
Effects of Ionizing Radiation and Long-Term Storage on Hydrated vs. Dried Cell Samples of Extremophilic Microorganisms

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
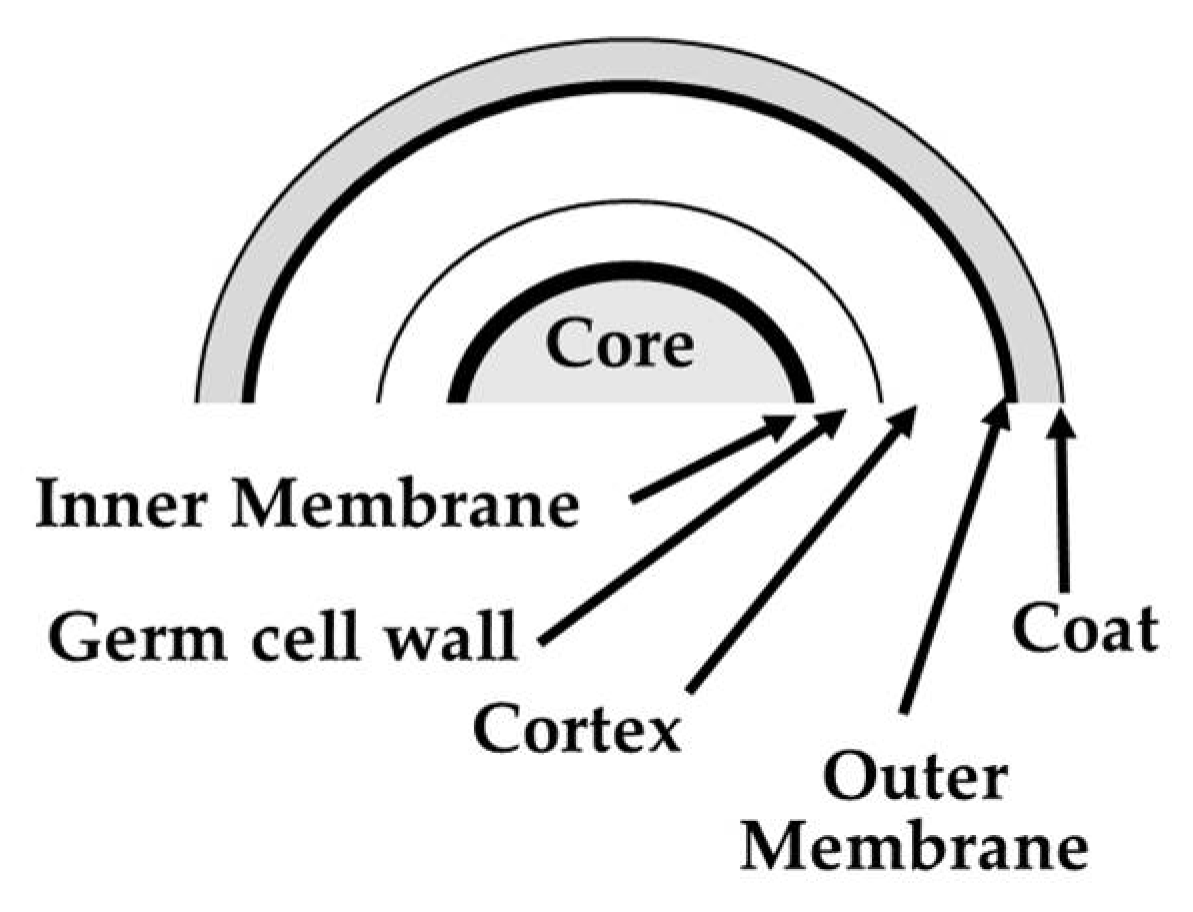
2.1. Production and Purification of Spores of Parageobacillus Thermantarcticus
2.2. Vegetative Cell Culture Conditions
2.3. Radiation Exposure Conditions
2.4. HZE Cross Section Calculations
2.5. Surface-Enhanced Raman Spectroscopy (SERS) Measurements
2.6. Synchrotron Infrared Microspectroscopy
3. Results
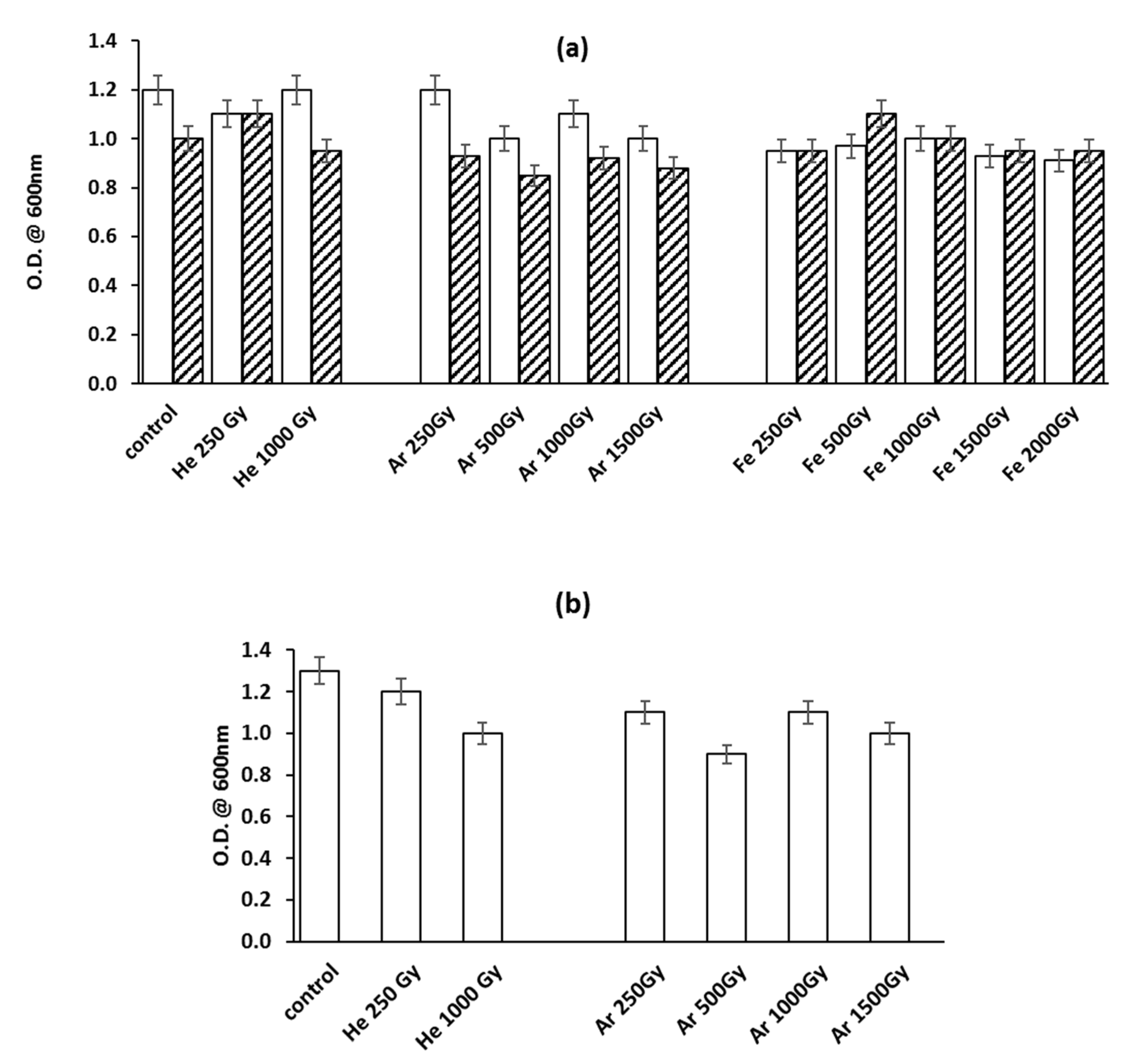
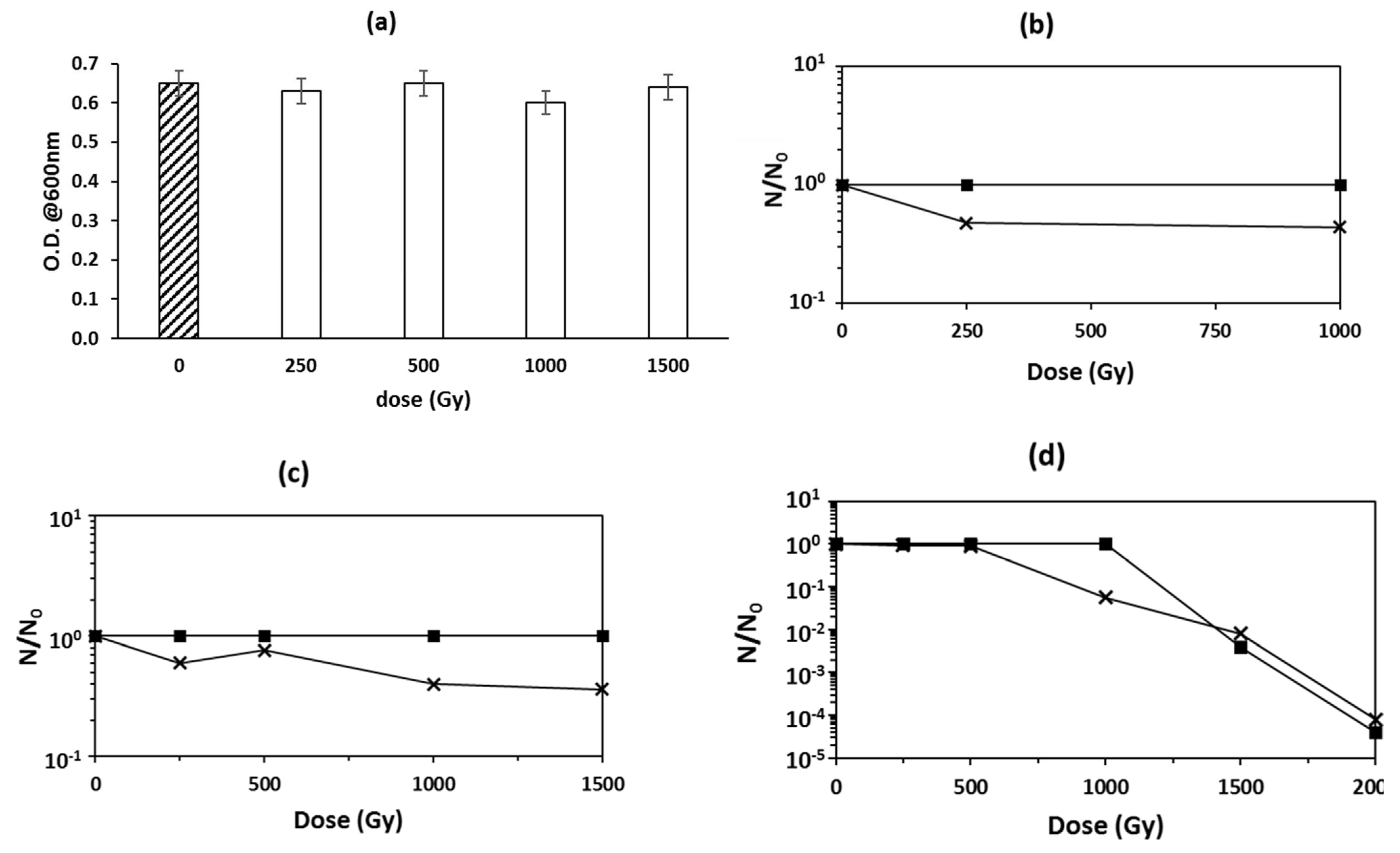
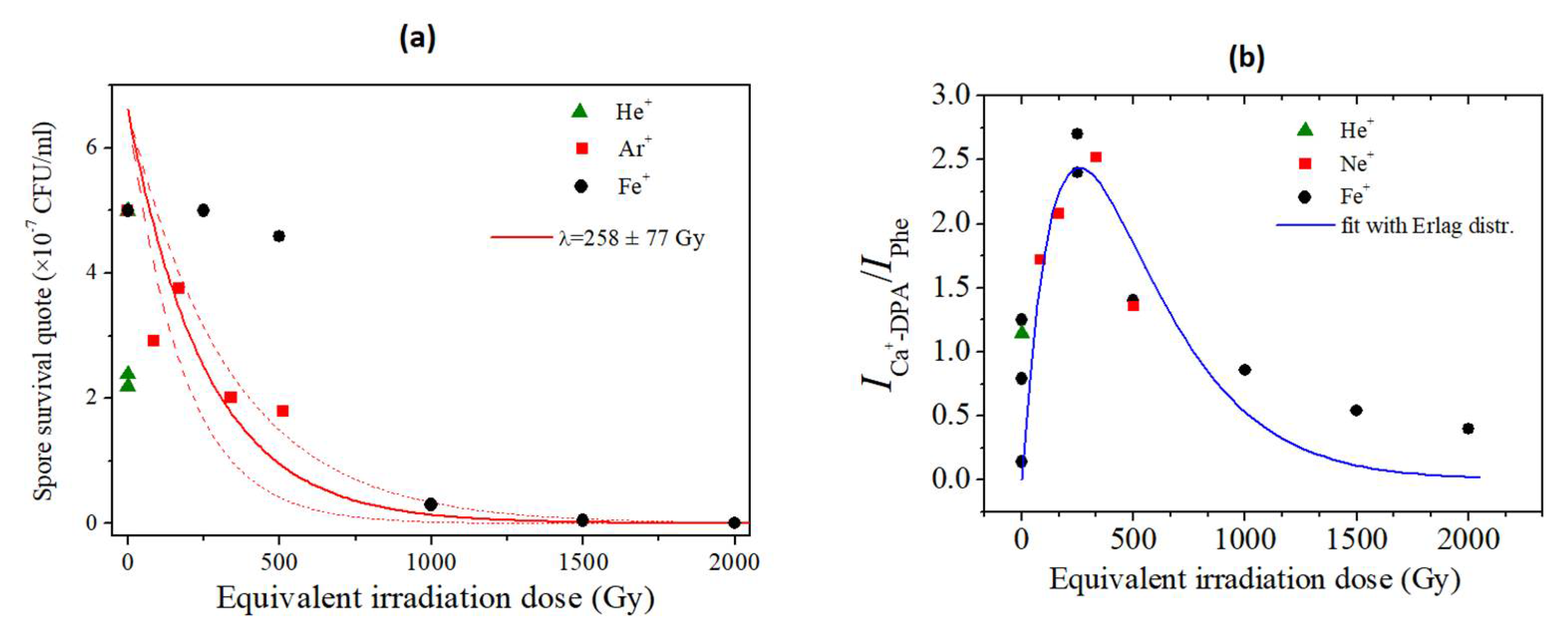
3.1. Microorganisms’ Survival to Irradiation with HZE
3.2. Biological Effects of HZE on Bacterial Spores
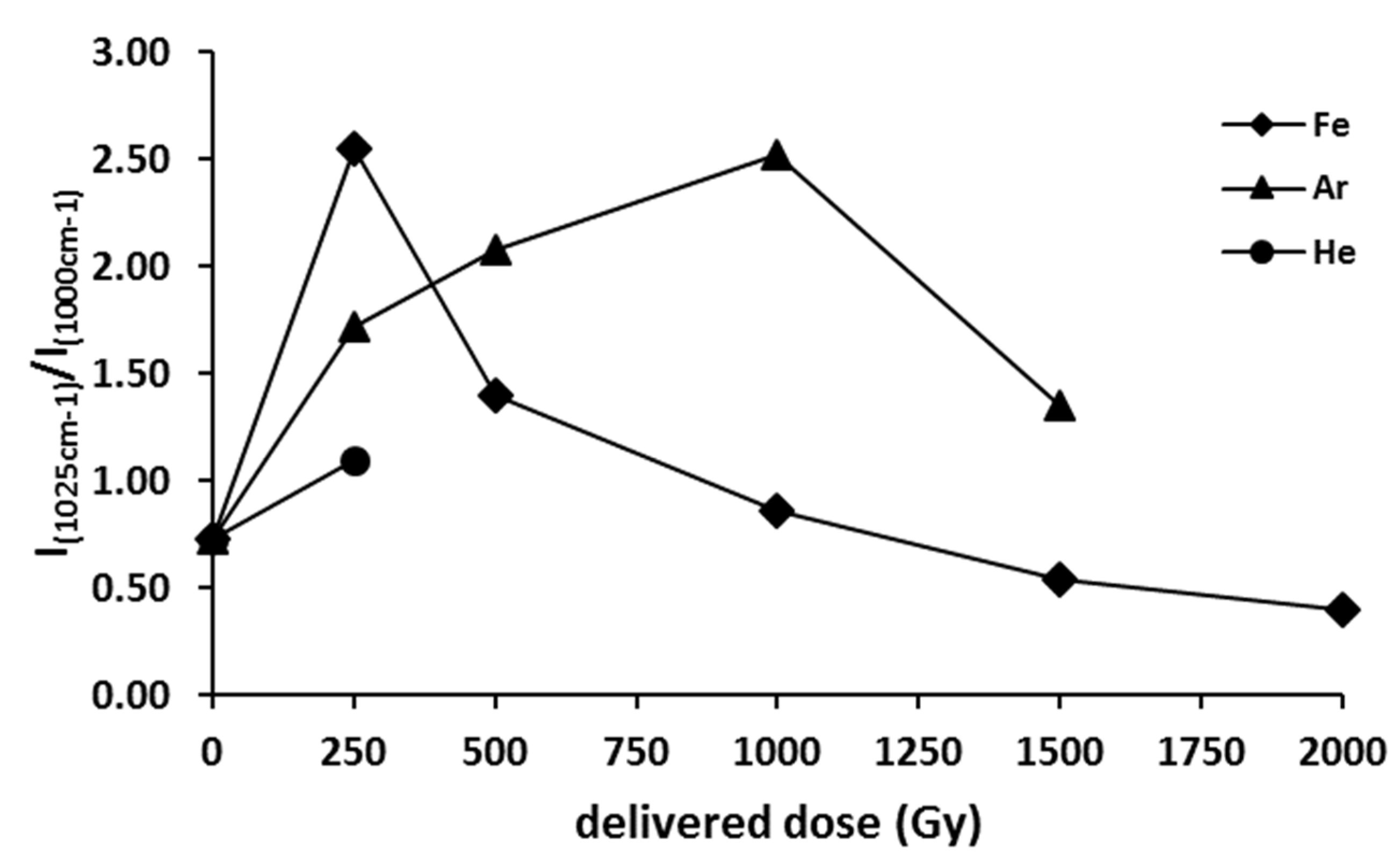
3.2.1. SERS analysis of spores of P. thermantarcticus after HZE ion irradiation
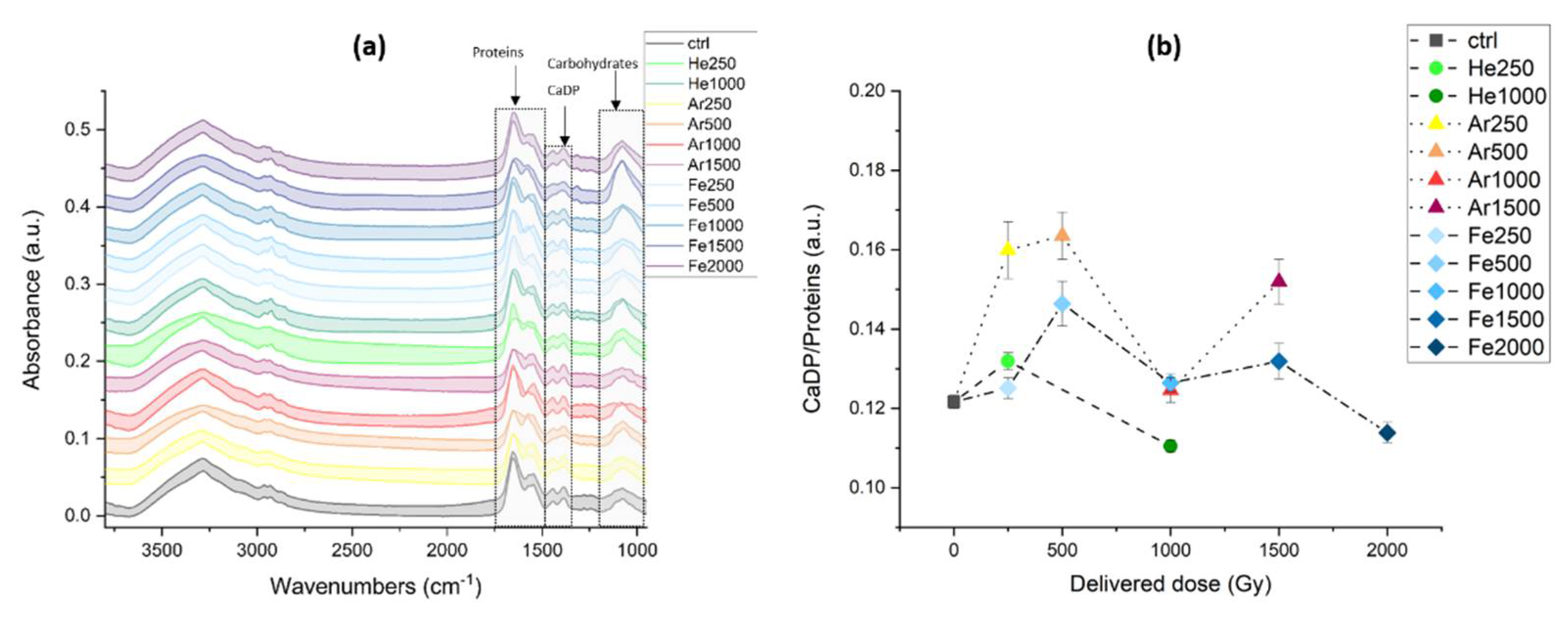
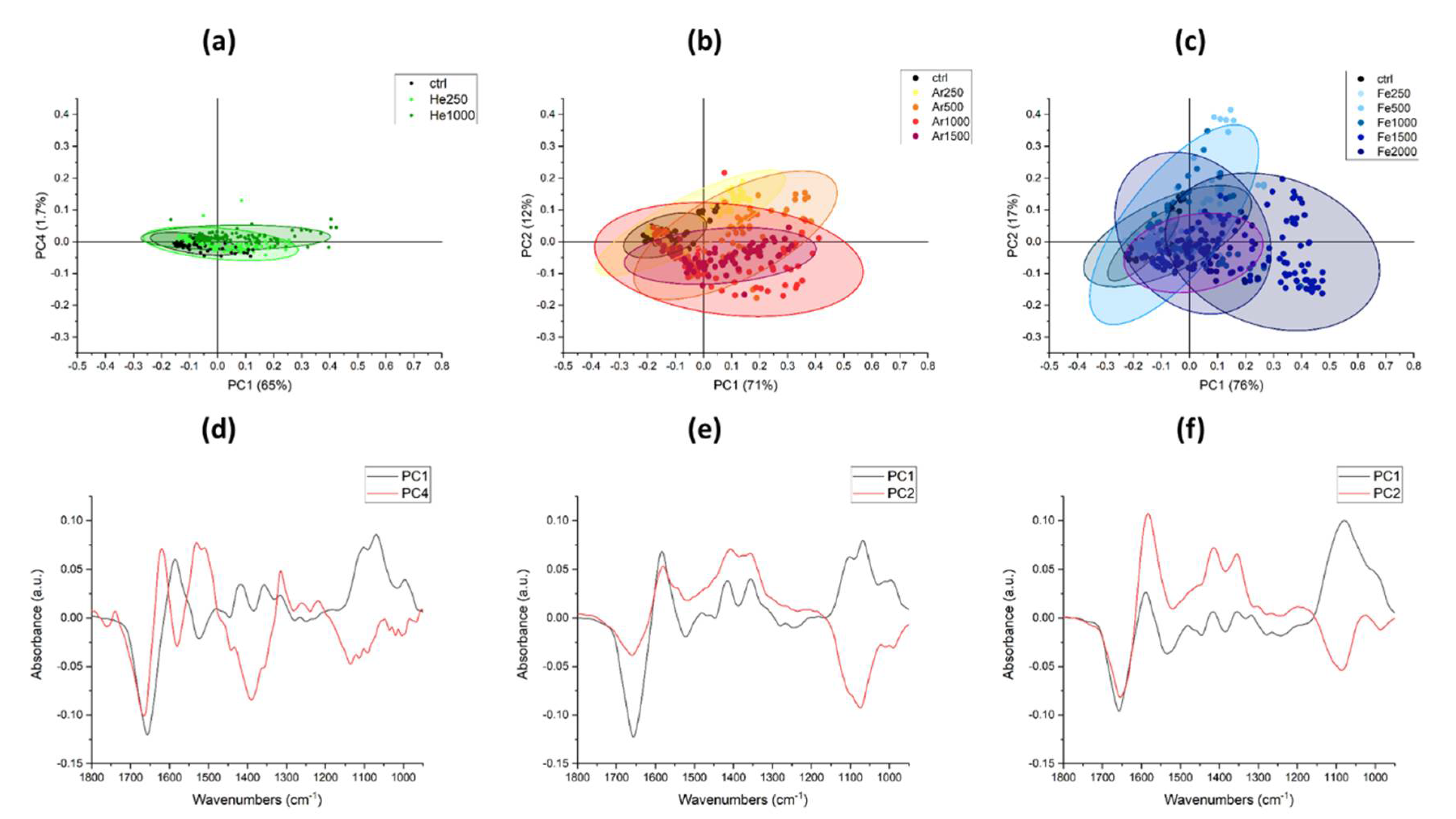
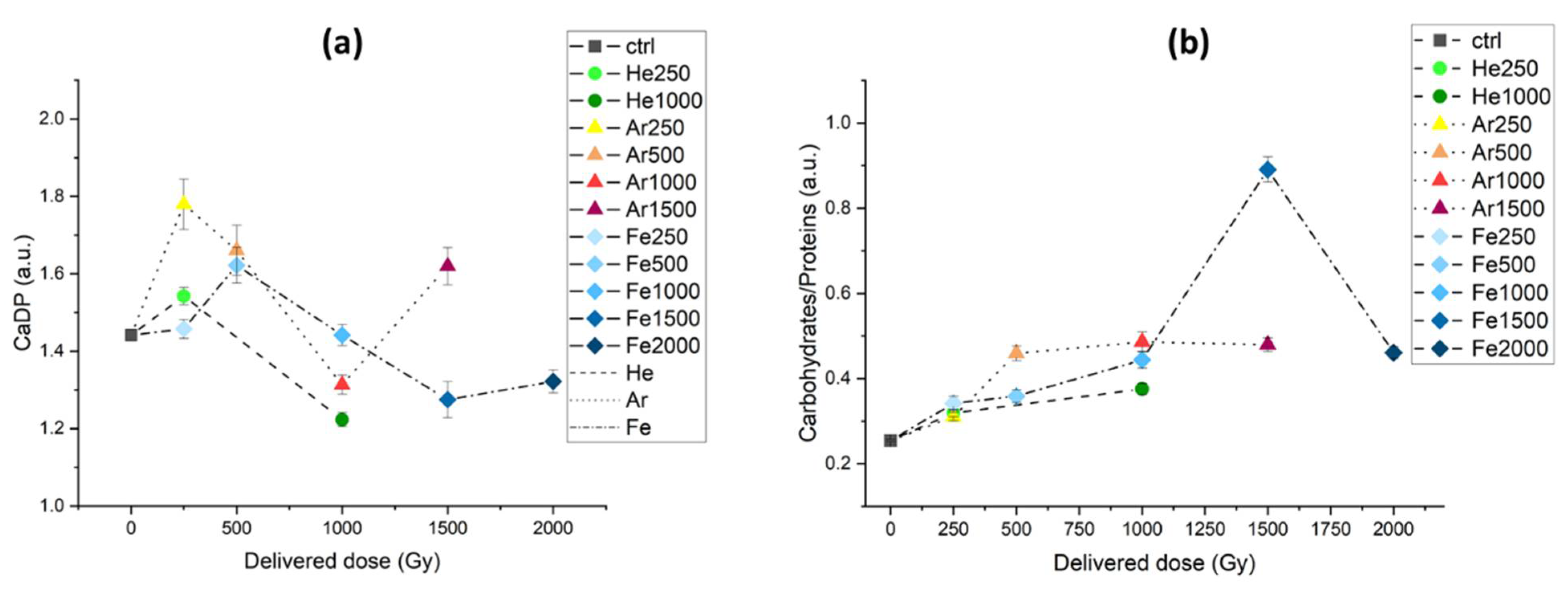
3.2.2. IR Microspectroscopy Analysis of Spores of P. thermantarcticus Spores after Irradiation with HZE in Solution
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

Appendix B
References
- Ferrari, F.; Szuszkiewicz, E. Cosmic Rays: A Review for Astrobiologists. Astrobiology 2009, 9, 413–436. [Google Scholar] [CrossRef] [PubMed]
- Horneck, G.; Klaus, D.M.; Mancinelli, R.L. Space microbiology. Microbiol. Mol. Biol. Rev. 2010, 74, 121–156. [Google Scholar] [CrossRef] [Green Version]
- Zammuto, V.; Rizzo, M.G.; De Plano, L.M.; Franco, D.; Guglielmino, S.; Caccamo, M.T.; Magazù, S.; Fujimori, A.; Lo Giudice, A.; Guglielmin, M.; et al. Effects of heavy ion particle irradiation on spore germination of bacillus spp. from extremely hot and cold environments. Life 2020, 10, 264. [Google Scholar]
- Moeller, R.; Raguse, M.; Leuko, S.; Berger, T.; Hellweg, C.E.; Fujimori, A.; Okayasu, R.; Horneck, G.; the STARLIFE Research Group. STARLIFE—An international Campaign to Study the Role of Galactic Cosmic Radiation in Astrobiological Model Systems. Astrobiology 2017, 17, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Durante, M.; Cucinotta, F.A. Physical basis of radiation protection in space travel. Rev. Mod. Phys. 2011, 83, 1245–1281. [Google Scholar] [CrossRef]
- Mastascusa, V.; Romano, I.; Di Donato, P.; Poli, A.; Della Corte, V.; Rotundi, A.; Bussoletti, E.; Quarto, M.; Pugliese, M.; Nicolaus, B. Extremophiles survival to simulated space conditions: An astrobiology model study. Orig. Life Evol. Biosph. 2014, 44, 231–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Donato, P.; Romano, I.; Mastascusa, V.; Poli, A.; Orlando, P.; Pugliese, M.; Nicolaus, B. Survival and Adaptation of the Thermophilic Species Geobacillus thermantarcticus in Simulated Spatial Conditions. Orig. Life Evol. Biosph. 2018, 48, 141–158. [Google Scholar] [CrossRef]
- Romano, I.; de Angelis, A.; Poli, A.; Ragni, P.; Lilla, L.; Zito, G.; Nicolaus, B.; De Luca, A.C.; Di Donato, P. Resistance and Raman spectroscopy analysis of Parageobacillus thermantarcticus spores after γ-ray exposure. Extremophiles 2018, 22, 931–941. [Google Scholar] [CrossRef]
- Romano, I.; Poli, A.; Finore, I.; Huertas, F.J.; Gambacorta, A.; Pelliccione, S.; Nicolaus, G.; Lama, L.; Nicolaus, B. Haloterrigena hispanica sp. nov., an extremely halophilic archaeon from Fuente de Piedra, southern Spain. Int. J. Syst. Evol. Microbiol. 2007, 57, 1499–1503. [Google Scholar] [CrossRef] [Green Version]
- Bauermeister, A.; Moeller, R.; Reitz, G.; Sommer, S.; Rettberg, P. Effect of Relative Humidity on Deinococcus radiodurans’ Resistance to Prolonged Desiccation, Heat, Ionizing, Germicidal, and Environmentally Relevant UV Radiation. Microb. Eco. 2011, 61, 715–722. [Google Scholar] [CrossRef]
- Paulino-Lima, I.; Janot-Pacheco, E.; Galante, D.; Cockell, C.; Olsson-Francis, K.; Brucato, J.R.; Baratta, G.; Strazzulla, G.; Merrigan, T.; McCullough, R.; et al. Survival of Deinococcus radiodurans Against Laboratory-Simulated Solar Wind Charged Particles. Astrobiology 2011, 11, 875–882. [Google Scholar] [CrossRef] [PubMed]
- Okayasu, R.; Okada, M.; Okabe, A.; Noguchi, M.; Takakura, K.; Takahashi, S. Repair of DNA Damage Induced by Accelerated Heavy Ions in Mammalian Cells Proficient and Deficient in the Non-homologous End-Joining Pathway. Radiat. Res. 2006, 165, 59–67. [Google Scholar] [CrossRef]
- Levalois, M.; Bogdanski, P.; Toulemonde, M. Induced damage by high energy heavy ion irradiation at the GANIL accelerator in semiconductor materials. Nucl. Inst. Methods Phys. Res. B 1992, 63, 14–20. [Google Scholar] [CrossRef]
- Lewis, I.R.; Edwards, H.G.M. Handbook of Raman Spectroscopy: From the Research Laboratory to the Process Line; Marcel Dekker: New York, NY, USA, 2001; pp. 1–1054. ISBN 0-8247-0557-2. [Google Scholar]
- Camerlingo, C.; Portaccio, M.; Taté, R.; Lepore, M.; Delfino, I. Fructose and pectin detection in fruit-based food products by Surface-Enhanced Raman Spectroscopy. Sensors 2017, 17, 839. [Google Scholar] [CrossRef] [Green Version]
- Camerlingo, C.; Lisitskiy, M.; Lepore, M.; Portaccio, M.; Montorio, D.; Del Prete, S.; Cennamo, G. characterization of human tear fluid by means of Surface-enhanced Raman spectroscopy. Sensors 2019, 19, 1177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ZENODO. Quasars/Orange-Spectroscopy: Release 0.5.8. Available online: https://zenodo.org/record/4617978#.YOxVYugzaUk (accessed on 12 July 2021).
- Toplak, M.; Birarda, G.; Read, S.; Sandt, C.; Rosendahl, S.M.; Vaccari, L.; Demšar, J.; Borondics, F. Infrared Orange: Connecting Hyperspectral Data with Machine Learning. Synchrotron Radiat. News 2017, 30, 40–45. [Google Scholar] [CrossRef]
- Setlow, P. Summer meeting 2013—When the sleepers wake: The germination of spores of Bacillus species. J. Appl. Microbiol. 2013, 115, 1251–1268. [Google Scholar] [CrossRef] [PubMed]
- Farquharson, S.; Shende, C.; Gift, A.; Inscore, F. Detection of Bacillus Spores by Surface-Enhanced Raman Spectroscopy. 2012. Available online: http://www.intechopen.com/books/bioterrorism/detection-ofbacillus-spores-by-surface-enhanced-raman-spectroscopy (accessed on 26 July 2021).
- Gautam, R.; Chandrasekar, B.; Deobagkar-Lele, M.; Rakshit, S.; Umapathy, S.; Nandi, D. Identification of early biomarkers during acetaminophen-induced hepatotoxicity by Fourier transform infrared microspectroscopy. PLoS ONE 2012, 7, e45521. [Google Scholar] [CrossRef] [Green Version]
- Li, J.Y.; Ying, G.G.; Jones, K.C.; Martin, F.L. Real-world carbon nanoparticle exposures induce brain and gonadal alterations in zebrafish (Danio rerio) as determined by biospectroscopy techniques. Analyst 2015, 140, 2687–2695. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Verma, T.; Mukherjee, R.; Ariese, F.; Somasundaram, K.; Umapathy, S. Raman and infrared microspectroscopy: Towards quantitative evaluation for clinical research by ratiometric analysis. Chem. Soc. Rev. 2016, 45, 1879–1900. [Google Scholar] [CrossRef]
- Belskaya, L.V. Use of IR spectroscopy in cancer diagnosis. A review. J. Appl. Spectrosc. 2019, 86, 187–205. [Google Scholar] [CrossRef]
- Zucchiatti, P.; Latella, K.; Birarda, G.; Vaccari, L.; Rossi, B.; Gessini, A.; Masciovecchio, C.; D’Amico, F. The quality is in the eye of the beholder: The perspective of FTIR and UV resonant Raman spectroscopies on extracted nucleic acids. J. Raman Spectrosc. 2018, 49, 1056–1065. [Google Scholar] [CrossRef]
- Johnson, T.; Williams, S.; Valentine, N.; Su, Y. The hydration number n of calcium dipicolinate trihydrate, CaDP·nH2O, and its effect on the IR spectra of sporulated Bacillus bacteria. Vib. Spectrosc. 2010, 53, 28–33. [Google Scholar] [CrossRef]
- Gómez-Ordóñez, E.; Rupérez, P. FTIR-ATR spectroscopy as a tool for polysaccharide identification in edible brown and red seaweeds. Food Hydrocoll. 2011, 25, 1514–1520. [Google Scholar] [CrossRef]
- Marcotte, L.; Kegelaer, G.; Sandt, C.; Barbeau, J.; Lafleur, M. An alternative infrared spectroscopy assay for the quantification of polysaccharides in bacterial samples. Anal. Biochem. 2007, 361, 7–14. [Google Scholar] [CrossRef]
- Jackson, M.; Mantsch, H.H. The Use and Misuse of FTIR Spectroscopy in the Determination of Protein Structure. Crit. Rev. Biochem. Mol. Biol. 1995, 30, 95–120. [Google Scholar] [CrossRef]
- Reineke, K.; Schlumbach, K.; Baier, D.; Mathys, A.; Knorr, D. The release of dipicolinic acid—The rate-limiting step of Bacillus endospore inactivation during the high pressure thermal sterilization process. Int. J. Food Microbiol. 2013, 162, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Cucinotta, F.; Durante, M. Cancer risk from exposure to galactic cosmic rays: Implications for space exploration by human beings. Lancet Oncol. 2006, 7, 431–435. [Google Scholar] [CrossRef] [Green Version]
- Ross, S.M. Special Random Variables. In Introduction to Probability and Statistics for Engineers and Scientists; Sheldon, M.R., Ed.; Elsevier: Amsterdam, The Netherlands, 2004; pp. 141–200. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ion | Energy (MeV/n) | LET (keV/µm) | Intensity (pps) | Dose Rate (Gy/min) | Delivered Doses (Gy)/Irradiation’s Duration (min) |
|---|---|---|---|---|---|
| He | 150 | 2.2 | 1.2 × 1010 | 4.2 | 250/59.5, 1000/82.6 |
| Ar | 500 | 90 | 2.4 × 108 | 6.8 | 250/36.8, 500/73.5, 1000/147.1, 1500/220 |
| Fe | 500 | 200 | 2.5 × 108 | 12.1 | 250, 500, 1000, 1500, 2000 |
| ion | M1(amu) | Z1 | Normalized σ |
|---|---|---|---|
| He | 4 | 2 | 4 × 10−4 |
| Ar | 40 | 18 | 0.34 |
| Fe | 55.9 | 26 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Romano, I.; Camerlingo, C.; Vaccari, L.; Birarda, G.; Poli, A.; Fujimori, A.; Lepore, M.; Moeller, R.; Di Donato, P. Effects of Ionizing Radiation and Long-Term Storage on Hydrated vs. Dried Cell Samples of Extremophilic Microorganisms. Microorganisms 2022, 10, 190. https://doi.org/10.3390/microorganisms10010190
Romano I, Camerlingo C, Vaccari L, Birarda G, Poli A, Fujimori A, Lepore M, Moeller R, Di Donato P. Effects of Ionizing Radiation and Long-Term Storage on Hydrated vs. Dried Cell Samples of Extremophilic Microorganisms. Microorganisms. 2022; 10(1):190. https://doi.org/10.3390/microorganisms10010190
Chicago/Turabian StyleRomano, Ida, Carlo Camerlingo, Lisa Vaccari, Giovanni Birarda, Annarita Poli, Akira Fujimori, Maria Lepore, Ralf Moeller, and Paola Di Donato. 2022. "Effects of Ionizing Radiation and Long-Term Storage on Hydrated vs. Dried Cell Samples of Extremophilic Microorganisms" Microorganisms 10, no. 1: 190. https://doi.org/10.3390/microorganisms10010190