chromatogr. A 2003, 944, 13–36. [Google Scholar] [CrossRef]
- Pucci, V.; Di Palma, S.; Alfieri, A.; Bonelli, F.; Monteagudo, E. A novel strategy for reducing phospholipids-based matrix effect in LC-ESI-MS bioanalysis by means of HybridSPE. J. Pharm. Biomed. Anal. 2009, 50, 867–871. [Google Scholar] [CrossRef]
- Bollag, G.; Tsai, J.; Zhang, J.; Zhang, C.; Ibrahim, P.; Nolop, K.; Hirth, P. Vemurafenib: The first drug approved for BRAF-mutant cancer. Nat. Rev. Drug Discov. 2012, 11, 873–886. [Google Scholar] [CrossRef]
- Baidoo, E.E.; Benke, P.I.; Keasling, J.D. Mass spectrometry-based microbial metabolomics: Techniques, analysis, and applications. In Microbial Metabolomics. Methods in Molecular Biology; Baidoo, E., Ed.; Humana Press: New York, NY, USA, 2018; pp. 11–69. [Google Scholar] [CrossRef]
- Zeng, X.; Zhang, P.; He, W.; Qin, C.; Chen, S.; Tao, L.; Wang, Y.; Tan, Y.; Gao, D.; Wang, B.; et al. NPASS: Natural product activity and species source database for natural product research, discovery and tool development. Nucleic Acids Res. 2017, 46, D1217–D1222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Santen, J.A.; Jacob, G.; Singh, A.L.; Aniebok, V.; Balunas, M.J.; Bunsko, D.; Neto, F.C.; Castaño-Espriu, L.; Chang, C.; Clark, T.N.; et al. The natural products atlas: An open access knowledge base for microbial natural products discovery. ACS Cennt. Sci. 2019, 5, 1824–1833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sorokina, M.; Steinbeck, C. Review on natural products databases: Where to find data in 2020. J. Cheminform. 2020, 12, 1–51. [Google Scholar] [CrossRef] [Green Version]
- Spengler, B. Mass spectrometry imaging of biomolecular information. Anal. Chem. 2015, 87, 64–82. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Balan, P.; Vertes, A. Molecular imaging of growth, metabolism, and antibiotic inhibition in bacterial colonies by laser ablation electrospray ionization mass spectrometry. Angew. Chem. Int. Ed. Engl. 2016, 55, 15035–15039. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, B.; Walsh, C.; Boyce, G.; Nemes, P. Microprobe MS imaging of live tissues, cells, and bacterial colonies using LAESI. In Advances in MALDI and Laser-Induced Soft Ionization Mass Spectrometry; Cramer, R., Ed.; Springer: Cham, Switzerland, 2016; pp. 149–167. [Google Scholar] [CrossRef]
- Netzker, T.; Fischer, J.; Weber, J.; Mattern, D.J.; König, C.C.; Valiante, V.; Schroeckh, V.; Brakhage, A.A. Microbial communication leading to the activation of silent fungal secondary metabolite gene clusters. Front. Microbiol. 2015, 6, 299. [Google Scholar] [CrossRef]
- Cooks, R.G.; Ouyang, Z.; Takats, Z.; Wiseman, J.M. Detection Technologies. Ambient mass spectrometry. Science 2006, 311, 1566–1570. [Google Scholar] [CrossRef]
Figure 1.
Development of fungal studies involving fungal metabolites and mass spectrometry. Publications from 1985 to 2020 involving metabolites (blue bars) and mass spectrometry (orange bars) in mycological studies. The search was carried out in the portal of the Web of Science using the following search terms: (i) “fungal OR fungi AND metabolite*”; and (ii) “fungal OR fungi AND mass spectromet*”.
Figure 1.
Development of fungal studies involving fungal metabolites and mass spectrometry. Publications from 1985 to 2020 involving metabolites (blue bars) and mass spectrometry (orange bars) in mycological studies. The search was carried out in the portal of the Web of Science using the following search terms: (i) “fungal OR fungi AND metabolite*”; and (ii) “fungal OR fungi AND mass spectromet*”.

Figure 2.
Workflow of studies involving mass spectrometry and identification of fungal chemical compounds. A summary of the steps involving mass spectrometry used in microfungal chemical studies, from sample preparation to analysis. Micro-fungi are used as an example and sample techniques are also shown.
Figure 2.
Workflow of studies involving mass spectrometry and identification of fungal chemical compounds. A summary of the steps involving mass spectrometry used in microfungal chemical studies, from sample preparation to analysis. Micro-fungi are used as an example and sample techniques are also shown.


{kind=link}
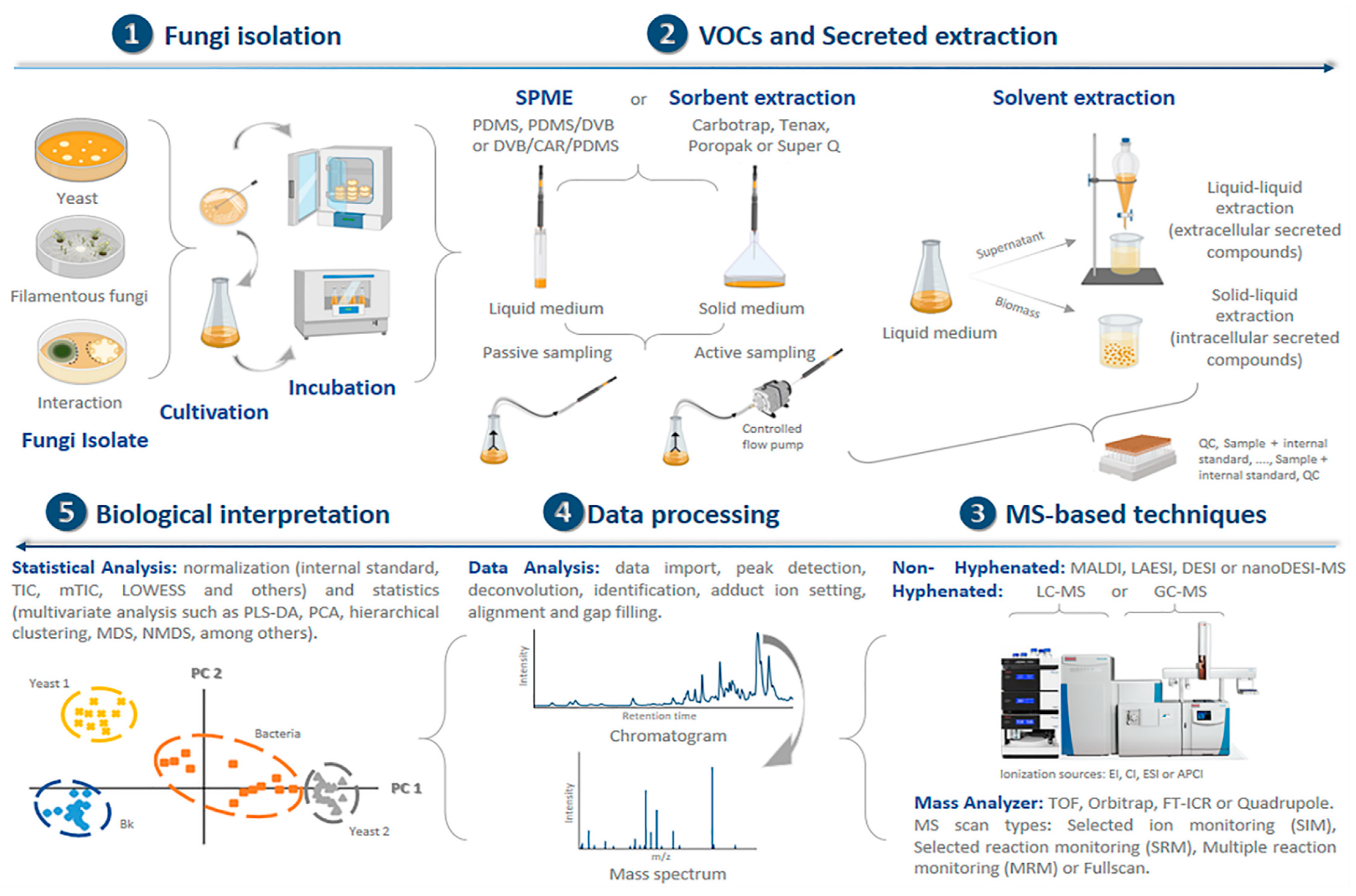{kind=link}
Table 1.
Mass spectrometry-based strategies used in mycological studies.
| Application | MS System | Associated MS Database | Sample Processing | Targeted Chemical Compounds | References |
|---|---|---|---|---|---|
| Biotyping | MALDI-TOF-MS, MALDI-TOF-IC MS | Biotyper, SARAMIS, MBT Compass, BDAL | solvent extraction (ethanol-formic acid, trifluoroacetic acid or formic acid/acetonitrile) | peptides or proteins | [73,74,75,76,77,78,79,80,81,82,83,84,85,86,87,88,89,90,91,92] |
| Biotyping | Rapid evaporative ionization mass spectrometry (REIMS) | In-house database | no pre-processing | fatty acids, lipids | [93,94] |
| Biotyping | PCR electrospray ionization-mass spectrometry (PCR/ESI-MS) | PLEX-ID system, GenBank | no pre-processing | proteins, polysaccharides | [95,96,97] |
| Biotyping | Paper spray mass spectrometry (PS-MS) | In-house database | disruption analysis (cell lysis method) | lipids | [98] |
| Metabolomics | PTR-MS | NIST library, chemical standards | no pre-processing | alcohols, aldehydes, esters, ketones, lipids, organic acids, terpenes, among other VOCs | [99,100,101,102] |
| Metabolomics | SIFT-MS | Precursor–VOC reaction | no pre-processing | alcohols, aldehydes, ketones, among other VOCs | [103,104,105] |
| Metabolomics | SESI-MS | In-house database | no pre-processing | fatty acids, among other VOCs | [106] |
| Metabolomics | LDI-MS | In-house database | solvent extraction (e.g., dichloromethane) | anthraquinones, aromatics, chromones, depsides, depsidones, dibenzofurans, diphenylethers, lipids, naphthoquinone, polyols, xanthones. | [107] |
| Metabolomics | DESI-MS | In-house database | no pre-processing | anthraquinones, peptaibols, phomopsinones, pyrroloindoles | [108,109,110,111] |
| Biotyping | Gas chromatography-combustion-isotope ratio mass spectrometry (GC-C-IRMS) | In-house database, a mixed raster of 14 amino acids with known ẟ-13C | solvent extraction (e.g., dichloromethane) | amino acids | [112] |
| Biotyping and Metabolomics | GC-MS, GC-IMS, GC-FID Ionization sources: EI, CI, APCI Mass analyzer: Q or TOF | NIST library, LTPRIs, chemical standards | -liquid–liquid extraction (ethyl acetate–methanol, ethyl acetate–water) -solid–liquid extraction (methanol, dichloromethane or methanol-chloroform-water) -passive sampling, SPME (PA, PDMS, DVB, CAR/PDMS, DVB/PDMS or DVB/CAR/PDMS), SPE (C18) or sorbent (PTFE, Carbotrap, Tenax or bentonite) -active sampling, sorbent or SPME | alcohols, aldehydes, alkanes, alkenes, amino acids, carboxylic acids, carbohydrates, esters, ketones, fatty acids, lipids, organic acids, terpenes, among other VOCs | [14,99,100,102,113,114,115,116,117,118,119,120,121,122,123,124,125,126,127,128,129,130] |
| Metabolomics | LC-MS | ACD IntelliXtract, in-house database | direct analysis (droplet−LMJ−SSP) | anthraquinones, alkaloids | [131,132] |
| Metabolomics | LC-MS Ionization sources: ESI or APCI Mass analyzer: ion-trap, orbitrap, Q, TOF | Antibase, LDB, GNPS, ACD Chemfolder, NIST, DNP, Mycosynthetix, ORFs, Peptaibol, chemical standards | -liquid–liquid extraction (ethyl acetate, methanol-chloroform or ethyl acetate:acetic acid) -solid–liquid extraction (acetone, methanol, dichloromethane or dichloromethane:methanol) -cleanup by SPE (C18) | anthraquinones, aromatics, chromanones, chromones, depsides, depsidones, dibenzofurans, diphenylethers, lipids, naphthoquinones, peptides, polyols, xanthones. | [133,134,135,136,137,138,139,140,141,142,143,144,145,146,147,148,149,150] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Calla-Quispe, E.; Fuentes-Rivera, H.L.; Ramírez, P.; Martel, C.; Ibañez, A.J. Mass Spectrometry: A Rosetta Stone to Learn How Fungi Interact and Talk. Life 2020, 10, 89. https://doi.org/10.3390/life10060089
AMA Style
Calla-Quispe E, Fuentes-Rivera HL, Ramírez P, Martel C, Ibañez AJ. Mass Spectrometry: A Rosetta Stone to Learn How Fungi Interact and Talk. Life. 2020; 10(6):89. https://doi.org/10.3390/life10060089
Chicago/Turabian StyleCalla-Quispe, Erika, Hammerly Lino Fuentes-Rivera, Pablo Ramírez, Carlos Martel, and Alfredo J. Ibañez. 2020. "Mass Spectrometry: A Rosetta Stone to Learn How Fungi Interact and Talk" Life 10, no. 6: 89. https://doi.org/10.3390/life10060089
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.