
Prevention of Stress-Induced Depressive-like Behavior by Saffron Extract Is Associated with Modulation of Kynurenine Pathway and Monoamine Neurotransmission

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Treatment
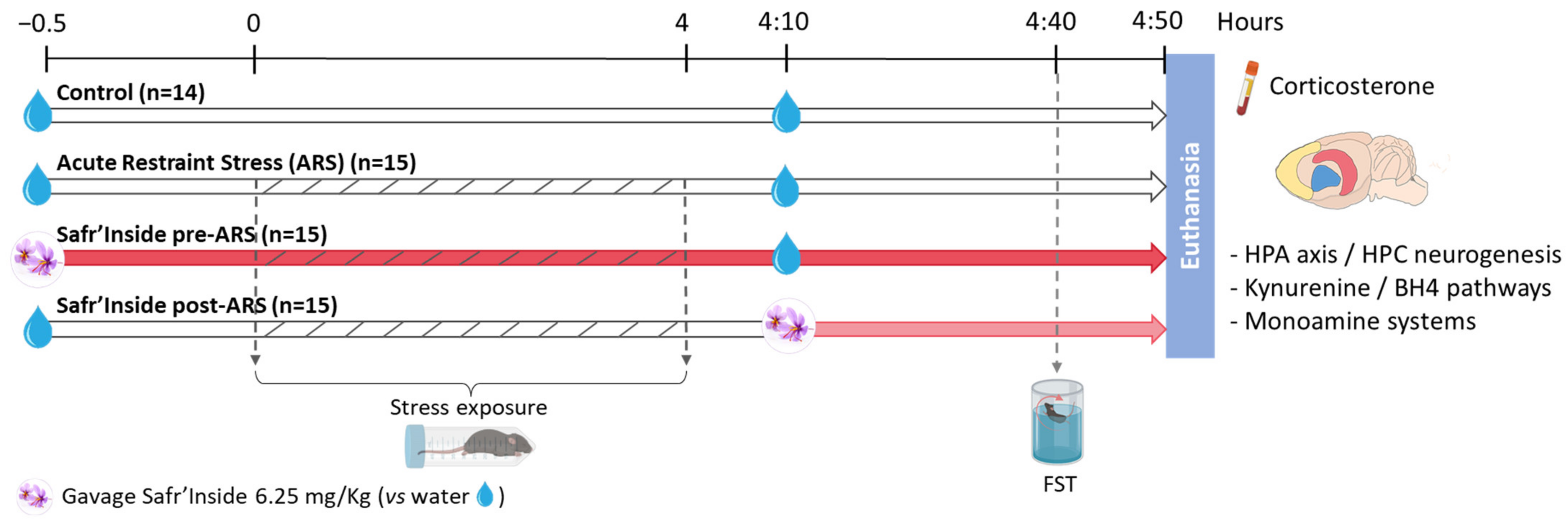
2.2. Experimental Design
2.3. Behavioral Measures
2.4. Tissues Sampling
2.5. Enzyme Immunoassays (EIA)
2.6. High Performance Liquid Chromatography Coupled to Electrochemical Detection (HPLC-EC)
2.7. Real-Time Quantitative PCR (RT-qPCR)
2.8. Western Blotting (WB)
2.9. Statistical Analyses
3. Results
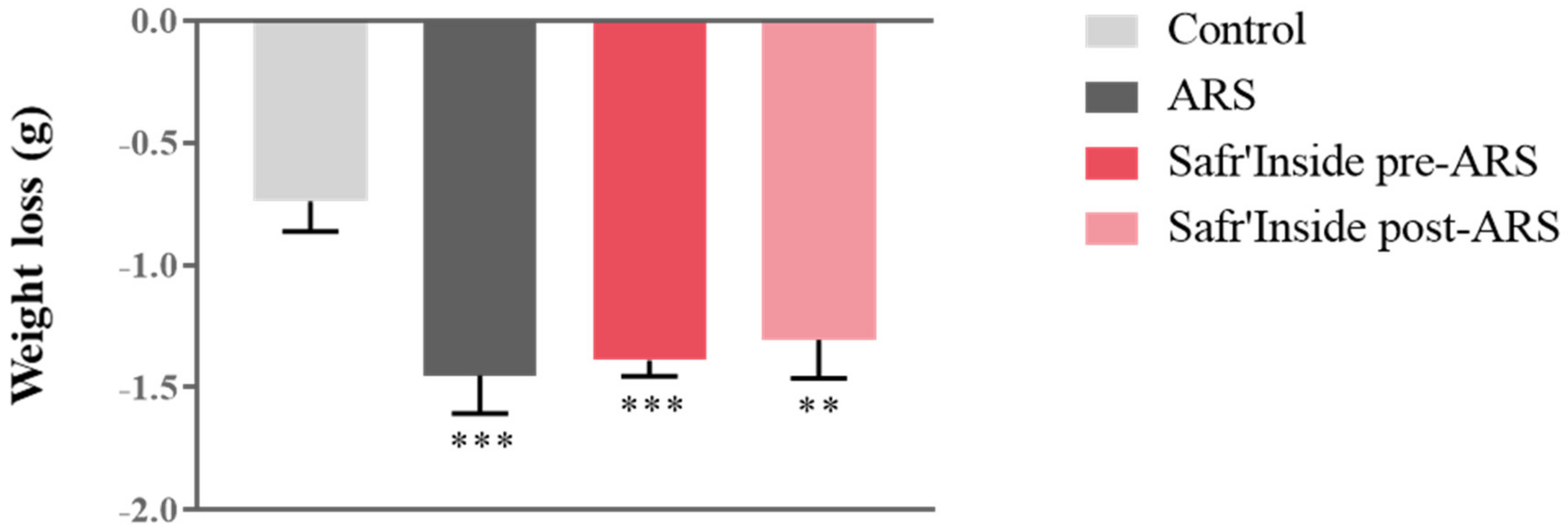
3.1. Safr’Inside Administration Does Not Modify ARS-Induced Weight Loss
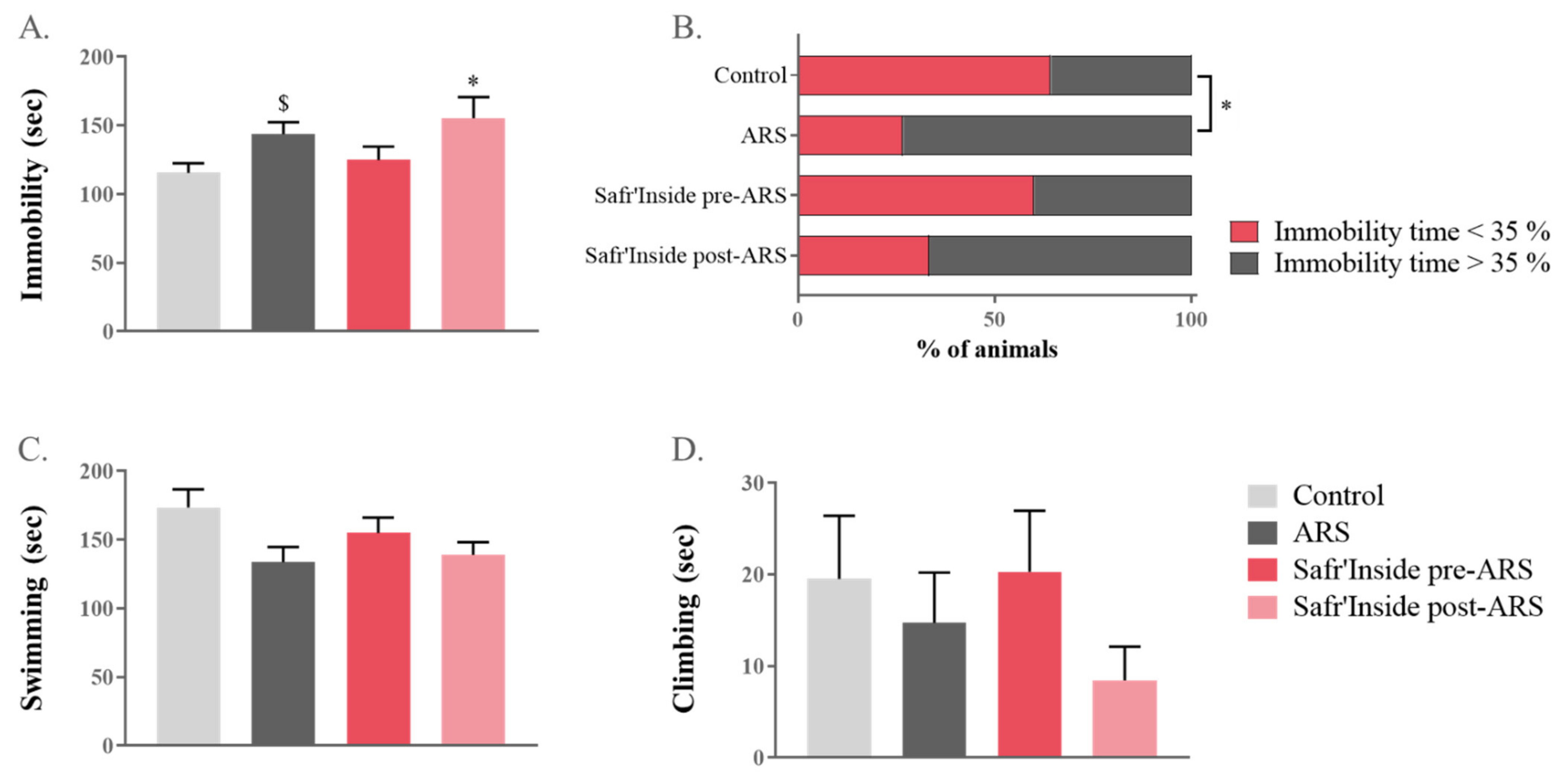
3.2. Safr’Inside Administration Reduces ARS-Induced Depressive-like Behavior Only When Given before Stress Exposure
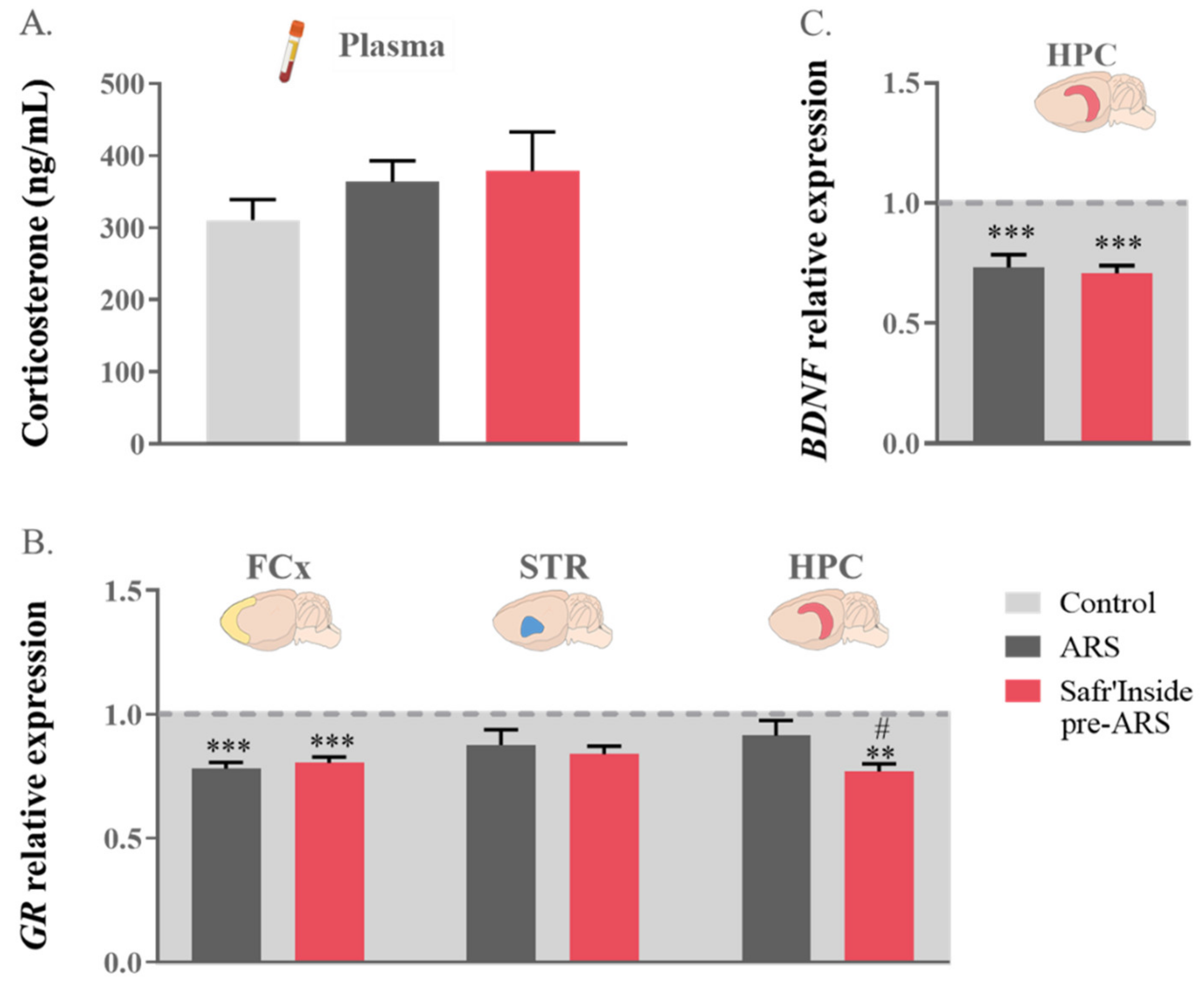
3.3. Safr’Inside Administration Only Slightly Changes HPA Axis Function and Related Neurobiological Targets
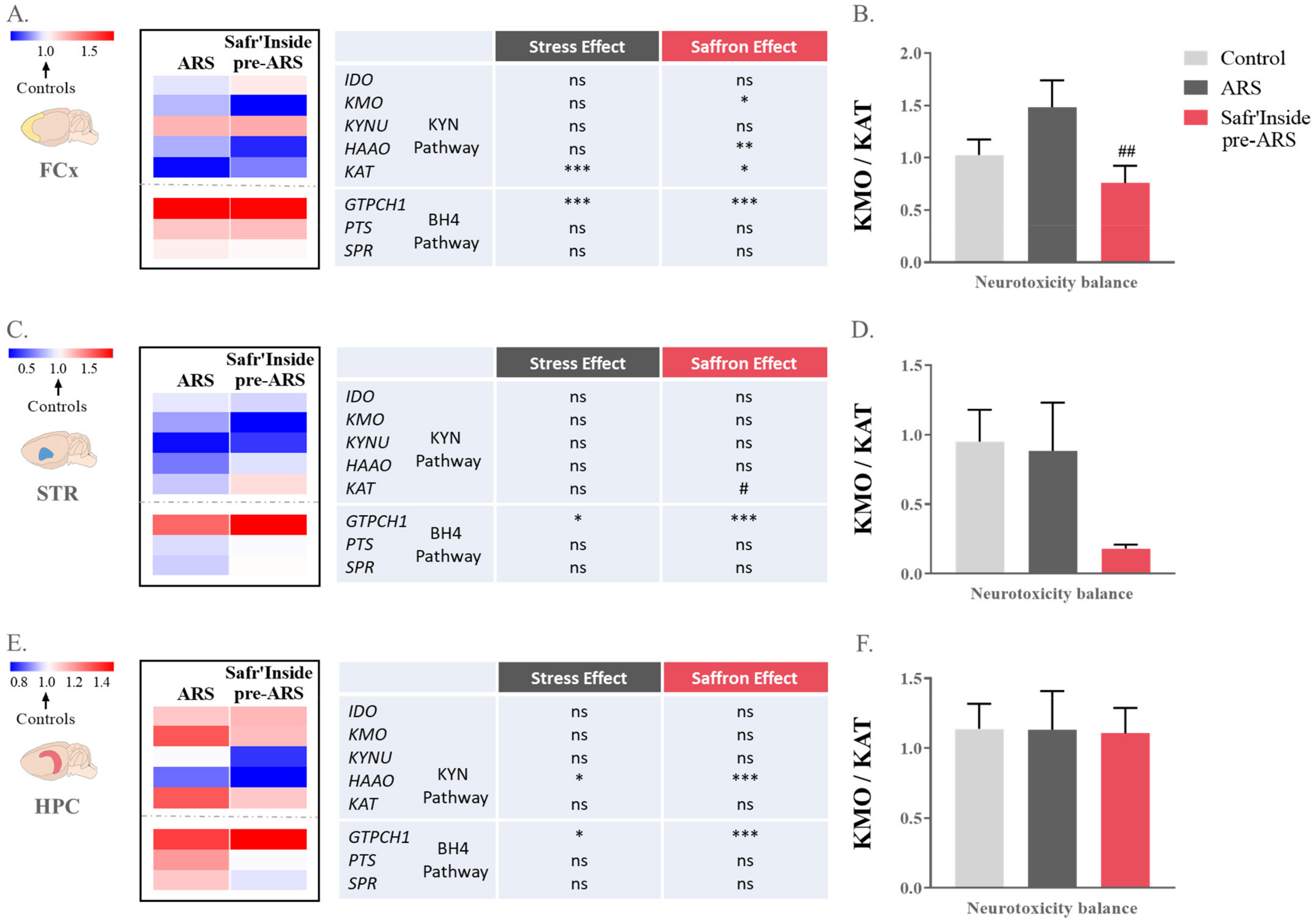
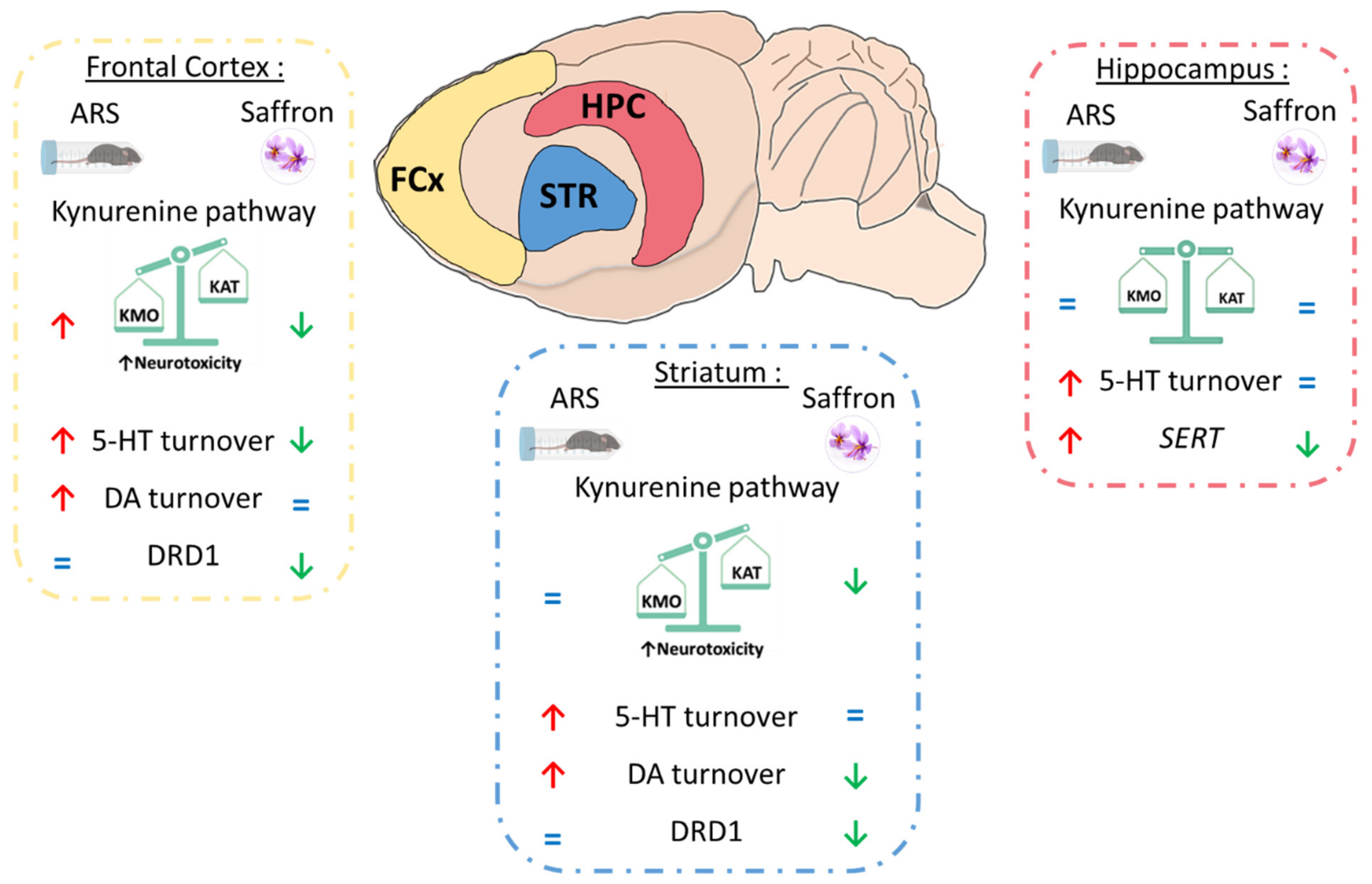
3.4. Safr’Inside Administration Positively Regulates the Kynurenine Pathway
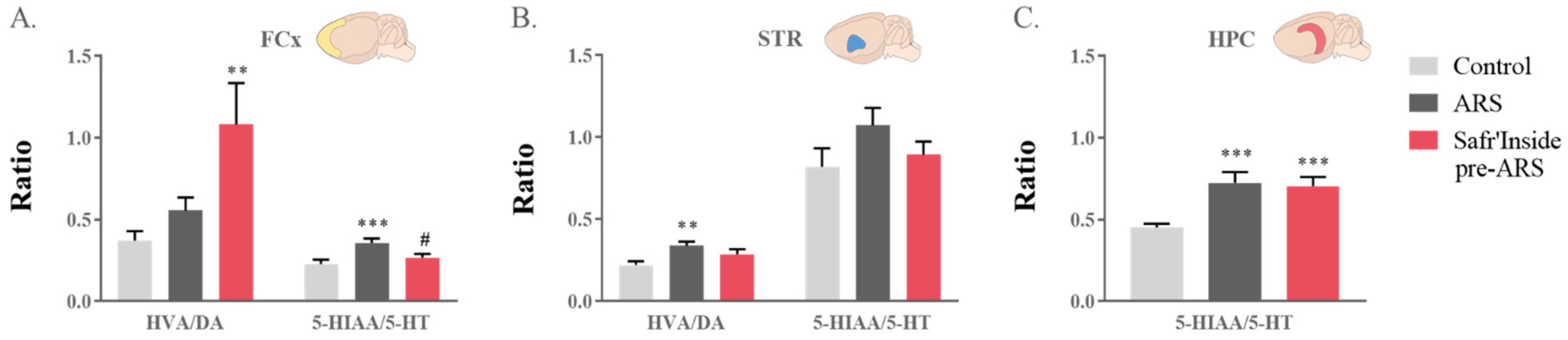
3.5. Safr’Inside Administration Partially Prevents ARS-Induced Alterations of Neurotransmission
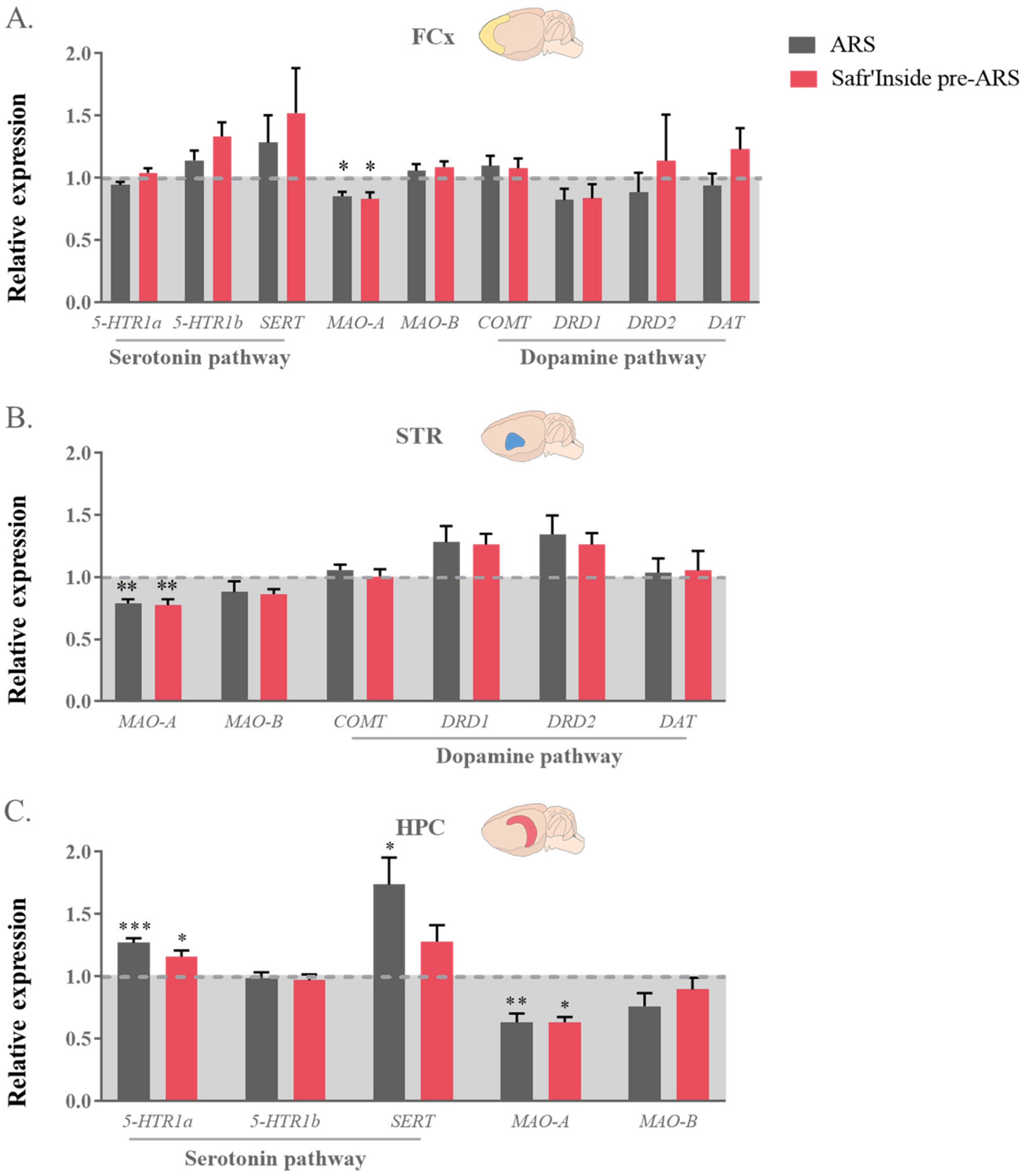
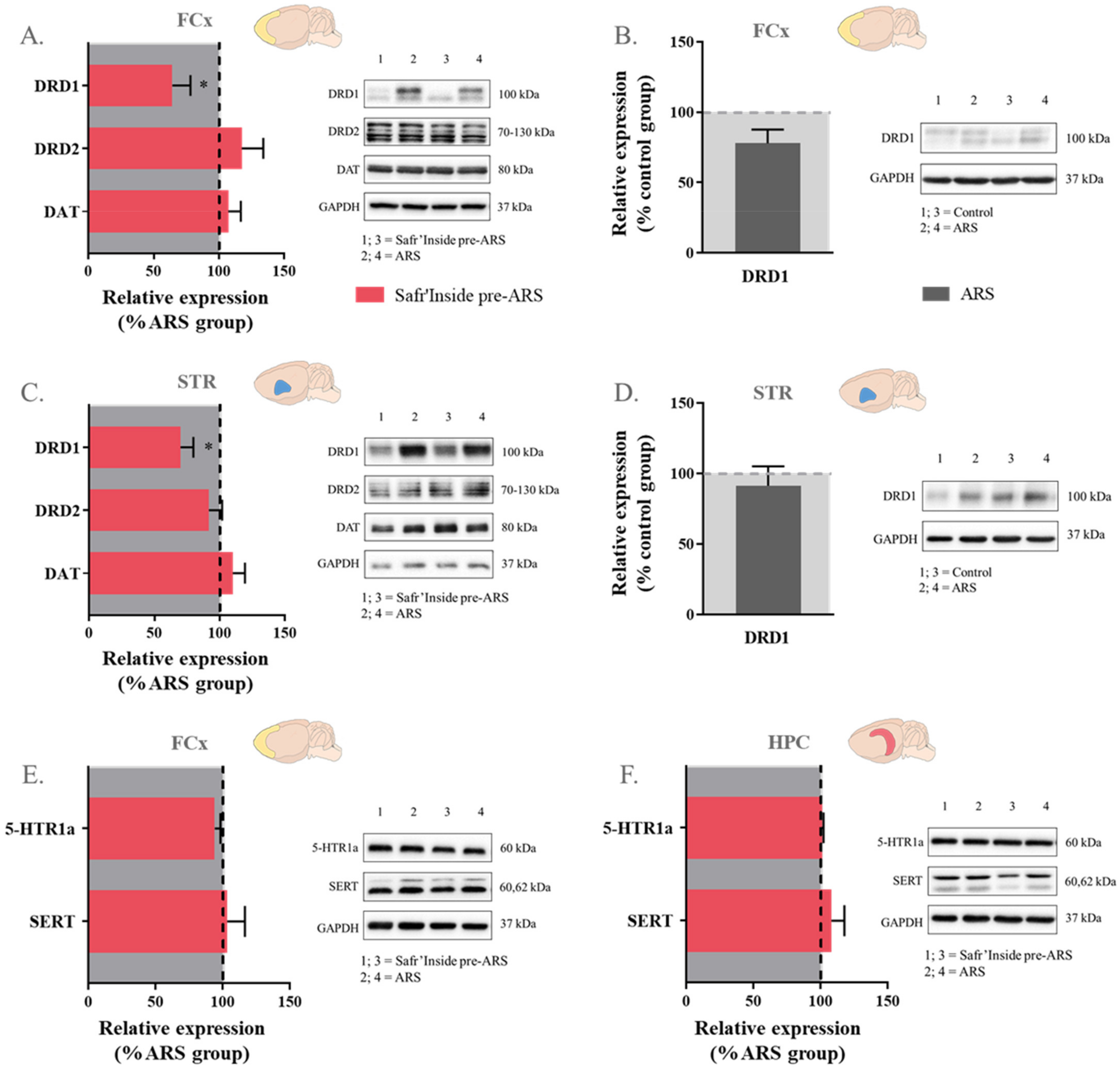
3.6. Safr’Inside Administration Modulates Key Elements of Monoaminergic Systems
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Depression and Other Common Mental Disorders. Available online: http://www.who.int/mental_health/management/depression/prevalence_global_health_estimates/en/ (accessed on 7 January 2021).
- Beurel, E.; Toups, M.; Nemeroff, C.B. The Bidirectional Relationship of Depression and Inflammation: Double Trouble. Neuron 2020, 107, 234–256. [Google Scholar] [CrossRef]
- Capuron, L.; Lasselin, J.; Castanon, N. Role of Adiposity-Driven Inflammation in Depressive Morbidity. Neuropsychopharmacology 2017, 42, 115–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, D.L.; Charney, D.S.; Lewis, L.; Golden, R.N.; Gorman, J.M.; Krishnan, K.R.R.; Nemeroff, C.B.; Bremner, J.D.; Carney, R.M.; Coyne, J.C.; et al. Mood Disorders in the Medically Ill: Scientific Review and Recommendations. Biol. Psychiatry 2005, 58, 175–189. [Google Scholar] [CrossRef] [PubMed]
- Plotsky, P.M.; Owens, M.J.; Nemeroff, C.B. Psychoneuroendocrinology of Depression. Hypothalamic-Pituitary-Adrenal Axis. Psychiatr. Clin. N. Am. 1998, 21, 293–307. [Google Scholar] [CrossRef]
- Kendler, K.S.; Karkowski, L.M.; Prescott, C.A. Causal Relationship between Stressful Life Events and the Onset of Major Depression. Am. J. Psychiatry 1999, 156, 837–841. [Google Scholar] [CrossRef] [PubMed]
- Gilman, S.E.; Trinh, N.-H.; Smoller, J.W.; Fava, M.; Murphy, J.M.; Breslau, J. Psychosocial Stressors and the Prognosis of Major Depression: A Test of Axis IV. Psychol. Med. 2013, 43, 303–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rush, A.J.; Trivedi, M.H.; Wisniewski, S.R.; Nierenberg, A.A.; Stewart, J.W.; Warden, D.; Niederehe, G.; Thase, M.E.; Lavori, P.W.; Lebowitz, B.D.; et al. Acute and Longer-Term Outcomes in Depressed Outpatients Requiring One or Several Treatment Steps: A STAR*D Report. Am. J. Psychiatry 2006, 163, 1905–1917. [Google Scholar] [CrossRef]
- Hollon, S.D.; Thase, M.E.; Markowitz, J.C. Treatment and Prevention of Depression. Psychol. Sci. Public Interest 2002, 3, 39–77. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, J.P.R.; Medvedev, I.O.; Caron, M.G. The 5-HT Deficiency Theory of Depression: Perspectives from a Naturalistic 5-HT Deficiency Model, the Tryptophan Hydroxylase 2Arg439His Knockin Mouse. Philos. Trans. R. Soc. B 2012, 367, 2444–2459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeon, S.W.; Kim, Y.-K. Molecular Neurobiology and Promising New Treatment in Depression. Int. J. Mol. Sci. 2016, 17, 381. [Google Scholar] [CrossRef] [Green Version]
- Dean, J.; Keshavan, M. The Neurobiology of Depression: An Integrated View. Asian J. Psychiatry 2017, 27, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Morris, M.C.; Compas, B.E.; Garber, J. Relations among Posttraumatic Stress Disorder, Comorbid Major Depression, and HPA Function: A Systematic Review and Meta-Analysis. Clin. Psychol. Rev. 2012, 32, 301–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pariante, C.M.; Lightman, S.L. The HPA Axis in Major Depression: Classical Theories and New Developments. Trends Neurosci. 2008, 31, 464–468. [Google Scholar] [CrossRef]
- Zunszain, P.A.; Anacker, C.; Cattaneo, A.; Carvalho, L.A.; Pariante, C.M. Glucocorticoids, Cytokines and Brain Abnormalities in Depression. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2011, 35, 722–729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Sa, C.; Duman, R.S. Antidepressants and Neuroplasticity. Bipolar Disord. 2002, 4, 183–194. [Google Scholar] [CrossRef]
- Levchuk, L.A.; Meeder, E.M.G.; Roschina, O.V.; Loonen, A.J.M.; Boiko, A.S.; Michalitskaya, E.V.; Epimakhova, E.V.; Losenkov, I.S.; Simutkin, G.G.; Bokhan, N.A.; et al. Exploring Brain Derived Neurotrophic Factor and Cell Adhesion Molecules as Biomarkers for the Transdiagnostic Symptom Anhedonia in Alcohol Use Disorder and Comorbid Depression. Front. Psychiatry 2020, 11, 296. [Google Scholar] [CrossRef] [Green Version]
- Duman, R.S. Neuronal Damage and Protection in the Pathophysiology and Treatment of Psychiatric Illness: Stress and Depression. Dialogues Clin. Neurosci. 2009, 11, 239–255. [Google Scholar]
- Raison, C.L.; Capuron, L.; Miller, A.H. Cytokines Sing the Blues: Inflammation and the Pathogenesis of Depression. Trends Immunol. 2006, 27, 24–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capuron, L.; Castanon, N. Role of Inflammation in the Development of Neuropsychiatric Symptom Domains: Evidence and Mechanisms. Curr. Top. Behav. Neurosci. 2017, 31, 31–44. [Google Scholar] [CrossRef]
- Dantzer, R.; O’Connor, J.C.; Freund, G.G.; Johnson, R.W.; Kelley, K.W. From Inflammation to Sickness and Depression: When the Immune System Subjugates the Brain. Nat. Rev. Neurosci. 2008, 9, 46–56. [Google Scholar] [CrossRef] [Green Version]
- Vancassel, S.; Capuron, L.; Castanon, N. Brain Kynurenine and BH4 Pathways: Relevance to the Pathophysiology and Treatment of Inflammation-Driven Depressive Symptoms. Front. Neurosci. 2018, 12, 499. [Google Scholar] [CrossRef]
- Fanet, H.; Ducrocq, F.; Tournissac, M.; Oummadi, A.; Lo, A.; Bourrassa, P.; De Smedt-Peyrusse, V.; Azzougen, B.; Capuron, L.; Layé, S.; et al. Tetrahydrobiopterin Administration Facilitates Amphetamine-Induced Dopamine Release and Motivation in Mice. Behav. Brain Res. 2020, 379, 112348. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-S.; Shen, C.-Y.; Jiang, J.-G. Antidepressant Active Ingredients from Herbs and Nutraceuticals Used in TCM: Pharmacological Mechanisms and Prospects for Drug Discovery. Pharm. Res 2019, 150, 104520. [Google Scholar] [CrossRef] [PubMed]
- Manosso, L.M.; Moretti, M.; Rodrigues, A.L.S. Nutritional Strategies for Dealing with Depression. Food Funct. 2013, 4, 1776. [Google Scholar] [CrossRef] [PubMed]
- Shahmansouri, N.; Farokhnia, M.; Abbasi, S.-H.; Kassaian, S.E.; Noorbala Tafti, A.-A.; Gougol, A.; Yekehtaz, H.; Forghani, S.; Mahmoodian, M.; Saroukhani, S.; et al. A Randomized, Double-Blind, Clinical Trial Comparing the Efficacy and Safety of Crocus sativus L. with Fluoxetine for Improving Mild to Moderate Depression in Post Percutaneous Coronary Intervention Patients. J. Affect. Disord. 2014, 155, 216–222. [Google Scholar] [CrossRef]
- Mazidi, M.; Shemshian, M.; Mousavi, S.H.; Norouzy, A.; Kermani, T.; Moghiman, T.; Sadeghi, A.; Mokhber, N.; Ghayour-Mobarhan, M.; Ferns, G.A.A. A Double-Blind, Randomized and Placebo-Controlled Trial of Saffron (Crocus sativus L.) in the Treatment of Anxiety and Depression. J. Complementary Integr. Med. 2016, 13, 195–199. [Google Scholar] [CrossRef]
- Bathaie, S.Z.; Hoshyar, R.; Miri, H.; Sadeghizadeh, M. Anticancer Effects of Crocetin in Both Human Adenocarcinoma Gastric Cancer Cells and Rat Model of Gastric Cancer. Biochem. Cell Biol. 2013, 91, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Razavi, B.M.; Hosseinzadeh, H. Saffron: A Promising Natural Medicine in the Treatment of Metabolic Syndrome: Saffron as a Natural Medicine in Metabolic Syndrome. J. Sci. Food Agric. 2017, 97, 1679–1685. [Google Scholar] [CrossRef] [PubMed]
- Hosseinzadeh, H.; Noraei, N.B. Anxiolytic and Hypnotic Effect of Crocus sativus Aqueous Extract and Its Constituents, Crocin and Safranal, in Mice. Phytother. Res. 2009, 23, 768–774. [Google Scholar] [CrossRef] [PubMed]
- Noorbala, A.A.; Akhondzadeh, S.; Tahmacebi-Pour, N.; Jamshidi, A.H. Hydro-Alcoholic Extract of Crocus sativus L. versus Fluoxetine in the Treatment of Mild to Moderate Depression: A Double-Blind, Randomized Pilot Trial. J. Ethnopharmacol. 2005, 97, 281–284. [Google Scholar] [CrossRef]
- Talaei, A.; Hassanpour Moghadam, M.; Sajadi Tabassi, S.A.; Mohajeri, S.A. Crocin, the Main Active Saffron Constituent, as an Adjunctive Treatment in Major Depressive Disorder: A Randomized, Double-Blind, Placebo-Controlled, Pilot Clinical Trial. J. Affect. Disord. 2015, 174, 51–56. [Google Scholar] [CrossRef]
- Hausenblas, H.A.; Saha, D.; Dubyak, P.J.; Anton, S.D. Saffron (Crocus sativus L.) and Major Depressive Disorder: A Meta-Analysis of Randomized Clinical Trials. J. Integr. Med. 2013, 11, 377–383. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Han, T.; Zhu, Y.; Zheng, C.-J.; Ming, Q.-L.; Rahman, K.; Qin, L.-P. Antidepressant Properties of Bioactive Fractions from the Extract of Crocus sativus L. J. Nat. Med. 2010, 64, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Hosseinzadeh, H.; Karimi, G.; Niapoor, M. Antidepressant Effect of Crocus sativus L. Stigma Extracts and their Constituents, Crocin and Safranal, in Mice. Acta Hortic. 2004, 435–445. [Google Scholar] [CrossRef]
- Amin, B.; Nakhsaz, A.; Hosseinzadeh, H. Evaluation of the Antidepressant-like Effects of Acute and Sub-Acute Administration of Crocin and Crocetin in Mice. Avicenna J. Phytomed. 2015, 5, 458–468. [Google Scholar] [PubMed]
- Pitsikas, N.; Boultadakis, A.; Georgiadou, G.; Tarantilis, P.A.; Sakellaridis, N. Effects of the Active Constituents of Crocus sativus L., Crocins, in an Animal Model of Anxiety. Phytomedicine 2008, 15, 1135–1139. [Google Scholar] [CrossRef] [PubMed]
- Monchaux De Oliveira, C.; Pourtau, L.; Vancassel, S.; Pouchieu, C.; Capuron, L.; Gaudout, D.; Castanon, N. Saffron Extract-Induced Improvement of Depressive-Like Behavior in Mice Is Associated with Modulation of Monoaminergic Neurotransmission. Nutrients 2021, 13, 904. [Google Scholar] [CrossRef]
- Ettehadi, H.; Mojabi, S.N.; Ranjbaran, M.; Shams, J.; Sahraei, H.; Hedayati, M.; Asefi, F. Aqueous Extract of Saffron (Crocus sativus) Increases Brain Dopamine and Glutamate Concentrations in Rats. J. Behav. Brain Sci. 2013, 3, 315–319. [Google Scholar] [CrossRef] [Green Version]
- Boskabady, M.H.; Farkhondeh, T. Antiinflammatory, Antioxidant, and Immunomodulatory Effects of Crocus sativus L. and Its Main Constituents. Phytother. Res. 2016, 30, 1072–1094. [Google Scholar] [CrossRef]
- Ghadrdoost, B.; Vafaei, A.A.; Rashidy-Pour, A.; Hajisoltani, R.; Bandegi, A.R.; Motamedi, F.; Haghighi, S.; Sameni, H.R.; Pahlvan, S. Protective Effects of Saffron Extract and Its Active Constituent Crocin against Oxidative Stress and Spatial Learning and Memory Deficits Induced by Chronic Stress in Rats. Eur. J. Pharmacol. 2011, 667, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Hooshmandi, Z.; Rohani, A.H.; Eidi, A.; Fatahi, Z.; Golmanesh, L.; Sahraei, H. Reduction of Metabolic and Behavioral Signs of Acute Stress in Male Wistar Rats by Saffron Water Extract and Its Constituent Safranal. Pharm. Biol. 2011, 49, 947–954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cryan, J.F.; Valentino, R.J.; Lucki, I. Assessing Substrates Underlying the Behavioral Effects of Antidepressants Using the Modified Rat Forced Swimming Test. Neurosci. Biobehav. Rev. 2005, 29, 547–569. [Google Scholar] [CrossRef] [PubMed]
- Halataei, B.-S.; Khosravi, M.; Arbabian, S.; Sahraei, H.; Golmanesh, L.; Zardooz, H.; Jalili, C.; Ghoshooni, H. Saffron (Crocus sativus) Aqueous Extract and Its Constituent Crocin Reduces Stress-Induced Anorexia in Mice. Phytother. Res. 2011, 25, 1833–1838. [Google Scholar] [CrossRef] [PubMed]
- Domingues, M.; Casaril, A.M.; Birmann, P.T.; Bampi, S.R.; Lourenço, D.d.A.; Vieira, B.M.; Dapper, L.H.; Lenardão, E.J.; Sonego, M.; Collares, T.; et al. Effects of a Selanylimidazopyridine on the Acute Restraint Stress-Induced Depressive- and Anxiety-like Behaviors and Biological Changes in Mice. Behav. Brain Res. 2019, 366, 96–107. [Google Scholar] [CrossRef]
- Casaril, A.M.; Domingues, M.; Bampi, S.R.; de Andrade Lourenço, D.; Padilha, N.B.; Lenardão, E.J.; Sonego, M.; Seixas, F.K.; Collares, T.; Nogueira, C.W.; et al. The Selenium-Containing Compound 3-((4-Chlorophenyl)Selanyl)-1-Methyl-1H-Indole Reverses Depressive-like Behavior Induced by Acute Restraint Stress in Mice: Modulation of Oxido-Nitrosative Stress and Inflammatory Pathway. Psychopharmacology 2019, 236, 2867–2880. [Google Scholar] [CrossRef] [PubMed]
- Sousa, F.S.S.; Birmann, P.T.; Balaguez, R.; Alves, D.; Brüning, C.A.; Savegnago, L. α-(Phenylselanyl) Acetophenone Abolishes Acute Restraint Stress Induced-Comorbid Pain, Depression and Anxiety-Related Behaviors in Mice. Neurochem. Int. 2018, 120, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Turner, P.V.; Brabb, T.; Pekow, C.; Vasbinder, M.A. Administration of Substances to Laboratory Animals: Routes of Administration and Factors to Consider. J. Am. Assoc. Lab. Anim. Sci. 2011, 50, 600–613. [Google Scholar]
- Nestler, E.J.; Hyman, S.E. Animal Models of Neuropsychiatric Disorders. Nat. Neurosci. 2010, 13, 1161–1169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- André, C.; Dinel, A.-L.; Ferreira, G.; Layé, S.; Castanon, N. Diet-Induced Obesity Progressively Alters Cognition, Anxiety-like Behavior and Lipopolysaccharide-Induced Depressive-like Behavior: Focus on Brain Indoleamine 2,3-Dioxygenase Activation. Brain Behav. Immun. 2014, 41, 10–21. [Google Scholar] [CrossRef]
- Hornung, J.-P. The Human Raphe Nuclei and the Serotonergic System. J. Chem. Neuroanat. 2003, 26, 331–343. [Google Scholar] [CrossRef]
- Björklund, A.; Dunnett, S.B. Dopamine Neuron Systems in the Brain: An Update. Trends Neurosci. 2007, 30, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Ducrocq, F.; Walle, R.; Contini, A.; Oummadi, A.; Caraballo, B.; van der Veldt, S.; Boyer, M.-L.; Aby, F.; Tolentino-Cortez, T.; Helbling, J.-C.; et al. Causal Link between N-3 Polyunsaturated Fatty Acid Deficiency and Motivation Deficits. Cell Metab. 2020, 31, 755–772.e7. [Google Scholar] [CrossRef] [PubMed]
- Outlier Calculator. Available online: https://www.graphpad.com/quickcalcs/Grubbs1.cfm (accessed on 13 September 2021).
- O’Connor, J.C.; Lawson, M.A.; André, C.; Moreau, M.; Lestage, J.; Castanon, N.; Kelley, K.W.; Dantzer, R. Lipopolysaccharide-Induced Depressive-like Behavior Is Mediated by Indoleamine 2,3-Dioxygenase Activation in Mice. Mol. Psychiatry 2009, 14, 511–522. [Google Scholar] [CrossRef] [Green Version]
- Fu, X.-Y.; Li, H.-Y.; Jiang, Q.-S.; Cui, T.; Jiang, X.-H.; Zhou, Q.-X.; Qiu, H.-M. Infliximab Ameliorating Depression-like Behavior through Inhibiting the Activation of the IDO–HAAO Pathway Mediated by Tumor Necrosis Factor-α in a Rat Model. NeuroReport 2016, 27, 953–959. [Google Scholar] [CrossRef]
- Gibney, S.M.; Fagan, E.M.; Waldron, A.-M.; O’Byrne, J.; Connor, T.J.; Harkin, A. Inhibition of Stress-Induced Hepatic Tryptophan 2,3-Dioxygenase Exhibits Antidepressant Activity in an Animal Model of Depressive Behaviour. Int. J. Neuropsychopharmacol. 2014, 17, 917–928. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.T.; Choi, J.H.; Chang, J.W.; Kim, S.W.; Hwang, O. Immobilization Stress Causes Increases in Tetrahydrobiopterin, Dopamine, and Neuromelanin and Oxidative Damage in the Nigrostriatal System: BH4 and DA Overproduction and Oxidative Damage. J. Neurochem. 2005, 95, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Livingston, M.G.; Livingston, H.M. Monoamine Oxidase Inhibitors. An Update on Drug Interactions. Drug Saf. 1996, 14, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Feighner, J.P. Mechanism of Action of Antidepressant Medications. J. Clin. Psychiatry 1999, 60 (Suppl. 4), 4–11, discussion 12–13. [Google Scholar] [PubMed]
- Zarate, C.A.; Payne, J.L.; Singh, J.; Quiroz, J.A.; Luckenbaugh, D.A.; Denicoff, K.D.; Charney, D.S.; Manji, H.K. Pramipexole for Bipolar II Depression: A Placebo-Controlled Proof of Concept Study. Biol. Psychiatry 2004, 56, 54–60. [Google Scholar] [CrossRef] [Green Version]
- Thase, M.E. Therapeutic Alternatives for Difficult-to-Treat Depression: A Narrative Review of the State of the Evidence. CNS Spectr. 2004, 9, 808–821. [Google Scholar] [CrossRef]
- Millan, M.J. Dual- and Triple-Acting Agents for Treating Core and Co-Morbid Symptoms of Major Depression: Novel Concepts, New Drugs. Neurotherapeutics 2009, 6, 53–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bang-Andersen, B.; Ruhland, T.; Jørgensen, M.; Smith, G.; Frederiksen, K.; Jensen, K.G.; Zhong, H.; Nielsen, S.M.; Hogg, S.; Mørk, A.; et al. Discovery of 1-[2-(2,4-Dimethylphenylsulfanyl)Phenyl]Piperazine (Lu AA21004): A Novel Multimodal Compound for the Treatment of Major Depressive Disorder. J. Med. Chem. 2011, 54, 3206–3221. [Google Scholar] [CrossRef] [PubMed]
- Dale, E.; Pehrson, A.L.; Jeyarajah, T.; Li, Y.; Leiser, S.C.; Smagin, G.; Olsen, C.K.; Sanchez, C. Effects of Serotonin in the Hippocampus: How SSRIs and Multimodal Antidepressants Might Regulate Pyramidal Cell Function. CNS Spectr. 2016, 21, 143–161. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, N.C.; Gould, G.G.; Koek, W.; Daws, L.C. Ontogeny of SERT Expression and Antidepressant-like Response to Escitalopram in Wild-Type and SERT Mutant Mice. J. Pharmacol. Exp. Ther. 2016, 358, 271–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pesarico, A.P.; Stangherlin, E.C.; Mantovani, A.C.; Zeni, G.; Nogueira, C.W. 7-Fluoro-1,3-Diphenylisoquinoline-1-Amine Abolishes Depressive-like Behavior and Prefrontal Cortical Oxidative Damage Induced by Acute Restraint Stress in Mice. Physiol. Behav. 2015, 149, 294–302. [Google Scholar] [CrossRef]
- Ghalandari-Shamami, M.; Nourizade, S.; Yousefi, B.; Vafaei, A.A.; Pakdel, R.; Rashidy-Pour, A. Beneficial Effects of Physical Activity and Crocin Against Adolescent Stress Induced Anxiety or Depressive-Like Symptoms and Dendritic Morphology Remodeling in Prefrontal Cortex in Adult Male Rats. Neurochem. Res. 2019, 44, 917–929. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Li, Q.; Jiang, S.; Xu, Z.; Jiang, Y.; Liu, L.; Jiang, J.; Tong, Y.; Wang, P. Crocetin Ameliorates Chronic Restraint Stress-Induced Depression-like Behaviors in Mice by Regulating MEK/ERK Pathways and Gut Microbiota. J. Ethnopharmacol. 2021, 268, 113608. [Google Scholar] [CrossRef] [PubMed]
- Ghajar, A.; Neishabouri, S.; Velayati, N.; Jahangard, L.; Matinnia, N.; Haghighi, M.; Ghaleiha, A.; Afarideh, M.; Salimi, S.; Meysamie, A.; et al. Crocus sativus L. versus Citalopram in the Treatment of Major Depressive Disorder with Anxious Distress: A Double-Blind, Controlled Clinical Trial. Pharmacopsychiatry 2017, 50, 152–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orio, L.; Alen, F.; Ballesta, A.; Martin, R.; Gomez de Heras, R. Antianhedonic and Antidepressant Effects of Affron®, a Standardized Saffron (Crocus sativus L.) Extract. Molecules 2020, 25, 3207. [Google Scholar] [CrossRef] [PubMed]
- Jackson, P.A.; Forster, J.; Khan, J.; Pouchieu, C.; Dubreuil, S.; Gaudout, D.; Moras, B.; Pourtau, L.; Joffre, F.; Vaysse, C.; et al. Effects of Saffron Extract Supplementation on Mood, Well-Being, and Response to a Psychosocial Stressor in Healthy Adults: A Randomized, Double-Blind, Parallel Group, Clinical Trial. Front. Nutr. 2021, 7, 606124. [Google Scholar] [CrossRef]
- Ghasemi, T.; Abnous, K.; Vahdati, F.; Mehri, S.; Razavi, B.; Hosseinzadeh, H. Antidepressant Effect of Crocus sativus Aqueous Extract and Its Effect on CREB, BDNF, and VGF Transcript and Protein Levels in Rat Hippocampus. Drug Res. 2014, 65, 337–343. [Google Scholar] [CrossRef]
- USFDA. Guidance for Industry: Estimating the Maximum Safe Starting Dose in Adult Healthy Volunteer; US Food and Drug Administration: Rockville, MD, USA, 2005; Volume 30.
- Duman, R.S.; Aghajanian, G.K.; Sanacora, G.; Krystal, J.H. Synaptic Plasticity and Depression: New Insights from Stress and Rapid-Acting Antidepressants. Nat. Med. 2016, 22, 238–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morsink, M.C.; Steenbergen, P.J.; Vos, J.B.; Karst, H.; Joëls, M.; Kloet, E.R.D.; Datson, N.A. Acute Activation of Hippocampal Glucocorticoid Receptors Results in Different Waves of Gene Expression Throughout Time. J. Neuroendocrinol. 2006, 18, 239–252. [Google Scholar] [CrossRef] [PubMed]
- Vahdati Hassani, F.; Naseri, V.; Razavi, B.M.; Mehri, S.; Abnous, K.; Hosseinzadeh, H. Antidepressant Effects of Crocin and Its Effects on Transcript and Protein Levels of CREB, BDNF, and VGF in Rat Hippocampus. DARU J. Pharm. Sci. 2014, 22, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roustazade, R.; Radahmadi, M.; Yazdani, Y. Therapeutic Effects of Saffron Extract on Different Memory Types, Anxiety, and Hippocampal BDNF and TNF-α Gene Expressions in Sub-Chronically Stressed Rats. Nutr. Neurosci. 2021, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Miller, B.R.; Hen, R. The Current State of the Neurogenic Theory of Depression and Anxiety. Curr. Opin. Neurobiol. 2015, 30, 51–58. [Google Scholar] [CrossRef] [Green Version]
- Savitz, J. The Kynurenine Pathway: A Finger in Every Pie. Mol. Psychiatry 2020, 25, 131–147. [Google Scholar] [CrossRef] [PubMed]
- Parrott, J.M.; Redus, L.; O’Connor, J.C. Kynurenine Metabolic Balance Is Disrupted in the Hippocampus Following Peripheral Lipopolysaccharide Challenge. J. Neuroinflamm. 2016, 13, 124. [Google Scholar] [CrossRef] [Green Version]
- Savitz, J.; Drevets, W.C.; Wurfel, B.E.; Ford, B.N.; Bellgowan, P.S.F.; Victor, T.A.; Bodurka, J.; Teague, T.K.; Dantzer, R. Reduction of Kynurenic Acid to Quinolinic Acid Ratio in Both the Depressed and Remitted Phases of Major Depressive Disorder. Brain Behav. Immun. 2015, 46, 55–59. [Google Scholar] [CrossRef] [Green Version]
- Sadeghnia, H.R.; Kamkar, M.; Assadpour, E.; Taher, M.; Ghorbani, A. Protective Effect of Safranal, a Constituent of Crocus sativus, on Quinolinic Acid-Induced Oxidative Damage in Rat Hippocampus. Iran. J. Basic Med. Sci. 2013, 16, 73. [Google Scholar]
- Georgiadou, G.; Grivas, V.; Tarantilis, P.A.; Pitsikas, N. Crocins, the Active Constituents of Crocus sativus L., Counteracted Ketamine-Induced Behavioural Deficits in Rats. Psychopharmacology 2014, 231, 717–726. [Google Scholar] [CrossRef] [PubMed]
- Verdonk, F.; Petit, A.-C.; Abdel-Ahad, P.; Vinckier, F.; Jouvion, G.; de Maricourt, P.; De Medeiros, G.F.; Danckaert, A.; Van Steenwinckel, J.; Blatzer, M.; et al. Microglial Production of Quinolinic Acid as a Target and a Biomarker of the Antidepressant Effect of Ketamine. Brain Behav. Immun. 2019, 81, 361–373. [Google Scholar] [CrossRef]
- Leonard, B.E. HPA and Immune Axes in Stress: Involvement of the Serotonergic System. Neuroimmunomodulation 2006, 13, 268–276. [Google Scholar] [CrossRef] [PubMed]
- Leonard, B.E. SSRI Differentiation: Pharmacology and Pharmacokinetics. Hum. Psychopharmacol. 1995, 10 (Suppl. S3), S149–S158. [Google Scholar] [CrossRef] [PubMed]
- Couch, Y.; Anthony, D.C.; Dolgov, O.; Revischin, A.; Festoff, B.; Santos, A.I.; Steinbusch, H.W.; Strekalova, T. Microglial Activation, Increased TNF and SERT Expression in the Prefrontal Cortex Define Stress-Altered Behaviour in Mice Susceptible to Anhedonia. Brain Behav. Immun. 2013, 29, 136–146. [Google Scholar] [CrossRef]
- Filipenko, M.L.; Beilina, A.G.; Alekseyenko, O.V.; Dolgov, V.V.; Kudryavtseva, N.N. Repeated Experience of Social Defeats Increases Serotonin Transporter and Monoamine Oxidase A MRNA Levels in Raphe Nuclei of Male Mice. Neurosci. Lett. 2002, 321, 25–28. [Google Scholar] [CrossRef]
- Flügge, G.; Van Kampen, M.; Mijnster, M.J. Perturbations in Brain Monoamine Systems during Stress. Cell Tissue Res. 2004, 315, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Blanchard, D.C.; Cholvanich, P.; Blanchard, R.J.; Clow, D.W.; Hammer, R.P.; Rowlett, J.K.; Bardo, M.T. Serotonin, but Not Dopamine, Metabolites Are Increased in Selected Brain Regions of Subordinate Male Rats in a Colony Environment. Brain Res. 1991, 568, 61–66. [Google Scholar] [CrossRef]
- Beitia, G.; Garmendia, L.; Azpiroz, A.; Vegas, O.; Brain, P.F.; Arregi, A. Time-Dependent Behavioral, Neurochemical, and Immune Consequences of Repeated Experiences of Social Defeat Stress in Male Mice and the Ameliorative Effects of Fluoxetine. Brain Behav. Immun. 2005, 19, 530–539. [Google Scholar] [CrossRef]
- Higuchi, Y.; Soga, T.; Parhar, I.S. Regulatory Pathways of Monoamine Oxidase A during Social Stress. Front. Neurosci. 2017, 11, 604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pusceddu, M.M.; Barboza, M.; Keogh, C.E.; Schneider, M.; Stokes, P.; Sladek, J.A.; Kim, H.J.D.; Torres-Fuentes, C.; Goldfild, L.R.; Gillis, S.E.; et al. Nod-like Receptors Are Critical for Gut-Brain Axis Signalling in Mice. J. Physiol. 2019, 597, 5777–5797. [Google Scholar] [CrossRef] [PubMed]
- Carhart-Harris, R.L.; Nutt, D.J. Serotonin and Brain Function: A Tale of Two Receptors. J. Psychopharmacol. 2017, 31, 1091–1120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willner, P. Dopamine and Depression: A Review of Recent Evidence. I. Empirical Studies. Brain Res. Rev. 1983, 6, 211–224. [Google Scholar] [CrossRef]
- Dailly, E.; Chenu, F.; Renard, C.E.; Bourin, M. Dopamine, Depression and Antidepressants. Fundam. Clin. Pharm. 2004, 18, 601–607. [Google Scholar] [CrossRef] [PubMed]
- Alghasham, A.; Rasheed, N. Stress-Mediated Modulations in Dopaminergic System and Their Subsequent Impact on Behavioral and Oxidative Alterations: An Update. Pharm. Biol. 2014, 52, 368–377. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Powell, D.K.; Wang, H.; Gold, B.T.; Corbly, C.R.; Joseph, J.E. Functional Dissociation in Frontal and Striatal Areas for Processing of Positive and Negative Reward Information. J. Neurosci. 2007, 27, 4587–4597. [Google Scholar] [CrossRef] [PubMed]
- Forbes, E.E.; Hariri, A.R.; Martin, S.L.; Silk, J.S.; Moyles, D.L.; Fisher, P.M.; Brown, S.M.; Ryan, N.D.; Birmaher, B.; Axelson, D.A.; et al. Altered Striatal Activation Predicting Real-World Positive Affect in Adolescent Major Depressive Disorder. Am. J. Psychiatry 2009, 166, 64–73. [Google Scholar] [CrossRef] [Green Version]
- Dunlop, B.W.; Nemeroff, C.B. The Role of Dopamine in the Pathophysiology of Depression. Arch. Gen. Psychiatry 2007, 64, 327. [Google Scholar] [CrossRef]
- Metz, V.G.; Segat, H.J.; Dias, V.T.; Barcelos, R.C.S.; Maurer, L.H.; Stiebe, J.; Emanuelli, T.; Burger, M.E.; Pase, C.S. Omega-3 Decreases D1 and D2 Receptors Expression in the Prefrontal Cortex and Prevents Amphetamine-Induced Conditioned Place Preference in Rats. J. Nutr. Biochem. 2019, 67, 182–189. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| FCx (nmoles/g) | STR (nmoles/g) | HPC (pmoles/g) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Control | ARS | Safr’Inside | Control | ARS | Safr’Inside | Control | ARS | Safr’Inside | |
| [DA] | 4.3 ± 1.1 | 4.1 ± 0.5 | 3.7 ± 1.0 | 49 ± 5.8 | 47.2 ± 3.4 | 42.9 ± 4.1 | 144 ± 33 | 125 ± 18 | 93 ± 15 |
| [DOPAC] | 1.2 ± 0.2 | 1.4 ± 0.2 | 0.95 ± 0.1 | 11.1 ± 1.2 | 9.5 ± 1 | 7.7 ± 0.7 | n.d | n.d | n.d |
| [HVA] | 1.1 ± 0.2 | 1.9 ± 0.2 ** | 1.7 ± 0.2 * | 11 ± 1.3 | 15.8 ± 1.5 * | 11.3 ± 1.1 | n.d | n.d | n.d |
| [5-HT] | 1.2 ± 0.05 | 1.4 ± 0.08 | 1.3 ± 0.7 | 0.75 ± 0.1 | 0.84 ± 0.09 | 0.97 ± 0.1 | 986 ± 121 | 1082 ± 134 | 938 ± 119 |
| [5-HIAA] | 0.27 ± 0.03 | 0.49 ± 0.04 *** | 0.37 ± 0.04 | 0.52 ± 0.06 | 0.88 ± 0.1 ** | 0.83 ± 0.08 ** | 401 ± 38 | 698 ± 55 *** | 593 ± 43 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monchaux De Oliveira, C.; De Smedt-Peyrusse, V.; Morael, J.; Vancassel, S.; Capuron, L.; Gaudout, D.; Pourtau, L.; Castanon, N. Prevention of Stress-Induced Depressive-like Behavior by Saffron Extract Is Associated with Modulation of Kynurenine Pathway and Monoamine Neurotransmission. Pharmaceutics 2021, 13, 2155. https://doi.org/10.3390/pharmaceutics13122155
Monchaux De Oliveira C, De Smedt-Peyrusse V, Morael J, Vancassel S, Capuron L, Gaudout D, Pourtau L, Castanon N. Prevention of Stress-Induced Depressive-like Behavior by Saffron Extract Is Associated with Modulation of Kynurenine Pathway and Monoamine Neurotransmission. Pharmaceutics. 2021; 13(12):2155. https://doi.org/10.3390/pharmaceutics13122155
Chicago/Turabian StyleMonchaux De Oliveira, Camille, Véronique De Smedt-Peyrusse, Jennifer Morael, Sylvie Vancassel, Lucile Capuron, David Gaudout, Line Pourtau, and Nathalie Castanon. 2021. "Prevention of Stress-Induced Depressive-like Behavior by Saffron Extract Is Associated with Modulation of Kynurenine Pathway and Monoamine Neurotransmission" Pharmaceutics 13, no. 12: 2155. https://doi.org/10.3390/pharmaceutics13122155