
Vitamin A5/X, a New Food to Lipid Hormone Concept for a Nutritional Ligand to Control RXR-Mediated Signaling

 , and
, and 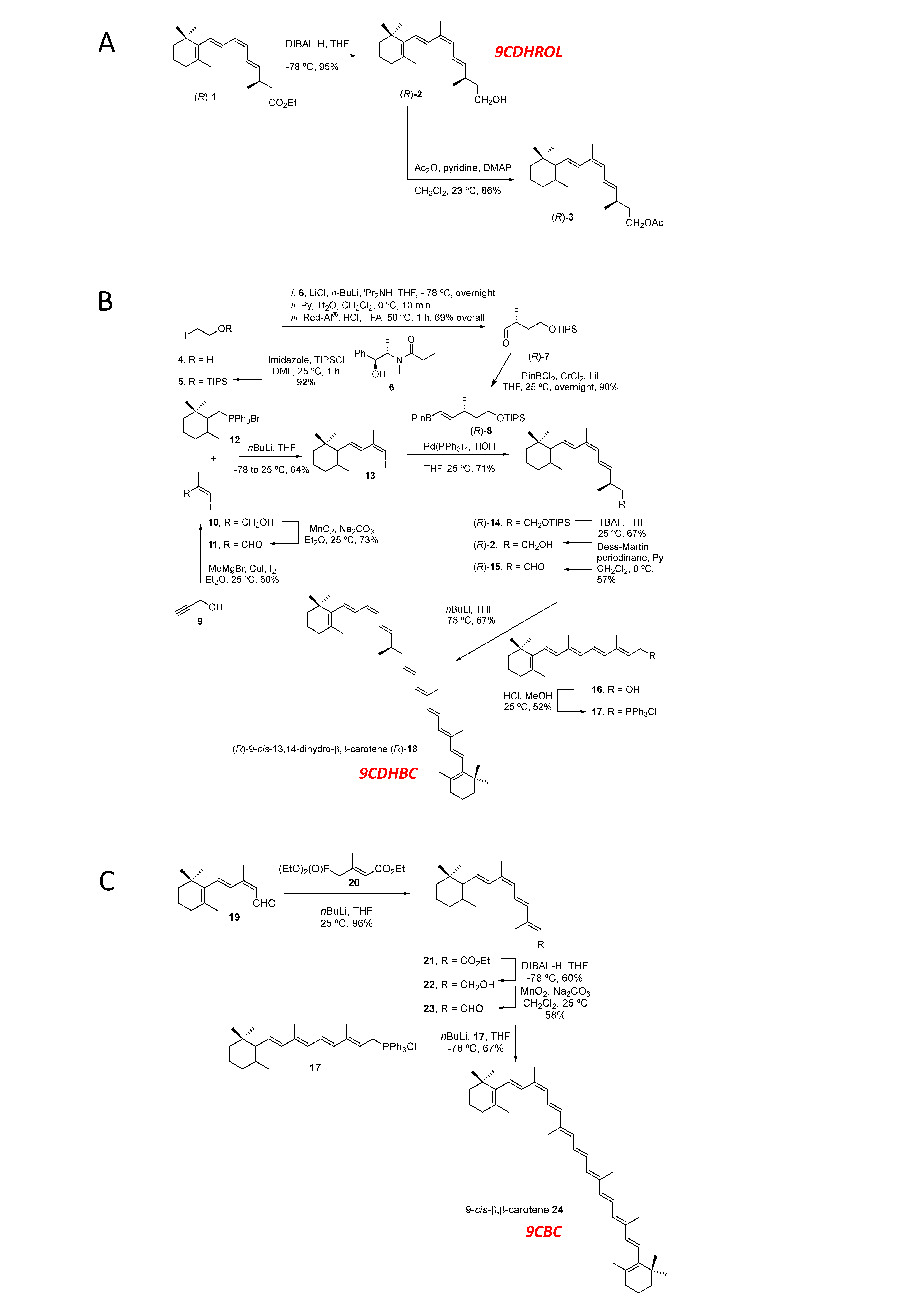{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Organic Chemical Synthesis
2.1.1. Synthesis of (R)-9-cis-13,14-Dihydroretinol (R)-(2) (9CDHROL) and of the Corresponding Acetate (R)-(3)
2.1.2. Synthesis of (R)-9-cis-13,14-Dihydro-β,β-Carotene (9CDHBC; Figure 1B, including Numbering)
2.1.3. Synthesis of 9-cis-β,β-Carotene (9CBC; Figure 1C, including Numbering)
2.2. Samples Used for LC-MS Analysis
2.3. Animal Experiments
2.3.1. Animals
2.3.2. Animal Treatments
2.3.3. Memory Analysis
2.4. Cell Cultures and Pharmacological Treatments
2.5. Liquid Chromatography-Mass Spectrometry (LC-MS) Analysis—Combined Retinoid and Carotenoid Analysis
2.6. Statistical Analysis
3. Results
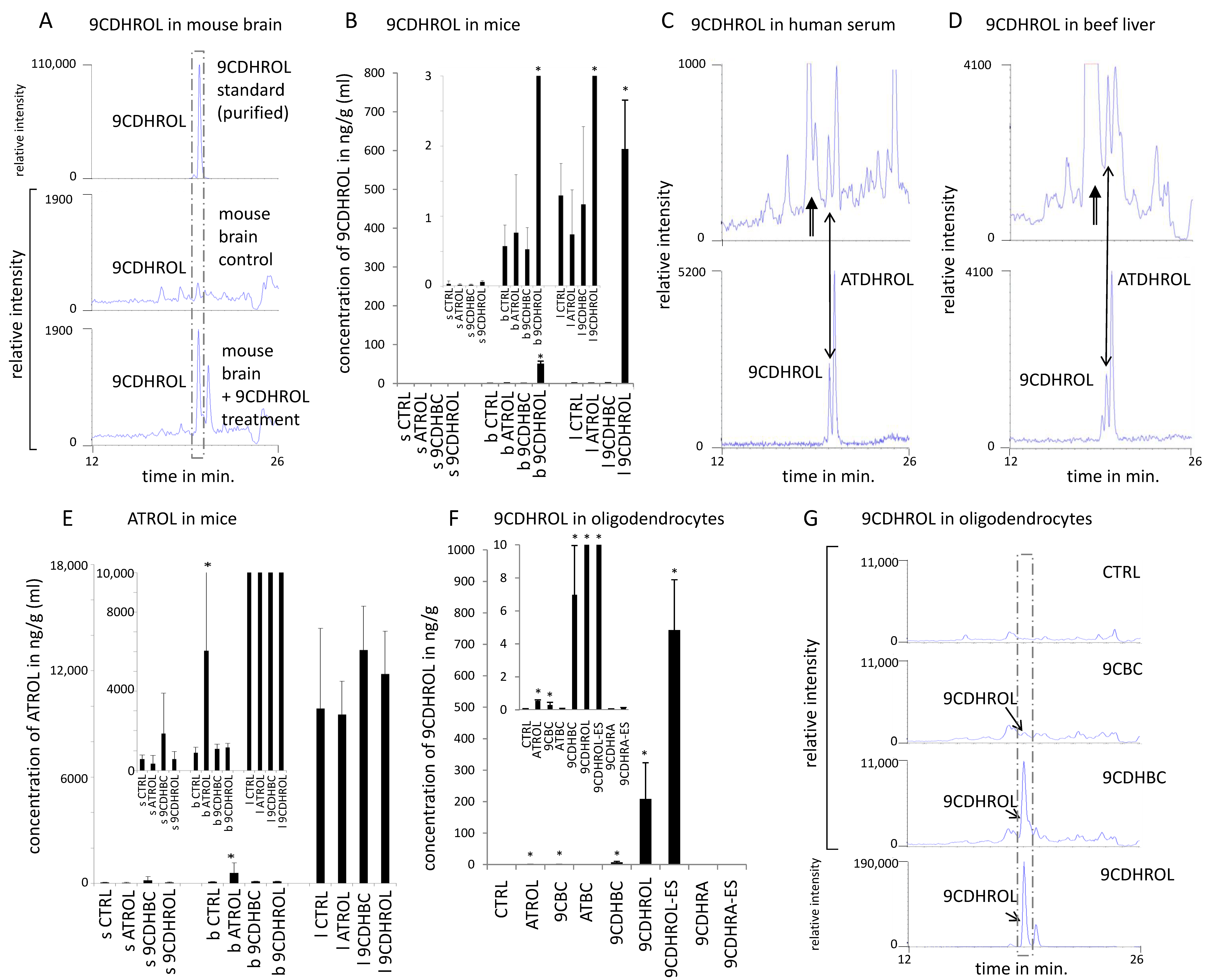
3.1. Identification of Endogenous 9CDHROL in Mouse, Human and the Human Food Matrix
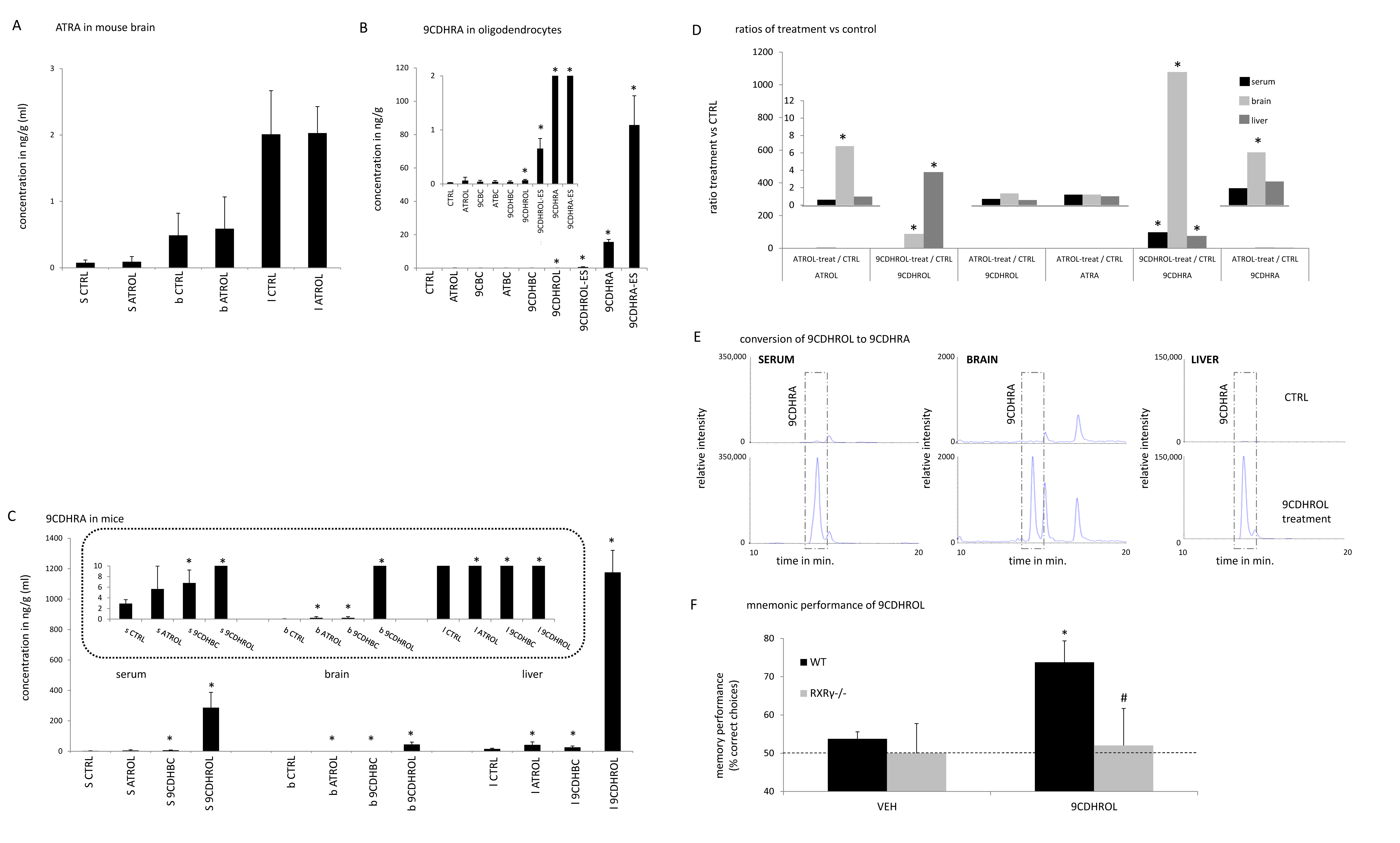
3.2. The Vitamin A5/X Concept: 9CDHROL, but Not ATROL, Is an Efficient Precursor of 9CDHRA in Mice
3.3. Vitamin A5/X-Specific Activity: RXR-Mediated Signaling Specific Activity of 9CDHROL
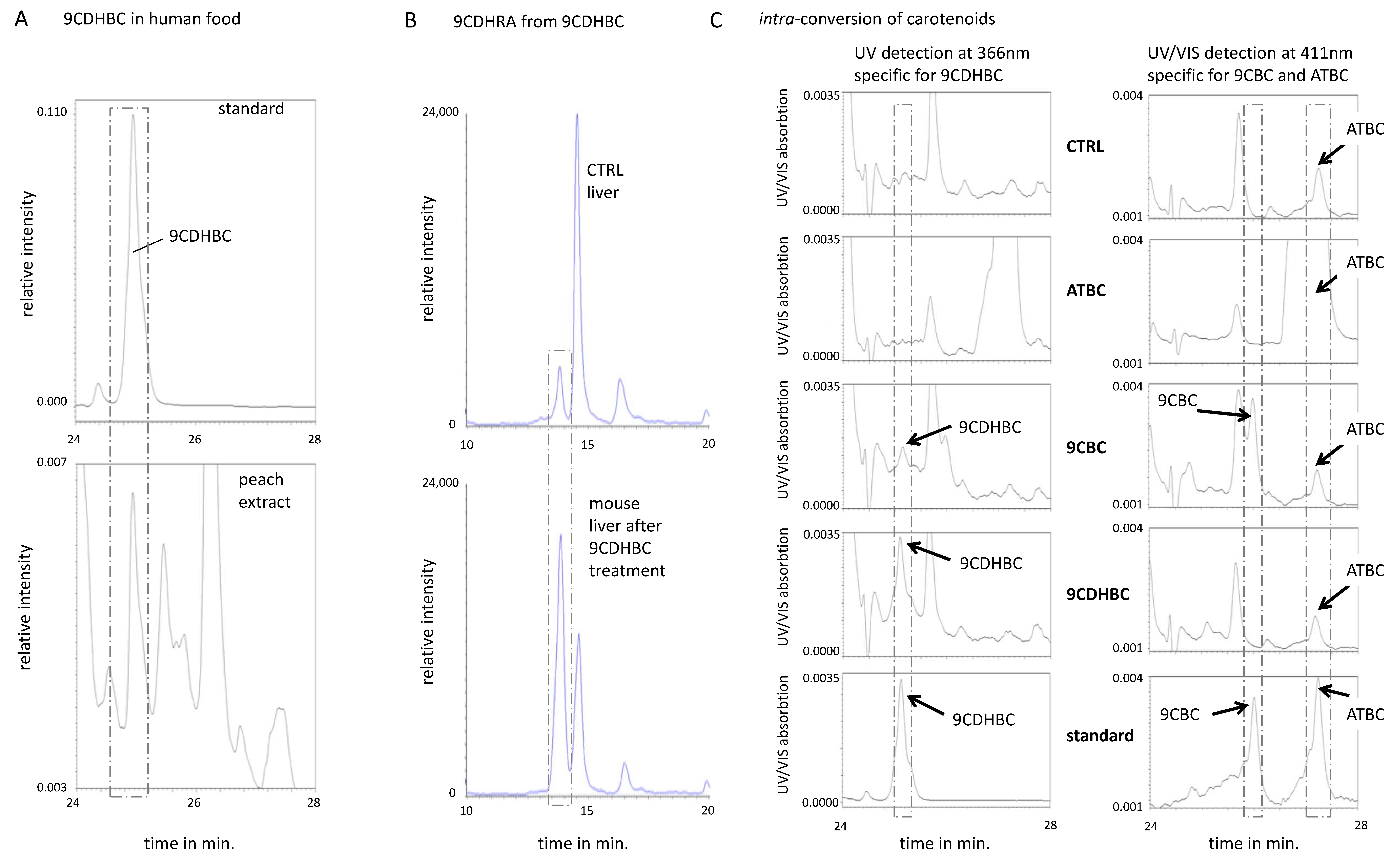
3.4. The Provitamin A5/X Concept: 9CDHBC Is a New Carotenoid of Plant Origin and an Efficient Precursor of 9CDHRA in Mice
3.5. Identification of 9-cis-13,14-Dihydroretinyl- and 9-cis-13,14-Dihydroretinoyl-Esters as Precursors for 9CDHRA in Cultured Oligodendrocytes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Desvergne, B. RXR: From partnership to leadership in metabolic regulations. Vitam. Horm. 2007, 75, 1–32. [Google Scholar] [CrossRef] [PubMed]
- Szántó, A.; Narkar, V.; Shen, Q.; Uray, I.P.; Davies, P.J.A.; Nagy, L. Retinoid X receptors: X-ploring their (patho)physiological functions. Cell Death Differ. 2004, 11, S126–S143. [Google Scholar] [CrossRef] [Green Version]
- Mangelsdorf, D.J.; Thummel, C.; Beato, M.; Herrlich, P.; Schütz, G.; Umesono, K.; Blumberg, B.; Kastner, P.; Mark, M.; Chambon, P.; et al. The nuclear receptor superfamily: The second decade. Cell 1995, 83, 835–839. [Google Scholar] [CrossRef] [Green Version]
- Mangelsdorf, D.J.; Evans, R.M. The RXR heterodimers and orphan receptors. Cell 1995, 83, 841–850. [Google Scholar] [CrossRef] [Green Version]
- Evans, R.M.; Mangelsdorf, D.J. Nuclear receptors, RXR, and the big bang. Cell 2014, 157, 255–266. [Google Scholar] [CrossRef] [Green Version]
- Petkovich, M.; Brand, N.J.; Krust, A.; Chambon, P. A human retinoic acid receptor which belongs to the family of nuclear receptors. Nat. Cell Biol. 1987, 330, 444–450. [Google Scholar] [CrossRef]
- Giguere, V.; Ong, E.S.; Segui, P.; Evans, R.M. Identification of a receptor for the morphogen retinoic acid. Nature 1987, 330, 624–629. [Google Scholar] [CrossRef]
- Heyman, R.A.; Mangelsdorf, D.J.; Dyck, J.A.; Stein, R.B.; Eichele, G.; Evans, R.M. Thaller C: 9-cis-retinoic acid is a high affinity ligand for the retinoid X receptor. Cell 1992, 68, 397–406. [Google Scholar] [CrossRef]
- Levin, A.A.; Sturzenbecker, L.J.; Kazmer, S.; Bosakowski, T.; Huselton, C.; Allenby, G.; Speck, J.; Kratzeisen, C.; Rosenberger, M.; Lovey, A.; et al. 9-cis-retinoic acid stereoisomer binds and activates the nuclear receptor RXR alpha. Nature 1992, 355, 359–361. [Google Scholar] [CrossRef]
- De Lera, A.R.; Krezel, W.; Rühl, R. An endogenous mammalian retinoid X receptor ligand, at last! ChemMedChem 2016, 11, 1027–1037. [Google Scholar] [CrossRef]
- Rühl, R.; Krzyzosiak, A.; Niewiadomska-Cimicka, A.; Rochel, N.; Szeles, L.; Vaz, B.; Wietrzych-Schindler, M.; Alvarez, S.; Szklenar, M.; Nagy, L.; et al. 9-cis-13,14-dihydroretinoic acid is an endogenous retinoid acting as RXR ligand in mice. PLoS Genet. 2015, 11, e1005213. [Google Scholar] [CrossRef] [Green Version]
- Rühl, R.; Krezel, W.; de Lera, A.R. 9-Cis-13,14-dihydroretinoic acid, a new endogenous mammalian ligand of retinoid X receptor and the active ligand of a potential new vitamin A category: Vitamin A5. Nutr. Rev. 2018, 76, 929–941. [Google Scholar] [CrossRef]
- Krężel, W.; Rühl, R.; de Lera, A.R. Alternative retinoid X receptor (RXR) ligands. Mol. Cell. Endocrinol. 2019, 491, 110436. [Google Scholar] [CrossRef]
- Krezel, W.; Ghyselinck, N.; Samad, T.A.; Dupe, V.; Kastner, P.; Borrelli, E.; Chambon, P. Impaired locomotion and dopamine signaling in retinoid receptor mutant mice. Science 1998, 279, 863–867. [Google Scholar] [CrossRef]
- Wietrzych, M.; Meziane, H.; Sutter, A.; Ghyselinck, N.; Chapman, P.F.; Chambon, P.; Krezel, W. Working memory deficits in retinoid X receptor gamma-deficient mice. Learn. Mem. 2005, 12, 318–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nomoto, M.; Takeda, Y.; Uchida, S.; Mitsuda, K.; Enomoto, H.; Saito, K.; Choi, T.; Watabe, A.M.; Kobayashi, S.; Masushige, S.; et al. Dysfunction of the RAR/RXR signaling pathway in the forebrain impairs hippocampal memory and synaptic plasticity. Mol. Brain 2012, 5, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiang, M.Y.; Misner, D.; Kempermann, G.; Schikorski, T.; Giguere, V.; Sucov, H.M.; Gage, F.H.; Ste-vens, C.F.; Evans, R.M. An essential role for retinoid receptors RARbeta and RXRgamma in long-term potentiation and depression. Neuron 1998, 21, 1353–1361. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, C.K.; Volland, J.; Hamscher, G.; Nau, H. Characterization of a new endogenous vitamin A metabolite. Biochim. Biophys. Acta BBA Mol. Cell Biol. Lipids 2002, 1583, 237–251. [Google Scholar] [CrossRef]
- Moise, A.R.; Kuksa, V.; Imanishi, Y.; Palczewski, K. Identification of All-trans-Retinol: All-trans-13,14-dihydroretinol Saturase. J. Biol. Chem. 2004, 279, 50230–50242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moise, A.R.; Dominguez, M.; Alvarez, S.; Alvarez, R.; Schupp, M.; Cristancho, A.G.; Kiser, P.D.; de Lera, A.R.; Lazar, M.A.; Palczewski, K. Stereospecificity of retinol saturase: Absolute configuration, synthesis, and biological evaluation of dihydroretinoids. J. Am. Chem. Soc. 2008, 130, 1154–1155. [Google Scholar] [CrossRef] [Green Version]
- Koukal, P.; Kotora, M. Enantioselective Allylation of (2E,4E)-2,4-Dimethylhexadienal: Synthesis of (5R,6S)-(+)-Pteroenone. Chemistry 2016, 21, 7408–7412. [Google Scholar] [CrossRef]
- Okitsu, T.; Iwatsuka, K.; Wada, A. Caesium fluoride-promoted Stille coupling reaction: An efficient synthesis of 9Z-retinoic acid and its analogues using a practical building block. Chem. Commun. Camb. 2008, 47, 6330–6332. [Google Scholar] [CrossRef]
- Englert, G. ChemInform Abstract: Carbon-13 nmr. Study of cis-trans isomeric vitamins a, carotenoids, and related compounds. Chem. Inf. 1976, 7. [Google Scholar] [CrossRef]
- Zanoun, A.; Bouazit, S.; Belaidi, A.; Vergoten, G. The SASIBA force field for some retinal conformers. J. Mol. Struct. THEOCHEM 2006, 777, 23. [Google Scholar] [CrossRef]
- Sher, I.; Tzameret, A.; Peri-Chen, S.; Edelshtain, V.; Ioffe, M.; Sayer, A.; Buzhansky, L.; Gazit, E.; Rotenstreich, Y. Synthetic 9-cis-beta-carotene inhibits photoreceptor degeneration in cultures of eye cups from rpe65rd12 mouse model of retinoid cycle defect. Sci. Rep. 2018, 8, 6130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koyama, Y.; Hosoni, M.; Hashimoto, H.; Shimamura, T. 1H NMR specrta of the all-trans 7-cis, 9-cis, 13-cis and 15-cis isomers of ß-carotene: Elongation of the double bound and shortening of the single bond towards the center of the conjugated chain as revealed by vicinal coupling constant. J. Mol. Struct. 1989, 193, 12. [Google Scholar] [CrossRef]
- Wietrzych-Schindler, M.; Szyszka-Niagolov, M.; Ohta, K.; Endo, Y.; Perez, E.; de Lera, A.R.; Cham-bon, P.; Krezel, W. Retinoid x receptor gamma is implicated in docosahexaenoic acid modulation of despair behaviors and working memory in mice. Biol. Psychiatry 2011, 69, 788–794. [Google Scholar] [CrossRef] [PubMed]
- Feutz, A.C.; Pham-Dinh, D.; Allinquant, B.; Miehe, M.; Ghandour, M.S. An immortalized jimpy oligodendrocyte cell line: Defects in cell cycle and cAMP pathway. Glia 2001, 34, 241–252. [Google Scholar] [CrossRef] [PubMed]
- Rühl, R. Method to determine 4-oxo-retinoic acids, retinoic acids and retinol in serum and cell extracts by liquid chromatography/diode-array detection atmospheric pressure chemical ionisation tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2006, 20, 2497–2504. [Google Scholar] [CrossRef]
- Stoney, P.N.; McCaffery, P. A vitamin on the mind: New discoveries on control of the brain by Vitamin A. Hidden Hunger 2016, 115, 98–108. [Google Scholar] [CrossRef]
- Shearer, K.D.; Stoney, P.N.; Morgan, P.J.; McCaffery, P.J. A vitamin for the brain. Trends Neurosci. 2012, 35, 733–741. [Google Scholar] [CrossRef] [PubMed]
- D’Ambrosio, D.N.; Clugston, R.D.; Blaner, W.S. Vitamin A metabolism: An update. Nutrients 2011, 3, 63–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blomhoff, R.; Blomhoff, H.K. Overview of retinoid metabolism and function. J. Neurobiol. 2006, 66, 606–630. [Google Scholar] [CrossRef]
- Bohn, T.; Desmarchelier, C.; El, S.N.; Keijer, J.; van Schothorst, E.M.; Rühl, R.; Borel, P. B-Carotene in the Human body—Metabolic acitivation pathways—from Digestion to Tissue Distribution. Proc. Nutr. Soc. 2019, 78, 68–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.K.; Jarjour, A.A.; Nait Oumesmar, B.; Kerninon, C.; Williams, A.; Krezel, W.; Kagechika, H.; Bauer, J.; Zhao, C.; Baron-Van Evercooren, A.; et al. Retinoid X receptor gamma signaling accelerates CNS remyelination. Nat. Neurosci. 2011, 14, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Stahl, W.; Schwarz, W.; Sundquist, A.R.; Sies, H. Cis-trans isomers of lycopene and beta-carotene in human serum and tissues. Arch. Biochem. Biophys. 1992, 294, 173–177. [Google Scholar] [CrossRef]
- Khoo, H.E.; Prasad, K.N.; Kong, K.W.; Jiang, Y.; Ismail, A. Carotenoids and their isomers: Color pigments in fruits and vegetables. Molecules 2011, 16, 1710–1738. [Google Scholar] [CrossRef]
- Lessin, J.W.; Catigani, G.L.; Schwartz, S.J. Quantification of cis-trans isomers of Provitamin A carotenoids in fresh and processed fruits and vegetables. J. Agric. Food Chem. 1997, 45, 3728–3732. [Google Scholar] [CrossRef]
- Ben-Amotz, A.; Fishler, R. Analysis of carotenoids with emphasis on 9-cis-beta-carotene in vegetables and fruits commonly consumed in israel. Food Chem. 1998, 62, 515–520. [Google Scholar] [CrossRef]
- Abuauf, H.; Haider, I.; Jia, K.P.; Ablazov, A.; Mi, J.; Blilou, I.; Al-Babili, S. The Arabidopsis DWARF27 gene encodes an all-trans-/9-cis-beta-carotene isomerase and is induced by auxin, abscisic acid and phosphate deficiency. Plant Sci. 2018, 277, 33–42. [Google Scholar] [CrossRef] [Green Version]
- Aman, R.; Schieber, A.; Carle, R. Effects of heating and illumination on trans-cis isomerization and degradation of beta-carotene and lutein in isolated spinach chloroplasts. J. Agric. Food Chem. 2005, 53, 9512–9518. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.M.; Boileau, A.C.; Boileau, T.W.M.; Williams, A.W.; Swanson, K.S.; Heintz, K.A.; Erdman, J.W. Review of animal models in carotenoid research. J. Nutr. 1999, 129, 2271–2277. [Google Scholar] [CrossRef] [PubMed]
- Forman, B.M.; Umesono, K.; Chen, J.; Evans, R.M. Unique response pathways are established by allosteric interactions among nuclear hormone receptors. Cell 1995, 81, 541–550. [Google Scholar] [CrossRef] [Green Version]
- Von Lintig, J.; Vogt, K. Filling the gap in vitamin A research. Molecular identification of an enzyme cleaving beta-carotene to retinal. J. Biol. Chem. 2000, 275, 11915–11920. [Google Scholar] [CrossRef] [Green Version]
- Kiefer, C.; Hessel, S.; Lampert, J.M.; Vogt, K.; Lederer, M.O.; Breithaupt, D.E.; von Lintig, J. Identification and characterization of a mammalian enzyme catalyzing the asymmetric oxidative cleav-age of provitamin A. J. Biol. Chem. 2001, 276, 14110–14116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Lintig, J.; Hessel, S.; Isken, A.; Kiefer, C.; Lampert, J.M.; Voolstra, O.; Vogt, K. Towards a better understanding of carotenoid metabolism in animals. Biochim. Biophys. Acta 2005, 1740, 122–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maeda, T.; Perusek, L.; Amengual, J.; Babino, D.; Palczewski, K.; von Lintig, J. Dietary 9-cis-beta,beta-carotene fails to rescue vision in mouse models of leber congenital amaurosis. Mol. Pharmacol. 2011, 80, 943–952. [Google Scholar] [CrossRef] [Green Version]
- Blaner, W.S. Vitamin A signaling and homeostasis in obesity, diabetes, and metabolic disorders. Pharmacol. Ther. 2019, 197, 153–178. [Google Scholar] [CrossRef]
- Landrier, J.F.; Kasiri, E.; Karkeni, E.; Mihaly, J.; Beke, G.; Weiss, K.; Lucas, R.; Aydemir, G.; Salles, J.; Walrand, S.; et al. Reduced adiponectin expression after high-fat diet is associated with selective up-regulation of ALDH1A1 and further retinoic acid receptor signaling in adipose tissue. FASEB J. 2017, 31, 203–211. [Google Scholar] [CrossRef] [Green Version]
- Arnold, S.L.; Kent, T.; Hogarth, C.A.; Schlatt, S.; Prasad, B.; Haenisch, M.; Walsh, T.; Muller, C.H.; Griswold, M.D.; Amory, J.K.; et al. Importance of ALDH1A enzymes in determining human testicular retinoic acid concentrations. J. Lipid Res. 2015, 56, 342–357. [Google Scholar] [CrossRef] [Green Version]
- Napoli, J.L. Physiological insights into all-trans-retinoic acid biosynthesis. Biochim. Biophys. Acta BBA Mol. Cell Biol. Lipids 2012, 1821, 152–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krzyzosiak, A.; Szyszka-Niagolov, M.; Wietrzych, M.; Gobaille, S.; Muramatsu, S.; Krezel, W. Retinoid X receptor gamma control of affective behaviors involves dopaminergic signaling in mice. Neuron 2010, 66, 908–920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Neerven, S.; Kampmann, E.; Mey, J. RAR/RXR and PPAR/RXR signaling in neurological and psychiatric diseases. Prog. Neurobiol. 2008, 85, 433–451. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krężel, W.; Rivas, A.; Szklenar, M.; Ciancia, M.; Alvarez, R.; de Lera, A.R.; Rühl, R. Vitamin A5/X, a New Food to Lipid Hormone Concept for a Nutritional Ligand to Control RXR-Mediated Signaling. Nutrients 2021, 13, 925. https://doi.org/10.3390/nu13030925
Krężel W, Rivas A, Szklenar M, Ciancia M, Alvarez R, de Lera AR, Rühl R. Vitamin A5/X, a New Food to Lipid Hormone Concept for a Nutritional Ligand to Control RXR-Mediated Signaling. Nutrients. 2021; 13(3):925. https://doi.org/10.3390/nu13030925
Chicago/Turabian StyleKrężel, Wojciech, Aurea Rivas, Monika Szklenar, Marion Ciancia, Rosana Alvarez, Angel R. de Lera, and Ralph Rühl. 2021. "Vitamin A5/X, a New Food to Lipid Hormone Concept for a Nutritional Ligand to Control RXR-Mediated Signaling" Nutrients 13, no. 3: 925. https://doi.org/10.3390/nu13030925