
Increased Indoleamine 2,3-Dioxygenase 1 (IDO-1) Activity and Inflammatory Responses during Chikungunya Virus Infection

 ,
,  , , , , ,
, , , , ,  and
and
Abstract
:1. Introduction
2. Results
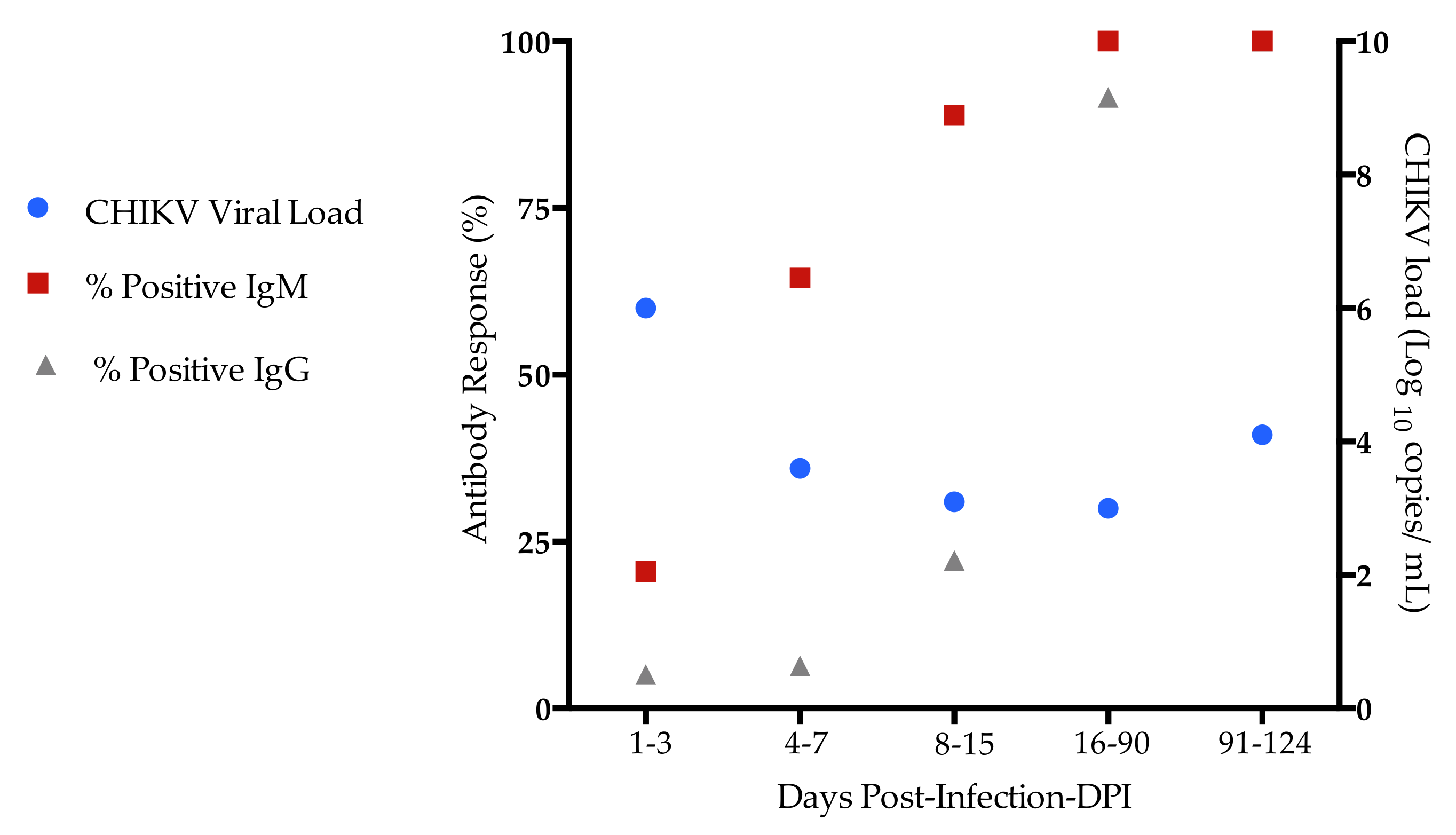
2.1. Demographic, Clinical, and Laboratorial Characteristics of Chikungunya-Infected Patients
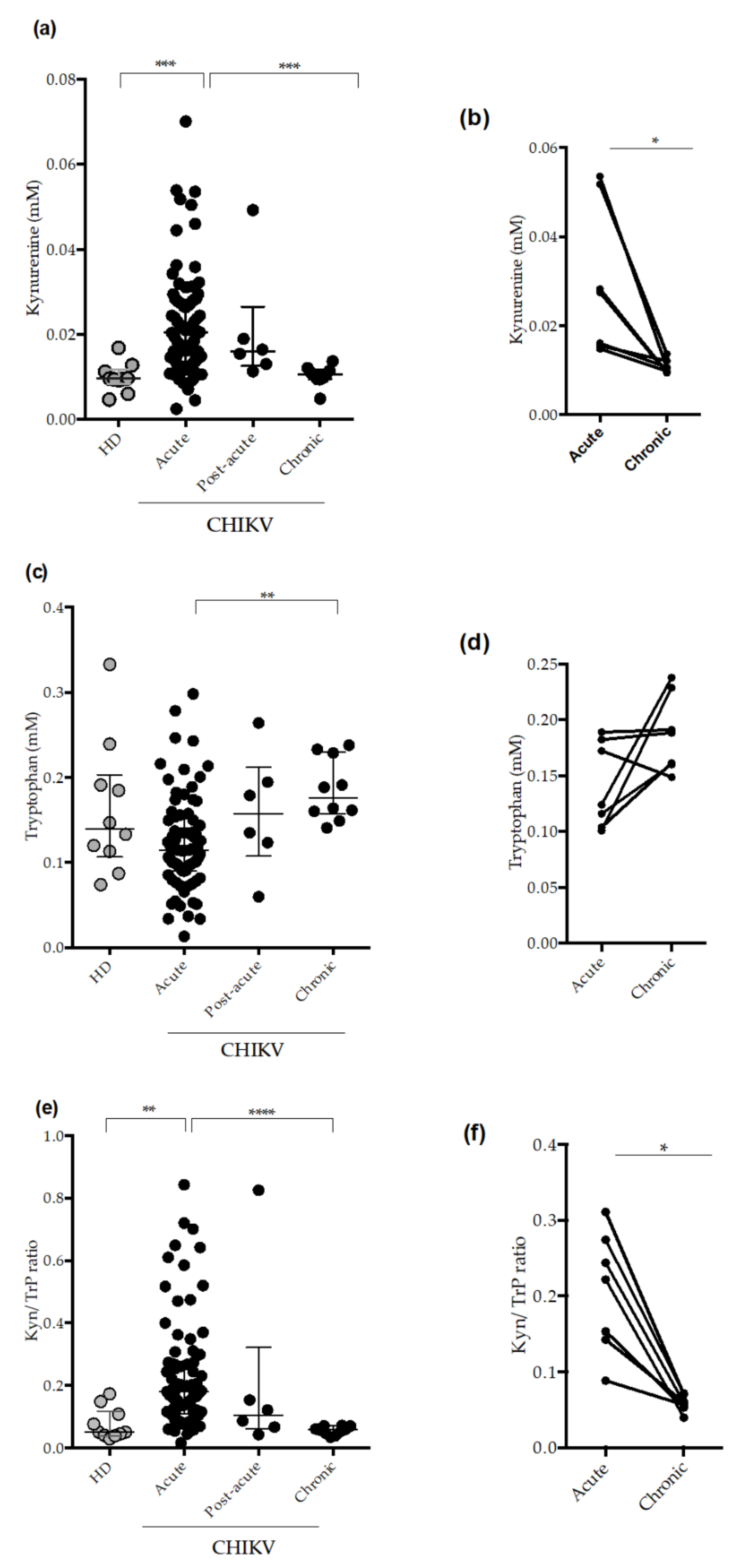
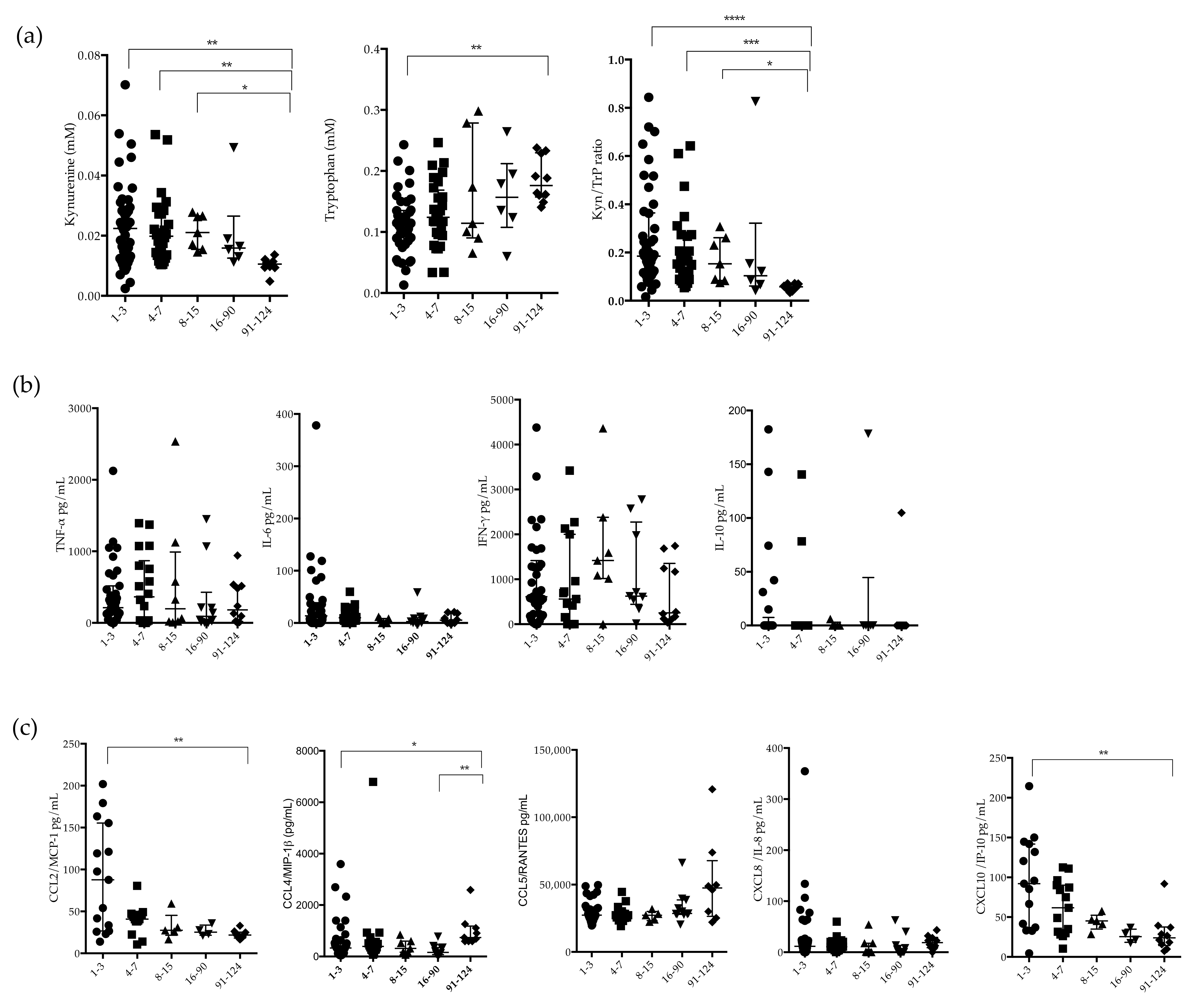
2.2. Circulating Plasma Levels of Kynurenune, Tryptophan and Indoleamine 2,3 Dioxygenase (IDO-1) Activity in CHIKV-Infected Patients
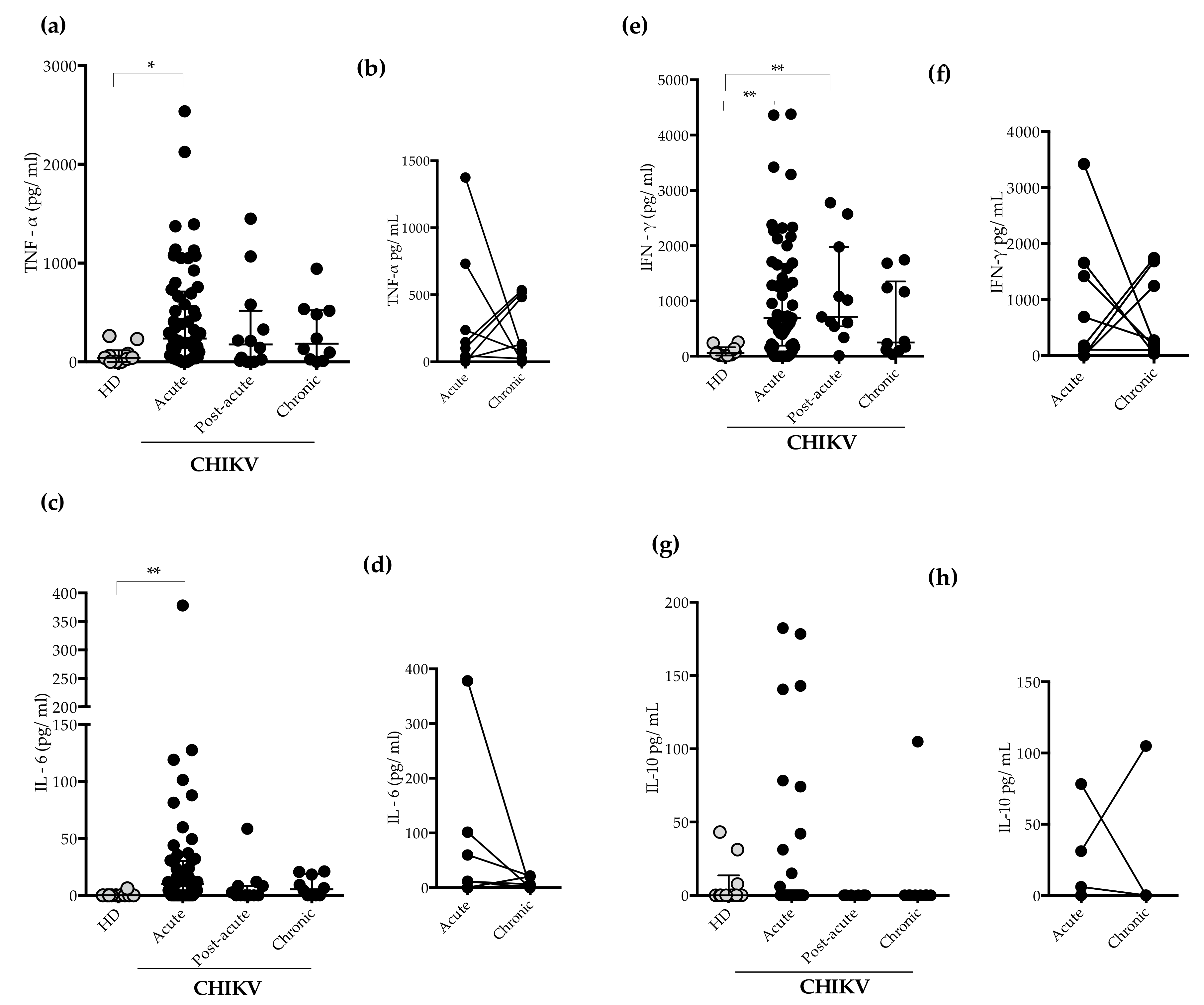
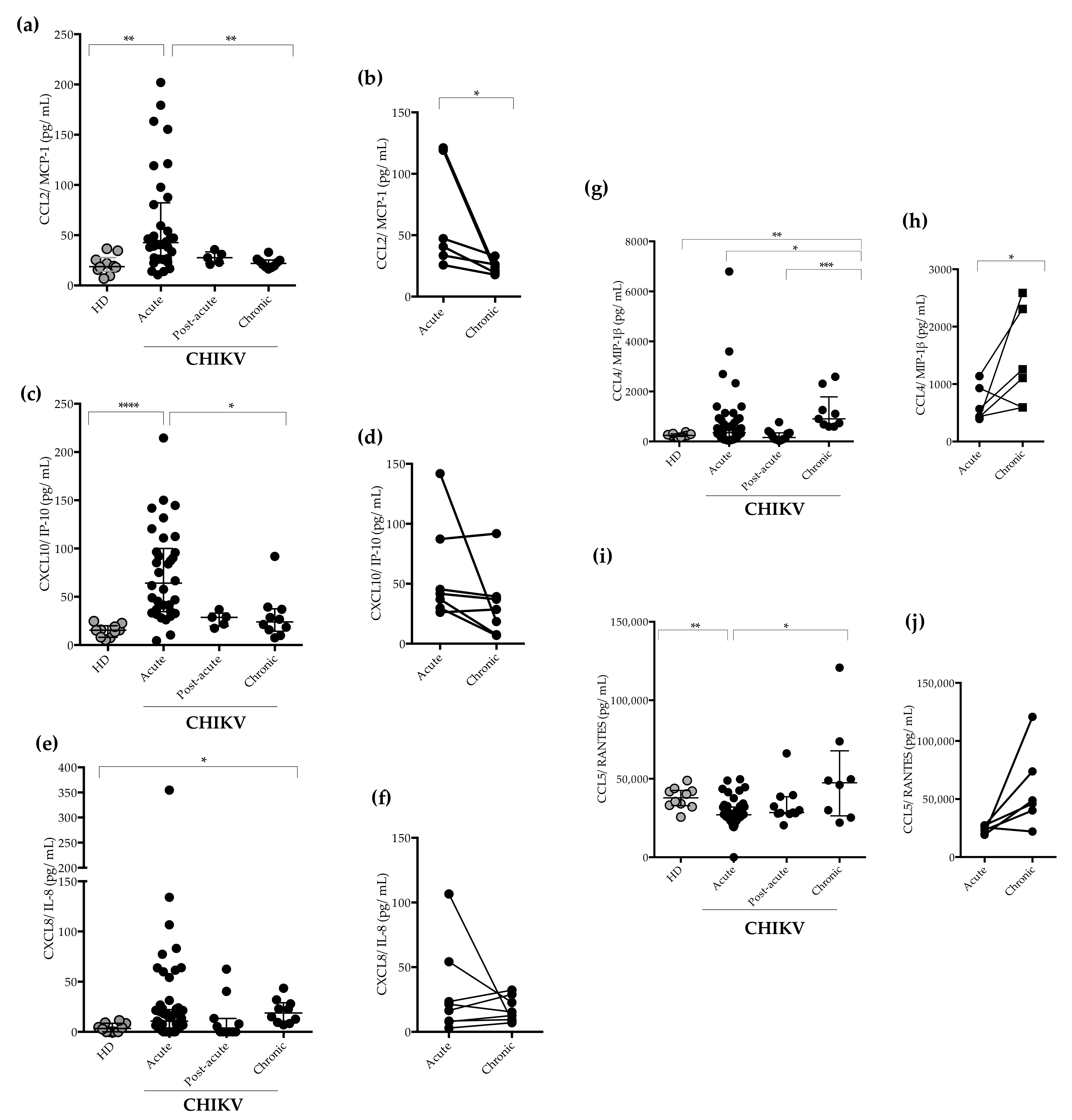
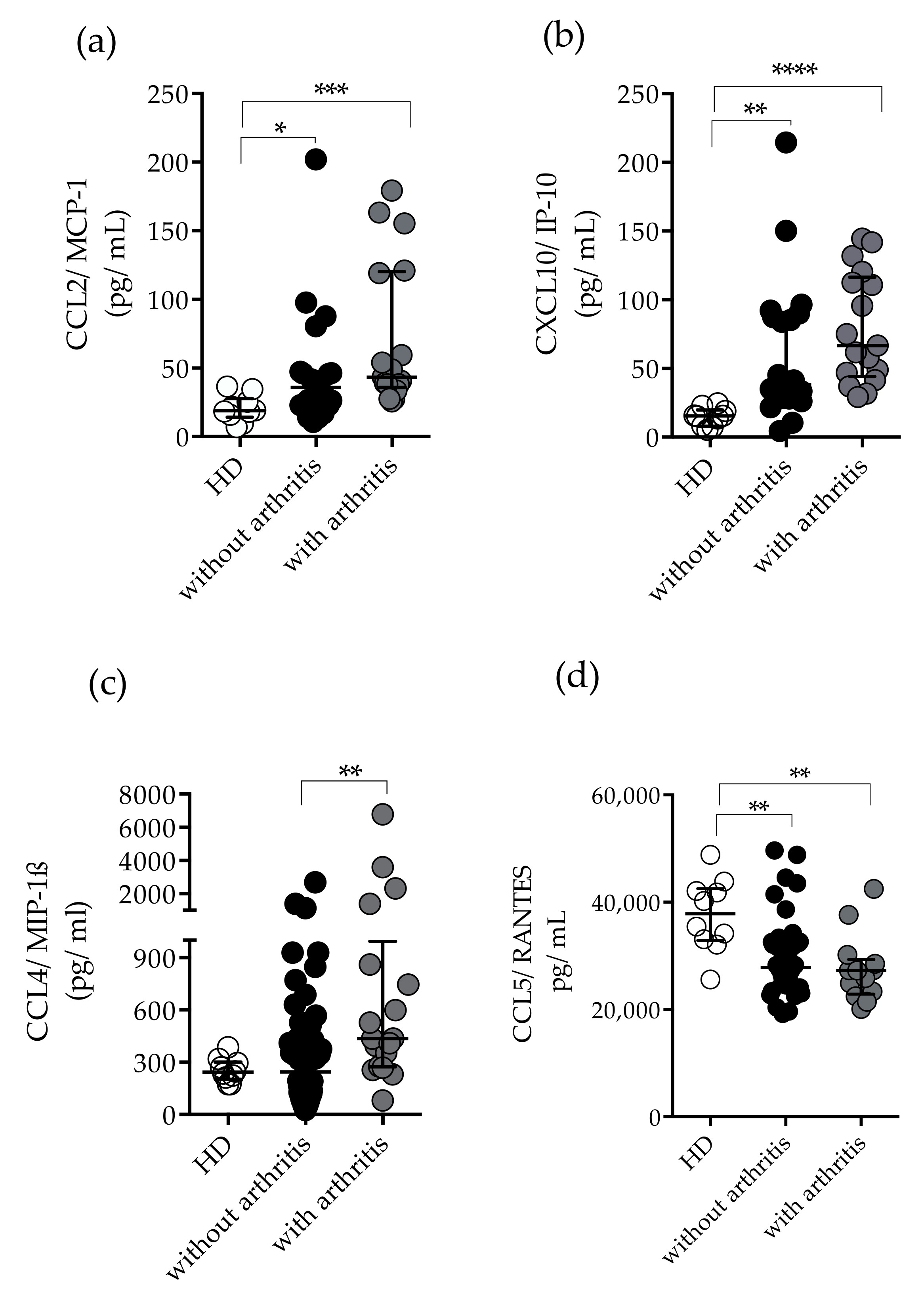
2.3. Measurement of Plasma Levels of Cytokines and Chemokines
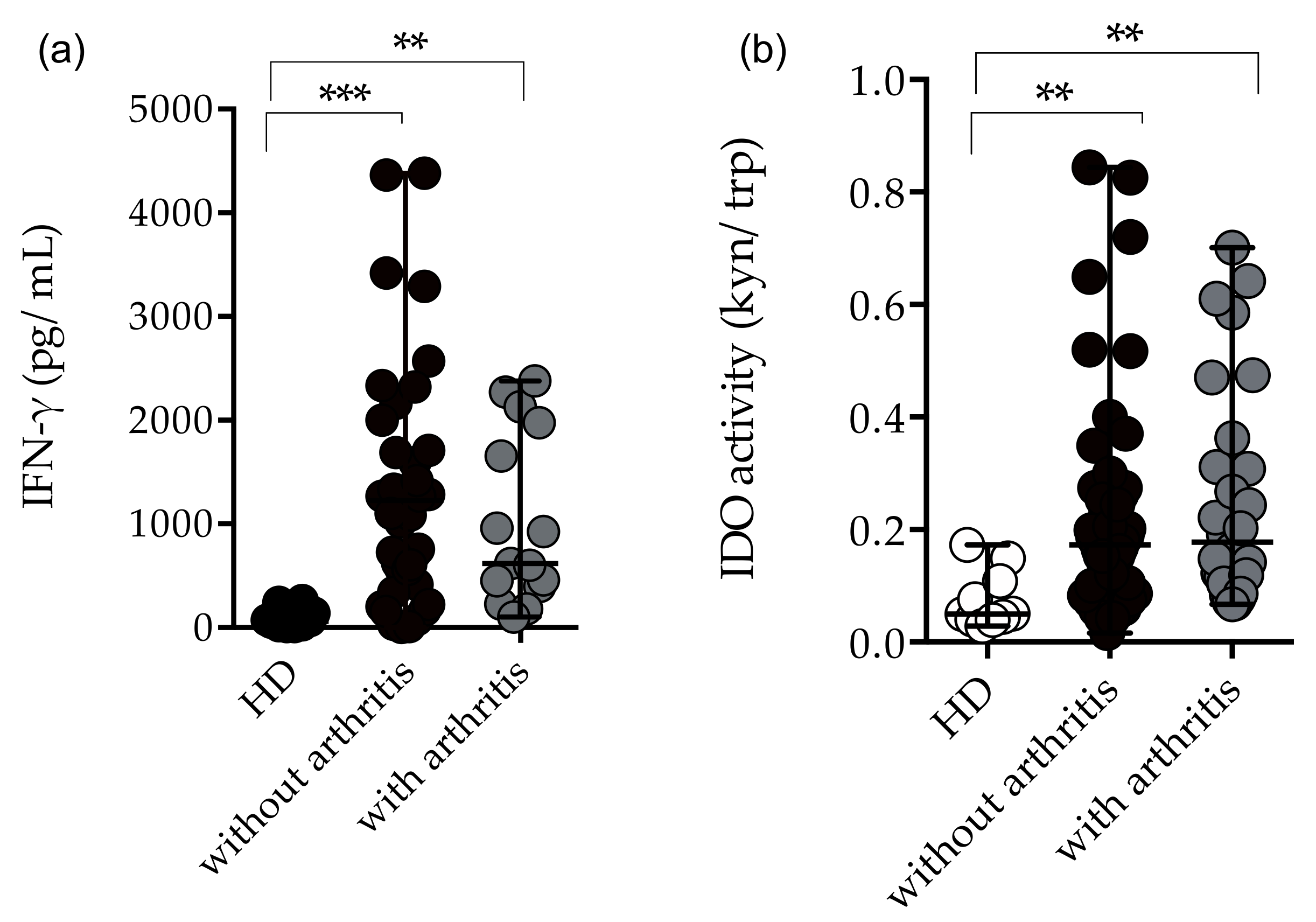
2.4. Immunological Profile in CHIKV-Infected Patients with Arthritis and Without Arthritis
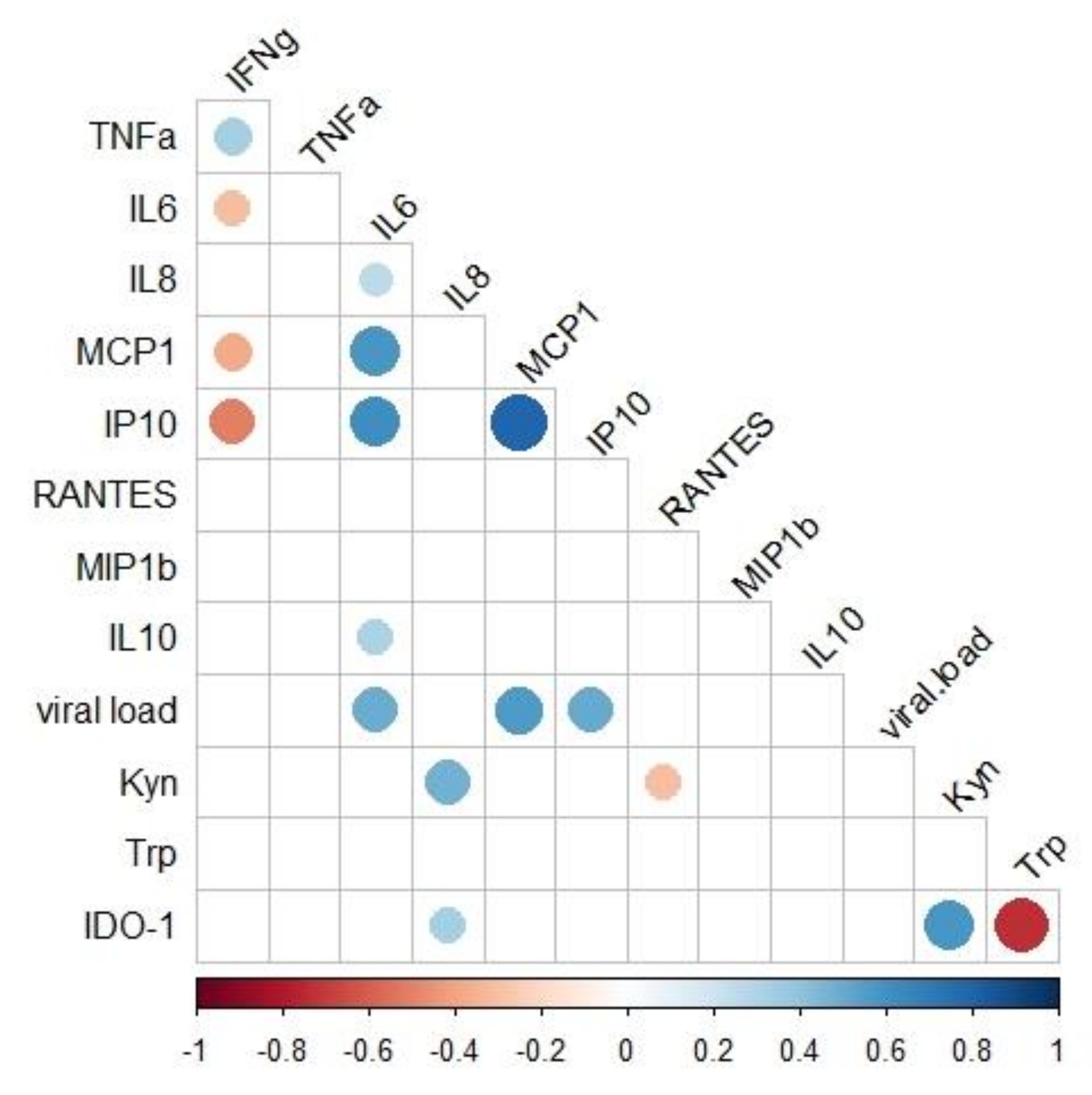
2.5. Associations of Immunological and Virologic Factors
3. Discussion
4. Materials and Methods
4.1. Study Subjects
4.2. Ethics Statement
4.3. Laboratorial Diagnosis
4.4. Measurement of Plasma Cytokines and Chemokines
4.5. Measurement of the IDO-1 Enzymatic Activity
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen recognition and innate immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medzhitov, R. The spectrum of inflammatory responses. Science 2021, 374, 1070–1075. [Google Scholar] [CrossRef] [PubMed]
- Munn, D.H.; Mellor, A.L. Indoleamine 2,3 dioxygenase and metabolic control of immune responses. Trends Immunol. 2013, 34, 137–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mellor, A.L.; Munn, D.H. Tryptophan catabolism and T-cell tolerance: Immunosuppression by starvation? Immunol. Today 1999, 20, 469–473. [Google Scholar] [CrossRef]
- Fallarino, F.; Grohmann, U.; You, S.; McGrath, B.C.; Cavener, D.R.; Vacca, C.; Orabona, C.; Bianchi, R.; Belladonna, M.L.; Volpi, C.; et al. The combined effects of tryptophan starvation and tryptophan catabolites down-regulate T cell receptor zeta-chain and induce a regulatory phenotype in naive T cells. J. Immunol. 2006, 176, 6752–6761. [Google Scholar] [CrossRef]
- Nunes, M.R.; Faria, N.R.; de Vasconcelos, J.M.; Golding, N.; Kraemer, M.U.; de Oliveira, L.F.; Azevedo Rdo, S.; da Silva, D.E.; da Silva, E.V.; da Silva, S.P.; et al. Emergence and potential for spread of Chikungunya virus in Brazil. BMC Med. 2015, 13, 102. [Google Scholar] [CrossRef] [Green Version]
- Azeredo, E.L.; Dos Santos, F.B.; Barbosa, L.S.; Souza, T.M.A.; Badolato-Correa, J.; Sanchez-Arcila, J.C.; Nunes, P.C.G.; de-Oliveira-Pinto, L.M.; de Filippis, A.M.; Dal Fabbro, M.; et al. Clinical and Laboratory Profile of Zika and Dengue Infected Patients: Lessons Learned From the Co-circulation of Dengue, Zika and Chikungunya in Brazil. PLoS Curr. 2018, 10, 5843488. [Google Scholar] [CrossRef]
- De Souza, T.M.A.; Ribeiro, E.D.; Correa, V.C.E.; Damasco, P.V.; Santos, C.C.; de Bruycker-Nogueira, F.; Chouin-Carneiro, T.; Faria, N.; Nunes, P.C.G.; Heringer, M.; et al. Following in the Footsteps of the Chikungunya Virus in Brazil: The First Autochthonous Cases in Amapa in 2014 and Its Emergence in Rio de Janeiro during 2016. Viruses 2018, 10, 623. [Google Scholar] [CrossRef] [Green Version]
- Burt, F.J.; Chen, W.; Miner, J.J.; Lenschow, D.J.; Merits, A.; Schnettler, E.; Kohl, A.; Rudd, P.A.; Taylor, A.; Herrero, L.J.; et al. Chikungunya virus: An update on the biology and pathogenesis of this emerging pathogen. Lancet Infect. Dis. 2017, 17, e107–e117. [Google Scholar] [CrossRef]
- Lima, M.E.S.; Bachur, T.P.R.; Aragao, G.F. Guillain-Barre syndrome and its correlation with dengue, Zika and chikungunya viruses infection based on a literature review of reported cases in Brazil. Acta Trop. 2019, 197, 105064. [Google Scholar] [CrossRef]
- Chusri, S.; Siripaitoon, P.; Hirunpat, S.; Silpapojakul, K. Case reports of neuro-Chikungunya in southern Thailand. Am. J. Trop. Med. Hyg. 2011, 85, 386–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lima, E.C.B.; Januario, A.M.S.; Fortuna, E.G.; Real, M.E.C.; Lacerda, H.R. Chikungunya encephalitis: Report of a fatal case in Northeastern Brazil. Rev. Inst. Med. Trop. Sao Paulo 2020, 62, e40. [Google Scholar] [CrossRef] [PubMed]
- Morrison, T.E.; Oko, L.; Montgomery, S.A.; Whitmore, A.C.; Lotstein, A.R.; Gunn, B.M.; Elmore, S.A.; Heise, M.T. A mouse model of chikungunya virus-induced musculoskeletal inflammatory disease: Evidence of arthritis, tenosynovitis, myositis, and persistence. Am. J. Pathol. 2011, 178, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Couderc, T.; Lecuit, M. Chikungunya virus pathogenesis: From bedside to bench. Antivir. Res. 2015, 121, 120–131. [Google Scholar] [CrossRef]
- Chow, A.; Her, Z.; Ong, E.K.; Chen, J.M.; Dimatatac, F.; Kwek, D.J.; Barkham, T.; Yang, H.; Renia, L.; Leo, Y.S.; et al. Persistent arthralgia induced by Chikungunya virus infection is associated with interleukin-6 and granulocyte macrophage colony-stimulating factor. J. Infect Dis. 2011, 203, 149–157. [Google Scholar] [CrossRef] [Green Version]
- Ng, L.F.; Chow, A.; Sun, Y.J.; Kwek, D.J.; Lim, P.L.; Dimatatac, F.; Ng, L.C.; Ooi, E.E.; Choo, K.H.; Her, Z.; et al. IL-1beta, IL-6, and RANTES as biomarkers of Chikungunya severity. PLoS ONE 2009, 4, e4261. [Google Scholar] [CrossRef]
- Sanchez-Arcila, J.C.; Badolato-Correa, J.; de Souza, T.M.A.; Paiva, I.A.; Barbosa, L.S.; Nunes, P.C.G.; Lima, M.; Dos Santos, F.B.; Damasco, P.V.; da Cunha, R.V.; et al. Clinical, Virological, and Immunological Profiles of DENV, ZIKV, and/or CHIKV-Infected Brazilian Patients. Intervirology 2020, 63, 33–45. [Google Scholar] [CrossRef]
- Krishnan, S.M.; Mahalingam, J.; Sabarimurugan, S.; Muthu, T.; Venkidasamy, B.; Krishnasamy, K.; Sharma, A.; Ramalingam, S. Comparison of Cytokine Expression Profile in Chikungunya and Dengue Co-Infected and Mono-Infected Patients’ Samples. Pathogens 2021, 10, 166. [Google Scholar] [CrossRef]
- Becerra, A.; Warke, R.V.; Xhaja, K.; Evans, B.; Evans, J.; Martin, K.; de Bosch, N.; Rothman, A.L.; Bosch, I. Increased activity of indoleamine 2,3-dioxygenase in serum from acutely infected dengue patients linked to gamma interferon antiviral function. J. Gen. Virol. 2009, 90, 810–817. [Google Scholar] [CrossRef]
- Sun, X.; Hua, S.; Chen, H.R.; Ouyang, Z.; Einkauf, K.; Tse, S.; Ard, K.; Ciaranello, A.; Yawetz, S.; Sax, P.; et al. Transcriptional Changes during Naturally Acquired Zika Virus Infection Render Dendritic Cells Highly Conducive to Viral Replication. Cell Rep. 2017, 21, 3471–3482. [Google Scholar] [CrossRef] [Green Version]
- Fialho, L.G.; Torrentes-Carvalho, A.; Cunha, R.V.D.; Faria, N.R.; Cipitelli, M.D.C.; Gandini, M.; de Oliveira-Pinto, L.; De Azeredo, E.; Kubelka, C.F. Induced nitric oxide synthase (iNOS) and indoleamine 2,3-dioxygenase (IDO) detection in circulating monocyte subsets from Brazilian patients with Dengue-4 virus. Virol. Rep. 2017, 7, 9–19. [Google Scholar] [CrossRef]
- Ministerio da Saúde. Óbito por arboviroses no Brasil, 2008 a 2019. Distrito Federal, Brasil. Bol. Epidemiológico 2020, 51, 2020. [Google Scholar]
- Ministério da Saúde. Monitoramento dos casos de arboviroses urbanas causados por vírus transmitidos pelo mosquito Aedes (dengue, chikungunya e zika), semanas epidemioloígicas 1 a 47, 2021. Distrito Federal, Brasil. Bol. Epidemiológico 2021, 52, 2020. [Google Scholar]
- Ministério da Saúde. Febre da Chikungunya Manejo Clínico; Distrito Federal, Brasil. Ministério da Saúde: Brasília, Brasil, 2015.
- Brito, C.A.A.; Freitas, A.R.R.; Said, R.F.; Falcao, M.B.; Cunha, R.V.D.; Siqueira, A.M.; Teixeira, M.G.; Ribeiro, G.S.; Brito, M.C.M.; Cavalcanti, L.P.G. Classification of chikungunya cases: A proposal. Rev. Soc. Brasil Med. Trop. 2020, 53, e20200529. [Google Scholar] [CrossRef] [PubMed]
- Johnson, B.W.; Russell, B.J.; Goodman, C.H. Laboratory Diagnosis of Chikungunya Virus Infections and Commercial Sources for Diagnostic Assays. J. Infect Dis. 2016, 214 (Suppl. S5), S471–S474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Souza Sales, J.; Lara, F.A.; Amadeu, T.P.; de Oliveira Fulco, T.; da Costa Nery, J.A.; Sampaio, E.P.; Pinheiro, R.O.; Sarno, E.N. The role of indoleamine 2, 3-dioxygenase in lepromatous leprosy immunosuppression. Clin. Exp. Immunol. 2011, 165, 251–263. [Google Scholar] [CrossRef]
- Chirathaworn, C.; Chansaenroj, J.; Poovorawan, Y. Cytokines and Chemokines in Chikungunya Virus Infection: Protection or Induction of Pathology. Pathogens 2020, 9, 415. [Google Scholar] [CrossRef]
- Jacob-Nascimento, L.C.; Carvalho, C.X.; Silva, M.M.O.; Kikuti, M.; Anjos, R.O.; Fradico, J.R.B.; Campi-Azevedo, A.C.; Tauro, L.B.; Campos, G.S.; Moreira, P.; et al. Acute-Phase Levels of CXCL8 as Risk Factor for Chronic Arthralgia Following Chikungunya Virus Infection. Front. Immunol. 2021, 12, 744183. [Google Scholar] [CrossRef]
- Hoarau, J.J.; Jaffar Bandjee, M.C.; Krejbich Trotot, P.; Das, T.; Li-Pat-Yuen, G.; Dassa, B.; Denizot, M.; Guichard, E.; Ribera, A.; Henni, T.; et al. Persistent chronic inflammation and infection by Chikungunya arthritogenic alphavirus in spite of a robust host immune response. J. Immunol. 2010, 184, 5914–5927. [Google Scholar] [CrossRef] [Green Version]
- Young, A.R.; Locke, M.C.; Cook, L.E.; Hiller, B.E.; Zhang, R.; Hedberg, M.L.; Monte, K.J.; Veis, D.J.; Diamond, M.S.; Lenschow, D.J. Dermal and muscle fibroblasts and skeletal myofibers survive chikungunya virus infection and harbor persistent RNA. PLoS Pathog. 2019, 15, e1007993. [Google Scholar] [CrossRef]
- Labadie, K.; Larcher, T.; Joubert, C.; Mannioui, A.; Delache, B.; Brochard, P.; Guigand, L.; Dubreil, L.; Lebon, P.; Verrier, B.; et al. Chikungunya disease in nonhuman primates involves long-term viral persistence in macrophages. J. Clin. Invest. 2010, 120, 894–906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacKenzie, C.R.; Heseler, K.; Muller, A.; Daubener, W. Role of indoleamine 2,3-dioxygenase in antimicrobial defence and immuno-regulation: Tryptophan depletion versus production of toxic kynurenines. Curr. Drug. Metab. 2007, 8, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Bodaghi, B.; Goureau, O.; Zipeto, D.; Laurent, L.; Virelizier, J.L.; Michelson, S. Role of IFN-gamma-induced indoleamine 2,3 dioxygenase and inducible nitric oxide synthase in the replication of human cytomegalovirus in retinal pigment epithelial cells. J. Immunol. 1999, 162, 957–964. [Google Scholar] [PubMed]
- Adams, O.; Besken, K.; Oberdorfer, C.; MacKenzie, C.R.; Takikawa, O.; Daubener, W. Role of indoleamine-2,3-dioxygenase in alpha/beta and gamma interferon-mediated antiviral effects against herpes simplex virus infections. J. Virol. 2004, 78, 2632–2636. [Google Scholar] [CrossRef] [Green Version]
- Obojes, K.; Andres, O.; Kim, K.S.; Daubener, W.; Schneider-Schaulies, J. Indoleamine 2,3-dioxygenase mediates cell type-specific anti-measles virus activity of gamma interferon. J. Virol. 2005, 79, 7768–7776. [Google Scholar] [CrossRef] [Green Version]
- Geisler, S.; Lytton, S.D.; Toan, N.L.; Nghia, T.H.; Nam, N.M.; Hung, H.V.; Son, N.T.; Anh, D.T.; Tuyen, H.T.; Tien, T.V.; et al. Neopterin levels and Kyn/Trp ratios were significantly increased in dengue virus patients and subsequently decreased after recovery. Int. J. Infect. Dis. 2020, 91, 162–168. [Google Scholar] [CrossRef] [Green Version]
- Boasso, A.; Shearer, G.M. How does indoleamine 2,3-dioxygenase contribute to HIV-mediated immune dysregulation. Curr. Drug. Metab. 2007, 8, 217–223. [Google Scholar] [CrossRef]
- Yuan, F.L.; Li, X.; Lu, W.G.; Xu, R.S.; Zhao, Y.Q.; Li, C.W.; Li, J.P.; Chen, F.H. Regulatory T cells as a potent target for controlling bone loss. Biochem. Biophys. Res. Commun. 2010, 402, 173–176. [Google Scholar] [CrossRef]
- Zaiss, M.M.; Frey, B.; Hess, A.; Zwerina, J.; Luther, J.; Nimmerjahn, F.; Engelke, K.; Kollias, G.; Hunig, T.; Schett, G.; et al. Regulatory T cells protect from local and systemic bone destruction in arthritis. J. Immunol. 2010, 184, 7238–7246. [Google Scholar] [CrossRef]
- Lee, W.W.; Teo, T.H.; Her, Z.; Lum, F.M.; Kam, Y.W.; Haase, D.; Renia, L.; Rotzschke, O.; Ng, L.F. Expanding regulatory T cells alleviates chikungunya virus-induced pathology in mice. J. Virol. 2015, 89, 7893–7904. [Google Scholar] [CrossRef] [Green Version]
- Amaral, J.K.; Bilsborrow, J.B.; Schoen, R.T. Chronic Chikungunya Arthritis and Rheumatoid Arthritis: What They Have in Common. Am. J. Med. 2020, 133, e91–e97. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.Y.; Lee, S.H.; Jung, S.M.; Park, S.H.; Jung, B.H.; Ju, J.H. Downregulation of Tryptophan-related Metabolomic Profile in Rheumatoid Arthritis Synovial Fluid. J. Rheumatol. 2015, 42, 2003–2011. [Google Scholar] [CrossRef] [PubMed]
- Bertazzo, A.; Punzi, L.; Bertazzolo, N.; Pianon, M.; Pozzuoli, A.; Costa, C.V.; Allegri, G. Tryptophan catabolism in synovial fluid of various arthropathies and its relationship with inflammatory cytokines. Adv. Exp. Med. Biol. 1999, 467, 565–570. [Google Scholar] [PubMed]
- Criado, G.; Simelyte, E.; Inglis, J.J.; Essex, D.; Williams, R.O. Indoleamine 2,3 dioxygenase-mediated tryptophan catabolism regulates accumulation of Th1/Th17 cells in the joint in collagen-induced arthritis. Arthritis Rheum. 2009, 60, 1342–1351. [Google Scholar] [CrossRef]
- Forrest, C.M.; Kennedy, A.; Stone, T.W.; Stoy, N.; Darlington, L.G. Kynurenine and neopterin levels in patients with rheumatoid arthritis and osteoporosis during drug treatment. Adv. Exp. Med. Biol. 2003, 527, 287–295. [Google Scholar]
- Ferreira, A.S.; Baldoni, N.R.; Cardoso, C.S.; Oliveira, C.D.L. Biomarkers of severity and chronification in chikungunya fever: A systematic review and meta-analysis. Rev. Inst. Med. Trop. Sao Paulo 2021, 63, e16. [Google Scholar] [CrossRef]
- Wauquier, N.; Becquart, P.; Nkoghe, D.; Padilla, C.; Ndjoyi-Mbiguino, A.; Leroy, E.M. The acute phase of Chikungunya virus infection in humans is associated with strong innate immunity and T CD8 cell activation. J. Infect. Dis. 2011, 204, 115–123. [Google Scholar] [CrossRef]
- Lee, A.J.; Ashkar, A.A. The Dual Nature of Type I and Type II Interferons. Front. Immunol. 2018, 9, 2061. [Google Scholar] [CrossRef] [Green Version]
- Kokkonen, H.; Soderstrom, I.; Rocklov, J.; Hallmans, G.; Lejon, K.; Rantapaa Dahlqvist, S. Up-regulation of cytokines and chemokines predates the onset of rheumatoid arthritis. Arthritis Rheum. 2010, 62, 383–391. [Google Scholar] [CrossRef]
- Teo, T.H.; Lum, F.M.; Claser, C.; Lulla, V.; Lulla, A.; Merits, A.; Renia, L.; Ng, L.F. A pathogenic role for CD4+ T cells during Chikungunya virus infection in mice. J. Immunol. 2013, 190, 259–269. [Google Scholar] [CrossRef] [Green Version]
- Chopra, A.; Anuradha, V.; Ghorpade, R.; Saluja, M. Acute Chikungunya and persistent musculoskeletal pain following the 2006 Indian epidemic: A 2-year prospective rural community study. Epidemiol. Infect. 2012, 140, 842–850. [Google Scholar] [CrossRef] [PubMed]
- Schilte, C.; Staikowsky, F.; Couderc, T.; Madec, Y.; Carpentier, F.; Kassab, S.; Albert, M.L.; Lecuit, M.; Michault, A. Chikungunya virus-associated long-term arthralgia: A 36-month prospective longitudinal study. PLoS Negl. Trop. Dis. 2013, 7, e2137. [Google Scholar] [CrossRef]
- Lohachanakul, J.; Phuklia, W.; Thannagith, M.; Thonsakulprasert, T.; Ubol, S. High concentrations of circulating interleukin-6 and monocyte chemotactic protein-1 with low concentrations of interleukin-8 were associated with severe chikungunya fever during the 2009-2010 outbreak in Thailand. Microbiol. Immunol. 2012, 56, 134–138. [Google Scholar] [CrossRef] [PubMed]
- Ninla-Aesong, P.; Mitarnun, W.; Noipha, K. Proinflammatory Cytokines and Chemokines as Biomarkers of Persistent Arthralgia and Severe Disease After Chikungunya Virus Infection: A 5-Year Follow-Up Study in Southern Thailand. Viral Immunol. 2019, 32, 442–452. [Google Scholar] [CrossRef]
- Deshmane, S.L.; Kremlev, S.; Amini, S.; Sawaya, B.E. Monocyte chemoattractant protein-1 (MCP-1): An overview. J. Interferon. Cytokine Res. 2009, 29, 313–326. [Google Scholar] [CrossRef]
- Neville, L.F.; Mathiak, G.; Bagasra, O. The immunobiology of interferon-gamma inducible protein 10 kD (IP-10): A novel, pleiotropic member of the C-X-C chemokine superfamily. Cytokine Growth Factor Rev. 1997, 8, 207–219. [Google Scholar] [CrossRef]
- Rulli, N.E.; Rolph, M.S.; Srikiatkhachorn, A.; Anantapreecha, S.; Guglielmotti, A.; Mahalingam, S. Protection from arthritis and myositis in a mouse model of acute chikungunya virus disease by bindarit, an inhibitor of monocyte chemotactic protein-1 synthesis. J. Infect Dis. 2011, 204, 1026–1030. [Google Scholar] [CrossRef]
- Maurer, M.; von Stebut, E. Macrophage inflammatory protein-1. Int. J. Biochem. Cell. Biol. 2004, 36, 1882–1886. [Google Scholar] [CrossRef]
- Thiberville, S.D.; Moyen, N.; Dupuis-Maguiraga, L.; Nougairede, A.; Gould, E.A.; Roques, P.; de Lamballerie, X. Chikungunya fever: Epidemiology, clinical syndrome, pathogenesis and therapy. Antivir. Res. 2013, 99, 345–370. [Google Scholar] [CrossRef]
- Maghazachi, A.A. Role of chemokines in the biology of natural killer cells. Curr. Top Microbiol. Immunol. 2010, 341, 37–58. [Google Scholar]
- Thanapati, S.; Das, R.; Tripathy, A.S. Phenotypic and functional analyses of NK and NKT-like populations during the early stages of chikungunya infection. Front. Microbiol. 2015, 6, 895. [Google Scholar] [CrossRef] [PubMed]
- Petitdemange, C.; Becquart, P.; Wauquier, N.; Beziat, V.; Debre, P.; Leroy, E.M.; Vieillard, V. Unconventional repertoire profile is imprinted during acute chikungunya infection for natural killer cells polarization toward cytotoxicity. PLoS Pathog. 2011, 7, e1002268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teo, T.H.; Her, Z.; Tan, J.J.; Lum, F.M.; Lee, W.W.; Chan, Y.H.; Ong, R.Y.; Kam, Y.W.; Leparc-Goffart, I.; Gallian, P.; et al. Caribbean and La Reunion Chikungunya Virus Isolates Differ in Their Capacity To Induce Proinflammatory Th1 and NK Cell Responses and Acute Joint Pathology. J. Virol. 2015, 89, 7955–7969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, D.; Zlotnik, A. The biology of chemokines and their receptors. Annu. Rev. Immunol. 2000, 18, 217–242. [Google Scholar] [CrossRef]
- Campbell, B.M.; Charych, E.; Lee, A.W.; Moller, T. Kynurenines in CNS disease: Regulation by inflammatory cytokines. Front. Neurosci. 2014, 8, 12. [Google Scholar] [CrossRef] [Green Version]
- Agere, S.A.; Akhtar, N.; Watson, J.M.; Ahmed, S. RANTES/CCL5 Induces Collagen Degradation by Activating MMP-1 and MMP-13 Expression in Human Rheumatoid Arthritis Synovial Fibroblasts. Front. Immunol. 2017, 8, 1341. [Google Scholar] [CrossRef] [Green Version]
- Lanciotti, R.S.; Calisher, C.H.; Gubler, D.J.; Chang, G.J.; Vorndam, A.V. Rapid detection and typing of dengue viruses from clinical samples by using reverse transcriptase-polymerase chain reaction. J. Clin. Microbiol. 1992, 30, 545–551. [Google Scholar] [CrossRef] [Green Version]
- Lanciotti, R.S.; Kosoy, O.L.; Laven, J.J.; Velez, J.O.; Lambert, A.J.; Johnson, A.J.; Stanfield, S.M.; Duffy, M.R. Genetic and serologic properties of Zika virus associated with an epidemic, Yap State, Micronesia, 2007. Emerg. Infect. Dis. 2008, 14, 1232–1239. [Google Scholar] [CrossRef]
- Vieira, C.J.; Silva, D.J.; Barreto, E.S.; Siqueira, C.E.; Colombo, T.E.; Ozanic, K.; Schmidt, D.J.; Drumond, B.P.; Mondini, A.; Nogueira, M.L.; et al. Detection of Mayaro virus infections during a dengue outbreak in Mato Grosso, Brazil. Acta Trop. 2015, 147, 12–16. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chikungunya n (%) | |||
|---|---|---|---|
| Acute Phase n = 78 | Post-Acute Phase n = 12 | Chronic Phase n = 10 | |
| Gender | |||
| Female n (%) | 45 (57.7) | 7 (58.3) | 6 (60) |
| Male | 33 (42.3) | 5 (41.7) | 4 (40) |
| Age | |||
| Years 1 | 45 (28–57) | 54 (36.7–65) | 60 (52–66.2) |
| Days after disease onset | 3 (2.0–5.5) | 30 (20.5–51) | 91 (91–110) |
| Laboratorial Diagnosis | |||
| RT-PCR | 78 (100) | 5 (41) | 8 (80) |
| CHIKV Viral Load | |||
| Ct values 1 | 24.8 (18–32.6) | 35.1 (26.1–36.6) | 30.1 (27.5–30.9) |
| Viral load Log copies /mL | 4.9 (3.1–6.2) | 3.0 (2.9–4.1) | 4.1 (3.7–4.2) |
| Anti-CHIKV antibodies | |||
| IgM | 35 (44.8) | 12 (100) | 10 (100) |
| IgG | 6 (7.6) | 12 (100) | 10 (100) |
| Signs/Symptoms | |||
| Fever | 72 (92) | 3 (25) | 0 |
| Headache | 57 (73) | 8 (66) | 0 |
| Running nose | 6 (7.6) | 0 | 0 |
| Cough | 7 (8.9) | 0 | 0 |
| Retroorbital pain | 38 (48) | 4 (33) | 0 |
| Myalgia | 54 (69) | 6 (50) | 5 (50) |
| Low back pain | 48 (61) | 4 (33) | 0 |
| Arthralgia | 73 (93) | 12 (100) | 10 (100) |
| Polyarthritis | 25 (32) | 4 (33) | 6 (60) |
| Edema | 40 (51) | 7 (58) | 6 (60) |
| Anorexia | 53 (67) | 5 (41) | 0 |
| Prostration | 59 (75) | 6 (50) | 5 (50) |
| Dizziness | 23 (29) | 2 (16) | 0 |
| Nausea | 40 (51) | 3 (25) | 0 |
| Vomiting | 17 (21) | 2 (16) | 0 |
| Abdominal pain | 13 (16) | 0 | 0 |
| Pruritus | 34 (43) | 2 (16) | 0 |
| Exanthema | 41 (52) | 2 (16) | 0 |
| Paresthesia | 8 (10) | 2 (16) | 2 (20) |
| Conjunctival hyperemia | 26 (33) | 0 | 0 |
| Hospitalization | |||
| 3 (3.5) | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alves de Souza, T.M.; Fernandes-Santos, C.; Araújo da Paixão de Oliveira, J.; Tomé, L.C.T.; Fiestas-Solórzano, V.E.; Nunes, P.C.G.; Guimaraes, G.M.C.; Sánchez-Arcila, J.C.; Paiva, I.A.; de Souza, L.J.; et al. Increased Indoleamine 2,3-Dioxygenase 1 (IDO-1) Activity and Inflammatory Responses during Chikungunya Virus Infection. Pathogens 2022, 11, 444. https://doi.org/10.3390/pathogens11040444
Alves de Souza TM, Fernandes-Santos C, Araújo da Paixão de Oliveira J, Tomé LCT, Fiestas-Solórzano VE, Nunes PCG, Guimaraes GMC, Sánchez-Arcila JC, Paiva IA, de Souza LJ, et al. Increased Indoleamine 2,3-Dioxygenase 1 (IDO-1) Activity and Inflammatory Responses during Chikungunya Virus Infection. Pathogens. 2022; 11(4):444. https://doi.org/10.3390/pathogens11040444
Chicago/Turabian StyleAlves de Souza, Thiara Manuele, Caroline Fernandes-Santos, Jéssica Araújo da Paixão de Oliveira, Larissa Cristina Teixeira Tomé, Victor Edgar Fiestas-Solórzano, Priscila Conrado Guerra Nunes, Gabriel Macedo Costa Guimaraes, Juan Camilo Sánchez-Arcila, Iury Amâncio Paiva, Luís Jose de Souza, and et al. 2022. "Increased Indoleamine 2,3-Dioxygenase 1 (IDO-1) Activity and Inflammatory Responses during Chikungunya Virus Infection" Pathogens 11, no. 4: 444. https://doi.org/10.3390/pathogens11040444