Molecular Typing, Antibiogram and PCR-RFLP Based Detection of Aeromonas hydrophila Complex Isolated from Oreochromis niloticus

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fish Sampling
2.2. Clinical Examination
2.3. Bacteriological Examination
2.4. Molecular Typing of A. hydrophila Complex and its Virulence Genes
2.5. Restriction Fragment Length Polymorphism (RFLP) of A. hyrophila Complex
2.6. Antimicrobial Susceptibility Testing
2.7. Pathogenicity Test
2.7.1. Acclimation Period
2.7.2. Challenge Trial
2.8. Statistical Analysis
3. Results
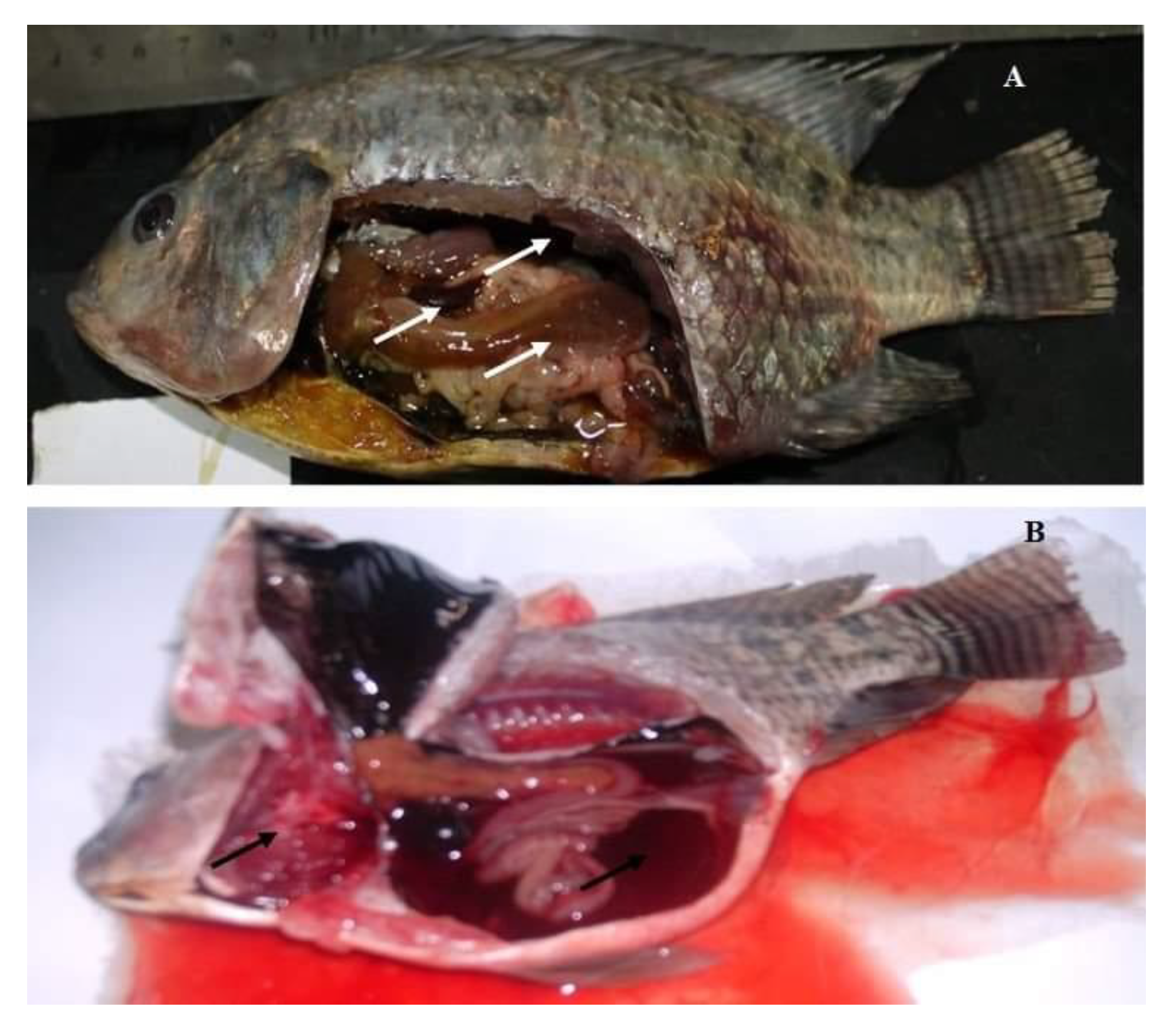
3.1. Clinical Findings
3.2. Bacteriological Examination
3.3. Prevalence of A. hydrophila Complex among Naturally Infected O. niloticus and Their Intensities in Different Internal Organs
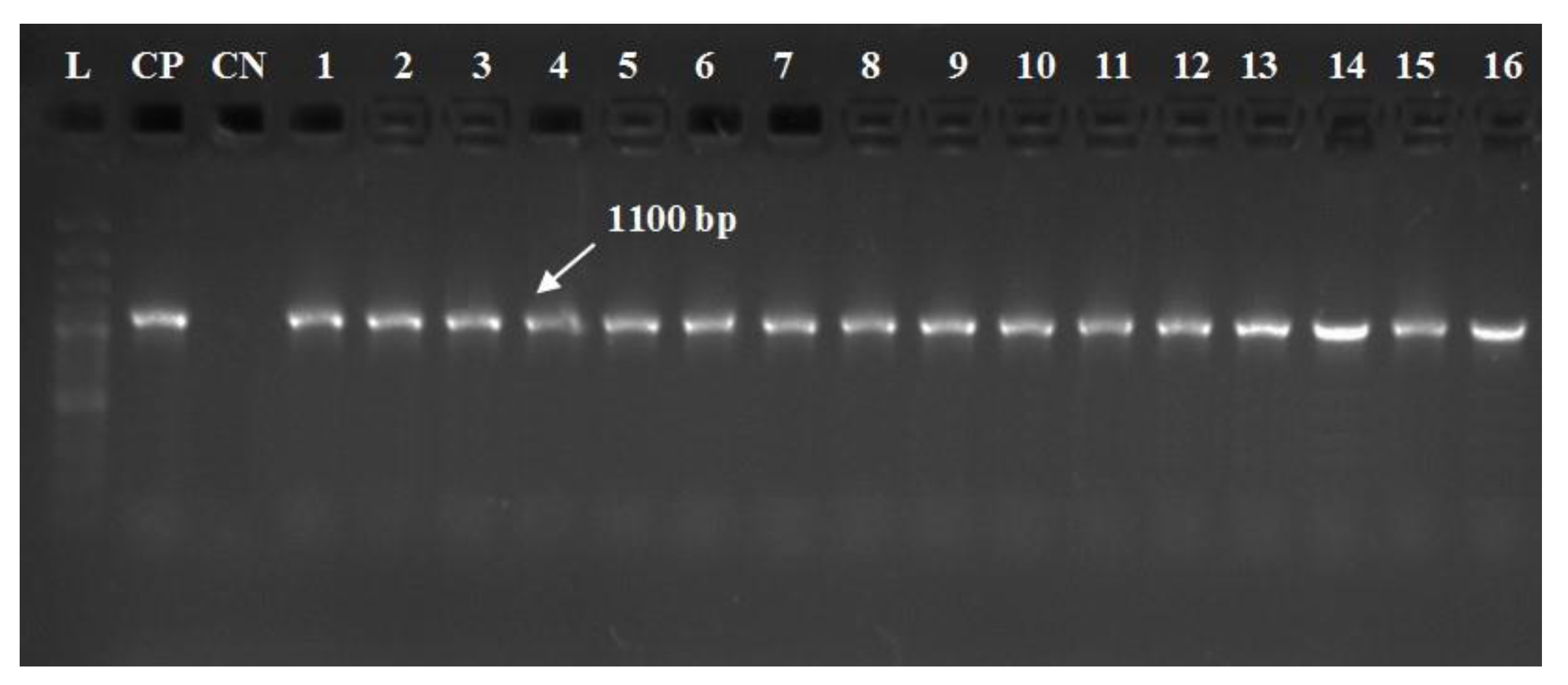
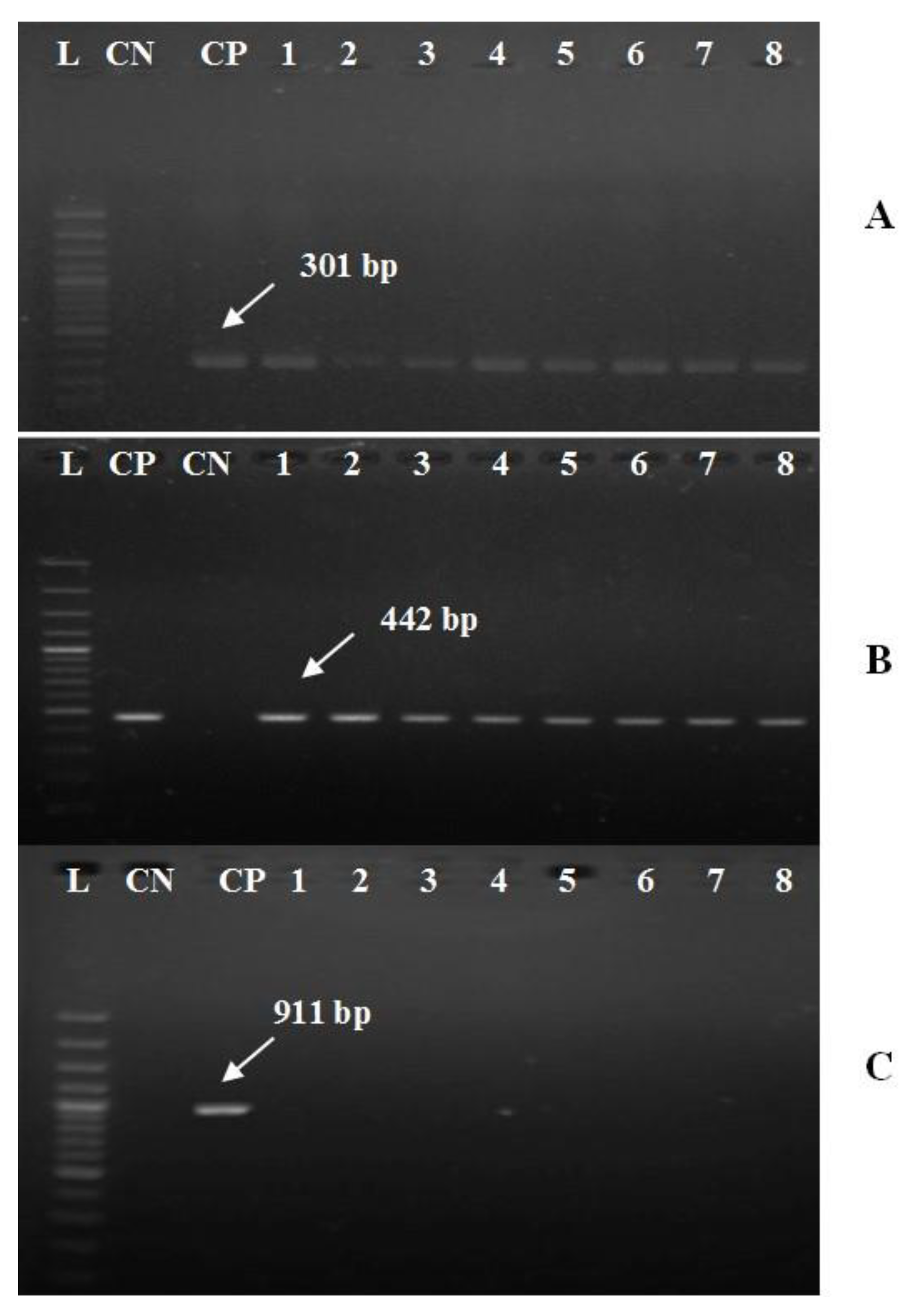
3.4. Molecular Typing of A. hydrophila Complex and Their Virulent Genes
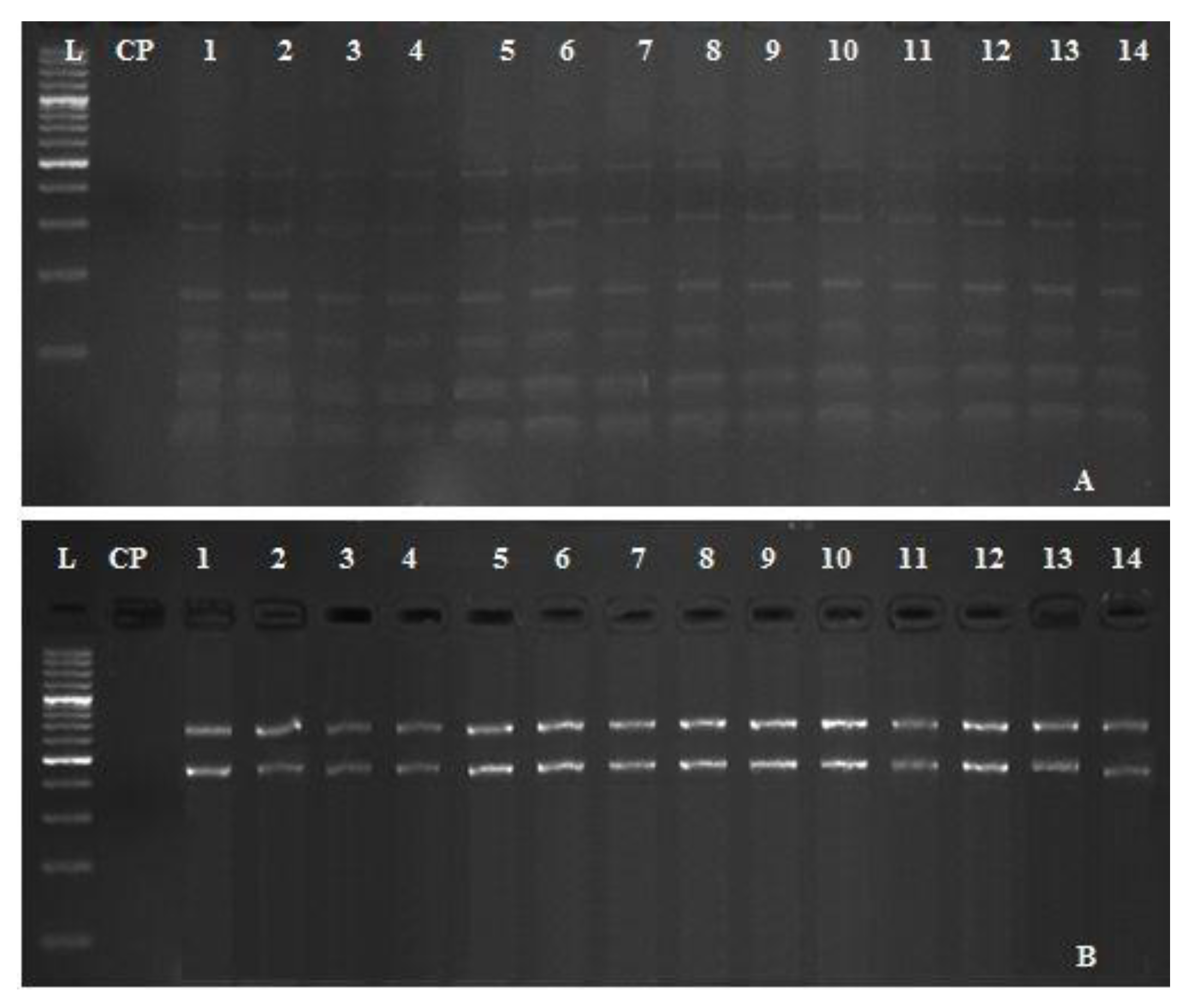
3.5. Restriction Fragment Length Polymorphisms (RFLP) of A. hydrophila Complex
3.6. Antimicrobial Susceptibility Testing
3.7. Pathogenicity (Challenge Trial)
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Buentello, J.A.; Reyes-Becerril, M.; de Jesús Romero-Geraldo, M.; de Jesús Ascencio-Valle, F. Effects of dietary arginine on hematological parameters and innate immune function of channel catfish. J. Aquat. Anim. Health 2007, 19, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Pakingking, R.; Palma, P.; Usero, R. Quantitative and qualitative analyses of the bacterial microbiota of tilapia (Oreochromis niloticus) cultured in earthen ponds in the Philippines. World J. Microbiol. Biotechnol. 2015, 31, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Soliman, N.F.; Yacout, D.M. Aquaculture in Egypt: Status, constraints and potentials. Aquac. Int. 2016, 24, 1201–1227. [Google Scholar] [CrossRef]
- Toranzo, A.E.; Magariños, B.; Romalde, J.L. A review of the main bacterial fish diseases in mariculture systems. Aquaculture 2005, 246, 37–61. [Google Scholar] [CrossRef]
- El-Sayed, M.; Algammal, A.; Abouel-Atta, M.; Mabrok, M.; Emam, A. Pathogenicity, genetic typing, and antibiotic sensitivity of Vibrio alginolyticus isolated from Oreochromis niloticus and Tilapia zillii. Rev. Med. Vet. 2019, 170, 80–86. [Google Scholar]
- Pridgeon, J.W.; Klesius, P.H. Molecular identification and virulence of three Aeromonas hydrophila isolates cultured from infected channel catfish during a disease outbreak in west Alabama (USA) in 2009. Dis. Aquat. Org. 2011, 94, 249–253. [Google Scholar] [CrossRef] [Green Version]
- Chandran, M.; Aruna, B.; Logambal, S.; Michael, R.D. Immunisation of Indian major carps against Aeromonas hydrophila by intraperitoneal injection. Fish Shellfish Immunol. 2002, 13, 1–9. [Google Scholar] [CrossRef]
- Llobrera, A.T.; Gacutan, R.Q. Aeromonas hydrophila associated with ulcerative disease epizootic in Laguna de Bay, Philippines. Aquaculture 1987, 67, 273–278. [Google Scholar] [CrossRef]
- Yambot, A. Isolation of Aeromonas hydrophila from Oreochromis niloticus during fish disease outbreaks in the Philippines. Asian Fish. Sci. 1998, 10, 347–354. [Google Scholar]
- Esteve, C.; Biosca, E.G.; Amaro, C. Virulence of Aeromonas hydrophila and some other bacteria isolated from European eels Anguilla anguilla reared in fresh water. Dis. Aquat. Org. 1993, 16, 15–20. [Google Scholar] [CrossRef]
- Maji, S.; Mali, P.; Joardar, S. Immunoreactive antigens of the outer membrane protein of Aeromonas hydrophila, isolated from goldfish, Carassius auratus (Linn.). Fish Shellfish Immunol. 2006, 20, 462–473. [Google Scholar] [CrossRef] [PubMed]
- Dias, M.K.; Sampaio, L.S.; Proietti-Junior, A.A.; Yoshioka, E.T.; Rodrigues, D.P.; Rodriguez, A.F.; Ribeiro, R.A.; Faria, F.S.; Ozório, R.O.; Tavares-Dias, M. Lethal dose and clinical signs of Aeromonas hydrophila in Arapaima gigas (Arapaimidae), the giant fish from Amazon. Vet. Microbiol. 2016, 188, 12–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamy, B.; Laurent, F.; Verdier, I.; Decousser, J.-W.; Lecaillon, E.; Marchandin, H.; Roger, F.; Tigaud, S.; de Montclos, H.; Kodjo, A. Accuracy of 6 commercial systems for identifying clinical Aeromonas isolates. Diagn. Microbiol. Infect. Dis. 2010, 67, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Soler, L.; Marco, F.; Vila, J.; Chacón, M.; Guarro, J.; Figueras, M. Evaluation of two miniaturized systems, MicroScan W/A and BBL Crystal E/NF, for identification of clinical isolates of Aeromonas spp. J. Clin. Microbiol. 2003, 41, 5732–5734. [Google Scholar] [CrossRef] [Green Version]
- Awan, M.B.; Ahmed, M.M.; Bari, A.; Saad, A.M. Biochemical characterization of the Aeromonas species isolated from food and environment. Pak. J. Physiol. 2005, 1, 1–2. [Google Scholar]
- Jagoda, S.D.S.; Wijewardana, T.; Arulkanthan, A.; Igarashi, Y.; Tan, E.; Kinoshita, S.; Watabe, S.; Asakawa, S. Characterization and antimicrobial susceptibility of motile Aeromonads isolated from freshwater ornamental fish showing signs of septicaemia. Dis. Aquat. Org. 2014, 109, 127–137. [Google Scholar] [CrossRef] [Green Version]
- Beaz-Hidalgo, R.; Alperi, A.; Buján, N.; Romalde, J.L.; Figueras, M.J. Comparison of phenotypical and genetic identification of Aeromonas strains isolated from diseased fish. Syst. Appl. Microbiol. 2010, 33, 149–153. [Google Scholar] [CrossRef]
- Mabrok, M.; Chokmangmeepisarn, P.; LaFrentz, B.; Kayansamruaj, P.; Dong, H.; Rodkhum, C. Development of a species-specific polymerase chain reaction for highly sensitive detection of Flavobacterium columnare targeting chondroitin AC lyase gene. Aquaculture 2019, 521, 734597. [Google Scholar] [CrossRef]
- Enany, M.E.; Algammal, A.M.; Shagar, G.I.; Hanora, A.M.; Elfeil, W.K.; Elshaffy, N.M. Molecular typing and evaluation of Sidr honey inhibitory effect on virulence genes of MRSA strains isolated from catfish in Egypt. Pak. J. Pharm. Sci. 2018, 31, 1865–1870. [Google Scholar]
- Eid, H.M.; Algammal, A.M.; Elfeil, W.K.; Youssef, F.M.; Harb, S.M.; Abd-Allah, E.M. Prevalence, molecular typing, and antimicrobial resistance of bacterial pathogens isolated from ducks. Vet. World 2019, 12, 677. [Google Scholar] [CrossRef]
- Khor, W.C.; Puah, S.M.; Tan, J.A.M.A.; Puthucheary, S.; Chua, K.H. Phenotypic and genetic diversity of Aeromonas species isolated from fresh water lakes in Malaysia. PLoS ONE 2015, 10, e0145933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakroun, C.; Urdaci, M.C.; Faure, D.; Grimont, F.; Bernardet, J.-F. Random amplified polymorphic DNA analysis provides rapid differentiation among isolates of the fish pathogen Flavobacterium psychrophilum and among Flavobacterium species. Dis. Aquat. Org. 1997, 31, 187–196. [Google Scholar] [CrossRef]
- Fadel, A.; Mabrok, M.; Aly, S. Epizootics of Pseudomonas anguilliseptica among cultured seabream (Sparus aurata) populations: Control and treatment strategies. Microb. Pathog. 2018, 121, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Austin, B.; Austin, D. Characteristics of the pathogens: Gram-negative bacteria. Bact. Fish Pathog. Dis. Farmed Wild Fish 2007, 81–150. [Google Scholar] [CrossRef]
- Noga, E.J. Fish Disease: Diagnosis and Treatment; John Wiley & Sons: Hoboken, NJ, USA, 2010. [Google Scholar]
- Shotts, E.B.; Rimler, R. Medium for the isolation of Aeromonas hydrophila. Appl. Environ. Microbiol. 1973, 26, 550–553. [Google Scholar] [CrossRef] [Green Version]
- Quinn, P.J.; Markey, B.K.; Leonard, F.C.; Hartigan, P.; Fanning, S.; Fitzpatrick, E. Veterinary Microbiology and Microbial Disease; John Wiley & Sons: Hoboken, NJ, USA, 2011. [Google Scholar]
- Yanez, M.; Catalán, V.; Apraiz, D.; Figueras, M.; Martinez-Murcia, A. Phylogenetic analysis of members of the genus Aeromonas based on gyrB gene sequences. Int. J. Syst. Evol. Microbiol. 2003, 53, 875–883. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Ni, X.; Liu, Y.; Lu, C. Detection of three virulence genes alt, ahp and aerA in Aeromonas hydrophila and their relationship with actual virulence to zebrafish. J. Appl. Microbiol. 2011, 110, 823–830. [Google Scholar] [CrossRef]
- Alderman, D.; Smith, P. Development of draft protocols of standard reference methods for antimicrobial agent susceptibility testing of bacteria associated with fish diseases. Aquaculture 2001, 196, 211–243. [Google Scholar] [CrossRef]
- NCCLS. Performance Standards for Antimicrobial Susceptibility Testing. In 15th informational supplement M100-S15; National Committee for Clinical Laboratory Standards Wayne: Wayne, PA, USA, 2005. [Google Scholar]
- El-Bahar, H.M.; Ali, N.G.; Aboyadak, I.M.; Khalil, S.A.E.S.; Ibrahim, M.S. Virulence genes contributing to Aeromonas hydrophila pathogenicity in Oreochromis niloticus. Int. Microbiol. 2019, 22, 479–490. [Google Scholar] [CrossRef]
- Tschirner, M.; Kloas, W. Increasing the sustainability of aquaculture systems: Insects as alternative protein source for fish diets. GAIA Ecol. Perspect. Sci. Soc. 2017, 26, 332–340. [Google Scholar] [CrossRef]
- De Silva, S.S.; Nguyen, T.T.; Ingram, B. Fish reproduction in relation to aquaculture. Fish Reprod. 2008, 535–575. Available online: dro.deakin.edu.au/eserv/DU:30021451/desilva-fishproduction-2008.pdf (accessed on 1 March 2020).
- Yardimci, B.; AYDIN, Y. Pathological findings of experimental Aeromonas hydrophila infection in Nile tilapia (Oreochromis niloticus). Ank. Üniv. Vet. Fak. Derg. 2011, 58, 47–54. [Google Scholar]
- Zhang, D.; Xu, D.-H.; Shoemaker, C. Experimental induction of motile Aeromonas septicemia in channel catfish (Ictalurus punctatus) by waterborne challenge with virulent Aeromonas hydrophila. Aquac. Rep. 2016, 3, 18–23. [Google Scholar] [CrossRef] [Green Version]
- Ibrahem, M.; Mostafa, M.; Arab, R.; Rezk, M. Prevalence of Aeromonas hydrophila infection in wild and cultured tilapia nilotica (O. niloticus) in Egypt. In Proceedings of the 8th International Symposium on Tilapia in Aquaculture, Cairo, Egypt, 12–14 October 2008; pp. 1257–1271. [Google Scholar]
- Aberoum, A.; Jooyandeh, H. A review on occurrence and characterization of the Aeromonas species from marine fishes. World J. Fish Mar. Sci. 2010, 2, 519–523. [Google Scholar]
- HäUnninen, M.L. Phenotypic characteristics of the three hybridization groups of Aeromonas hydrophila complex isolated from different sources. J. Appl. Bacteriol. 1994, 76, 455–462. [Google Scholar] [CrossRef]
- Abbott, S.L.; Cheung, W.K.; Janda, J.M. The genus Aeromonas: Biochemical characteristics, atypical reactions, and phenotypic identification schemes. J. Clin. Microbiol. 2003, 41, 2348–2357. [Google Scholar] [CrossRef] [Green Version]
- Simmons, J.H.; Gibson, S. Bacterial and mycotic diseases of nonhuman primates. In Nonhuman Primates in Biomedical Research; Elsevier Inc.: Amsterdam, The Netherlands, 2012; pp. 105–172. [Google Scholar]
- Goñi-Urriza, M.; Arpin, C.; Capdepuy, M.; Dubois, V.; Caumette, P.; Quentin, C. Type II topoisomerase quinolone resistance-determining regions of Aeromonas caviae, A. hydrophila, and A. sobria complexes and mutations associated with quinolone resistance. Antimicrob. Agents Chemother. 2002, 46, 350–359. [Google Scholar] [CrossRef] [Green Version]
- Kusdarwati, R.; Dinda, N.; Nurjanah, I. Antimicrobial resistance prevalence of Aeromonas hydrophila isolates from motile Aeromonas septicemia disease. In Proceedings of the IOP Conference Series: Earth and Environmental Science, Batu City, Indonesia, 7–9 November 2017; p. 012076. [Google Scholar]
- Stratev, D.; Odeyemi, O.A. Antimicrobial resistance of Aeromonas hydrophila isolated from different food sources: A mini-review. J. Infect. Public Health 2016, 9, 535–544. [Google Scholar] [CrossRef] [Green Version]
- Alcaide, E.; Blasco, M.-D.; Esteve, C. Mechanisms of quinolone resistance in Aeromonas species isolated from humans, water and eels. Res. Microbiol. 2010, 161, 40–45. [Google Scholar] [CrossRef]
- Tacão, M.; Moura, A.; Alves, A.; Henriques, I.; Saavedra, M.J.; Correia, A. Evaluation of 16S rDNA-and gyrB-DGGE for typing members of the genus Aeromonas. FEMS Microbiol. Lett. 2005, 246, 11–18. [Google Scholar] [CrossRef] [Green Version]
- González-Serrano, C.; Santos, J.; García-López, M.; Otero, A. Virulence markers in Aeromonas hydrophila and Aeromonas veronii biovar sobria isolates from freshwater fish and from a diarrhoea case. J. Appl. Microbiol. 2002, 93, 414–419. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.; Somvanshi, P.; Rathore, G.; Kapoor, D.; Mishra, B. Gene cloning, expression and homology modeling of hemolysin gene from Aeromonas hydrophila. Protein Expr. Purif. 2009, 65, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Aguilera-Arreola, M.G.; Hernández-Rodríguez, C.; Zúñiga, G.; Figueras, M.J.; Castro-Escarpulli, G. Aeromonas hydrophila clinical and environmental ecotypes as revealed by genetic diversity and virulence genes. FEMS Microbiol. Lett. 2005, 242, 231–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talon, D.; Dupont, M.; Lesne, J.; Thouverez, M.; Michel-Briand, Y. Pulsed-field gel electrophoresis as an epidemiological tool for clonal identification of Aeromonas hydrophila. J. Appl. Bacteriol. 1996, 80, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Nieto, T.; Santos, Y.; Rodriguez, L.; Ellis, A. An extracellular acetylcholinesterase produced by Aeromonas hydrophila is a major lethal toxin for fish. Microb. Pathog. 1991, 11, 101–110. [Google Scholar] [CrossRef]
- Rey, A.; Verjan, N.; Ferguson, H.; Iregui, C. Pathogenesis of Aeromonas hydrophila strain KJ99 infection and its extracellular products in two species of fish. Vet. Rec. 2009, 164, 493–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Primers Sequences 5′–3′ | Amplicon Size (bp) | PCR Conditions | Reference | ||||
|---|---|---|---|---|---|---|---|---|
| No of Cycles | Denaturation | Annealing | Extension | Final Extension | ||||
| gyrB | F: TCCGGCGGTCTGCACGGCGT R: TTGTCCGGGTTGTACTCGTC | 1100 | 35 | 94 °C for 30 s | 55 °C for 30 s | 72 °C for 1 min | 72 °C for 10 min | [28] |
| aerA | F: AACCGAACTCTCCAT R: CGCCTTGTCCTTGTA | 301 | 30 | 54 °C for 30 s | [29] | |||
| alt | F: TGACCCAGTCCTGG R: GGTGATCGATCACC | 442 | 30 | 60 °C for 30 s | ||||
| ahp | F: ATTGGATCCCTGCCTA R: GCTAAGCTTGCATCCG | 911 | 30 | 56 °C for 30 s | ||||
| Enzymes | Acronym | Sequence | Site Length | Frequency | Cut Positions |
|---|---|---|---|---|---|
| Digestive enzyme I | Eco31I | GGTCTC | 6 | 1 | 690 |
| Digestive enzyme II | EcorII | CCWGG | 5 | 5 | 104, 221, 506, 983, 1055 |
| Test | A. hydrophila |
|---|---|
| Gram stain | Gram-negative |
| Motility | Motile |
| Cell shape | Straight rods |
| Oxidase | + |
| Catalase | + |
| O/F | fermentative |
| H2S | − |
| Indole | + |
| Methyl red test | − |
| Voges Proskauer reaction | + |
| Vibrostatic agent or Novobiocin disc | R |
| Casein, Gelatin, Starch | + |
| Urea | − |
| Esculin hydrolysis Growth on | + |
| 0–3% (w/v) sodium chloride | + |
| 5% (w/v) sodium chloride | − |
| Production of acid and/or gas | |
| Glucose | + |
| Fructose | + |
| Salicin | + |
| Arabinose | V |
| Sorbitol | − |
| Sucrose | + |
| Mannitol | + |
| Inositol | + |
| Raffinose | − |
| Maltose | + |
| Trehalose | + |
| Farm No | Locality | No of Fish | Prevalence of. A. hydrophila Complex among Naturally Infected Fish | Chi-Square Value and Significance | ||
|---|---|---|---|---|---|---|
| No | % | |||||
| 1 | Abbassa | 25 | 4 | 16 | 8.93 NS P > 0.05 | 1.7107 NS P > 0.05 |
| 2 | 25 | 6 | 24 | |||
| 3 | 25 | 3 | 12 | |||
| 4 | 25 | 7 | 28 | |||
| 5 | 25 | - | - | |||
| 125 | 20 | |||||
| 6 | El-Hossania | 25 | 4 | 16 | 11.6758 * P < 0.05 | |
| 7 | 25 | 3 | 12 | |||
| 8 | 25 | 6 | 24 | |||
| 9 | 25 | - | - | |||
| 10 | 25 | - | - | |||
| Total | 250 | 33 | 13.2 | |||
| Fish Species | Total No. of Isolates | Prevalence of Isolates in Different Organs | |||||
|---|---|---|---|---|---|---|---|
| Liver | Kidney | Spleen | |||||
| No | % | No | % | No | % | ||
| Oreochromis niloticus | 61 | 33 | 54.1 | 16 | 26.3 | 12 | 19.6 |
| Chi-square value and significance | 18.3443 * P < 0.0001 | ||||||
| Antimicrobial Agents | Concentration µg | Susceptibility Patterns | |||||
|---|---|---|---|---|---|---|---|
| Sensitive | Intermediate | Resistant | |||||
| No | % | No | % | No | % | ||
| Chloramphenicol | 30 | 55 | 90.1 | 6 | 9.9 | 0 | 0 |
| Trimethoprim/Sulphamethoxazol | 1.25/23.75 | 51 | 83.6 | 4 | 6.5 | 6 | 9.9 |
| Amoxicillin | 25 | 0 | 0 | 0 | 0 | 61 | 100 |
| Streptomycin | 10 | 11 | 18 | 13 | 21.3 | 37 | 60.7 |
| Erythromycin | 15 | 0 | 0 | 37 | 60.7 | 24 | 39.3 |
| Tetracycline | 30 | 0 | 0 | 6 | 9.9 | 55 | 90.1 |
| Ciprofloxacin | 5 | 61 | 100 | 0 | 0 | 0 | 0 |
| Nalidixic acid | 30 | 0 | 0 | 6 | 9.9 | 55 | 90.1 |
| Chi-square value and significance | 389.65 * P < 0.0001 | 132.17 * P < 0.0001 | 306.51 * P < 0.0001 | ||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Algammal, A.M.; Mohamed, M.F.; Tawfiek, B.A.; Hozzein, W.N.; El Kazzaz, W.M.; Mabrok, M. Molecular Typing, Antibiogram and PCR-RFLP Based Detection of Aeromonas hydrophila Complex Isolated from Oreochromis niloticus. Pathogens 2020, 9, 238. https://doi.org/10.3390/pathogens9030238
Algammal AM, Mohamed MF, Tawfiek BA, Hozzein WN, El Kazzaz WM, Mabrok M. Molecular Typing, Antibiogram and PCR-RFLP Based Detection of Aeromonas hydrophila Complex Isolated from Oreochromis niloticus. Pathogens. 2020; 9(3):238. https://doi.org/10.3390/pathogens9030238
Chicago/Turabian StyleAlgammal, Abdelazeem M., Mohamed Fathi Mohamed, Basma A. Tawfiek, Wael N. Hozzein, Waleed M. El Kazzaz, and Mahmoud Mabrok. 2020. "Molecular Typing, Antibiogram and PCR-RFLP Based Detection of Aeromonas hydrophila Complex Isolated from Oreochromis niloticus" Pathogens 9, no. 3: 238. https://doi.org/10.3390/pathogens9030238