

Host–Parasite Interaction and Phylogenetic of a New Cnidarian Myxosporean (Endocnidozoa: Myxobolidae) Infecting a Valuative Commercialized Ornamental Fish from Pantanal Wetland Biome, Brazil
 , and
, and
Abstract
:1. Introduction
2. Material and Methods
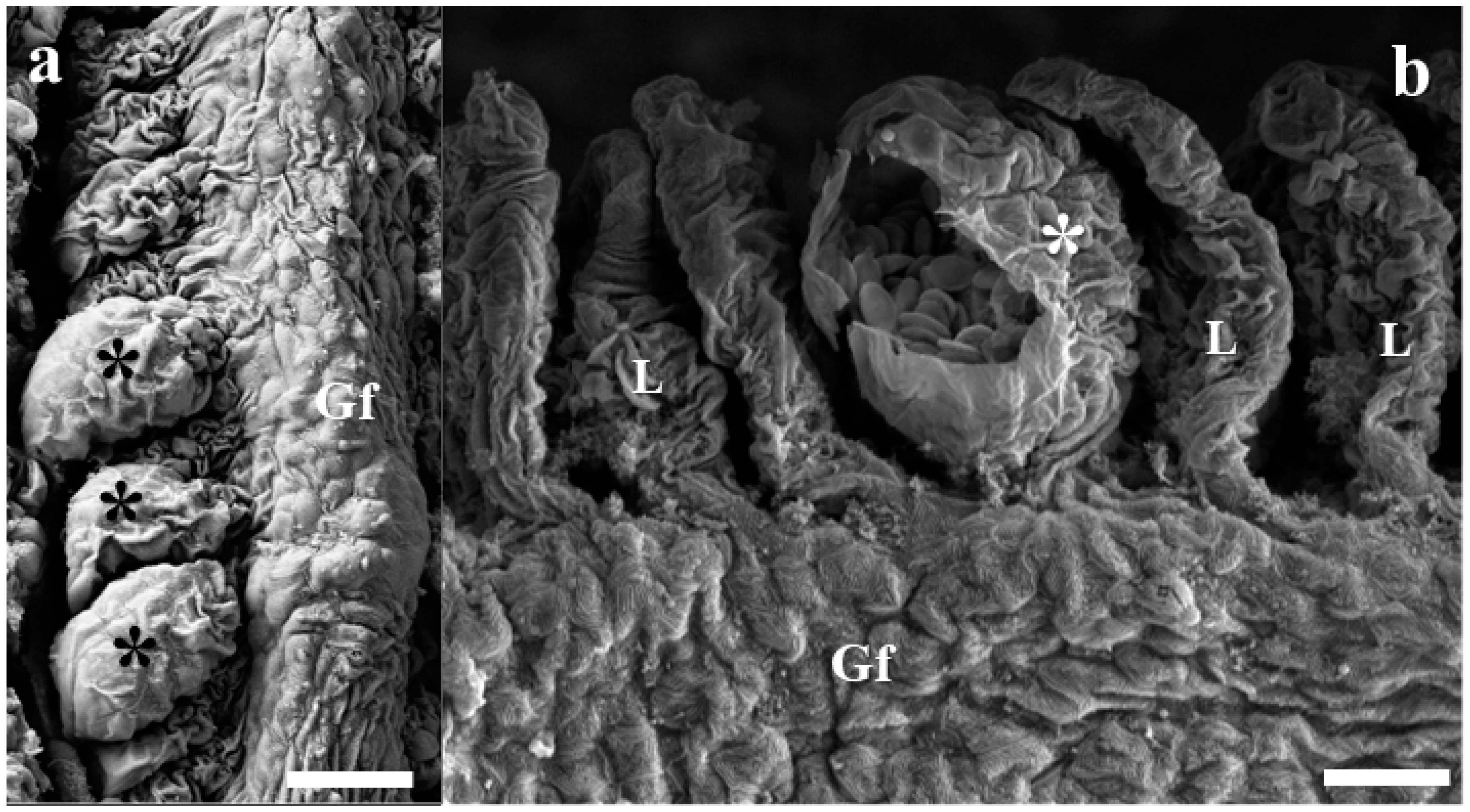
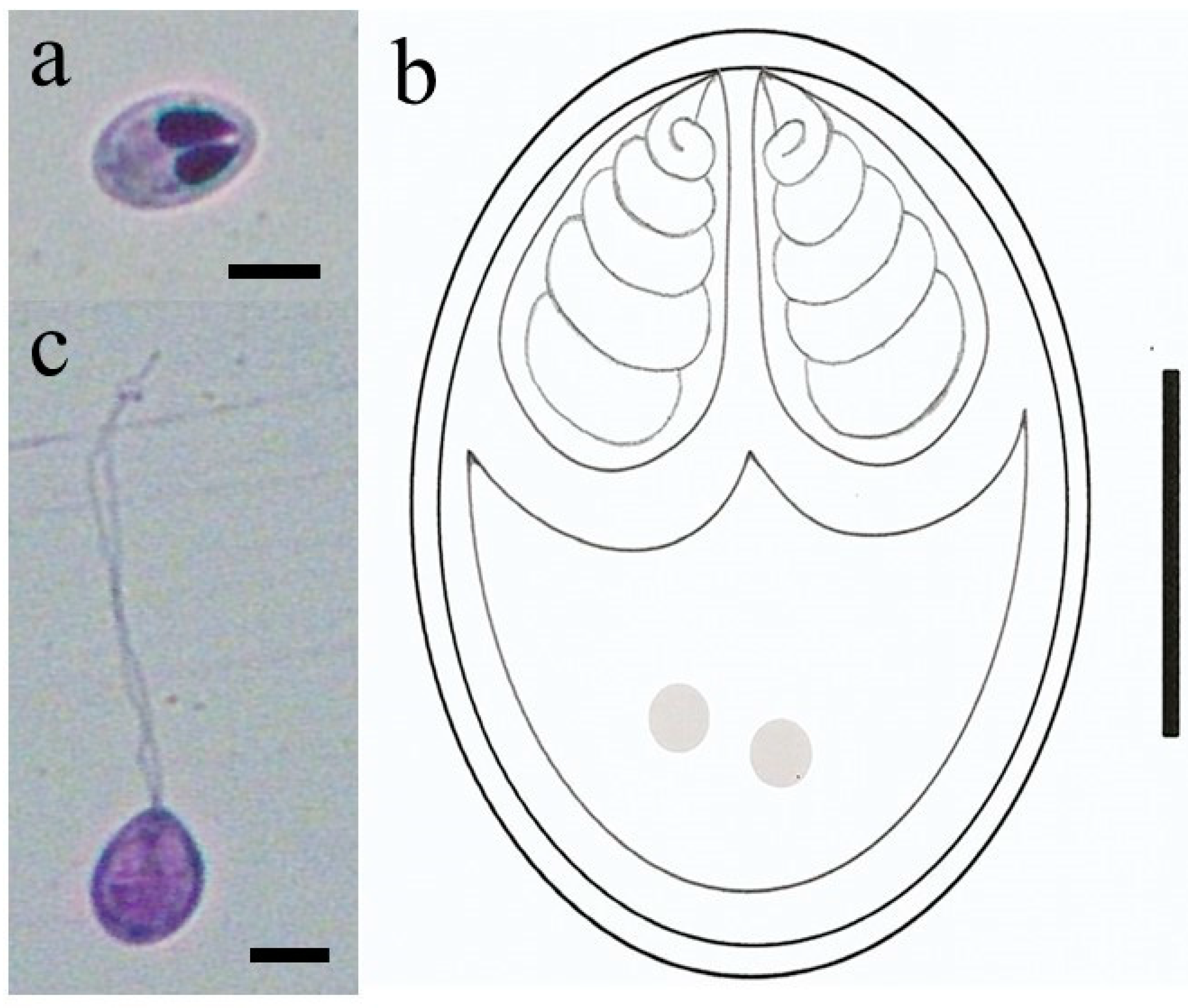
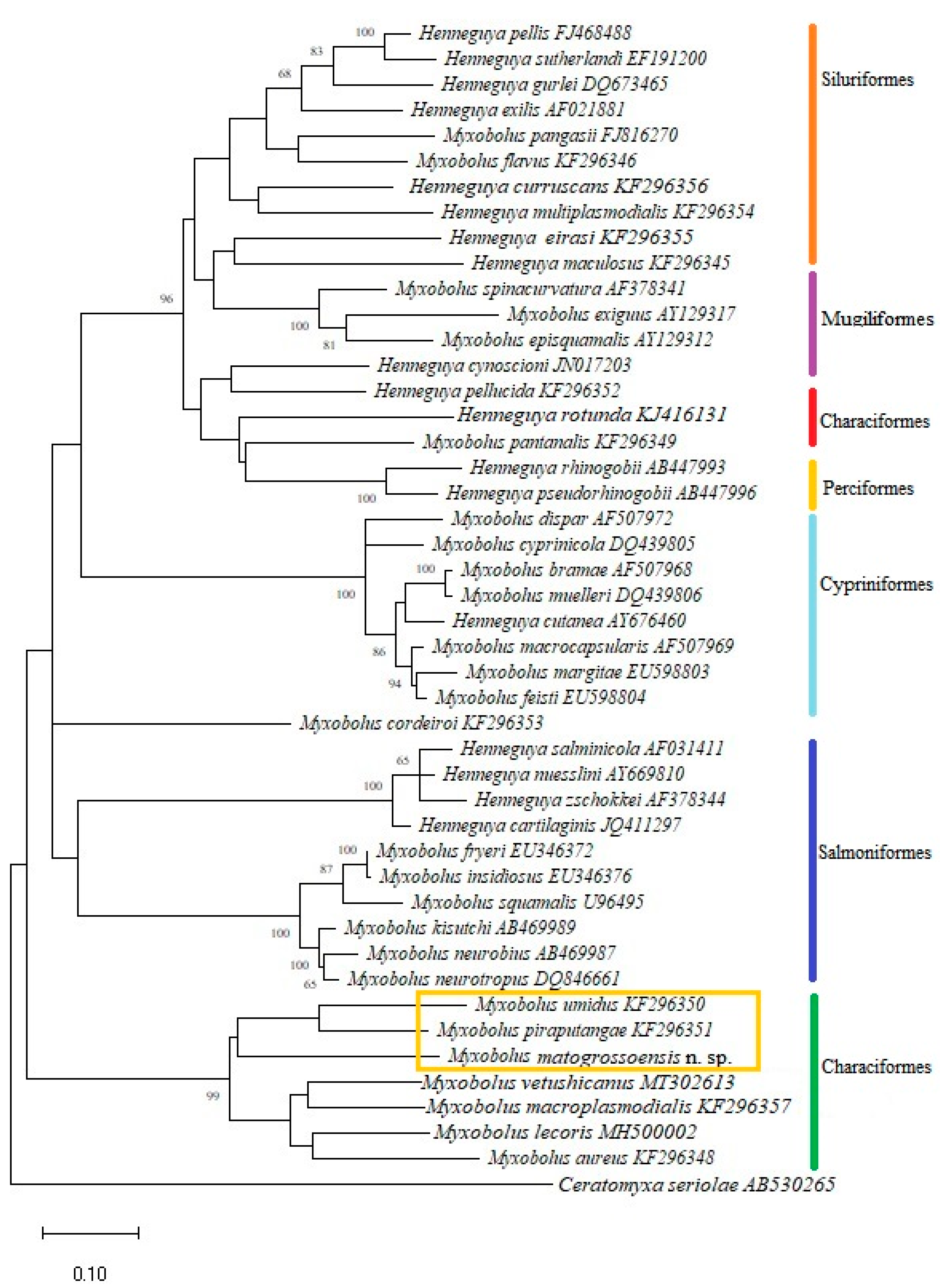
3. Results
- Phylum: Cnidaria Verrill, 1865
- Class: Myxosporea Bütschli, 1881
- Order: Bivalvulida Shulman, 1959
- Family: Myxobolidae Thélohan, 1892
- Genus: Myxobolus Bütschli, 1882
- Species: Myxobolus matogrossoensis n. sp.
- Type host: Hyphessobrycon eques (Characiformes: Characidae)
- Site of infection: Gills (Interlamellar-epithelial type, LE2)
- Gill plasmodium index (GPI): 1 (light infection)
- Category of plasmodium: Type A (visible under light microscope, size range 40–65 μm)
- Type of locality: Adjacent area of lakes of Paraguay River near the municipality Porto Murtinho, Mato Grosso do Sul State, Brazil (21°41′56″ S, 57°52′58″ W).
- Prevalence: From 30 examined fish, three were infected (10%).
- Type of material: Hapantotype (slides with stained myxospores) were deposited in the cnidarian collection of the Zoology Museum at the University of São Paulo—MZUSP, São Paulo, Brazil (slide no. MZUSP 8695). Partial 18S rDNA sequence gene was deposited in GenBank under accession number OP244900.
- Etymology: The specific name (M. matogrossoensis) is based on host species common name.
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Atkinson, S.D.; Bartholomew, J.L.; Lotan, T. Myxozoans: Ancient metazoan parasites find a home in phylum Cnidaria. Zoology 2018, 129, 66–68. [Google Scholar] [CrossRef] [PubMed]
- Okamura, B.; Gruhl, A.; Bartholomew, J.L. (Eds.) An introduction to Myxozoan evolution, ecology and development. In Myxozoan Evolution, Ecology and Development; Springer: Cham, Switzerland, 2015; pp. 1–20. [Google Scholar]
- Eiras, J.C.; Cruz, C.F.; Saraiva, A.; Adriano, E.A. Synopsis of the species of Myxobolus (Cnidaria, Myxozoa, Myxosporea) described between 2014 and 2020. Folia Parasitol. 2021, 68, 012. [Google Scholar] [CrossRef] [PubMed]
- Mathews, P.D.; Bonillo, C.; Rabet, N.; Lord, C.; Causse, R.; Keith, P.; Audebert, F. Phylogenetic analysis and characterization of a new parasitic cnidarian (Myxosporea: Myxobolidae) parasitizing skin of the giant mottled eel from the Solomon Islands. Infec. Genet. Evol. 2021, 94, 104986. [Google Scholar] [CrossRef] [PubMed]
- Fariya, N.; Kaur, H.; Singh, M.; Abidi, R.; El-Matbouli, M.; Kumar, G. Morphological and Molecular Characterization of a New Myxozoan, Myxobolus grassi sp. nov. (Myxosporea), Infecting the Grass Carp, Ctenopharyngodon idella in the Gomti River, India. Pathogens 2022, 11, 303. [Google Scholar] [CrossRef] [PubMed]
- Milanin, T.; Mathews, P.D.; Morandini, A.C.; Mertins, O.; Audebert, F.; Pereira, J.O.L.; Maia, A.A.M. Morphostructural data and phylogenetic relationships of a new cnidarian myxosporean infecting spleen of an economic and ecological important bryconid fish from Brazil. Microb. Pathog. 2021, 150, 104718. [Google Scholar] [CrossRef]
- Mathews, P.D.; Mertins, O.; Milanin, T.; Aguiar, J.C.; Gonzales-Flores, A.P.P.; Tavares, L.E.R.; Morandini, A.C. Ultrastructure, surface topography, morphology and histological observations of a new parasitic cnidarian of the marbled swamp eel from the world’s largest tropical wetland area, Pantanal, Brazil. Tissue Cell 2022, 79, 101909. [Google Scholar] [CrossRef]
- Florentino, A.C.; Petrere, M.; Freitas, C.E.D.C.; Toledo, J.J.; Mateus, L.; Súarez, Y.R.; Penha, J. Determinants of changes in fish diversity and composition in floodplain lakes in two basins in the Pantanal wetlands, Brazil. Environ. Biol. Fish. 2016, 99, 265–274. [Google Scholar] [CrossRef]
- Carriero, M.M.; Adriano, E.A.; Silva, M.R.M.; Ceccarelli, P.S.; Maia, A.A.M. Molecular phylogeny of the Myxobolus and Henneguya genera with several new south American species. PLoS ONE 2013, 8, e73713. [Google Scholar] [CrossRef]
- García-Alzate, C.A.; Urbano-Bonilla, A.; Taphorn, D.C. A new species of Hyphessobrycon (Characiformes, Characidae) from the upper Guaviare River, Orinoco River Basin, Colombia. ZooKeys 2017, 668, 123–138. [Google Scholar] [CrossRef]
- Pelicice, F.M.; Agostinho, A.A. Feeding ecology of fishes associated with Egeria spp. patches in a tropical reservoir, Brazil. Ecol. Freshw. Fish 2006, 15, 10–19. [Google Scholar] [CrossRef]
- Lom, J.; Arthur, J.R. A guideline for the preparation of species descriptions in Myxosporea. J. Fish Dis. 1989, 12, 151–156. [Google Scholar] [CrossRef]
- Kaur, H.; Attri, R. Morphological and molecular characterization of Henneguya bicaudi n. sp. (Myxosporea: Myxobolidae) infecting gills of Cirrhinus mrigala (Ham.) in Harike Wetland, Punjab (India). Parasitol. Res. 2015, 114, 4161–4167. [Google Scholar] [CrossRef] [PubMed]
- Kaur, H.; Katoch, A. Prevalence, site and tissue preference of myxozoan parasites infecting gills of cultured fish in Punjab (India). Dis. Aquat. Org. 2016, 118, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Molnár, K.; Eszterbauer, E.; Sczekely, C.; Benko, M.; Harrach, B. Morphological and molecular biological studies on intramuscular Myxobolus spp. of cyprinid fish. J. Fish Dis. 2002, 25, 643–652. [Google Scholar] [CrossRef]
- Barta, J.R.; Martin, D.S.; Liberator, P.A.; Dashkevicz, M.; Anderson, J.W.; Feighner, S.D.; Elbrecht, A.; Perkins-Barrow, A.; Jenkins, M.C.; Danforth, D.; et al. Phylogenetic relationships among eight Eimeria species infecting domestic fowl inferred using complete small subunit ribosomal DNA sequences. J. Parasitol. 1997, 83, 262–271. [Google Scholar] [CrossRef]
- Molnár, K. Site preference of fish myxosporeans in the gill. Dis. Aquat. Org. 2002, 48, 197–207. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.H.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA 6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Okamura, B.; Hartigan, A.; Naldoni, J. Extensive uncharted biodiversity: The parasite dimension. Integr. Comp. Biol. 2018, 58, 1132–1145. [Google Scholar] [CrossRef]
- Naldoni, J.; Arana, S.; Maia, A.A.M.; Silva, M.R.M.; Carriero, M.M.; Ceccarelli, P.S.; Tavares, L.E.R.; Adriano, E.A. Host–parasite–environment relationship, morphology and molecular analyses of Henneguya eirasi n. sp. parasite of two wild Pseudoplatystoma spp. in Pantanal wetland, Brazil. Vet. Parasitol. 2011, 177, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Fiala, I.; Bartosova-Sojkova, P.; Whipps, C.M. Classification and Phylogenetics of Myxozoa. In Myxozoan Evolution, Ecology and Development; Okamura, B., Gruhl, A., Bartholomew, J.L., Eds.; Springer: Cham, Switzerland, 2015; pp. 85–110. [Google Scholar]
- Rocha, S.; Azevedo, C.; Alves, Â.; Antunes, C.; Casal, G. Morphological and molecular characterization of myxobolids (Cnidaria, Myxozoa) infecting cypriniforms (Actinopterygii, Teleostei) endemic to the Iberian Peninsula. Parasite 2019, 26, 48. [Google Scholar] [CrossRef] [PubMed]
- Milanin, T.; Atkinson, S.D.; Silva, M.R.; Alves, R.G.; Maia, A.A.; Adriano, E.A. Occurrence of two novel actinospore types (Cnidaria: Myxosporea) in Brazilian fish farms, and the creation of a novel actinospore collective group, Seisactinomyxon. Acta Parasitol. 2017, 62, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Mathews, P.D.; Mertins, O.; Milanin, T.; Espinoza, L.L.; Flores-Gonzales, A.P.; Audebert, F.; Morandini, A.C. Molecular phylogeny and taxonomy of a new Myxobolus species from the endangered ornamental fish, Otocinclus cocama endemic to Peru: A host parasite coextinction approach. Acta Trop. 2020, 210, 105545. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Lovy, A.; Gu, Z.; Fiala, I. Phylogeny of Myxobolidae (Myxozoa) and the evolution of myxospore appendages in the Myxobolus clade. Int. J. Parasitol. 2019, 49, 523–530. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Myxobolus Species | SL | SW | ST | PCL | PCW | NC | Site of Infection |
| This study | 6.6 ± 0.4 | 3.5 ± 0.2 | - | 3.3 ± 0.2 | 1.8 ± 0.1 | 3–4 | Gill lamelae |
| M. sp. Mathews, Mertins, Milanin, Aguiar, Gonzales-Flores, Tavares & Morandini, 2022 | 22.7 ± 1.2 (21.5–23.9) | 12.5 ± 0.4 (12.1–12.9) | 11.3 ± 0.5 (10.8–11.8) | 4.6 ± 0.3 (3.9 ± 4.3) | 2.9 ± 0.1 (2.8–3.0) | 4–5 | Mandibule |
| M. cordeiroi. Adriano, Arana, Alves, Silva, Ceccarelli, Henrique-Silva & Maia, 2009 | 10.8 ± 0.5 | 7.1 ± 0.2 | 5.3 ± 0.3 | 5.2 ± 0.3 | 1.4 ± 0.1 | 5–6 | Gill arch, skin, body, eye |
| M. salminus. Adriano, Arana, Carriero, Naldoni, Ceccarelli & Maia, 2009 | 9.6–10.5 (10.1 ± 0.4) | 5.8–6.6 (6.1 ± 0.4) | 4.7–5.3 (5.0 ± 0.6) | 4.3–4.8 (4.6 ± 0.2) | 1.5–1.9 (1.7 ± 0.1) | 7–8 | Gill filament |
| M. oliveirai. Milanin, Eiras, Arana, Maia, Alves, Silva, Carriero, Ceccarelli & Adriano, 2010 | 11.2 ± 0.4 | 7.4 ± 0.5 | 4.6 ± 0.6 | 5.6 ± 0.2 | 2.3 ± 0.2 | 6–8 | Gill filament |
| M. brycon. Azevedo, Casal, Marques, Silva & Matos, 2011 | 6.9 ± 0.6 (6.5–7.2) | 3.9–4.8 (4.2 ± 0.5) | 1.9–2.8 (2.5 ± 0.7) | 3.8–4.7 (4.2 ± 0.6) | 1.7–2.5 (1.9 ± 0.6) | 8–9 | Gill filament |
| M. flavus. Carriero, Adriano, Silva, Ceccarelli, Maia, 2013 | 9.2 ± 0.2 | 6.5 ± 0.3 | 4.2 ± 0.2 | 4.5 ± 0.2 | 1.6 ± 0.1 | 4–5 | Gill arch |
| M. pantanalis. Carriero, Adriano, Silva, Ceccarelli, Maia, 2013 | 9.3 ± 0.4 | 6.5 ± 0.4 | - | 4.2 ± 0.5 | 2.0 ± 0.1 | 4 | Gill filament |
| M. aureus. Carriero, Adriano, Silva, Ceccarelli, Maia, 2013 | 12.6 ± 0.5 | 8.3 ± 0.3 | 5.5 ± 0.3 | 5.7 ± 0.3 | 2.9 ± 0.2 | 7–8 | Liver |
| M. umidus. Carriero, Adriano, Silva, Ceccarelli, Maia, 2013 | 13.5 ± 0.7 | 7.8 ± 0.4 | 7.7 ± 0.1 | 5.1 ± 0.4 | 2.7 ± 0.3 | 5 | Spleen |
| M. piraputungae. Carriero, Adriano, Silva, Ceccarelli, Maia, 2013 | 10.1 ± 0.5 | 8.7 ± 0.5 | 6.7 ± 0.3 | 5.2 ± 0.4 | 3.0 ± 0.3 | 4–5 | Kidney |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mathews, P.D.; Mertins, O.; Flores-Gonzales, A.P.P.; Espinoza, L.L.; Aguiar, J.C.; Milanin, T. Host–Parasite Interaction and Phylogenetic of a New Cnidarian Myxosporean (Endocnidozoa: Myxobolidae) Infecting a Valuative Commercialized Ornamental Fish from Pantanal Wetland Biome, Brazil. Pathogens 2022, 11, 1119. https://doi.org/10.3390/pathogens11101119
Mathews PD, Mertins O, Flores-Gonzales APP, Espinoza LL, Aguiar JC, Milanin T. Host–Parasite Interaction and Phylogenetic of a New Cnidarian Myxosporean (Endocnidozoa: Myxobolidae) Infecting a Valuative Commercialized Ornamental Fish from Pantanal Wetland Biome, Brazil. Pathogens. 2022; 11(10):1119. https://doi.org/10.3390/pathogens11101119
Chicago/Turabian StyleMathews, Patrick D., Omar Mertins, Anai P. P. Flores-Gonzales, Luis L. Espinoza, Julio C. Aguiar, and Tiago Milanin. 2022. "Host–Parasite Interaction and Phylogenetic of a New Cnidarian Myxosporean (Endocnidozoa: Myxobolidae) Infecting a Valuative Commercialized Ornamental Fish from Pantanal Wetland Biome, Brazil" Pathogens 11, no. 10: 1119. https://doi.org/10.3390/pathogens11101119