
Free Phenolic Compounds, Antioxidant Capacity and FT-NIR Survey of Debittered Lupinus mutabilis Seeds

 , , , , and
, , , , and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
2.2.1. Debittering and Milling
2.2.2. Free Phenolic Compounds
2.2.3. Antioxidant Capacity
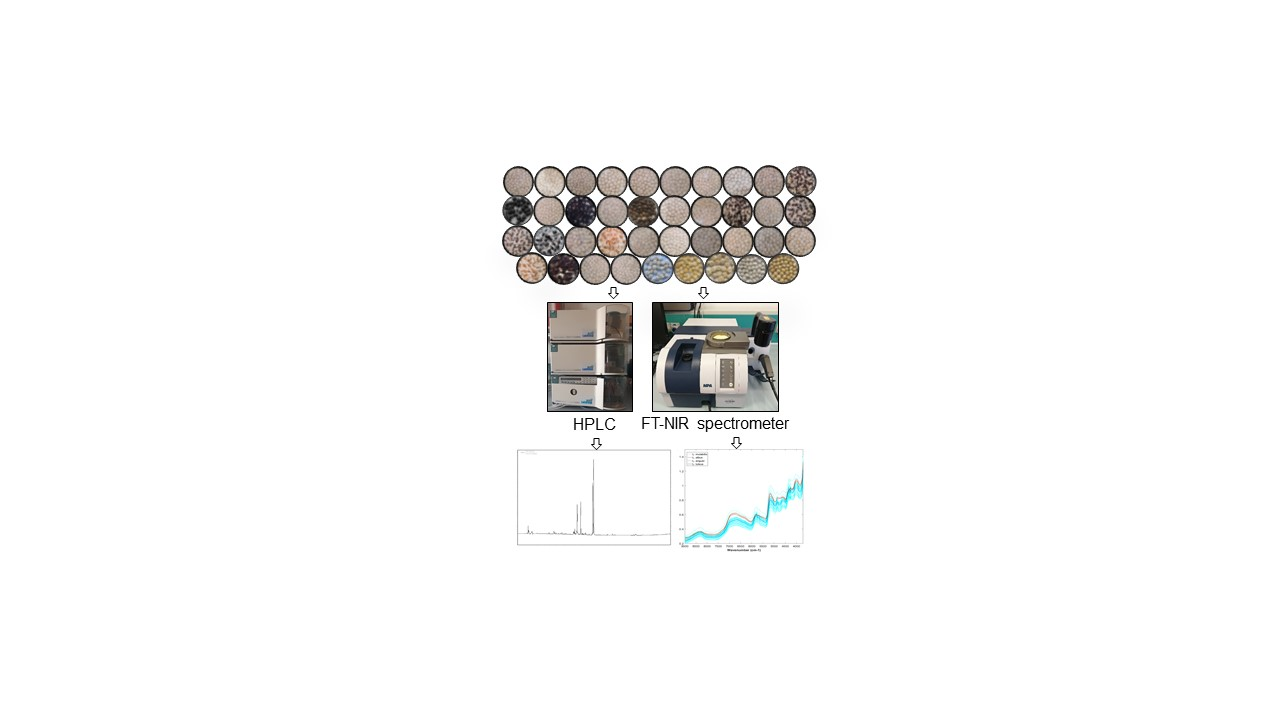
2.2.4. Near Infrared Spectroscopy
2.2.5. Statistical Analysis
3. Results
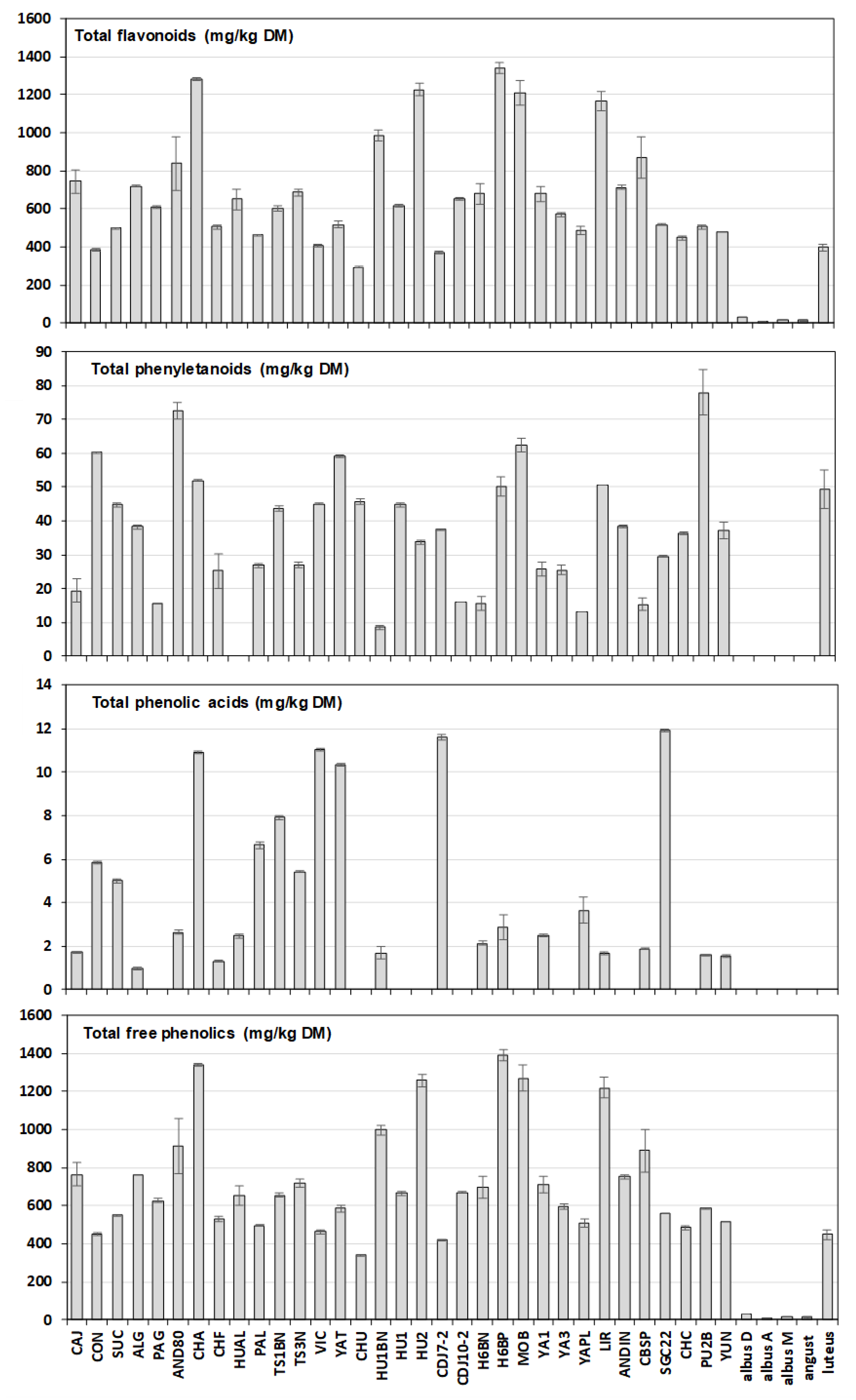
3.1. Phenolics Composition and Content
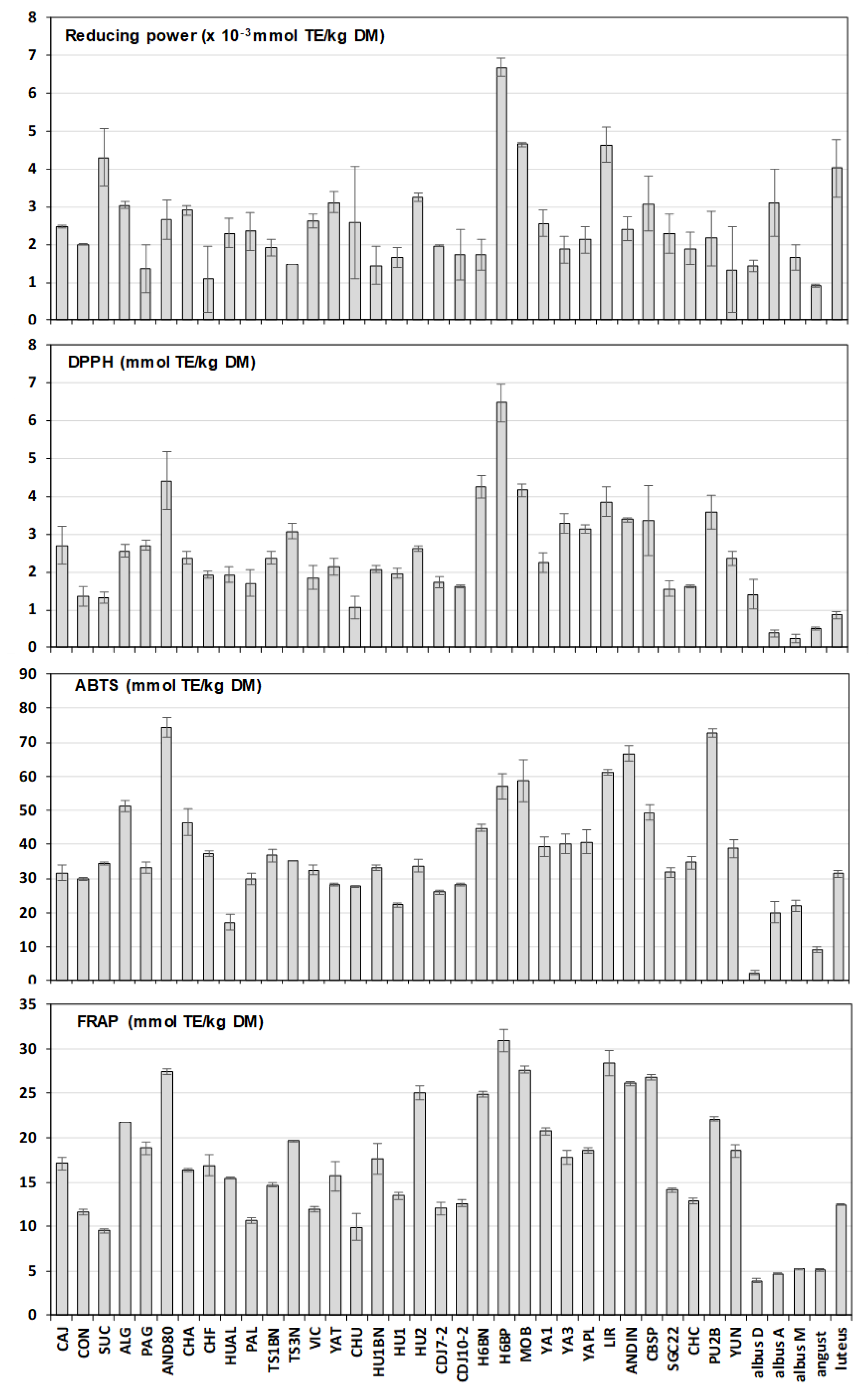
3.2. Antioxidant Capacity and Its Correlation with Phenolics Content
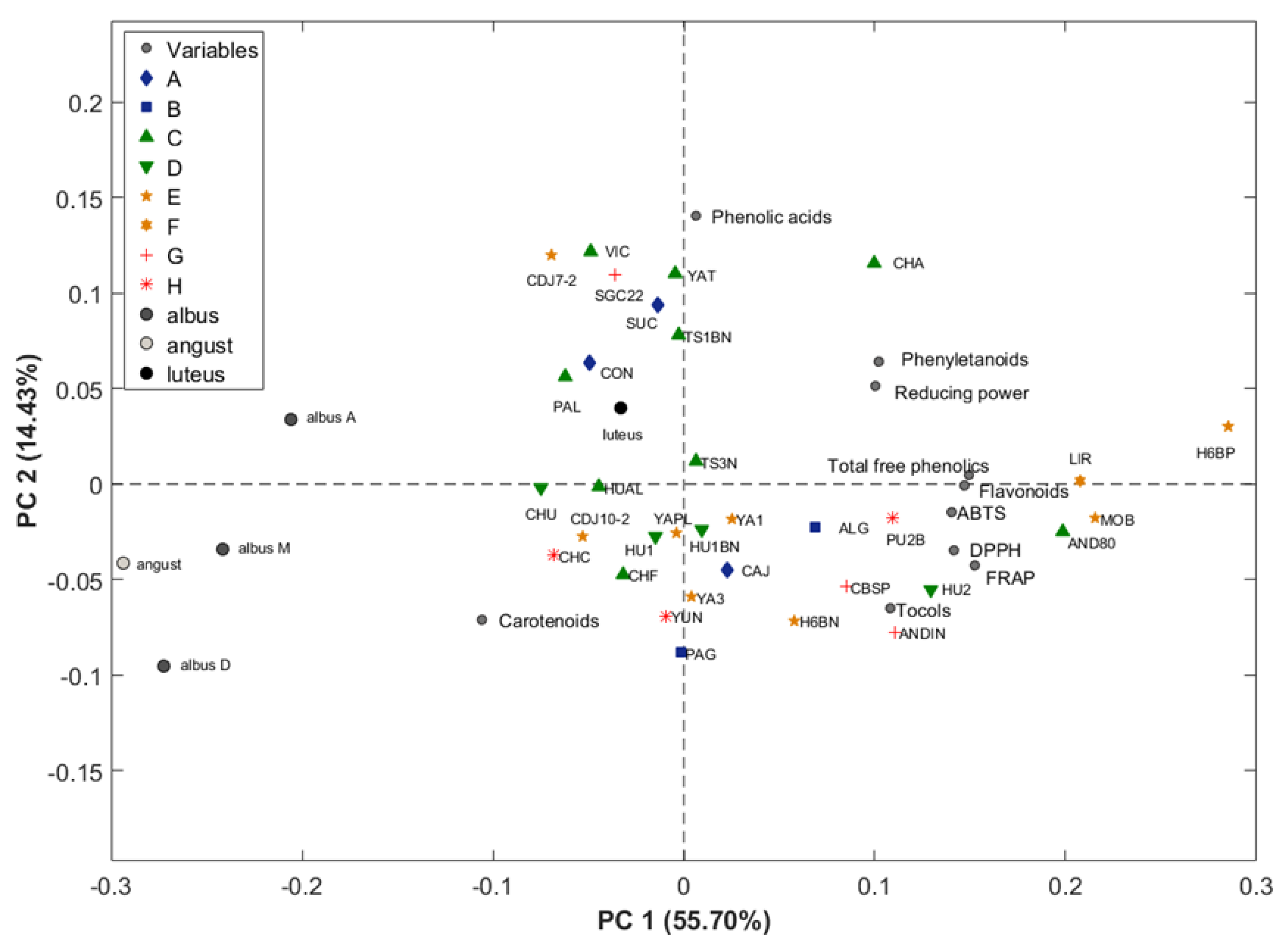
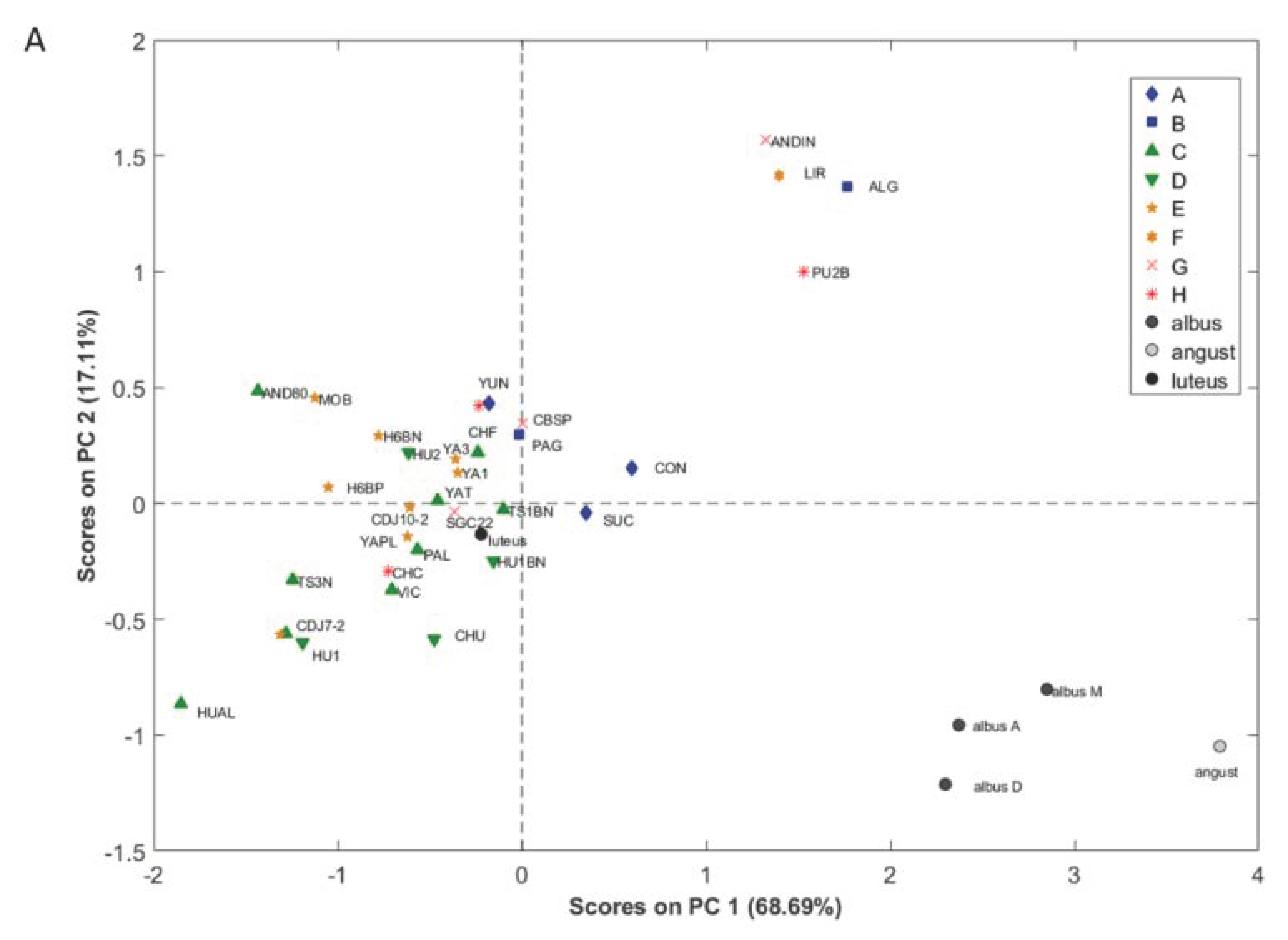
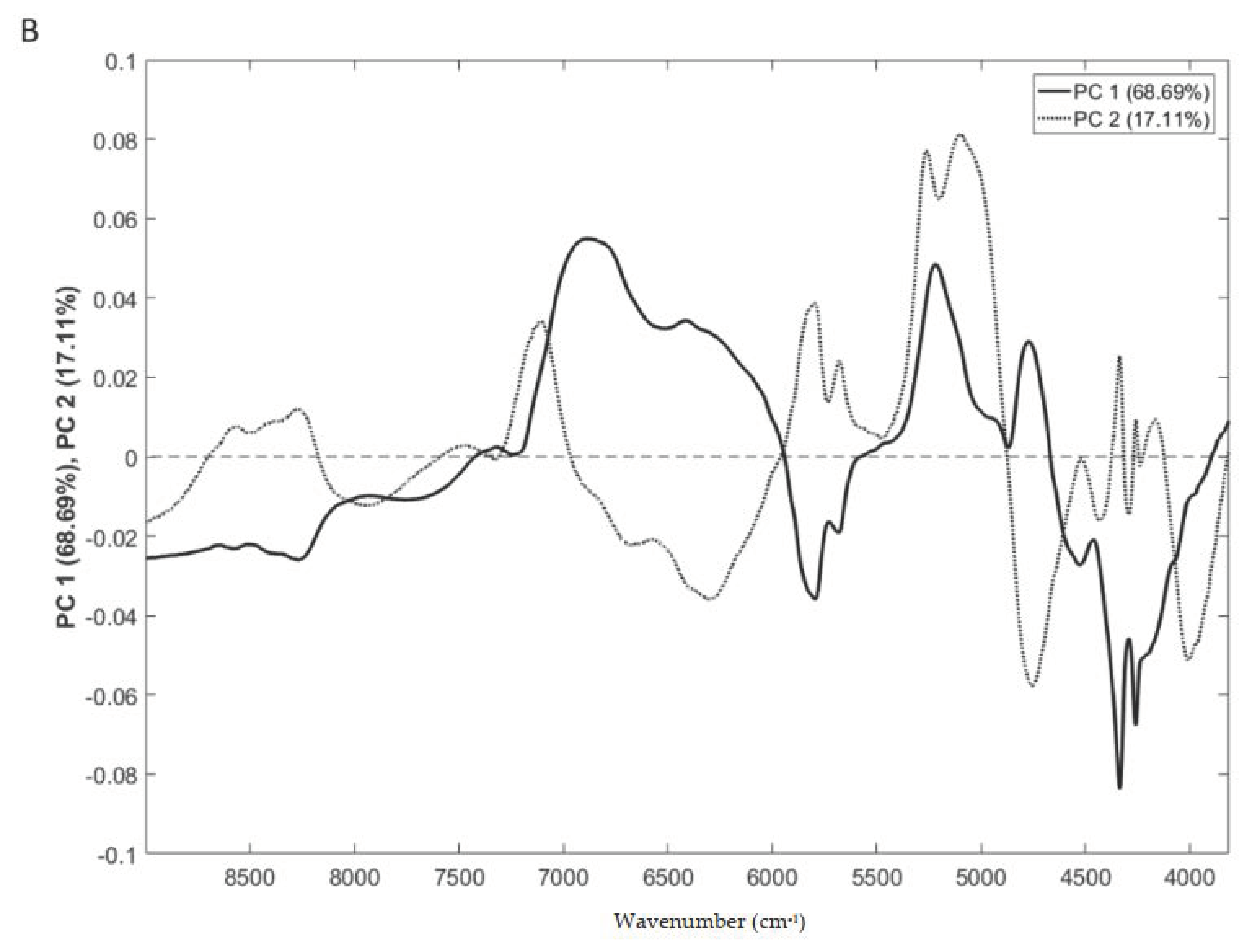
3.3. Geographical Origin
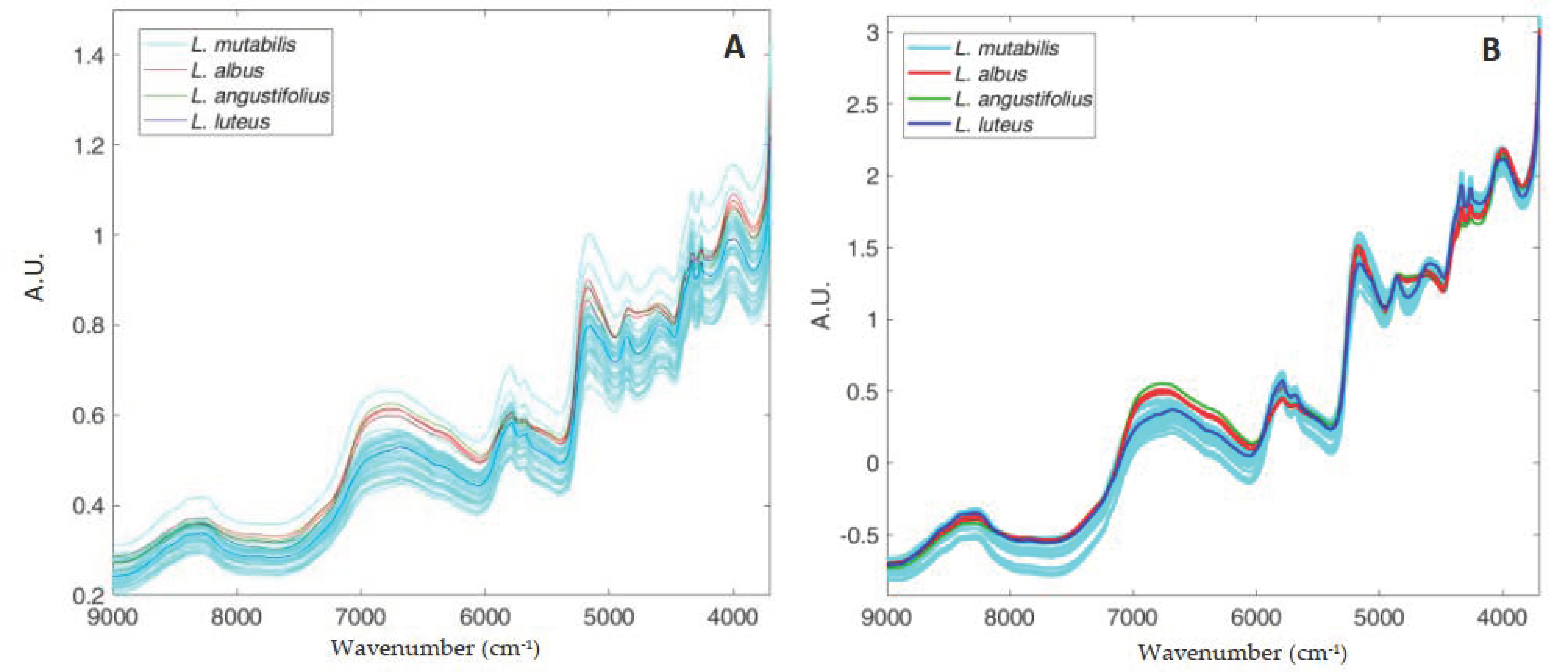
3.4. Near Infrared Spectra and Regression Models
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Musco, N.; Cutrignelli, M.I.; Calabrò, S.; Tudisco, R.; Infascelli, F.; Grazioli, R.; Lo Presti, V.; Gresta, F.; Chiofalo, B. Comparison of nutritional and antinutritional traits among different species (Lupinus albus L., Lupinus luteus L., Lupinus angustifolius L.) and varieties of lupin seeds. J. Anim. Physiol. Anim. Nutr. 2017, 101, 1227–1241. [Google Scholar] [CrossRef]
- Bähr, M.; Fechner, A.; Hasenkopf, K.; Mittermaier, S.; Jahreis, G. Chemical composition of dehulled seeds of selected lupin cultivars in comparison to pea and soya bean. LWT 2014, 59, 587–590. [Google Scholar] [CrossRef]
- Carvajal-Larenas, F.E.; Linnemann, A.R.; Nout, M.J.R.; Koziol, M.; Van Boekel, M.A.J.S. Lupinus mutabilis: Composition, uses, toxicology, and debittering. Crit. Rev. Food Sci. Nutr. 2016, 56, 1454–1487. [Google Scholar] [CrossRef]
- Ruiz-López, M.A.; Barrientos-Ramírez, L.; García-López, P.M.; Valdés-Miramontes, E.H.; Zamora-Natera, J.F.; Rodríguez-Macias, R.; Salcedo-Pérez, E.; Bañuelos-Pineda, J.; Vargas-Radillo, J.J. Nutritional and bioactive compounds in Mexican lupin beans species: A mini-review. Nutrients 2019, 11, 1785. [Google Scholar] [CrossRef] [PubMed]
- Boschin, G.; Arnoldi, A. Legumes are valuable sources of tocopherols. Food Chem. 2011, 127, 1199–1203. [Google Scholar]
- Briceño Berru, L.; Glorio-Paulet, P.; Basso, C.; Scarafoni, A.; Camarena, F.; Hidalgo, A.; Brandolini, A. Chemical composition, tocopherol and carotenoid content of seeds from different Andean lupin (Lupinus mutabilis) ecotypes. Plant Food Hum. Nutr. 2021, 76, 98–104. [Google Scholar] [CrossRef]
- Czubinski, J.; Grygier, A.; Siger, A. Lupinus mutabilis seed composition and its comparison with other lupin species. J. Food Compos. Anal. 2021, 99, 103875. [Google Scholar] [CrossRef]
- Brandolini, A.; Glorio-Paulet, P.; Estivi, L.; Locatelli, N.; Córdova-Ramos, J.S.; Hidalgo, A. Tocopherols, carotenoids and phenolics changes during Andean lupin (Lupinus mutabilis Sweet) seeds processing. J. Food Compos. Anal. 2022, 106, 104335. [Google Scholar] [CrossRef]
- Grela, E.; Kiczorowska, B.; Samolińska, W.; Matras, J.; Kiczorowski, P.; Rybiński, W.; Hanczakowska, E. Chemical composition of leguminous seeds: Part I—Content of basic nutrients, amino acids, phytochemical compounds, and antioxidant activity. Eur. Food Res. Technol. 2017, 243, 385–1395. [Google Scholar] [CrossRef]
- Nimse, S.; Pal, D. Free radicals, natural antioxidants and their reaction mechanisms. RSC Adv. 2015, 5, 27986. [Google Scholar] [CrossRef]
- Córdova-Ramos, J.S.; Glorio-Paulet, P.; Camarena, F.; Brandolini, A.; Hidalgo, A. Andean lupin (Lupinus mutabilis Sweet): Processing effects on chemical composition, heat damage and in vitro protein digestibility. Cereal Chem. 2020, 97, 827–835. [Google Scholar] [CrossRef]
- Carvajal-Larenas, F.E.; Van Boekel, M.J.A.S.; Koziol, M.; Nout, M.J.R.; Linnemann, A.R. Effect of processing on the diffusion of alkaloids and quality of Lupinus mutabilis Sweet. J. Food Process. Preserv. 2014, 38, 1461–1471. [Google Scholar] [CrossRef]
- Jiménez-Martínez, C.; Mora-Escobedo, R.; Cardador Martínez, A.; Muzquiz, M.; Pedrosa, M.M.; Dávila-Ortiz, G. Effect of aqueous, acid, and alkaline thermal treatments on antinutritional factors content and protein quality in Lupinus campestris seed flour. J. Agric. Food Chem. 2010, 58, 1741–1745. [Google Scholar] [CrossRef]
- Galek, R.; Sawicka-Sienkiewicz, E.; Zalewski, D.; Stawiński, S.; Spychała, K. Searching for low alkaloid forms in the Andean lupin (Lupinus mutabilis) collection. Czech J. Genet. Plant Breed. 2017, 53, 55–62. [Google Scholar] [CrossRef]
- Gulisano, A.; Alves, S.; Neves Martins, J.; Trindade, L.M. Genetics and breeding of Lupinus mutabilis: An emerging protein crop. Front. Plant Sci. 2019, 10, 1385. [Google Scholar] [CrossRef]
- Sbihi, H.M.; Nehdi, I.A.; Tan, C.P.; Al-Resayes, S.I. Bitter and sweet lupin (Lupinus albus L.) seeds and seed oils: A comparison study of their compositions and physicochemical properties. Ind. Crops Prod. 2013, 49, 573–579. [Google Scholar] [CrossRef]
- Córdova-Ramos, J.S.; Glorio-Paulet, P.; Hidalgo, A.; Camarena, F. Effect of technological process on antioxidant capacity and total phenolic content of Andean lupine (Lupinus mutabilis Sweet). Sci. Agropecu. 2020, 11, 157–165. [Google Scholar] [CrossRef]
- Villacrés, E.; Quelal, M.B.; Fernández, E.; Garcìa, G.; Cueva, G.; Rosell, C.M. Impact of debittering and fermentation processes on the antinutritional and antioxidant compounds in Lupinus mutabilis Sweet. LWT 2020, 131, 109745. [Google Scholar] [CrossRef]
- Estivi, L.; Buratti, S.; Fusi, D.; Benedetti, S.; Rodríguez, G.; Brandolini, A.; Hidalgo, A. Alkaloid content and taste profile assessed by electronic tongue of Lupinus albus seeds debittered by different methods. J. Food Comp. Anal. 2022, 114, 104810. [Google Scholar] [CrossRef]
- Dueñas, M.; Hernández, T.; Estrella, I.; Fernández, D. Germination as a process to increase the polyphenol content and antioxidant activity of lupin seeds (Lupinus angustifolius L.). Food Chem. 2009, 117, 599–607. [Google Scholar] [CrossRef]
- Zalewski, K.; Nitkiewicz, B.; Stolarski, M.; Amarowicz, R.; Okorskid, A. The influence of exogenous methyl jasmonate on the germination and, content and composition of flavonoids in extracts from seedlings of yellow and narrow-leafed lupine. J. Food Compos. Anal. 2020, 87, 103398. [Google Scholar] [CrossRef]
- Miller, J.C.; Miller, J.N. Statistics for Analytical Chemistry, 2nd ed.; Ellis Horwood Publishers: Chichester, UK, 1998. [Google Scholar]
- Oyaizu, M. Studies on products of browning reaction–antioxidant activities of products of browning reaction prepared from glucoamine. Jpn. J. Nutr. Diet. 1986, 44, 307–315. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Yilmaz, V.A.; Brandolini, A.; Hidalgo, A. Phenolic acids and antioxidant activity of wild, feral and domesticated diploid wheats. J. Cereal Sci. 2015, 64, 168–175. [Google Scholar] [CrossRef]
- Wold, S.; Ruhe, A.; Wold, H.; Dunn, W.J. The collinearity problem in linear regression. The partial least squares (PLS) approach to generalized inverses. J. Sci. Stat. Comp. 1984, 5, 735–743. [Google Scholar] [CrossRef]
- Nørgaard, L.; Saudland, A.; Wagner, J.; Nielsen, J.P.; Munck, L.; Engelsen, S.B. Interval partial least-squares regression (iPLS): A comparative chemometric study with an example from near-infrared spectroscopy. Appl. Spectrosc. 2000, 54, 413–419. [Google Scholar] [CrossRef]
- Gálvez-Ranilla, L.; Genovese, M.I.; Lajolo, F.M. Isoflavones and antioxidant capacity of Peruvian and Brazilian lupin cultivars. J. Food Compos. Anal. 2009, 22, 397–404. [Google Scholar] [CrossRef]
- Czubinski, J.; Wroblewska, K.; Czyzniejewski, M.; Górnaś, P.; Kachlicki, P.; Siger, A. Bioaccessibility of defatted lupin seed phenolic compounds in a standardized static in vitro digestion system. Food Res. Int. 2019, 116, 1126–1134. [Google Scholar] [CrossRef]
- Ferchichi, N.; Toukabri, W.; Vrhovsek, U.; Nouairi, I.; Angeli, A.; Masuero, D.; Mhamdi, R.; Trabelsi, D. Proximate composition, lipid and phenolic profiles, and antioxidant activity of different ecotypes of Lupinus albus, Lupinus luteus and Lupinus angustifolius. J. Food Meas. Charact. 2021, 15, 1241–1257. [Google Scholar] [CrossRef]
- Magalhães, S.C.Q.; Taveira, M.; Cabrita, A.R.; Fonseca, A.J.; Valentão, P.; Andrade, P.B. European marketable grain legume seeds: Further insight into phenolic compounds profiles. Food Chem. 2017, 215, 177–184. [Google Scholar] [CrossRef]
- Siger, A.; Czubinski, J.; Kachlicki, P.; Dwiecki, K.; Lampart-Szczapa, E.; Nogala-Kalucka, M. Antioxidant activity and phenolic content in three lupin species. J. Food Compos. Anal. 2012, 25, 190–197. [Google Scholar] [CrossRef]
- Karamać, M.; Orak, H.H.; Amarowicz, R.; Orak, A.; Piekoszewski, W. Phenolic contents and antioxidant capacities of wild and cultivated white lupin (Lupinus albus L.) seeds. Food Chem. 2018, 258, 1–7. [Google Scholar] [CrossRef]
- Król, A.; Amarowicz, R.; Weidner, S. Content of phenolic compounds and antioxidant properties in seeds of sweet and bitter cultivars of lupine (Lupinus angustifolius). Nat. Prod. Commun. 2018, 13, 1341–1344. [Google Scholar] [CrossRef]
- Lampart-Szczapa, E.; Korczak, J.; Nogala-Kalucka, M.; Zawirska-Wojtasiak, R. Antioxidant properties of lupin seed products. Food Chem. 2003, 83, 279–285. [Google Scholar] [CrossRef]
- Kalogeropoulos, N.; Chiou, A.; Ioannou, M.; Karathanos, V.T.; Hassapidou, M.; Andrikopoulos, N.K. Nutritional evaluation and bioactive microconstituents (phytosterols, tocopherols, polyphenols, triterpenic acids) in cooked dry legumes usually consumed in the Mediterranean countries. Food Chem. 2010, 121, 682–690. [Google Scholar] [CrossRef]
- Martínez-Villaluenga, C.; Zielinski, H.; Frias, J.; Piskuła, M.K.; Kozłowska, H.; Vidal-Valverde, C. Antioxidant capacity and polyphenolic content of high-protein lupin products. Food Chem. 2009, 112, 84–88. [Google Scholar] [CrossRef]
- Mohammed, M.A.; Mohamed, E.A.; Yagoub, A.E.A.; Mohamed, A.R.; Babiker, E.E. Effect of processing methods on alkaloids, phytate, phenolics, antioxidants activity and minerals of newly developed lupin (Lupinus albus L.) cultivar. J. Food Proc. Preserv. 2016, 41, e12960. [Google Scholar] [CrossRef]
- Khan, M.K.; Karnpanit, W.; Nasar-Abbas, S.M.; Huma, Z.; Jayasena, V. Phytochemical composition and bioactivities of lupin: A review. Int. J. Food Sci. Technol. 2015, 50, 2004–2012. [Google Scholar] [CrossRef]
- Chan, Y.T.; Hui, S. In-vitro assay as screening tools for determination of the antioxidant activities of quinolizidine (lupin) alkaloids. In Proceedings of the International Congress on Traditional Medicine, Beijing, China, 22–24 April 2000. [Google Scholar]
- Hassine, A.B.; Rocchetti, G.; Zhang, L.; Senizza, B.; Zengin, G.; Mahomoodally, M.F.; Ben-Attia, M.; Rouphael, Y.; Lucini, L.; El-Bok, S. Untargeted phytochemical profile, antioxidant capacity and enzyme inhibitory activity of cultivated and wild lupin seeds from Tunisia. Molecules 2021, 26, 3452. [Google Scholar] [CrossRef]
- Magalhães, S.C.Q.; Fernandes, F.; Cabrita, A.R.J.; Fonseca, A.J.M.; Valentão, P.; Andrade, P.B. Alkaloids in the valorization of European Lupinus spp. seeds crop. Ind. Crops Prod. 2017, 95, 286–295. [Google Scholar] [CrossRef]
- Tomar, M.; Bhardwaj, R.; Kumar, M.; Singh, S.P.; Krishnan, V.; Kansal, R.; Verma, R.; Yadav, V.K.; Ahlawat, S.P.; Rana, J.C.; et al. Development of NIR spectroscopy-based prediction models for nutritional profiling of pearl millet (Pennisetum glaucum (L.)) R. Br: A chemometrics approach. LWT 2021, 149, 111813. [Google Scholar] [CrossRef]
- Can, A.; Ayvaz, H.; Pala, Ç.U.; Condelli, N.; Galgano, F.; Tolve, R. The potential of near and mid-infrared spectroscopy for rapid quantification of oleuropein, total phenolics, total flavonoids and antioxidant activity in olive tree (Olea europaea) leaves. J. Food Meas. Charact. 2018, 12, 2747–2757. [Google Scholar] [CrossRef]
- Wang, J.; Liu, H.; Ren, G. Near-infrared spectroscopy (NIRS) evaluation and regional analysis of Chinese faba bean (Vicia faba L.). Crop J. 2014, 2, 28–37. [Google Scholar] [CrossRef]
- Goodchild, A.V.; El Haramein, F.J.; Abd El Moneim, A.; Makkar, H.P.S.; Williams, P.C. Prediction of phenolics and tannins in forage legumes by near infrared reflectance. J. Near Infrared Spectrosc. 1998, 6, 175–181. [Google Scholar] [CrossRef]
- Zhang, K.; Zhou, L.; Brady, M.; Xu, F.; Yu, J.; Wang, D. Fast analysis of high heating value and elemental compositions of sorghum biomass using near-infrared spectroscopy. Energy 2017, 118, 1353–1360. [Google Scholar] [CrossRef]
- Carbas, B.; Machado, N.; Oppolzer, D.; Queiroz, M.; Brites, C.; Rosa, E.A.; Barros, A.I. Prediction of phytochemical composition, in vitro antioxidant activity and individual phenolic compounds of common beans using MIR and NIR spectroscopy. Food Bioprocess Technol. 2020, 13, 962–977. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| L. mutabilis | L. albus | L. angustifolius | L. luteus | |
|---|---|---|---|---|
| Free phenolics content | ||||
| Genistein | 135.78 ± 10.49 | 2.21 ± 0.00 | nd | 40.39 ± 3.89 |
| (38.03–352.12) | (nd–13.26) | |||
| Genistein der * | 464.69 ± 25.30 | 0.82 ± 0.00 | nd | 230.00 ± 13.68 |
| (219.57–1062.84) | (nd–4.95) | |||
| Apigenin der | 13.02 ± 1.01 | 6.19 ± 0.79 | 14.82 ± 1.47 | 27.82 ± 0.39 |
| (2.70–27.99) | (6.79–17.31) | |||
| Catechin der | 6.11 ± 1.06 | nd | nd | nd |
| (nd–36.49) | ||||
| Diosmetin der | 64.69 ± 4.35 | nd | nd | 98.43 ± 5.47 |
| (7.23–168.19) | ||||
| Naringenin der | 5.36 ± 0.31 | nd | nd | 2.03 ± 0.07 |
| (nd–11.58) | ||||
| Tyrosol | 11.30 ± 1.58 | nd | nd | nd |
| (nd–49.21) | ||||
| Tyrosol der | 24.90 ± 2.31 | nd | nd | 49.42 ± 4.07 |
| (nd–60.27) | ||||
| Cinnamic acid der | 3.49 ± 0.47 | nd | nd | nd |
| (nd–11.92) | ||||
| Total free phenolics | 729.36 ± 34.74 | 9.23 ± 0.79 | 14.82 ± 1.47 | 448.09 ± 15.84 |
| (340.81–1393.32) | (6.79–31.27) | |||
| Antioxidant capacity | ||||
| Reducing power | 2.53 ± 0.15 | 1.03 ± 0.19 | 0.89 ± 0.02 | 4.01 ± 0.53 |
| (1.09–6.68) | (1.44–3.10) | |||
| ABTS | 40.17 ± 1.74 | 7.41 ± 3.03 | 9.23 ± 0.49 | 31.34 ± 0.80 |
| (17.25–74.27) | (2.36–21.98) | |||
| DPPH | 2.63 ± 0.14 | 0.34 ± 0.06 | 0.50 ± 0.02 | 0.86 ± 0.07 |
| (1.06–6.47) | (0.25–1.40) | |||
| FRAP | 18.41 ± 0.74 | 2.30 ± 0.06 | 5.08 ± 0.11 | 12.44 ± 0.04 |
| (9.48–30.90) | (3.90–5.21) | |||
| RP | ABTS | DPPH | FRAP | |
|---|---|---|---|---|
| Genistein | 0.35 * | 0.52 *** | 0.55 *** | 0.69 *** |
| Genistein der | 0.59 *** | 0.60 *** | 0.76 *** | 0.80 *** |
| Apigenin der | 0.38 * | 0.49 ** | 0.47 ** | 0.58 *** |
| Catechin der | 0.05 | 0.42 ** | 0.47 ** | 0.45 ** |
| Diosmetin der | 0.16 | 0.21 | 0.17 | 0.32 |
| Naringenin der | 0.10 | 0.45 ** | 0.34 * | 0.35 * |
| Tyrosol | 0.29 | 0.73 *** | 0.68 *** | 0.74 *** |
| Tyrosol der | 0.29 | 0.20 | −0.02 | −0.04 |
| Cinnamic acid der | 0.04 | −0.04 | −0.05 | −0.11 |
| Flavonoids | 0.61 *** | 0.45 ** | 0.63 *** | 0.71 *** |
| Phenylethanoids | 0.40 | 0.52 *** | 0.22 | 0.15 |
| Phenolic acids | 0.01 | −0.25 | −0.27 | −0.40 * |
| Total phenolics | 0.63*** | 0.48** | 0.64*** | 0.71*** |
| Area | ||||
|---|---|---|---|---|
| North | Centre-North | Centre-South | South | |
| N° Ecotypes | 5 | 13 | 9 | 6 |
| Flavonoids | 592.34 ± 67.65 | 699.20 ± 84.36 | 796.46 ± 116.43 | 589.92 ± 167.02 |
| Phenylethanoids | 35.68 ± 8.27 | 37.25 ± 5.51 | 32.99 ± 5.99 | 39.18 ± 20.89 |
| Phenolic acids | 2.70 ± 1.15 | 4.65 ± 1.19 | 2.72 ± 1.20 | 2.82 ± 4.54 |
| Total phenolics | 630.72 ± 60.76 | 741.10 ± 84.46 | 832.16 ± 120.66 | 631.91 ± 156.70 |
| RP | 2.63 ± 0.50 ab | 2.26 ± 0.19 b | 3.10 ± 0.59 a | 2.19 ± 0.58 b |
| ABTS | 36.17 ± 3.90 bc | 34.95 ± 3.84 c | 44.02 ± 4.24 ab | 49.02 ± 17.27 a |
| DPPH | 2.13 ± 0.32 b | 2.27 ± 0.22 b | 3.42 ± 0.50 a | 2.64 ± 0.92 ab |
| FRAP | 15.73 ± 2.27 b | 16.52 ± 1.42 b | 21.52 ± 2.29 a | 20.09 ± 5.93 ab |
| Parameter | Range | Variables (cm−1) | LVs | CAL | CV | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| R2 | RMSE | RMSPE | Bias | R2 | RMSE | RMSPE | Bias | RPD | ||||
| Total phenolics (mg/kg DM) | 300.00–1393.32 | 6823–6711, 6245–6133, 4509–4397, 4278–4050 | 7 | 0.92 | 78.50 | 12.1 | 4.55−12 | 0.84 | 112.40 | 17.4 | 1.11−2 | 3.0 |
| Flavonoids (mg/kg DM) | 295.00–1340.11 | 6823–6711, 6245–6133, 4509–4397, 4278–4050 | 7 | 0.92 | 81.00 | 13.3 | 3.75−12 | 0.83 | 116.80 | 19.1 | 7.20−2 | 2.8 |
| Carotenoids (mg/kg DM) | 0.68–7.14 | 4586–4494 | 3 | 0.89 | 0.42 | 21.2 | 8.88−16 | 0.86 | 0.51 | 25.8 | 4.33−2 | 2.4 |
| Tocols (mg/kg DM) | 103.22–378.22 | 6245–6017 | 3 | 0.90 | 18.20 | 6.4 | 1.71−13 | 0.86 | 23.80 | 8.4 | 9.72−2 | 2.5 |
| FRAP (mmol TE/kg DM) | 3.90–30.90 | 9000–3800 | 2 | 0.80 | 2.56 | 14.2 | −3.55−12 | 0.76 | 2.89 | 16.0 | 1.77−2 | 2.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Estivi, L.; Grassi, S.; Briceño-Berrú, L.; Glorio-Paulet, P.; Camarena, F.; Hidalgo, A.; Brandolini, A. Free Phenolic Compounds, Antioxidant Capacity and FT-NIR Survey of Debittered Lupinus mutabilis Seeds. Processes 2022, 10, 1637. https://doi.org/10.3390/pr10081637
Estivi L, Grassi S, Briceño-Berrú L, Glorio-Paulet P, Camarena F, Hidalgo A, Brandolini A. Free Phenolic Compounds, Antioxidant Capacity and FT-NIR Survey of Debittered Lupinus mutabilis Seeds. Processes. 2022; 10(8):1637. https://doi.org/10.3390/pr10081637
Chicago/Turabian StyleEstivi, Lorenzo, Silvia Grassi, Luis Briceño-Berrú, Patricia Glorio-Paulet, Felix Camarena, Alyssa Hidalgo, and Andrea Brandolini. 2022. "Free Phenolic Compounds, Antioxidant Capacity and FT-NIR Survey of Debittered Lupinus mutabilis Seeds" Processes 10, no. 8: 1637. https://doi.org/10.3390/pr10081637