The Influence of Light and Nutrient Starvation on Morphology, Biomass and Lipid Content in Seven Strains of Green Microalgae as a Source of Biodiesel


Abstract
:1. Introduction
2. Materials and Methods
2.1. Microalgae Strains
2.2. Experimental Setup
2.3. Analytical Methods
2.4. Lipid Extraction and Fatty Acid Profile Characterization
2.5. Evaluation of Biodiesel Properties
2.6. Microscopy
2.7. Statistical Analysis
3. Results and Discussion
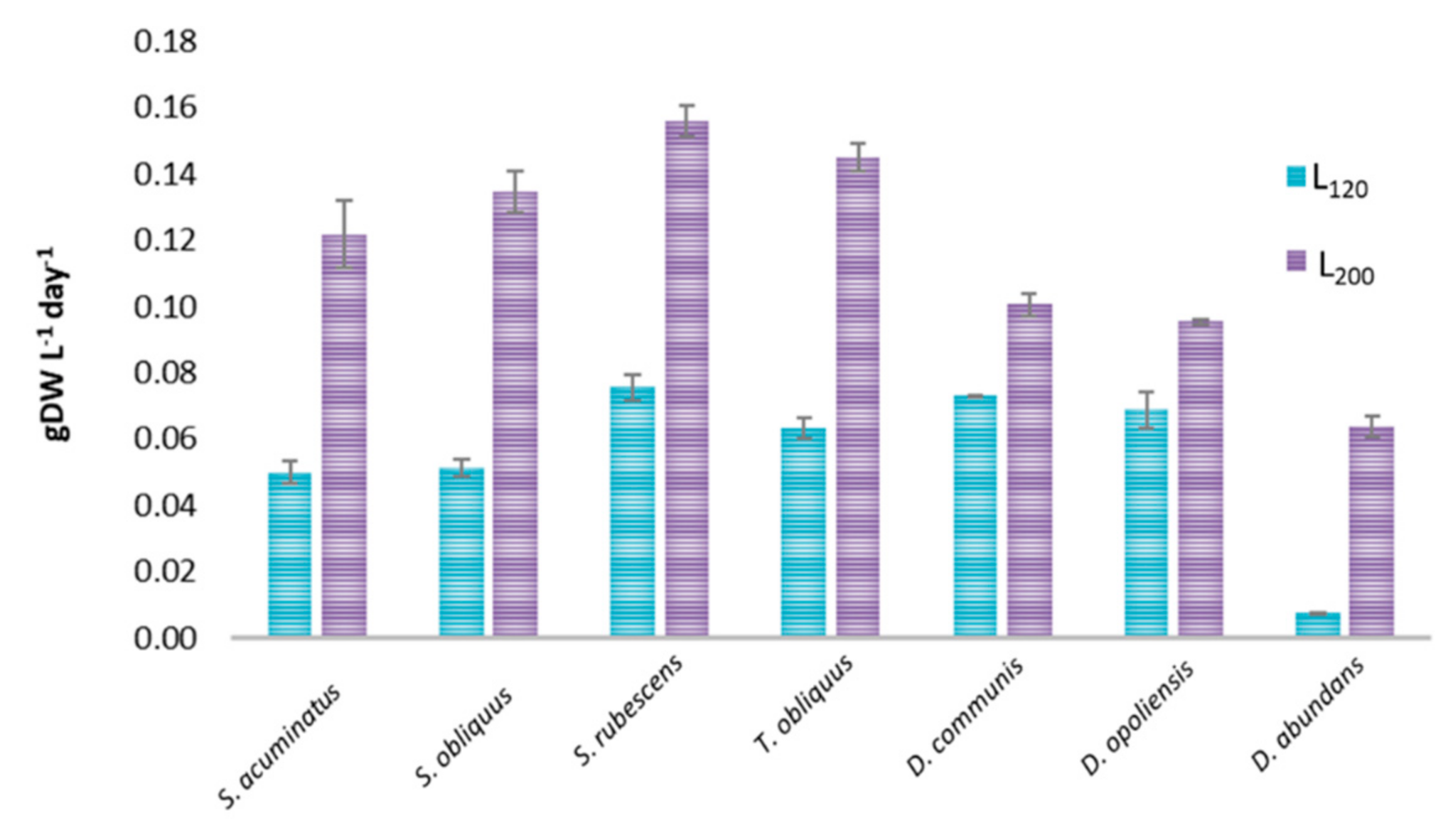
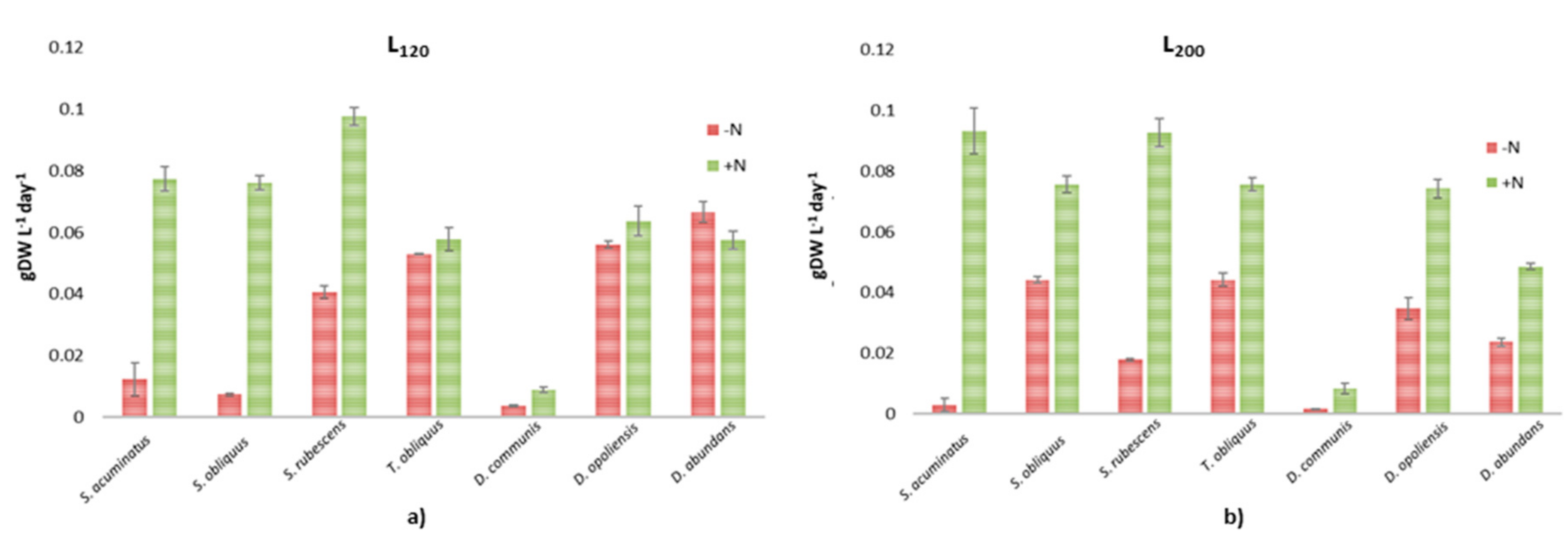
3.1. Growth Evaluation and Biomass Productivity
3.2. Lipid Yield and FAME Characterization
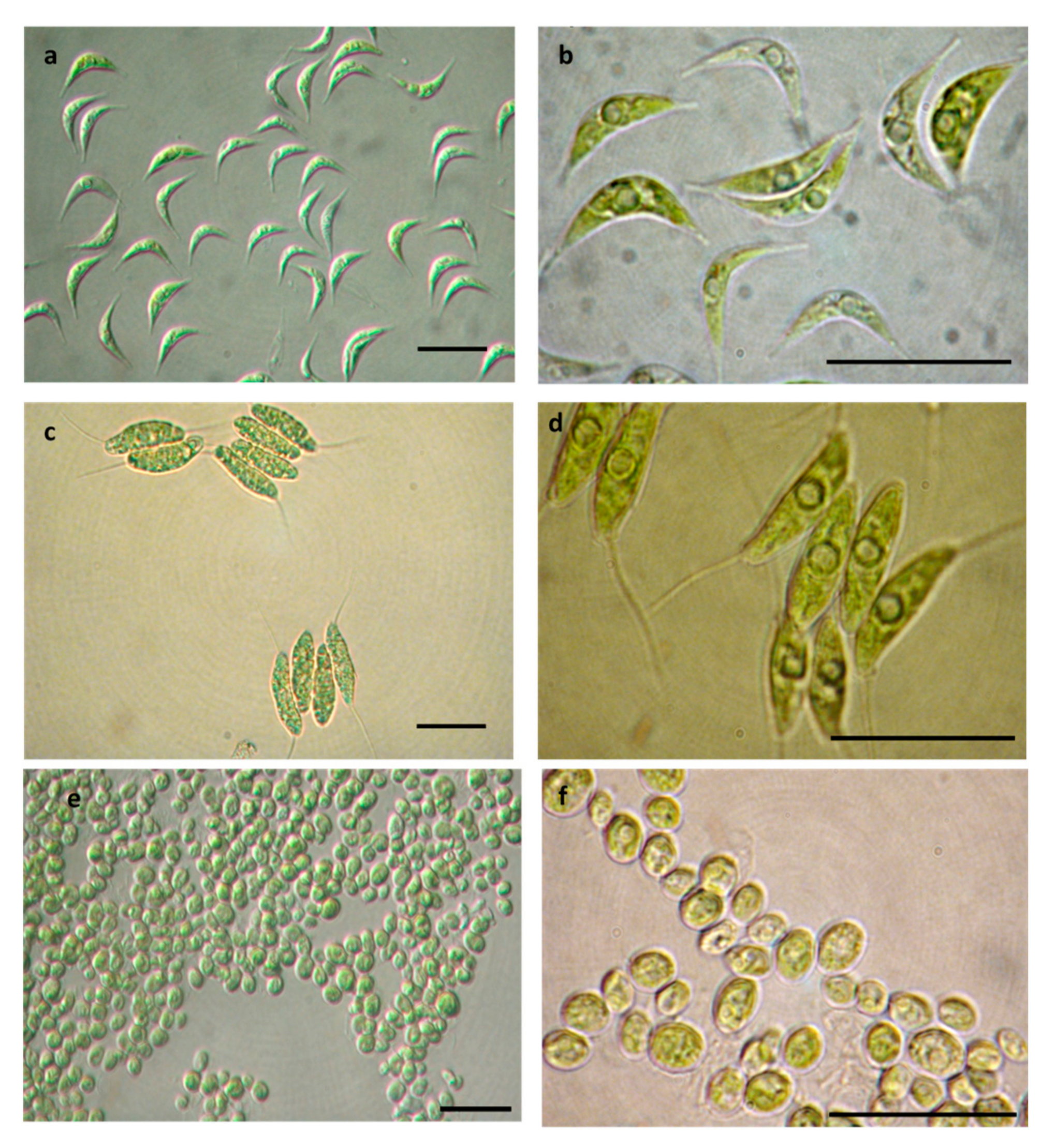
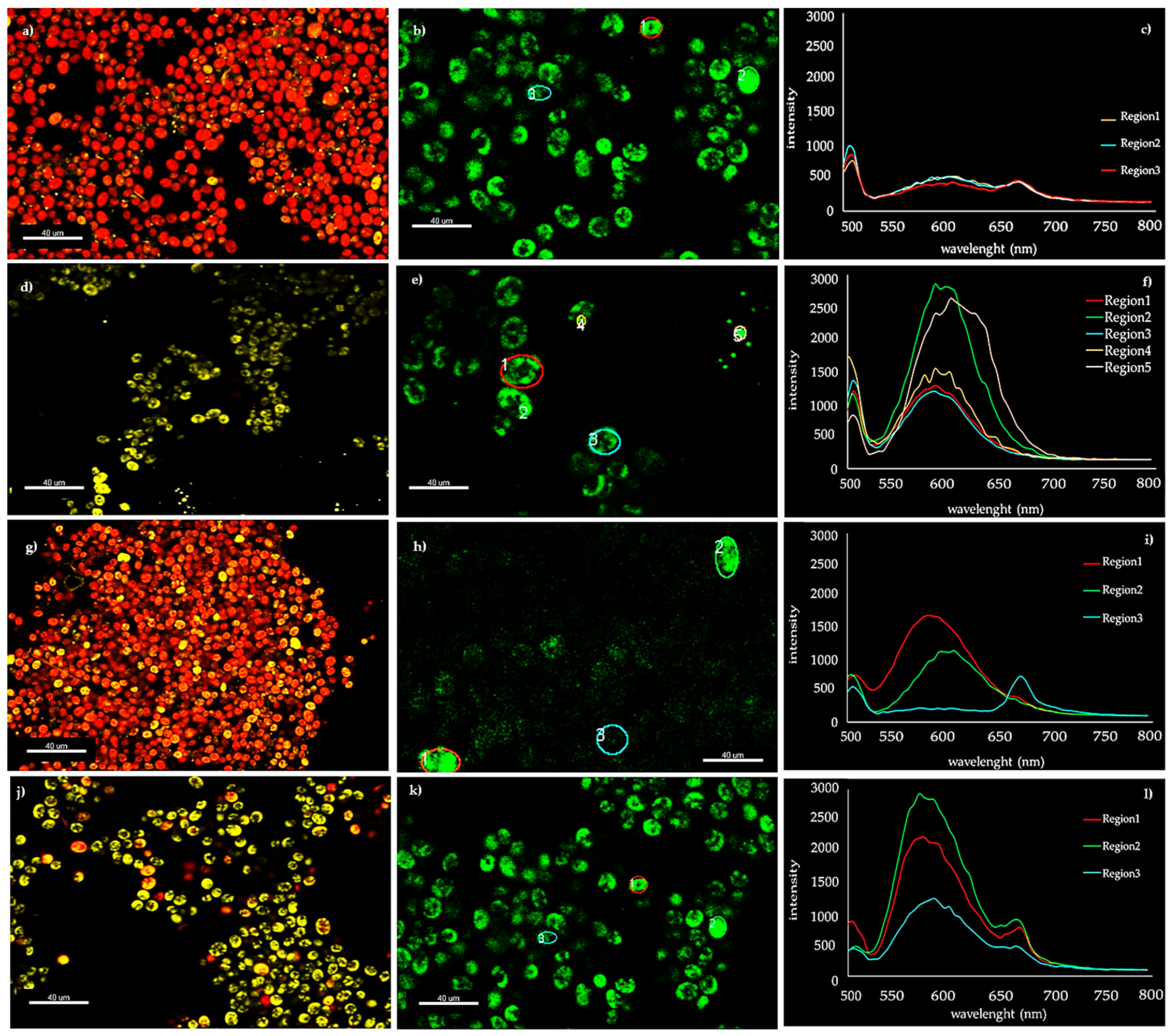
3.3. Microscopy Observations
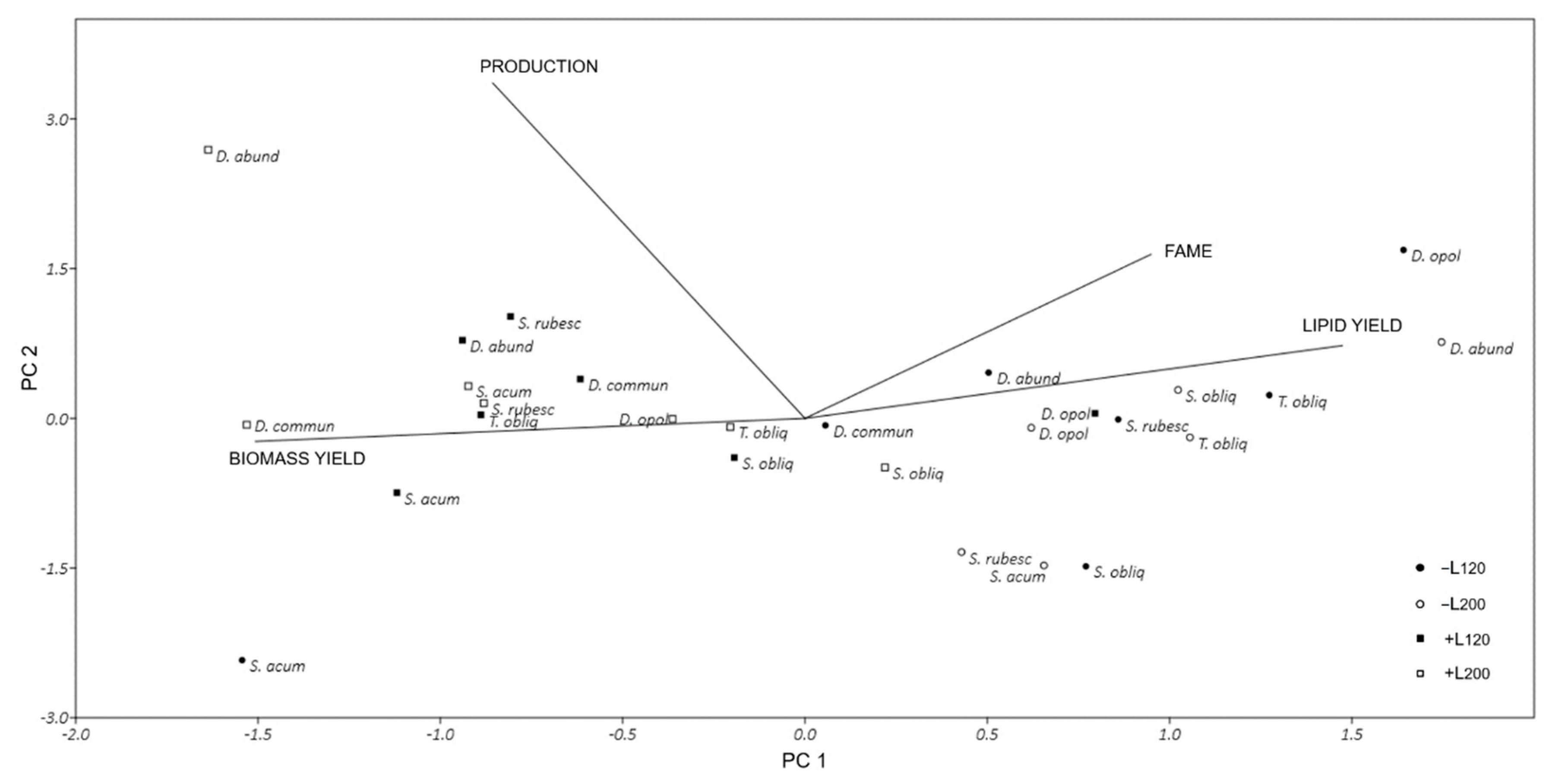
3.4. Multivariate Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Arora, N.; Tripathi, S.; Pruthi, P.A.; Poluri, K.M.; Pruthi, V. Assessing the robust growth and lipid-accumulating characteristics of Scenedesmus sp. for biodiesel production. Environ. Sci. Pollut. Res. 2019. [Google Scholar] [CrossRef] [PubMed]
- Maity, J.P.; Bundschuh, J.; Chen, C.Y.; Bhattacharya, P. Microalgae for third generation biofuel production, mitigation of greenhouse gas emissions and wastewater treatment: Present and future perspectives—A mini review. Energy 2014, 104–113. [Google Scholar] [CrossRef]
- Ghorbani, A.; Rahimpour, M.R.; Ghasemi, Y.; Raeissi, S. The biodiesel of microalgae as a solution for diesel demand in Iran. Energies 2018, 11, 950. [Google Scholar] [CrossRef] [Green Version]
- Nzayisenga, J.C.; Farge, X.; Groll, S.L.; Sellstedt, A. Effects of light intensity on growth and lipid production in microalgae grown in wastewater. Biotechnol. Biofuels 2020, 13. [Google Scholar] [CrossRef]
- Ramos, M.J.; Fernàndez, C.M.; Casa, A.; Rodrìguez, L.; Pérez, A. Influence of fatty acid composition of raw materials on biodiesel properties. Bioresour. Technol. 2009, 100, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Mata, T.M.; Martins, A.A.; Caetano, N.S. Microalgae for biodiesel production: A mini-review. Renew. Sustain. Energy Rev. 2010, 14, 217–232. [Google Scholar] [CrossRef] [Green Version]
- Akgul, F. Effects of nitrogen concentration on growth, biomass, and biochemical composition of Desmodesmus communis (E. Hegewald) E. Hegewald. Prep. Biochem. 98–105. [CrossRef]
- Rugnini, L.; Costa, G.; Congestri, R.; Antonaroli, S.; Sanità di Toppi, L.; Bruno, L. Phosphorus and metal removal combined with lipid production by the green microalga Desmodesmus sp.: An integrated approach. Plant. Physiol. Biochem. 2018, 125, 45–51. [Google Scholar] [CrossRef]
- Hernandez-Garcıa, A.; Velasquez-Orta, S.B.; Novelo, E.; Yanez-Noguez, I.; Monje-Ramırez, I.; Orta Ledesma, M.T. Wastewater-leachate treatment by microalgae: Biomass, carbohydrate and lipid production. Ecotoxico. Environ. Saf. 2019, 174, 435–444. [Google Scholar] [CrossRef]
- Aratboni, H.; Rafiei, N.; Garcia-Granados, R.; Alemzadeh, A.; Morones-Ramírez, J.R. Biomass and lipid induction strategies in microalgae for biofuel production and other applications. Microb. Cell Factories 2019, 18, 178. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Li, Z.; Hiltunen, E. Strategies for lipid production improvement in microalgae as a biodiesel feedstock. Biomed. Res. Int. 2016, 8. [Google Scholar] [CrossRef] [Green Version]
- Gouveia, L.; Oliveira, A.C.; Congestri, R.; Bruno, L.; Soare, A.T.; Menezes, R.S.; Filho, N.R.A.; Tzoveni, I. Biodiesel from microalgae. In Microalgae-Based Biofuels and Bioproducts, from Feedstock Cultivation to End-Products; Gonzalez-Fernadez, C., Munoz, R., Eds.; Woodhead Publishing Series in Energy: Duxford, UK, 2017. [Google Scholar]
- Pancha, I.; Chokshi, K.; George, B.; Ghosh, T.; Paliwal, C.; Maurya, R.; Mishra, S. Nitrogen stress triggered biochemical and morphological changes in the microalgae Scenedesmus sp. CCNM 1077. Bioresour. Technol. 2014, 156, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Griffith, M.J.; Van Hille, R.P.; Harrison, S.T.L. Lipid productivity, settling potential and fatty acid profile of 11 microalgal species grown under nitrogen replete and limited conditions. J. Appl. Phycol. 2012, 24, 989–1001. [Google Scholar] [CrossRef]
- Nascimento, I.A.; Marques, S.S.I.; Cabanelas, I.T.D.; Pereira, S.A.; Druzian, J.I.; de Souza, C.O.; Vich, D.V.; de Carvalho, G.C.; Nascimento, M.A. Screening microalgae strains for biodiesel production: Lipid productivity and estimation of fuel quality based on fatty acids profiles as selective criteria. Bioenerg. Res. 2013, 6, 1–13. [Google Scholar] [CrossRef]
- Gouveia, L.; Oliveira, A.C. Microalgae as raw material for biofuels production. J. Ind. Microbiol. Biotechnol. 2009, 6, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Rugnini, L.; Elwood, N.T.W.; Costa, G.; Falsetti, A.; Congestri, R.; Bruno, L. Scaling-up of wastewater bioremediation by Tetradesmus obliquus, sequential bio-treatments of nutrients and metals. Ecotoxicol. Environ. Saf. 2019, 172, 59–64. [Google Scholar] [CrossRef]
- Castenholz, R.W. Cyanobacteria. Oxygenic photosynthetic bacteria. In Bergey’s Manual of Systematic Bacteriology, 2nd ed.; Springer: New York, NY, USA, 2001; pp. 473–487. [Google Scholar]
- Starr, R.C.; Zeikus, J.A. UTEX—The culture collection of algae at the University of Texas at Austin. J. Phycol. 1987, 29, 1–106. [Google Scholar] [CrossRef]
- Bannon, C.D.; Breen, G.J.; Craske, J.D.; Trong, H.N.; Harper, N.L.; O’Rourke, K.L. Analysis of fatty acid methyl esters with high accuracy and reliability. iii. Literature review of and investigations into the development of rapid procedures for the methoxide-catalysed methanolysis of fats and oils. J. Chrom A 1982, 247, 71–89. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Greenspan, P.; Fowler, S.D. Spectrofluorometric studies of the lipid probe, nile red. J. Lipid Res. 1985, 26, 781–789. [Google Scholar]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Sforza, E.; Gris, B.; de Farias Silva, C.E.; Morosinotto, T.; Bertucco, A. Effects of light on cultivation of Scenedesmus Obliquus in batch and continuous flat plate photobioreactor. Chem. Eng. Trans. 2014, 38, 211–216. [Google Scholar]
- Wu, Y.H.; Yu, Y.; Hu, H.Y. Effects of initial phosphorus concentration and light intensity on biomass yield per phosphorus and lipid accumulation of Scenedesmus sp. LX1. Bioenergy Res. 2014. [Google Scholar] [CrossRef]
- Han, S.F.; Jin, W.; Abomohra., A.E.F.; Tu, R.; Zhou, X.; He, Z.; Chen, C.; Xie, G. Municipal wastewater enriched with trace metals for enhanced lipid production of the biodiesel-promising microalga Scenedesmus obliquus. BioEnergy Res. 2019. [Google Scholar] [CrossRef]
- Maheshwari, N.; Krishna, P.K.; Thakur, I.S.; Srivastava, S. Biological fixation of carbon dioxide and biodiesel production using microalgae isolated from sewage waste water. Environ. Sci. Pollut. Res. 2019, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Lang, I.; Hodac, L.; Friedl, T.; Feussner, I. Fatty acid profiles and their distribution patterns in microalgae: A comprehensive analysis of more than 2000 strains from the SAG culture collection. BMC Plant. Biol. 2011, 11, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quintana, N.; Van der Kooy, F.; Van de Rhee, M.D.; Voshol, G.P.; Verpoorte, R. Renewable energy from Cyanobacteria: Energy production optimization by metabolic pathway engineering. Appl. Microbiol. Biotechnol. 2011, 91, 471–490. [Google Scholar] [CrossRef] [Green Version]
- Bruno, L.; Di Pippo, F.; Antonaroli, S.; Gismondi, A.; Valentini, C.; Albertano, P. Characterization for biofilm-forming cyanobacteria for biomass and lipid production. J. Appl. Microbiol. 2012, 113, 1052–1064. [Google Scholar] [CrossRef] [Green Version]
- Mandotra, S.K.; Kumar, P.; Suseela, M.R.; Ramteke, P.W. Fresh water green microalga Scenedesmus abundans: A potential feedstock for high quality biodiesel production. Bioresour. Technol. 2014, 156, 42–47. [Google Scholar] [CrossRef]
- Pereira, H.; Barreira, L.; Custódio, L.; Alrokayan, S.; Mouffouk, F.; Varela, J.; Abu-Salah, K.M.; Ban-Hamadou, R. Isolation and fatty acid profile of selected microalgae strains from the Red sea for biofuel production. Energies 2013, 6, 2773–2783. [Google Scholar] [CrossRef] [Green Version]
- eur-lex.europa.eu. Available online: https://eur-lex.europa.eu/LexUriServ/LexUriServ.do?uri=OJ:L:2003:123:0042:0046:EN:PDF (accessed on 20 July 2020).
- ASTM. Standard specification for biodiesel fuel (B100) blend stock for distillate fuels. In Annual Book of ASTM Standard, Method D6751; ASTM International: West Conshohocken, PA, USA, 2008. [Google Scholar]
- Vishwakarma, R.; Dhar, D.W.; Saxena, S. Influence of nutrient formulations on growth, lipid yield, carbon partitioning and biodiesel quality potential of Botryococcus sp. and Chlorella sp. Environ. Sci. Pollut. Res. 2019, 26, 7589–7600. [Google Scholar] [CrossRef] [PubMed]
- Gismondi, A.; Di Pippo, F.; Bruno, L.; Antonaroli, S.; Congestri, R. Phosphorus removal coupled to bioenergy production by three cyanobacterial isolates in a biofilm dynamic growth system. Int. J. Phytoremediation 2016, 18, 869–876. [Google Scholar] [CrossRef] [PubMed]
- Komarek, J.; Foot, B. Chlorophyceae (Grünalgen), Ordung: Chlorococcales. In Das Phytoplankton des Sübwassers: Systematik und Biologie; Huber-Pestalozzi, G., Ed.; E. Schweizerbart’sche Verlagsbuchlandlung (Nägele u. Obermiller): Stuttgart, Germany, 1983; pp. 1–1044. [Google Scholar]
- Lurling, M. Phenotypic plasticity in the green algae Desmodesmus and Scenedesmus with special reference to the induction of defensive morphology. Ann. Limnol. Int. J. Lim. 2003, 39, 85–101. [Google Scholar] [CrossRef] [Green Version]
- El Semary, N.A. The polyphasic description of a Desmodesmus spp. Isolate with the potential of bioactive compounds production. Biotechnol. Agron. Soc. Environ. 2011, 15, 231–238. [Google Scholar]
- Yap, B.H.J.; Crawford, S.A.; Dagastine, R.R. Nitrogen deprivation of microalgae: Effect on cell size, cell wall thickness, cell strength, and resistance to mechanical disruption. J. Ind. Microbiol. Biotechnol. 2016, 43, 1671–1680. [Google Scholar] [CrossRef]
- Rumin, J.; Bonnefond, H.; Saint-Jeane, B.; Rouxel, C.; Sciandra, A.; Bernard, O.; Cadoret, J.P.; Bougaran, G. The use of fluorescent Nile red and BODIPY for lipid measurement in microalgae. Biotechnol. Biofuels 2015, 8, 42. [Google Scholar] [CrossRef] [Green Version]
- Sharma, J.; Kumar, S.K.; Bishnoi, N.R.; Pugazhendhi, A. Screening and enrichment of high lipid producing microalgal consortia. J. Photochem. Photobiol. B Biol. 2019, 192, 8–12. [Google Scholar] [CrossRef]
- Roldàn, M.; Thomas, F.; Castel, S.; Quesada, A.; Hernàndez-Mariné, M. Non-invasive pigment identification in single cells from living phototrophic biofilms by confocal imaging spectrofluorometry. Appl. Environ. Microbiol. 2004, 70, 3745–3750. [Google Scholar] [CrossRef] [Green Version]
- Gusbeth, C.A.; Eing, C.; Göttel, M.; Sträßner, R.; Wolfgang, F. Fluorescence diagnostics for lipid status monitoring of microalgae during cultivation. Int. J. Renew. Energy Biofuels 2016, 899698. [Google Scholar] [CrossRef] [Green Version]
- Ravindran, B.; Gupta, S.K.; Cho, W.-M.; Kim, J.K.; Lee, S.R.; Jeong, K.-H.; Lee, D.J.; Choi, H.-C. Microalgae potential and multiple roles—Current progress and future prospects—An overview. Sustainability 2016, 8, 1215. [Google Scholar] [CrossRef] [Green Version]
- De la Hoz Siegler, H.; Ben-Zvi, A.; McCaffrey, W.C. Improving the reliability of fluorescence-based neutral lipid content measurements in microalgal cultures. Algal Res. 2012, 1, 176–184. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. acuminatus | S. obliquus | S. rubescens | T. obliquus | D. abundans | D. communis | D. opoliensis | ||
|---|---|---|---|---|---|---|---|---|
| BBM-N/L120 | gDW L−1 | 0.08 ± 0.00 | 0.05 ± 0.02 | 0.36 ± 0.01 | 0.37 ± 0.01 | 0.52 ± 0.05 | 0.62 ± 0.03 | 0.73 ± 0.01 |
| FAME (%) | 4.56 | 13.01 | 18.80 | 22.26 | 22 | 8.67 | 23.00 | |
| BBM+N/L120 | gDW L−1 | 0.54 ± 0.01 | 0.53 ± 0.03 | 0.88 ± 0.04 | 0.40 ± 0.02 | 0.92 ± 0.09 | 0.70 ± 0.01 | 0.63 ± 0.03 |
| FAME (%) | 6.60 | 8.37 | 16.13 | 11.52 | 10.51 | 16.29 | 3.53 | |
| BBM-N/L200 | gDW L−1 | 0.02 ± 0.01 | 0.35 ± 0.02 | 0.14 ± 0.01 | 0.35 ± 0.01 | 0.27 ± 0.01 | 0.42 ± 0.02 | 0.28 ± 0.01 |
| FAME (%) | 16.88 | 7.04 | 11.92 | 14.79 | 34.60 | n.d. | 25.10 | |
| BBM+N/L200 | gDW L−1 | 0.75 ± 0.07 | 0.61 ± 0.01 | 0.74 ± 0.04 | 0.61 ± 0.02 | 1.41 ± 0.07 | 0.82 ± 0.02 | 0.58 ± 0.01 |
| FAME (%) | 12.24 | 14.34 | 8.34 | 8.85 | 17.88 | 3.93 | 13.75 |
| FAME | BBM-N/L120 | BBM+N/L120 | BBM-N/L200 | BBM+N/L200 | ||||
|---|---|---|---|---|---|---|---|---|
| T. obliquus | D. abundans | T. obliquus | D. abundans | T. obliquus | D. abundans | T. obliquus | D. abundans | |
| C14:0 | 0.00 | 0.00 | 1.58 | 0.71 | 0.00 | 0.47 | 0.94 | 0.44 |
| C16:0 | 81.75 | 46.18 | 55.07 | 31.79 | 62.94 | 73.71 | 50.90 | 32.72 |
| C16:1 (9) | 0.00 | 0.66 | 2.85 | 0.92 | 0.00 | 1.39 | 1.19 | 1.16 |
| C18:0 | 11.62 | 7.01 | 3.76 | 2.58 | 15.54 | 15.76 | 6.26 | 6.73 |
| C18:1 (trans-9) | 4.11 | 2.90 | 1.50 | 0.00 | 0.00 | 0.62 | 1.22 | 0.59 |
| C18:1 (cis-9) | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 42.29 |
| C18:2 (trans-9,12) | 2.52 | 0.00 | 0.00 | 8.85 | 0.00 | 0.00 | 31.45 | 0.00 |
| C18:2 (cis-9,12) | 0.00 | 0.00 | 0.00 | 13.83 | 0.00 | 0.00 | 6.55 | 6.36 |
| C18:3 (9,12,15) | 0.00 | 0.54 | 14.31 | 39.12 | 3.72 | 0.00 | 0.00 | 0.00 |
| C20:0 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 2.78 | 0.00 | 0.00 |
| C20:1 | 0.00 | 42.72 | 20.94 | 2.19 | 17.80 | 2.23 | 0.00 | 9.71 |
| C20:4 (5,8,11,14) | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 3.02 | 0.00 | 0.00 |
| SFA (%) | 93.37 | 53.19 | 60.41 | 35.09 | 78.48 | 92.73 | 58.10 | 39.89 |
| MUFA (%) | 4.11 | 46.27 | 25.29 | 3.11 | 17.80 | 4.25 | 2.41 | 53.75 |
| PUFA (%) | 2.52 | 0.54 | 14.31 | 61.80 | 3.72 | 3.02 | 38.00 | 6.36 |
| SV | 214.20 | 199.70 | 207.18 | 206.22 | 207.63 | 211.22 | 209.07 | 203.50 |
| IV | 8.08 | 39.56 | 60.42 | 150.86 | 24.74 | 12.46 | 39.56 | 59.16 |
| CN | 44.51 | 37.43 | 32.73 | 12.38 | 40.76 | 43.52 | 30.08 | 33.02 |
| CFPP (°C) | 27.46 | 9.04 | 6.73 | −2.43 | 27.71 | 40.19 | 9.04 | 4.37 |
| LCSF | 13.99 | 8.12 | 7.39 | 4.47 | 14.06 | 18.04 | 8.22 | 6.64 |
| DU | 9.15 | 47.35 | 53.90 | 126.71 | 25.24 | 10.29 | 76.41 | 66.48 |
| Growth Conditions | Length (µm) | ||
|---|---|---|---|
| Initial Phase | Stationary Phase | ||
| S. acuminatus | BBM-N/L120 | 20.55 ± 2.51 | 24.49 ± 2.90 |
| BBM+N/L120 | 19.68 ± 4.22 | 25.45 ± 3.09 | |
| BBM-N/L200 | 26.58 ± 3.71 | 26.75 ± 2.64 | |
| BBM+N/L200 | 25.72 ± 3.91 | 26.03 ± 3.92 | |
| S. obliquus | BBM-N/L120 | 12.69 ± 1.95 | 13.41 ± 3.18 |
| BBM+N/L120 | 15.05 ± 2.21 | 13.46 ± 2.58 | |
| BBM-N/L200 | 18.02 ± 2.83 | 14.73 ± 4.06 | |
| BBM+N/L200 | 16.22 ± 2.61 | 17.14 ± 4.30 | |
| S. rubescens | BBM-N/L120 | 12.04 ± 2.18 | 11.83 ± 2.36 |
| BBM+N/L120 | 12.07 ± 2.26 | 11.78 ± 2.30 | |
| BBM-N/L200 | 14.25 ± 3.01 | 13.58 ± 2.68 | |
| BBM+N/L200 | 13.72 ± 2.35 | 13.86 ± 1.74 | |
| T. obliquus | BBM-N/L120 | 13.84 ± 2.10 | 16.18 ± 2.41 |
| BBM+N/L120 | 16.23 ± 3.26 | 15.21 ± 1.28 | |
| BBM-N/L200 | 18.93 ± 2.27 | 18.28 ± 3.20 | |
| BBM+N L200 | 18.77 ± 3.14 | 15.27 ± 2.64 | |
| D. abundans | BBM-N/L120 | 5.85 ± 1.44 | 5.48 ± 1.08 |
| BBM+N/L120 | 6.33 ± 1.42 | 5.20 ± 1.15 | |
| BBM-N/L200 | 6.71 ± 1.54 | 8.13 ± 1.27 | |
| BBM+N/L200 | 6.32 ± 1.32 | 6.58 ± 1.16 | |
| D. communis | BBM-N/L120 | 22.74 ± 2.37 | 20.16 ± 2.63 |
| BBM+N/L120 | 22.76 ± 1.68 | 19.13 ± 2.42 | |
| BBM-N/L200 | 21.81 ± 3.65 | 21.65 ± 2.96 | |
| BBM+N/L200 | 21.23 ± 2.40 | 20.87 ± 2.64 | |
| D. opoliensis | BBM-N/L120 | 29.14 ± 3.08 | 29.88 ± 1.89 |
| BBM+N/L120 | 27.65 ± 3.69 | 24.02 ± 2.55 | |
| BBM-N/L200 | 21.55 ± 1.88 | 29.07 ± 1.68 | |
| BBM+N/L200 | 24.49 ± 2.42 | 24.17 ± 2.35 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rugnini, L.; Rossi, C.; Antonaroli, S.; Rakaj, A.; Bruno, L. The Influence of Light and Nutrient Starvation on Morphology, Biomass and Lipid Content in Seven Strains of Green Microalgae as a Source of Biodiesel. Microorganisms 2020, 8, 1254. https://doi.org/10.3390/microorganisms8081254
Rugnini L, Rossi C, Antonaroli S, Rakaj A, Bruno L. The Influence of Light and Nutrient Starvation on Morphology, Biomass and Lipid Content in Seven Strains of Green Microalgae as a Source of Biodiesel. Microorganisms. 2020; 8(8):1254. https://doi.org/10.3390/microorganisms8081254
Chicago/Turabian StyleRugnini, Lorenza, Catia Rossi, Simonetta Antonaroli, Arnold Rakaj, and Laura Bruno. 2020. "The Influence of Light and Nutrient Starvation on Morphology, Biomass and Lipid Content in Seven Strains of Green Microalgae as a Source of Biodiesel" Microorganisms 8, no. 8: 1254. https://doi.org/10.3390/microorganisms8081254