
Circulating Human Serum Metabolites Derived from the Intake of a Saffron Extract (Safr’InsideTM) Protect Neurons from Oxidative Stress: Consideration for Depressive Disorders

 , , ,
, , ,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Clinical Trial
2.2. Human Study Design and Pharmacokinetics of Absorption
2.3. Saffron Extract (SaE)
2.4. Human Neuron Cultures
2.5. Cell Viability
2.6. Reactive Oxygen Species (ROS) Measurement
2.7. Brain-Derived Neurotrophic (BDNF) Factor, Serotonin, 5-Hydroxyindoleacetic Acid (5-HIAA), and Dopamine Quantification
2.8. Real-Time RT-qPCR
2.9. Statistics
3. Results
3.1. Kinetic Profile of the SaE Absorption
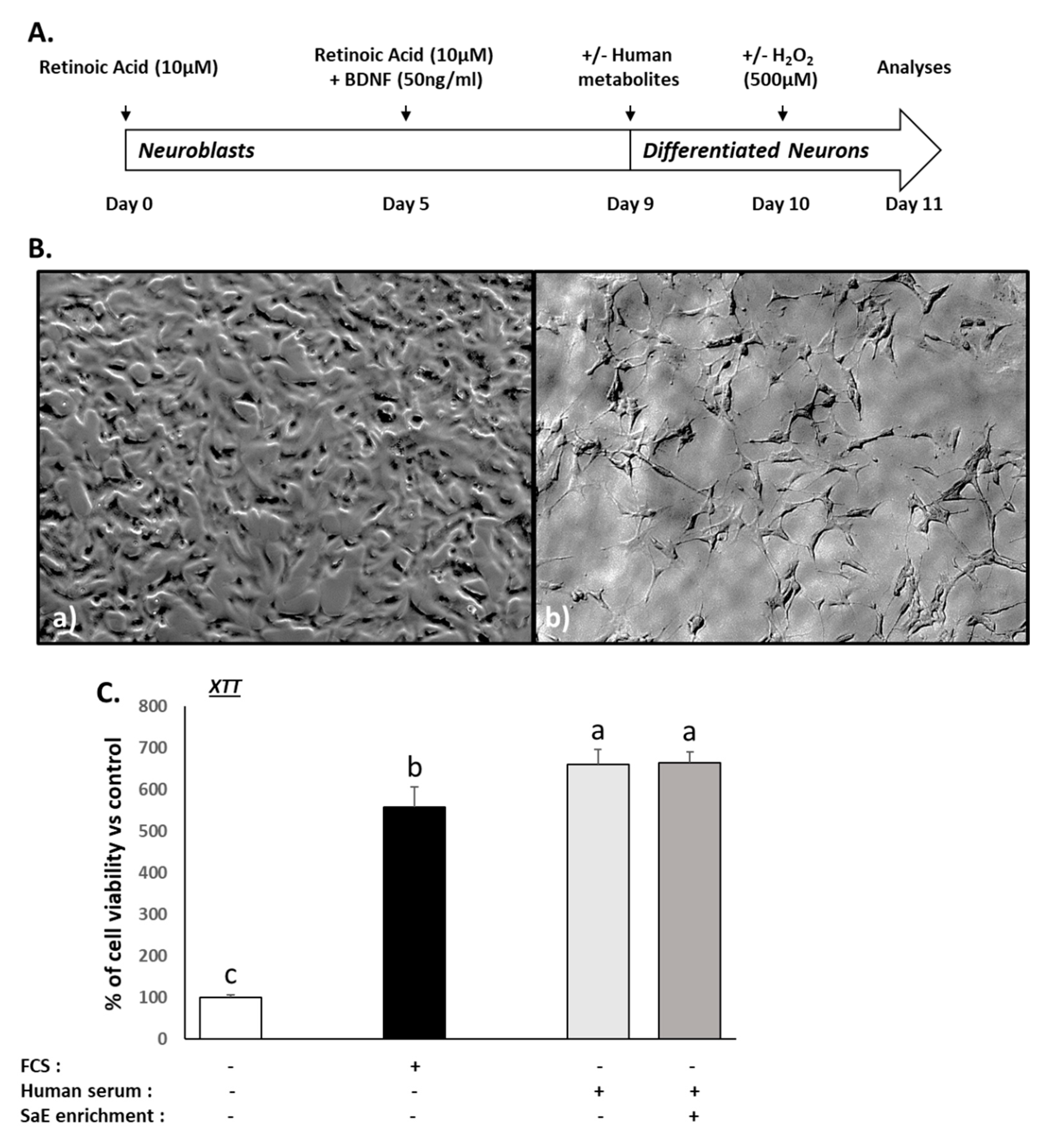
3.2. Validation of the Neuronal Differentiation and Ex Vivo Procedures
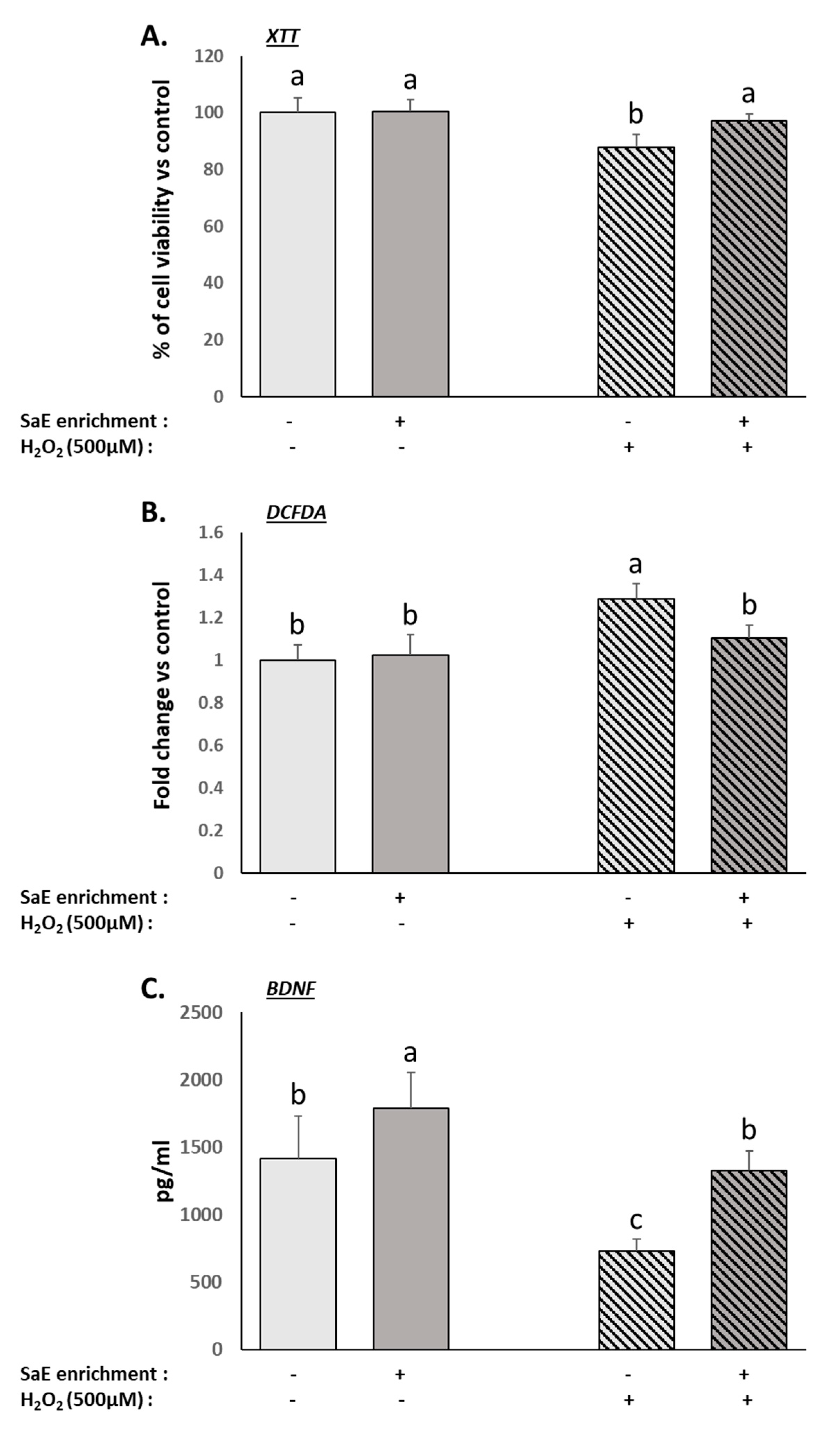
3.3. Human Serum Containing Metabolites from SaE Preserves Human Neurons from Oxidative-Stress-Induced Neurotoxicity
3.4. Human Serum Containing Metabolites from SaE Enhance Basal Activity of Serotonergic System and Protect It from Oxidative-Stress-Induced Alterations
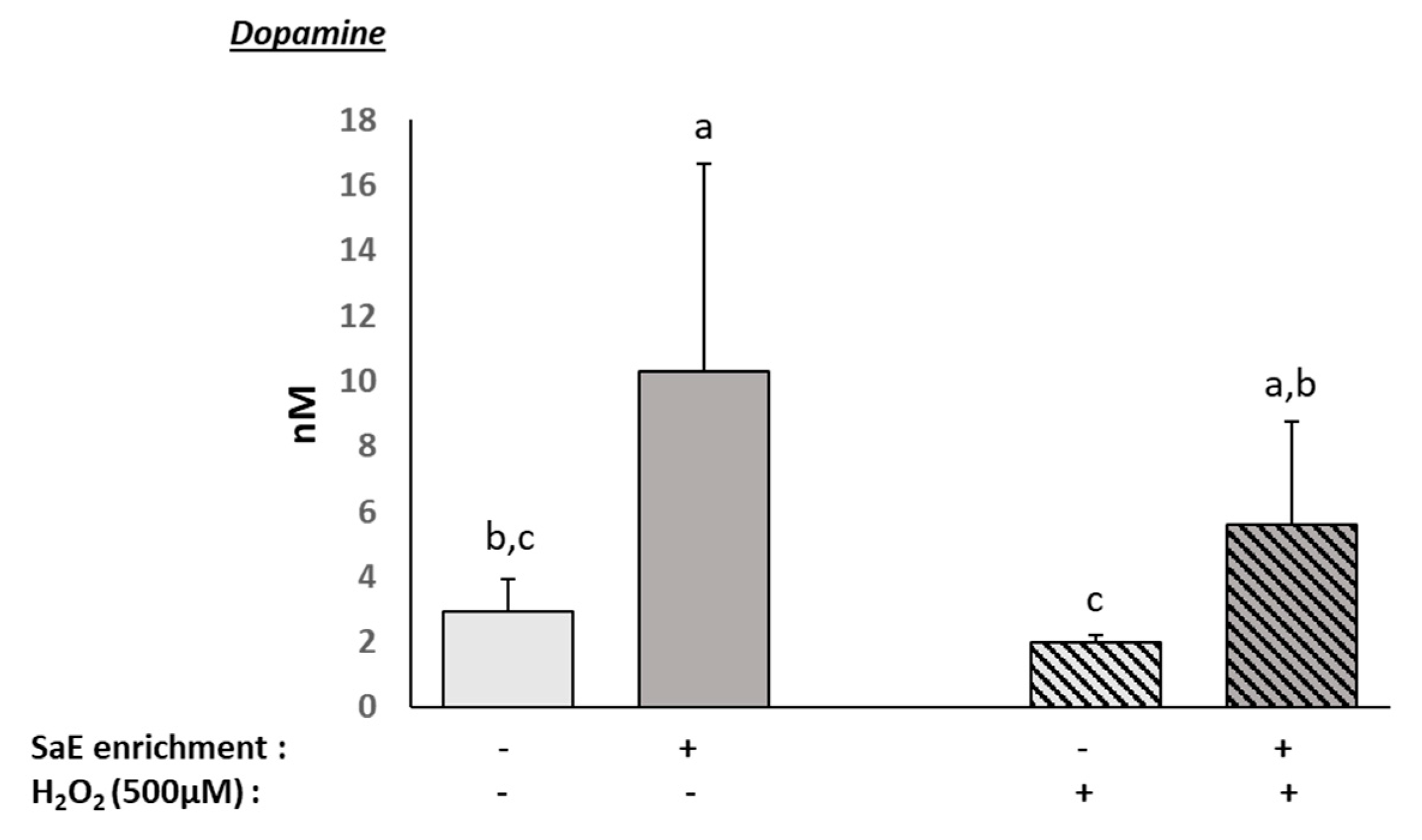
3.5. Human Serum Containing Metabolites from SaE Stimulates Dopamine Production
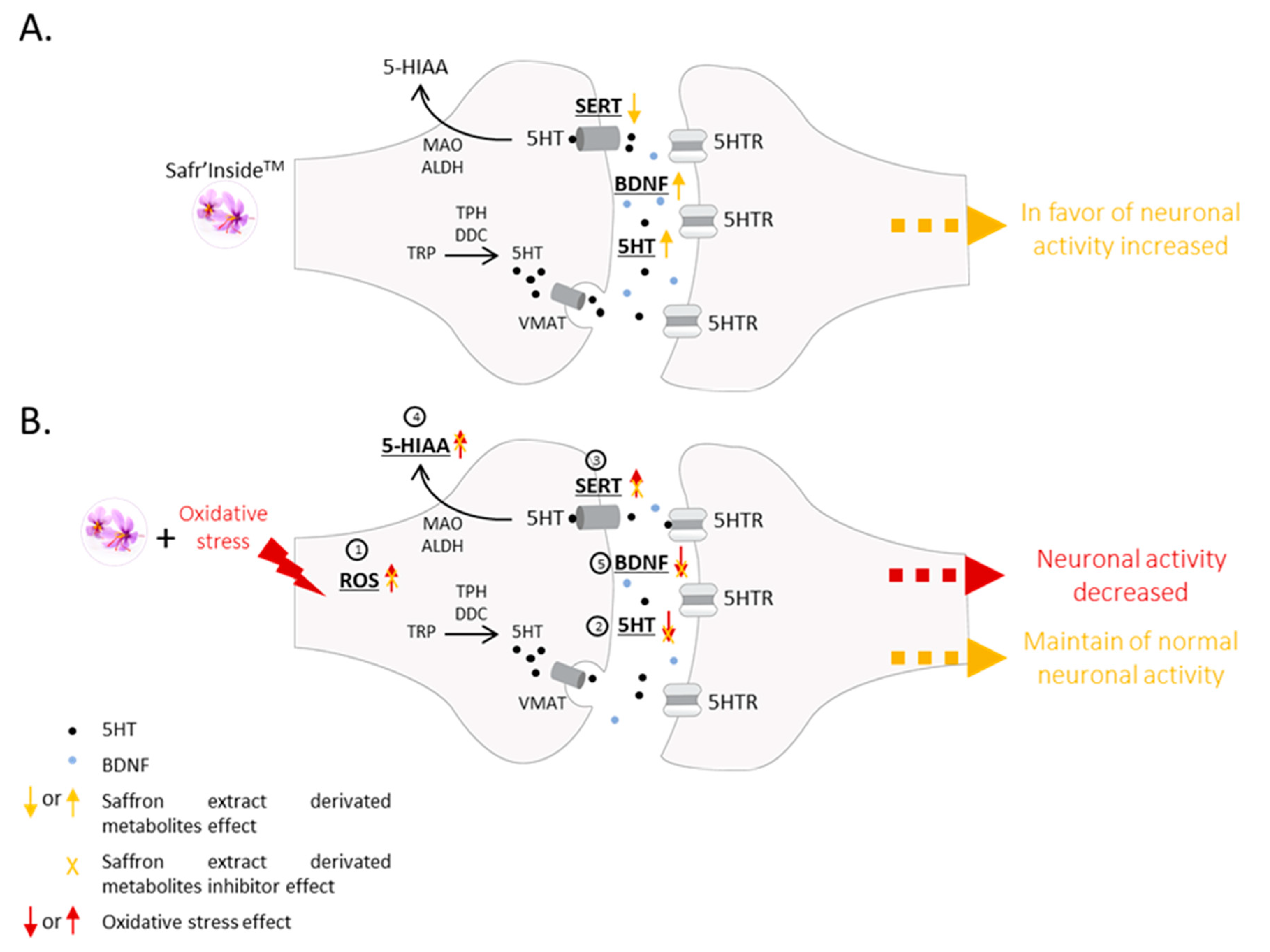
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Businaro, R.; Vauzour, D.; Sarris, J.; Münch, G.; Gyengesi, E.; Brogelli, L.; Zuzarte, P. Therapeutic Opportunities for Food Supplements in Neurodegenerative Disease and Depression. Front. Nutr. 2021, 8, 669846. [Google Scholar] [CrossRef] [PubMed]
- Michel, T.M.; Pulschen, D.; Thome, J. The role of oxidative stress in depressive disorders. Curr. Pharm. Des. 2012, 18, 5890–5899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hovatta, I.; Juhila, J.; Donner, J. Oxidative stress in anxiety and comorbid disorders. Neurosci. Res. 2010, 68, 261–275. [Google Scholar] [CrossRef] [PubMed]
- Maes, M.; Kubera, M.; Obuchowiczwa, E.; Goehler, L.; Brzeszcz, J. Depression’s multiple comorbidities explained by (neuro)inflammatory and oxidative & nitrosative stress pathways. Neuroendocr. Lett. 2011, 32, 7–24. [Google Scholar]
- Lindqvist, D.; Dhabhar, F.S.; James, S.J.; Hough, C.M.; Jain, F.A.; Bersani, F.S.; Reus, V.I.; Verhoeven, J.E.; Epel, E.S.; Mahan, L.; et al. Oxidative stress, inflammation and treatment response in major depression. Psychoneuroendocrinology 2017, 76, 197–205. [Google Scholar] [CrossRef] [Green Version]
- Black, C.N.; Bot, M.; Scheffer, P.G.; Cuijpers, P.; Penninx, B.W. Is depression associated with increased oxidative stress? A systematic review and meta-analysis. Psychoneuroendocrinology 2015, 51, 164–175. [Google Scholar] [CrossRef] [Green Version]
- Masood, A.; Nadeem, A.; Mustafa, S.J.; O’Donnell, J. Reversal of oxidative stress-induced anxiety by inhibition of phosphodiesterase-2 in mice. J. Pharmacol. Exp. Ther. 2008, 326, 369–379. [Google Scholar] [CrossRef] [Green Version]
- Salim, S.; Asghar, M.; Chugh, G.; Taneja, M.; Xia, Z.; Saha, K. Oxidative stress: A potential recipe for anxiety, hypertension and insulin resistance. Brain Res. 2010, 1359, 178–185. [Google Scholar] [CrossRef] [Green Version]
- Bouvier, E.; Brouillard, F.; Molet, J.; Claverie, D.; Cabungcal, J.H.; Cresto, N.; Doligez, N.; Rivat, C.; Do, K.Q.; Bernard, C.; et al. Nrf2-dependent persistent oxidative stress results in stress-induced vulnerability to depression. Mol. Psychiatry 2017, 22, 1701–1713. [Google Scholar] [CrossRef] [Green Version]
- Sen, S.; Duman, R.; Sanacora, G. Serum brain-derived neurotrophic factor, depression, and antidepressant medications: Meta-analyses and implications. Biol. Psychiatry 2008, 64, 527–532. [Google Scholar] [CrossRef] [Green Version]
- Jimenez-Fernandez, S.; Jimenez-Fernandez, S.; Gurpegui, M.; Diaz-Atienza, F.; Pérez-Costillas, L.; Gerstenberg, M.; Correll, C.U. Oxidative stress and antioxidant parameters in patients with major depressive disorder compared to healthy controls before and after antidepressant treatment: Results from a meta-analysis. J. Clin. Psychiatry 2015, 76, 1658–1667. [Google Scholar] [CrossRef] [PubMed]
- Bagur, M.J.; Salinas, G.L.A.; Jiménez-Monreal, A.M.; Chaouqi, S.; Llorens, S.; Martínez-Tomé, M.; Alonso, G.L. Saffron: An Old Medicinal Plant and a Potential Novel Functional Food. Molecules 2017, 23, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghasemi, T.; Abnous, K.; Vahdati, F.; Mehri, S.; Razavi, B.M.; Hosseinzadeh, H. Antidepressant Effect of Crocus sativus Aqueous Extract and its Effect on CREB, BDNF, and VGF Transcript and Protein Levels in Rat Hippocampus. Drug Res. 2015, 65, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Khaksarian, M.; Behzadifar, M.; Behzadifar, M.; Alipour, M.; Jahanpanah, F.; Re, T.S.; Firenzuoli, F.; Zerbetto, R.; Bragazzi, N.L. The efficacy of Crocus sativus (Saffron) versus placebo and Fluoxetine in treating depression: A systematic review and meta-analysis. Psychol. Res. Behav. Manag. 2019, 12, 297–305. [Google Scholar] [CrossRef] [Green Version]
- Kashani, L.; Esalatmanesh, S.; Eftekhari, F.; Salimi, S.; Foroughifar, T.; Etesam, F.; Safiaghdam, H.; Moazen-Zadeh, E.; Akhondzadeh, S. Efficacy of Crocus sativus (saffron) in treatment of major depressive disorder associated with post-menopausal hot flashes: A double-blind, randomized, placebo-controlled trial. Arch. Gynecol. Obstet. 2018, 297, 717–724. [Google Scholar] [CrossRef]
- Ahmadpanah, M.; Ramezanshams, F.; Ghaleiha, A.; Akhondzadeh, S.; Bahmani, D.S.; Brand, S. Crocus Sativus L. (saffron) versus sertraline on symptoms of depression among older people with major depressive disorders-a double-blind, randomized intervention study. Psychiatry Res. 2019, 282, 112613. [Google Scholar] [CrossRef]
- Jackson, P.A.; Forster, J.; Khan, J.; Pouchieu, C.; Dubreuil, S.; Gaudout, D.; Moras, B.; Pourtau, L.; Joffre, F.; Vaysse, C.; et al. Effects of Saffron Extract Supplementation on Mood, Well-Being, and Response to a Psychosocial Stressor in Healthy Adults: A Randomized, Double-Blind, Parallel Group, Clinical Trial. Front. Nutr. 2020, 7, 606124. [Google Scholar] [CrossRef]
- Heidari, S.; Mehri, S.; Shariaty, V.; Hosseinzadeh, A.H. Preventive effects of crocin on neuronal damages induced by D-galactose through AGEs and oxidative stress in human neuroblastoma cells (SH-SY5Y). J. Pharmacopunct. 2018, 21, 18–25. [Google Scholar] [CrossRef]
- Pan, P.K.; Qiao, L.Y.; Wen, X.N. Safranal prevents rotenone-induced oxidative stress and apoptosis in an in vitro model of Parkinson’s disease through regulating Keap1/Nrf2 signaling pathway. Cell. Mol. Biol. 2016, 62, 11–17. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Sun, Y.; Cheng, Z.; Guo, Y.; Liu, P.; Wen, Y. Crocin exerts anti-inflammatory and anti-arthritic effects on type II collagen-induced arthritis in rats. Pharm. Biol. 2018, 56, 209–216. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, M.; Betti, G.; Hensel, A. Saffron in phytotherapy: Pharmacology and clinical uses. Wien. Med. Wochenschr. 2007, 157, 315–319. [Google Scholar] [CrossRef] [PubMed]
- Surguchov, A.; Bernal, L.; Surguchev, A.A. Phytochemicals as Regulators of Genes Involved in Synucleinopathies. Biomolecules 2021, 11, 624. [Google Scholar] [CrossRef] [PubMed]
- Monchaux De Oliveira, C.; Pourtau, L.; Vancassel, S.; Pouchieu, C.; Capuron, L.; Gaudout, D.; Castanon, N. Saffron Extract-Induced Improvement of Depressive-Like Behavior in Mice Is Associated with Modulation of Monoaminergic Neurotransmission. Nutrients 2021, 13, 904. [Google Scholar] [CrossRef]
- Kleinnijenhuis, A.J.; Van Holthoon, F.L.; Maathuis, A.J.; Vanhoecke, B.; Prawitt, J.; Wauquier, F.; Wittrant, Y. Non-targeted and targeted analysis of collagen hydrolysates during the course of digestion and absorption. Anal. Bioanal. Chem. 2020, 412, 973–982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wauquier, F.; Boutin-Wittrant, L.; Viret, A.; Guilhaudis, L.; Oulyadi, H.; Bourafai-Aziez, A.; Charpentier, G.; Rousselot, G.; Cassin, E.; Descamps, S.; et al. Metabolic and Anti-Inflammatory Protective Properties of Human Enriched Serum Following Artichoke Leaf Extract Absorption: Results from an Innovative Ex Vivo Clinical Trial. Nutrients 2021, 13, 2653. [Google Scholar] [CrossRef]
- Wauquier, F.; Daneault, A.; Granel, H.; Prawitt, J.; Soulé, V.F.; Berger, J.; Pereira, B.; Guicheux, J.; Rochefort, G.Y.; Meunier, N.; et al. Human Enriched Serum Following Hydrolysed Collagen Absorption Modulates Bone Cell Activity: From Bedside to Bench and Vice Versa. Nutrients 2019, 11, 1249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wauquier, F.; Mevel, E.; Krisa, S.; Richard, T.; Valls, J.; Hornedo-Ortega, R.; Granel, H.; Boutin-Wittrant, L.; Urban, N.; Berger, J.; et al. Chondroprotective Properties of Human-Enriched Serum Following Polyphenol Extract Absorption: Results from an Exploratory Clinical Trial. Nutrients 2019, 11, 3071. [Google Scholar]
- Babaei, A.; Arshami, J.; Haghparast, A.; Daneshmesgharan, M. Effects of saffron (Crocus sativus) petal ethanolic extract on hematology, antibody response, and spleen histology in rats. Avicenna J. Phytomed. 2014, 4, 103–109. [Google Scholar]
- Li, J.; Lei, H.T.; Cao, L.; Mi, Y.N.; Li, S.; Cao, Y.X. Crocin alleviates coronary atherosclerosis via inhibiting lipid synthesis and inducing M2 macrophage polarization. Int. Immunopharmacol. 2018, 55, 120–127. [Google Scholar] [CrossRef]
- Nair, S.C.; Kurumboor, S.K.; Hasegawa, J.H. Saffron chemoprevention in biology and medicine: A review. Cancer Biother. Radiopharm. 1995, 10, 257–264. [Google Scholar] [CrossRef]
- Almodovar, P.; Briskey, D.; Rao, A.; Prodanov, M.; Inarejos-García, A.M. Bioaccessibility and Pharmacokinetics of a Commercial Saffron (Crocus sativus L.) Extract. Evid Based Complement Altern. Med. 2020, 2020, 1575730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayatollahi, H.; Javan, A.O.; Khajedaluee, M.; Shahroodian, M.; Hosseinzadeh, H. Effect of Crocus sativus L. (saffron) on coagulation and anticoagulation systems in healthy volunteers. Phytother. Res. 2014, 28, 539–543. [Google Scholar] [CrossRef] [PubMed]
- Modaghegh, M.H.; Shahabian, M.; Esmaeili, H.-A.; Rajbai, O.; Hosseinzadeh, H. Safety evaluation of saffron (Crocus sativus) tablets in healthy volunteers. Phytomedicine 2008, 15, 1032–1037. [Google Scholar] [CrossRef] [PubMed]
- Umigai, N.; Murakami, K.; Ulit, M.; Antonio, L.; Shirotori, M.; Morikawa, H.; Nakano, T. The pharmacokinetic profile of crocetin in healthy adult human volunteers after a single oral administration. Phytomedicine 2011, 18, 575–578. [Google Scholar] [CrossRef]
- Frank, A. Unusual purpura in artificial abortion. Dtsch. Med. Wochenschr. 1961, 86, 1618–1620. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Fei, F.; Zhen, L.; Zhu, X.; Wang, J.; Li, S.; Geng, J.; Sun, R.; Yu, X.; Chen, T.; et al. Sensitive analysis and simultaneous assessment of pharmacokinetic properties of crocin and crocetin after oral administration in rats. J. Chromatogr. B 2017, 1044–1045, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, N.R.; Kumar, R.; Gupta, A.; Meena, R.C.; Nanda, S.; Mishra, K.P.; Singh, S.B. Heat stress induced oxidative damage and perturbation in BDNF/ERK1/2/CREB axis in hippocampus impairs spatial memory. Behav. Brain Res. 2021, 396, 112895. [Google Scholar] [CrossRef]
- Ahmadimanesh, M.; Abbaszadegan, M.R.; Rad, D.M.; Moallem, S.M.H.; Mohammadpour, A.H.; Ghahremani, M.H.; Hosseini, F.F.; Behdani, F.; Manteghi, A.A.; Jowsey, P.; et al. Effects of selective serotonin reuptake inhibitors on DNA damage in patients with depression. J. Psychopharmacol. 2019, 33, 1364–1376. [Google Scholar] [CrossRef] [PubMed]
- Khanzode, S.D.; Dakhale, G.N.; Khanzode, S.S.; Saoji, A.; Palasodkar, R. Oxidative damage and major depression: The potential antioxidant action of selective serotonin re-uptake inhibitors. Redox Rep. 2003, 8, 365–370. [Google Scholar] [CrossRef]
- Moghadam, B.H.; Bagheri, R.; Roozbeh, B.; Ashtary-Larky, D.; Gaeini, A.A.; Dutheil, F.; Wong, A. Impact of saffron (Crocus Sativus Linn) supplementation and resistance training on markers implicated in depression and happiness levels in untrained young males. Physiol. Behav. 2021, 233, 113352. [Google Scholar] [CrossRef]
- Hosseini, A.; Razavi, B.M.; Hosseinzadeh, H. Pharmacokinetic Properties of Saffron and its Active Components. Eur. J. Drug Metab. Pharm. 2018, 43, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Wani, A.; Al Rihani, S.B.; Sharma, A.; Weadick, B.; Govindarajan, R.; Khan, S.U.; Sharma, P.R.; Dogra, A.; Nandi, U.; Reddy, C.N.; et al. Crocetin promotes clearance of amyloid-beta by inducing autophagy via the STK11/LKB1-mediated AMPK pathway. Autophagy 2021, 17, 3813–3832. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.H.; Xie, Y.; Huang, X.; Kadota, K.; Yao, X.S.; Yu, Y.; Chen, X.; Lu, A.; Yang, Z. Delivering Crocetin across the Blood-Brain Barrier by Using gamma-Cyclodextrin to Treat Alzheimer’s Disease. Sci. Rep. 2020, 10, 3654. [Google Scholar] [CrossRef] [PubMed]
- Tarafdar, A.; Pula, G. The Role of NADPH Oxidases and Oxidative Stress in Neurodegenerative Disorders. Int. J. Mol. Sci. 2018, 19, 3824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salim, S.; Sarraj, N.; Taneja, M.; Saha, K.; Tejada-Simon, M.V.; Chugh, G. Moderate treadmill exercise prevents oxidative stress-induced anxiety-like behavior in rats. Behav. Brain Res. 2010, 208, 545–552. [Google Scholar] [CrossRef]
- Solanki, N.; Salvi, A.; Patki, G.; Salim, S. Modulating Oxidative Stress Relieves Stress-Induced Behavioral and Cognitive Impairments in Rats. Int. J. Neuropsychopharmacol. 2017, 20, 550–561. [Google Scholar] [CrossRef] [PubMed]
- Camargo, A.; Dalmagro, A.P.; Rikel, L.; da Silva, E.B.; da Silva, K.A.B.S.; Zeni, A.L.B. Cholecalciferol counteracts depressive-like behavior and oxidative stress induced by repeated corticosterone treatment in mice. Eur. J. Pharmacol. 2018, 833, 451–461. [Google Scholar] [CrossRef]
- Dalmagro, A.P.; Camargo, A.; Pedron, N.B.; Garcia, S.A.; Zeni, A.L.B. Morus nigra leaves extract revokes the depressive-like behavior, oxidative stress, and hippocampal damage induced by corticosterone: A pivotal role of the phenolic syringic acid. Behav. Pharmacol. 2020, 31, 397–406. [Google Scholar] [CrossRef]
- Sharif, N.F.; Korade, Z.; Porter, N.A.; Harrison, F.E. Oxidative stress, serotonergic changes and decreased ultrasonic vocalizations in a mouse model of Smith-Lemli-Opitz syndrome. Genes Brain Behav. 2017, 16, 619–626. [Google Scholar] [CrossRef]
- Wang, Y.; Gu, Y.H.; Liu, M.; Bai, Y.; Wang, H.L. Fluoxetine protects against methamphetamineinduced lung inflammation by suppressing oxidative stress through the SERT/p38 MAPK/Nrf2 pathway in rats. Mol. Med. Rep. 2017, 15, 673–680. [Google Scholar] [CrossRef]
- Hong, I.S.; Lee, H.Y.; Kim, H.P. Anti-oxidative effects of Rooibos tea (Aspalathus linearis) on immobilization-induced oxidative stress in rat brain. PLoS ONE 2014, 9, e87061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, J.M.; Lee, B.D.; Sok, D.E.; Ma, J.Y.; Kim, M.R. Neuroprotective action of N-acetyl serotonin in oxidative stress-induced apoptosis through the activation of both TrkB/CREB/BDNF pathway and Akt/Nrf2/Antioxidant enzyme in neuronal cells. Redox Biol. 2017, 11, 592–599. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wauquier, F.; Boutin-Wittrant, L.; Pourtau, L.; Gaudout, D.; Moras, B.; Vignault, A.; Monchaux De Oliveira, C.; Gabaston, J.; Vaysse, C.; Bertrand, K.; et al. Circulating Human Serum Metabolites Derived from the Intake of a Saffron Extract (Safr’InsideTM) Protect Neurons from Oxidative Stress: Consideration for Depressive Disorders. Nutrients 2022, 14, 1511. https://doi.org/10.3390/nu14071511
Wauquier F, Boutin-Wittrant L, Pourtau L, Gaudout D, Moras B, Vignault A, Monchaux De Oliveira C, Gabaston J, Vaysse C, Bertrand K, et al. Circulating Human Serum Metabolites Derived from the Intake of a Saffron Extract (Safr’InsideTM) Protect Neurons from Oxidative Stress: Consideration for Depressive Disorders. Nutrients. 2022; 14(7):1511. https://doi.org/10.3390/nu14071511
Chicago/Turabian StyleWauquier, Fabien, Line Boutin-Wittrant, Line Pourtau, David Gaudout, Benjamin Moras, Adeline Vignault, Camille Monchaux De Oliveira, Julien Gabaston, Carole Vaysse, Karène Bertrand, and et al. 2022. "Circulating Human Serum Metabolites Derived from the Intake of a Saffron Extract (Safr’InsideTM) Protect Neurons from Oxidative Stress: Consideration for Depressive Disorders" Nutrients 14, no. 7: 1511. https://doi.org/10.3390/nu14071511