Towards a Sustainable One Health Approach to Crimean–Congo Hemorrhagic Fever Prevention: Focus Areas and Gaps in Knowledge

 , , and
, , and
Abstract
:1. Introduction
2. Surveillance
2.1. Human Surveillance
2.2. Animal Surveillance
2.3. Tick Surveillance
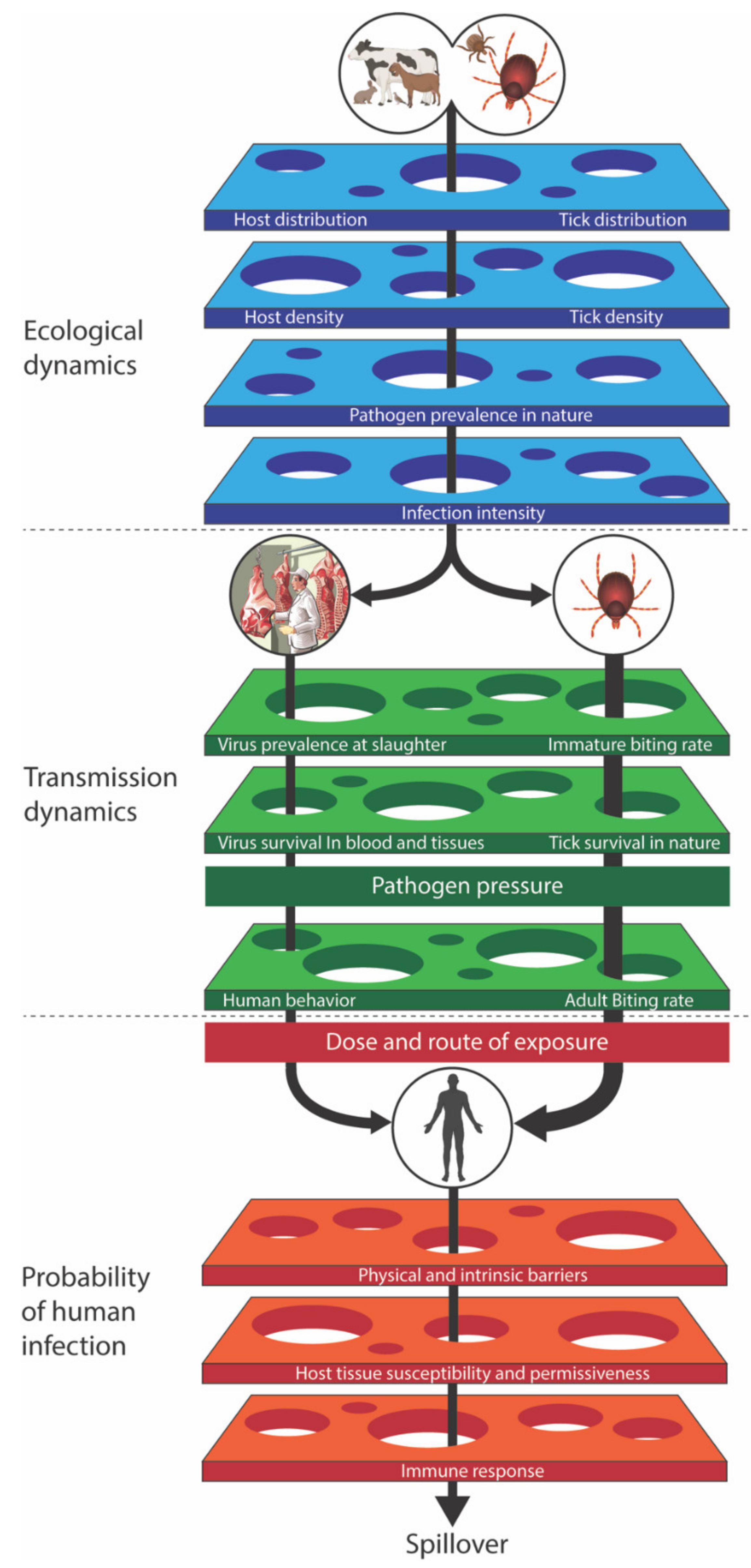
3. Risk Assessment
3.1. Modeling Tick-Borne Viruses
3.2. What Key Data Are Needed?
3.3. Important Features of CCHF Modeling
3.3.1. Multiple Host Species and the Dilution Effect
3.3.2. Non-Viremic Transmission through Co-Feeding
3.3.3. Transovarial Transmission
3.3.4. Sexual Transmission of the Virus in Ticks
3.3.5. Role of Other Tick Species: Vector Competence and Vectorial Capacity
4. Risk Reduction: Human-Targeted Approaches
4.1. Physical and Chemical
4.2. Behavior
4.3. Vaccination
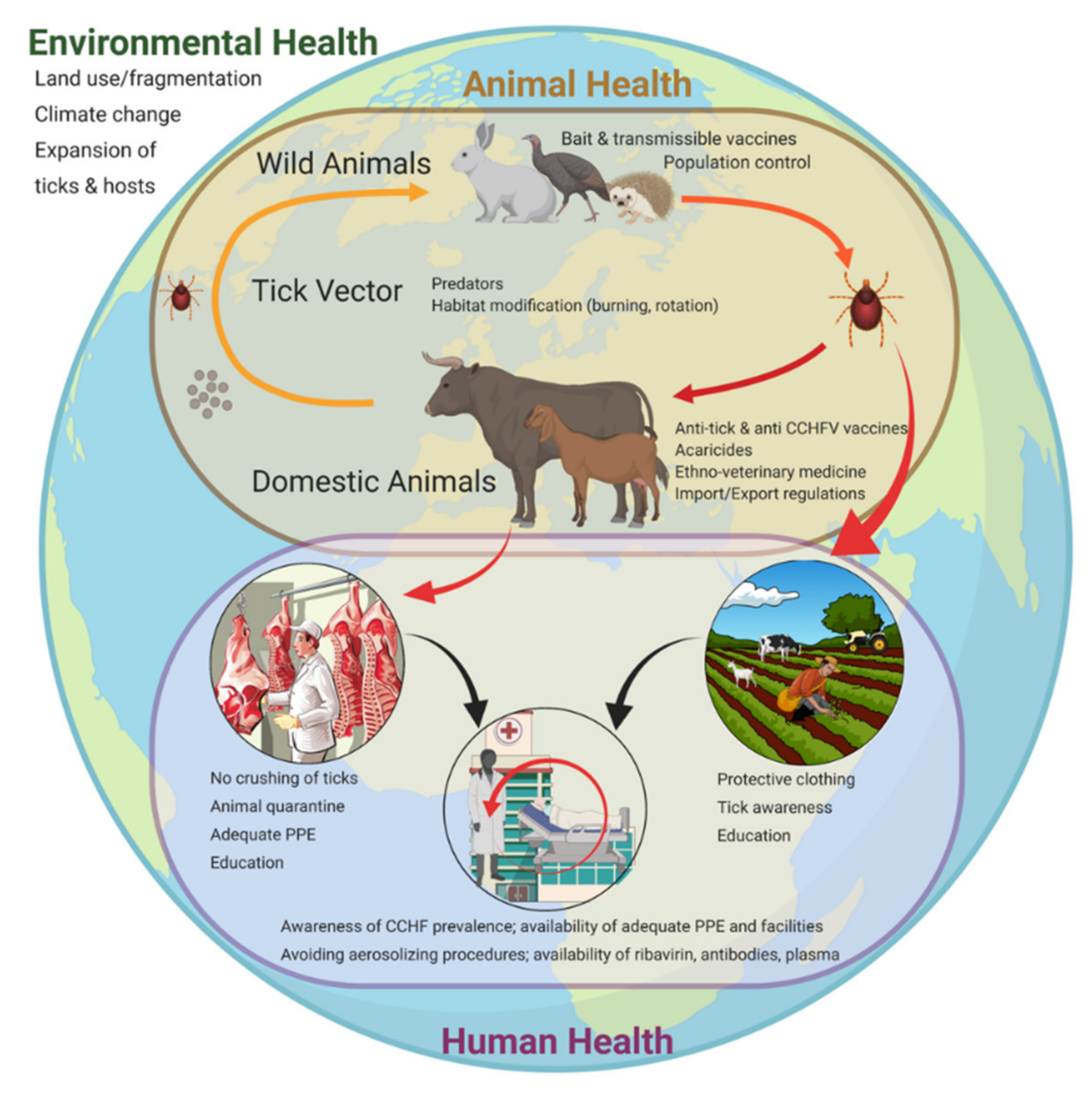
5. Risk Reduction: Animal-Targeted Approaches
5.1. Wild Animals
5.2. Domestic Animals
6. Risk Reduction: Tick-Targeted Approaches
6.1. Physical
6.2. Chemical
6.3. Biological
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Disclaimer
References
- Bente, D.A.; Forrester, N.L.; Watts, D.M.; McAuley, A.J.; Whitehouse, C.A.; Bray, M. Crimean-Congo hemorrhagic fever: History, epidemiology, pathogenesis, clinical syndrome and genetic diversity. Antivir. Res. 2013, 100, 159–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estrada-Peña, A.; Vatansever, Z.; Gargili, A.; Ergönul, Ö. The trend towards habitat fragmentation is the key factor driving the spread of Crimean-Congo haemorrhagic fever. Epidemiol. Infect. 2010, 138, 1194–1203. [Google Scholar] [CrossRef] [PubMed]
- Spengler, J.R.; Bente, D.A. Crimean-congo hemorrhagic fever in Spain—New arrival or silent resident? N. Engl. J. Med. 2017, 377. [Google Scholar] [CrossRef] [PubMed]
- Ansari, H.; Shahbaz, B.; Izadi, S.; Zeinali, M.; Tabatabaee, S.M.; Mahmoodi, M.; Holakouie-Naieni, K.; Mansournia, M.A. Crimean-Congo hemorrhagic fever and its relationship with climate factors in southeast Iran: A 13-year experience. J. Infect. Dev. Ctries. 2014, 8, 749–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duygu, F.; Sari, T.; Kaya, T.; Tavsan, O.; Naci, M. The relationship between Crimean-Congo hemorrhagic fever and climate: Does climate affect the number of patients? Acta Clin. Croat. 2018, 57, 443–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estrada-Peña, A.; Ruiz-Fons, F.; Acevedo, P.; Gortazar, C.; de la Fuente, J. Factors driving the circulation and possible expansion of Crimean-Congo haemorrhagic fever virus in the western Palearctic. J. Appl. Microbiol. 2013, 114, 278–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Records of Crimean-Congo Hemorrhagic Fever, Communicable Diseases Department; The Ministry of Health of Turkey: Ankara, Turkey, 2008.
- Blair, P.W.; Kuhn, J.H.; Pecor, D.B.; Apanaskevich, D.A.; Kortepeter, M.G.; Cardile, A.P.; Ramos, A.P.; Keshtkar-Jahromi, M. An emerging biothreat: Crimean-Congo hemorrhagic fever virus in southern and western Asia. Am. J. Trop. Med. Hyg. 2019, 100, 16–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spengler, J.R.; Estrada-Peña, A. Host preferences support the prominent role of Hyalomma ticks in the ecology of Crimean-Congo hemorrhagic fever. PLoS Negl. Trop. Dis. 2018, 12, e0006248. [Google Scholar] [CrossRef] [Green Version]
- Zinsstag, J.; Schelling, E.; Waltner-Toews, D.; Tanner, M. From “one medicine” to “one health” and systemic approaches to health and well-being. Prev. Vet. Med. 2011, 101, 148–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FAO; OIE; WHO. A Tripartite Guide to Addressing Zoonotic Diseases in Countries; World Health Organization (WHO); Food and Agriculture Organization of the United Nations (FAO); World Organisation for Animal Health (OIE): Geneva, Switzerland, 2019; ISBN 978-92-5-131236-0. [Google Scholar]
- Mertens, M.; Schmidt, K.; Ozkul, A.; Groschup, M.H. The impact of Crimean-Congo hemorrhagic fever virus on public health. Antivir. Res. 2013, 98, 248–260. [Google Scholar] [CrossRef]
- Estrada-Peña, A.; Zatansever, Z.; Gargili, A.; Aktas, M.; Uzun, R.; Ergonul, O.; Jongejan, F. Modeling the spatial distribution of Crimean-Congo hemorrhagic fever outbreaks in Turkey. Vector Borne Zoonotic Dis. 2007, 7, 667–678. [Google Scholar] [CrossRef] [PubMed]
- Formenty, P.; Schnepf, G.; Gonzalez-Martin, F.; Bi, Z. International surveillance and control of Crimean-Congo hemorrhagic fever outbreaks. In Crimean-Congo Hemorrhagic Fever: A Global Perspective; Springer: Dordrecht, The Netherlands, 2007; pp. 295–303. ISBN 9781402061066. [Google Scholar]
- WHO. WHO CCHF R&D Roadmap Meeting; World Health Organization (WHO): Geneva, Switzerland, 2018. [Google Scholar]
- Spengler, J.R.; Bente, D.A.; Bray, M.; Burt, F.; Hewson, R.; Korukluoglu, G.; Mirazimi, A.; Weber, F.; Papa, A. Second international conference on Crimean-Congo hemorrhagic fever. Antivir. Res. 2018, 150. [Google Scholar] [CrossRef] [PubMed]
- Bodur, H.; Akinci, E.; Ascioglu, S.; Öngürü, P.; Uyar, Y. Subclinical infections with Crimean-Congo hemorrhagic fever virus, Turkey. Emerg. Infect. Dis. 2012. [Google Scholar] [CrossRef] [PubMed]
- Gunes, T.; Engin, A.; Poyraz, O.; Elaldi, N.; Kaya, S.; Dokmetas, I.; Bakir, M.; Cinar, Z. Crimean-Congo hemorrhagic fever virus in high-risk population, Turkey. Emerg. Infect. Dis. 2009, 15, 461–464. [Google Scholar] [CrossRef] [PubMed]
- Ertugrul, B.; Kirdar, S.; Ersoy, O.S.; Ture, M.; Erol, N.; Ozturk, B.; Sakarya, S. The seroprevalence of Crimean-Congo haemorrhagic fever among inhabitants living in the endemic regions of Western Anatolia. Scand. J. Infect. Dis. 2012, 44, 276–281. [Google Scholar] [CrossRef]
- Nasirian, H. Crimean-Congo hemorrhagic fever (CCHF) seroprevalence: A systematic review and meta-analysis. Acta Trop. 2019, 196, 102–120. [Google Scholar] [CrossRef]
- Papadopoulos, O.; Koptopoulos, G. Isolation of Crimean-Congo hemorrhagic fever (CCHF) virus from Rhipicephalus bursa ticks in Greece. Acta Microbiolog. Hell. 1978, 23, 20–28. [Google Scholar]
- Papa, A.; Sidira, P.; Larichev, V.; Gavrilova, L.; Kuzmina, K.; Mousavi-Jazi, M.; Mirazimi, A.; Stroher, U.; Nichol, S. Crimean-Congo hemorrhagic fever virus, Greece. Emerg. Infect. Dis. 2014, 20, 288–290. [Google Scholar] [CrossRef] [Green Version]
- Salehi-Vaziri, M.; Baniasadi, V.; Jalali, T.; Mirghiasi, S.M.; Azad-Manjiri, S.; Zarandi, R.; Mohammadi, T.; Khakifirouz, S.; Fazlalipour, M. The first fatal case of Crimean-Congo hemorrhagic fever caused by the AP92-like strain of the Crimean-Congo hemorrhagic fever virus. Jpn. J. Infect. Dis. 2016, 69, 344–346. [Google Scholar] [CrossRef] [Green Version]
- Midilli, K.; Gargili, A.; Ergonul, O.; Elevli, M.; Ergin, S.; Turan, N.; Şengöz, G.; Ozturk, R.; Bakar, M. The first clinical case due to AP92 like strain of Crimean-Congo hemorrhagic fever virus and a field survey. BMC Infect. Dis. 2009, 9. [Google Scholar] [CrossRef] [Green Version]
- Vanhomwegen, J.; Alves, M.J.; Županc, T.A.; Bino, S.; Chinikar, S.; Karlberg, H.; Korukluoǧlu, G.; Korva, M.; Mardani, M.; Mirazimi, A.; et al. Diagnostic assays for Crimean-Congo hemorrhagic fever. Emerg. Infect. Dis. 2012, 18, 1958–1965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raabe, V.N. Diagnostic testing for Crimean-Congo hemorrhagic fever. J. Clin. Microbiol. 2020, 58. [Google Scholar] [CrossRef] [PubMed]
- Escadafal, C.; Ölschläger, S.; Avšič-Županc, T.; Papa, A.; Vanhomwegen, J.; Wölfel, R.; Mirazimi, A.; Teichmann, A.; Donoso-Mantke, O.; Niedrig, M. First international external quality assessment of molecular detection of Crimean-Congo hemorrhagic fever virus. PLoS Negl. Trop. Dis. 2012, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spengler, J.R.; Bergeron, É.; Rollin, P.E. Seroepidemiological studies of Crimean-Congo hemorrhagic fever virus in domestic and wild animals. PLoS Negl. Trop. Dis. 2016, 10, e0004210. [Google Scholar] [CrossRef] [Green Version]
- Hoogstraal, H. The epidemiology of tick-borne Crimean-Congo hemorrhagic fever in Asia, Europe, and Africa. J. Med. Entomol. 1979, 5, 307–417. [Google Scholar] [CrossRef]
- Spengler, J.R.; Estrada-Peña, A.; Garrison, A.R.; Schmaljohn, C.; Spiropoulou, C.F.; Bergeron, É.; Bente, D.A. A chronological review of experimental infection studies of the role of wild animals and livestock in the maintenance and transmission of Crimean-Congo hemorrhagic fever virus. Antivir. Res. 2016, 135. [Google Scholar] [CrossRef] [Green Version]
- Hawman, D.W.; Meade-White, K.; Haddock, E.; Habib, R.; Scott, D.; Thomas, T.; Rosenke, R.; Feldmann, H. Crimean-Congo hemorrhagic fever mouse model recapitulating human convalescence. J. Virol. 2019, 93. [Google Scholar] [CrossRef] [Green Version]
- Welch, S.R.; Scholte, F.E.M.; Spengler, J.R.; Ritter, J.M.; Coleman-Mccray, J.D.; Harmon, J.R.; Nichol, S.T.; Zaki, S.R.; Spiropoulou, C.F.; Bergeron, E. The Crimean-Congo hemorrhagic fever virus NSm protein is dispensable for growth in vitro and disease in IFNAR-/- mice. Microorganisms 2020, 8. [Google Scholar] [CrossRef]
- Chumakov, M. Contribution to 30 years of investigation of Crimean hemorrhagic fever. Translyatsia Instituta Polio-Virusnyh Entsefalitov Akademii Medicinskih NauK USSR, 1974; 22, 5–18. (In Russian). (In English, NAMRU-T950) [Google Scholar]
- Causey, O.R.; Kemp, G.E.; Madbouly, M.H.; David-West, T.S. Congo virus from domestic livestock, African hedgehog, and arthropods in Nigeria. Am. J. Trop. Med. Hyg. 1970, 19, 846–850. [Google Scholar] [CrossRef]
- Shanmugam, J.; Smirnova, S.E.; Chumakov, M.P. Presence of antibody to arboviruses of the Crimean haemorrhagic fever-Congo (CHF-Congo) group in human beings and domestic animals in India. Indian J. Med. Res. 1976, 64, 1403–1413. [Google Scholar] [PubMed]
- Goedhals, D.; Paweska, J.T.; Burt, F.J. Long-lived CD8+ T cell responses following Crimean-Congo haemorrhagic fever virus infection. PLoS Negl. Trop. Dis. 2017, 11. [Google Scholar] [CrossRef] [PubMed]
- Burt, F.J.; Samudzi, R.R.; Randall, C.; Pieters, D.; Vermeulen, J.; Knox, C.M. Human defined antigenic region on the nucleoprotein of Crimean-Congo hemorrhagic fever virus identified using truncated proteins and a bioinformatics approach. J. Virol. Methods 2013, 193, 706–712. [Google Scholar] [CrossRef] [PubMed]
- Burt, F.J.; Leman, P.A.; Abbott, J.C.; Swanepoel, R. Serodiagnosis of Crimean-Congo haemorrhagic fever. Epidemiol. Infect. 1994. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, J.-P.; Camicas, J.-L.; Cornet, J.-P.; Wilson, M.L. Biological and clinical responses of West African sheep to Crimean-Congo haemorrhagic fever virus experimental infection. Res. Virol. 1998, 149, 445–455. [Google Scholar] [CrossRef]
- Tignor, G.H.; Smith, A.L.; Casals, J.; Ezeokoli, C.D.; Okoli, J. Close relationship of Crimean hemorrhagic fever-Congo (CHF-C) virus strains by neutralizing antibody assays. Am. J. Trop. Med. Hyg. 1980, 29, 676–685. [Google Scholar] [CrossRef]
- Marriott, A.C.; Ward, V.K.; Higgs, S.; Nuttall, P.A. RNA probes detect nucleotide sequence homology between members of two different nairovirus serogroups. Virus Res. 1990, 16, 77–81. [Google Scholar] [CrossRef]
- Ward, V.K.; Marriott, A.C.; Polyzoni, T.; El-Ghorr, A.A.; Antoniadis, A.; Nuttall, P.A. Expression of the nucleocapsid protein of Dugbe virus and antigenic cross-reactions with other nairoviruses. Virus Res. 1992, 24, 223–229. [Google Scholar] [CrossRef]
- Kondratenko, V.; Blagoveshchenskaya, N.M.; Butenko, A.M.; Vyshnivetskaya, L.K.; Zarubina, L.V.; Milyutin, V.N. Results of virological investigation of ixodid ticks in Crimean hemorrhagic fever focus in Rostov Oblast. Mater 3 Oblast Nauchn Prakt Konf. 1970; 3, 29–35. (In Russian). (In English, NAMRU3-T524) [Google Scholar]
- Chumakov, M.P. A short story of the investigation of the virus of Crimean hemorrhagic fever. Translyatsia Instituta Polio-Virusnyh Entsefalitov Akademii Medicinskih NauK USSR, 1965; 7, 193–196. (In Russian). (In English, NAMRU3-TI89) [Google Scholar]
- Pak, T.P.; Daniyarov, O.A.; Kostyukov, M.A.; Bulychev, V.P.; Kuima, A.U. Ecology of Crimean hemorrhagic fever in Tadzhikistan. Mater Resp. Simp. Kamenyuki Belovezh Pushoha, 1974; 93–94. (In Russian). (In English, NAMRU3-T968). [Google Scholar]
- Swanepoel, R.; Struthers, J.K.; Shepherd, A.J.; McGillivray, G.M.; Nel, M.J.; Jupp, P.G. Crimean-Congo hemorrhagic fever in South Africa. Am. J. Trop. Med. Hyg. 1983, 32, 1407–1415. [Google Scholar] [CrossRef]
- Gargili, A.; Estrada-Peña, A.; Spengler, J.R.; Lukashev, A.; Nuttall, P.A.; Bente, D.A. The role of ticks in the maintenance and transmission of Crimean-Congo hemorrhagic fever virus: A review of published field and laboratory studies. Antivir. Res. 2017, 144. [Google Scholar] [CrossRef]
- Gonzalez, J.P.; Cornet, J.P.; Wilson, M.L.; Camicas, J.L. Crimean-Congo haemorrhagic fever virus replication in adult Hyalomma truncatum and Amblyomma variegatum ticks. Res. Virol. 1991, 142, 483–488. [Google Scholar] [CrossRef]
- Shepherd, A.J.; Swanepoel, R.; Cornel, A.J.; Mathee, O. Experimental studies on the replication and transmission of Crimean-Congo hemorrhagic fever virus in some African tick species. Am. J. Trop. Med. Hyg. 1989, 40, 326–331. [Google Scholar] [CrossRef]
- Kar, S.; Rodriguez, S.E.; Akyildiz, G.; Cajimat, M.N.B.; Bircan, R.; Mears, M.C.; Bente, D.A.; Keles, A.G. Crimean-Congo hemorrhagic fever virus in tortoises and Hyalomma aegyptium ticks in East Thrace, Turkey: Potential of a cryptic transmission cycle. Parasites Vectors 2020, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palomar, A.M.; Portillo, A.; Santibáñez, P.; Mazuelas, D.; Arizaga, J.; Crespo, A.; Gutiérrez, Ó.; Cuadrado, J.F.; Oteo, J.A. Crimean-congo hemorrhagic fever virus in ticks from migratory birds, Morocco. Emerg. Infect. Dis. 2013, 19, 260–263. [Google Scholar] [CrossRef] [PubMed]
- Chisholm, K.; Dueger, E.; Fahmy, N.T.; Samaha, H.A.T.; Zayed, A.; Abdel-Dayem, M.; Villinski, J.T. Crimean-Congo hemorrhagic fever virus in ticks from imported livestock, Egypt. Emerg. Infect. Dis. 2012, 18, 181–182. [Google Scholar] [CrossRef]
- De Liberato, C.; Frontoso, R.; Magliano, A.; Montemaggiori, A.; Autorino, G.L.; Sala, M.; Bosworth, A.; Scicluna, M.T. Monitoring for the possible introduction of Crimean-Congo haemorrhagic fever virus in Italy based on tick sampling on migratory birds and serological survey of sheep flocks. Prev. Vet. Med. 2018, 149, 47–52. [Google Scholar] [CrossRef]
- Buczek, A.M.; Buczek, W.; Buczek, A.; Bartosik, K. The potential role of migratory birds in the rapid spread of ticks and tick-borne pathogens in the changing climatic and environmental conditions in Europe. Int. J. Environ. Res. Public Health 2020, 17, 2117. [Google Scholar] [CrossRef] [Green Version]
- Leggiadro, R.J. Autochthonous Crimean-Congo hemorrhagic fever in Spain. Pediatr. Infect. Dis. J. 2017. [Google Scholar] [CrossRef]
- Estrada-Peña, A.; Palomar, A.M.; Santibáñez, P.; Sánchez, N.; Habela, M.A.; Portillo, A.; Romero, L.; Oteo, J.A. Crimean-Congo hemorrhagic fever virus in ticks, Southwestern Europe, 2010. Emerg. Infect. Dis. 2012, 18, 179–180. [Google Scholar] [CrossRef] [PubMed]
- Sureau, P.; Klein, J.M.; Casals, J.; Digoutte, J.; Salaun, J.; Piazak, N.; Calvo, M. Isolation of Thogoto, Wad Medani, Wanowrie and Crimean-Congo haemorrhagic fever viruses from ticks of domestic animals in Iran [Hyalomma anatolicum, Hyalomma asiaticum, Alveonasus lahorensis; isolates, epidemiology]. Ann. Virol. 1980, 131, 185–200. [Google Scholar]
- Wilson, M.L.; LeGuenno, B.; Guillaud, M.; Desoutter, D.; Gonzalez, J.P.; Camicas, J.L. Distribution of Crimean-Congo hemorrhagic fever viral antibody in Senegal: Environmental and vectorial correlates. Am. J. Trop. Med. Hyg. 1990, 43, 557–566. [Google Scholar] [CrossRef] [Green Version]
- Logan, T.M.; Linthicum, K.J.; Bailey, C.L.; Watts, D.M.; Moulton, J.R. Experimental transmission of Crimean-Congo hemorrhagic fever virus by Hyalomma truncatum koch. Am. J. Trop. Med. and Hyg. 1989, 40, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Şekercioĝlu, Ç.H. Guineafowl, Ticks and Crimean-Congo hemorrhagic fever in Turkey: The perfect storm? Trends Parasitol. 2013, 29, 1–2. [Google Scholar] [CrossRef]
- Plowright, R.K.; Parrish, C.R.; McCallum, H.; Hudson, P.J.; Ko, A.I.; Graham, A.L.; Lloyd-Smith, J.O. Pathways to zoonotic spillover. Nat. Rev. Microbiol. 2017, 15, 502–510. [Google Scholar] [CrossRef] [Green Version]
- Dietz, K. The estimation of the basic reproduction number for infectious diseases. Stat. Methods Med. Res. 1993, 2, 23–41. [Google Scholar] [CrossRef]
- Cooper, B.S. Mathematical modeling of Crimean-Congo hemorrhagic fever transmission. In Crimean-Congo Hemorrhagic Fever: A Global Perspective; Springer: Dordrecht, The Netherlands, 2007; pp. 187–203. ISBN 9781402061066. [Google Scholar]
- Medley, G.F.; Perry, B.; Young, A.A.S. Preliminary analysis of the transmission dynamics of Theileria parva in eastern Africa. Parasitology 1993, 106, 251–264. [Google Scholar] [CrossRef]
- Ak, Ç.; Ergönül, Ö.; Şencan, İ.; Torunoğlu, M.A.; Gönen, M. Spatiotemporal prediction of infectious diseases using structured Gaussian processes with application to Crimean-Congo hemorrhagic fever. PLoS Negl. Trop. Dis. 2018, 12. [Google Scholar] [CrossRef] [Green Version]
- Norman, R.; Bowers, R.G.; Begon, M.; Hudson, P.J. Persistence of tick-borne virus in the presence of multiple host species: Tick reservoirs and parasite mediated competition. J. Theor. Biol. 1999, 200, 111–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laurenson, M.K.; Norman, R.A.; Gilbert, L.; Reid, H.W.; Hudson, P.J. Identifying disease reservoirs in complex systems: Mountain hares as reservoirs of ticks and louping-ill virus, pathogens of red grouse. J. Anim. Ecol. 2003, 72, 177–185. [Google Scholar] [CrossRef] [Green Version]
- LoGiudice, K.; Ostfeld, R.S.; Schmidt, K.A.; Keesing, F. The ecology of infectious disease: Effects of host diversity and community composition on Lyme disease risk. Proc. Natl. Acad. Sci. USA 2003, 100, 567–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, L.D.; Davies, C.R.; Steele, G.M.; Nuttall, P.A. A novel mode of arbovirus transmission involving a nonviremic host. Science 1987, 237, 775–777. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S.W.; Linthicum, K.J.; Moulton, J.R. Transmission of Crimean-Congo hemorrhagic fever virus in two species of Hyalomma ticks from infected adults to cofeeding immature forms. Am. J. Trop. Med. Hyg. 1993, 48, 576–580. [Google Scholar] [CrossRef]
- Gonzalez, J.P.; Camicas, J.L.; Cornet, J.P.; Faye, O.; Wilson, M.L. Sexual and transovarian transmission of Crimean-Congo haemorrhagic fever virus in Hyalomma truncatum ticks. Res. Virol. 1992, 143, 23–28. [Google Scholar] [CrossRef]
- De la Fuente, J.; Antunes, S.; Bonnet, S.; Cabezas-Cruz, A.; Domingos, A.G.; Estrada-Peña, A.; Johnson, N.; Kocan, K.M.; Mansfield, K.L.; Nijhof, A.M.; et al. Tick-pathogen interactions and vector competence: identification of molecular drivers for tick-borne diseases. Front. Cell. Infect. Microbiol. 2017, 7. [Google Scholar] [CrossRef] [Green Version]
- Ergonul, O.O.; Celikbas, A.; Baykam, N.; Eren, S.; Dokuzoguz, B. Analysis of risk-factors among patients with Crimean-Congo haemorrhagic fever virus infection: Severity criteria revisited. Clin. Microbiol. Infect. 2006. [Google Scholar] [CrossRef] [Green Version]
- Mediannikov, O.; Diatta, G.; Fenollar, F.; Sokhna, C.; Trape, J.F.; Raoult, D. Tick-borne rickettsioses, neglected emerging diseases in rural Senegal. PLoS Negl. Trop. Dis. 2010, 4. [Google Scholar] [CrossRef] [Green Version]
- Kamani, J.; Baneth, G.; Apanaskevich, D.A.; Mumcuoglu, K.Y.; Harrus, S. Molecular detection of Rickettsia aeschlimannii in Hyalomma spp. ticks from camels (Camelus dromedarius) in Nigeria, West Africa. Med. Vet. Entomol. 2015, 29, 205–209. [Google Scholar] [CrossRef]
- Grandi, G.; Chitimia-Dobler, L.; Choklikitumnuey, P.; Strube, C.; Springer, A.; Albihn, A.; Jaenson, T.G.T.; Omazic, A. First records of adult Hyalomma marginatum and H. rufipes ticks (Acari: Ixodidae) in Sweden. Ticks TickBorne Dis. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Sedaghat, M.; Sarani, M.; Chinikar, S.; Telmadarraiy, Z.; Moghaddam, A.; Azam, K.; Nowotny, N.; Fooks, A.; Shahhosseini, N. Vector prevalence and detection of Crimean-Congo haemorrhagic fever virus in Golestan Province, Iran. J. Vector Borne Dis. 2017, 54, 353. [Google Scholar] [CrossRef]
- Coleman, N.; Coleman, S. Methods of tick removal: A systematic review of the literature. Australas. Med. J. 2017, 10, 53–62. [Google Scholar] [CrossRef]
- Akin Belli, A.; Dervis, E.; Kar, S.; Ergonul, O.; Gargili, A. Revisiting detachment techniques in human-biting ticks. J. Am. Acad. Dermatol. 2016, 75, 393–397. [Google Scholar] [CrossRef] [PubMed]
- Eisen, L. Pathogen transmission in relation to duration of attachment by Ixodes scapularis ticks. Ticks Tick Borne Dis. 2018, 9, 535–542. [Google Scholar] [CrossRef] [PubMed]
- Ebel, G.D.; Kramer, L.D. Short report: Duration of tick attachment required for transmission of Powassan virus by deer ticks. Am. J. Trop. Med. Hyg. 2004, 71. [Google Scholar] [CrossRef] [Green Version]
- Booth, T.F.; Steele, G.M.; Marriott, A.C.; Nuttall, P.A. Dissemination, replication, and trans-stadial persistence of Dugbe virus (nairovirus, bunyaviridae) in the tick vector Amblyomma variegatum. Am. J. Trop. Med. Hyg. 1991, 45, 146–157. [Google Scholar] [CrossRef]
- Leblebicioglu, H.; Sunbul, M.; Memish, Z.A.; Al-Tawfiq, J.A.; Bodur, H.; Ozkul, A.; Gucukoglu, A.; Chinikar, S.; Hasan, Z. Consensus report: Preventive measures for Crimean-Congo hemorrhagic fever during Eid-al-Adha festival. Int. J. Infect. Dis. 2015, 38, 9–15. [Google Scholar] [CrossRef] [Green Version]
- Mallhi, T.H.; Khan, Y.H.; Sarriff, A.; Khan, A.H. Crimean-Congo haemorrhagic fever virus and Eid-Ul-Adha festival in Pakistan. Lancet Infect. Dis. 2016, 16, 1332–1333. [Google Scholar] [CrossRef]
- Mallhi, T.H.; Khan, Y.H.; Tanveer, N.; Khan, A.H.; Qadir, M.I. Commentary: Surveillance of Crimean-Congo haemorrhagic fever in Pakistan. Front. Public Health 2017, 5. [Google Scholar] [CrossRef] [Green Version]
- Swanepoel, R.; Leman, P.A.; Burt, F.J.; Jardine, J.; Verwoerd, D.J.; Capua, I.; Brückner, G.K.; Burger, W.P. Experimental infection of ostriches with Crimean-Congo haemorrhagic fever virus. Epidemiol. Infect. 1998, 121, 427–432. [Google Scholar] [CrossRef]
- Papa, A.; Papadimitriou, E.; Christova, I. The Bulgarian vaccine Crimean-Congo haemorrhagic fever virus strain. Scand. J. Infect. Dis. 2011, 43, 225–229. [Google Scholar] [CrossRef] [PubMed]
- Tipih, T.; Burt, F.J. Crimean-Congo hemorrhagic fever virus: Advances in vaccine development. BioRes. Open Access 2020, 9, 137–150. [Google Scholar] [CrossRef] [PubMed]
- Dowall, S.D.; Carroll, M.W.; Hewson, R. Development of vaccines against Crimean-Congo haemorrhagic fever virus. Vaccine 2017, 35, 6015–6023. [Google Scholar] [CrossRef]
- Garrison, A.R.; Smith, D.R.; Golden, J.W. Animal models for Crimean-Congo hemorrhagic fever human disease. Viruses 2019, 11, 590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dowall, S.D.; Buttigieg, K.R.; Findlay-Wilson, S.J.D.; Rayner, E.; Pearson, G.; Miloszewska, A.; Graham, V.A.; Carroll, M.W.; Hewson, R. A Crimean-Congo hemorrhagic fever (CCHF) viral vaccine expressing nucleoprotein is immunogenic but fails to confer protection against lethal disease. Hum. Vaccines Immunother. 2016, 12, 519–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zivcec, M. Characterization of the Interferon αβ Receptor Knockout Mouse Model of Crimean-Congo Hemorrhagic Fever (CCHF) and Assessment of Adenovirus Based CCHF Virus Vaccine Efficacy and Correlates of Protection. Ph.D. Thesis, University of Manitoba, Winnipeg, MB, Canada, 2013. [Google Scholar]
- Dowall, S.D.; Graham, V.A.; Rayner, E.; Hunter, L.; Watson, R.; Taylor, I.; Rule, A.; Carroll, M.W. Protective effects of a modified Vaccinia Ankara-based vaccine candidate against Crimean-Congo haemorrhagic fever virus require both cellular and humoral responses. PLoS ONE 2016, 11. [Google Scholar] [CrossRef]
- Kortekaas, J.; Vloet, R.P.M.; Mcauley, A.J.; Shen, X.; Bosch, B.J.; de Vries, L.; Moormann, R.J.M.; Bente, D.A. Crimean-Congo hemorrhagic fever virus subunit vaccines induce high levels of neutralizing antibodies but no protection in STAT1 knockout mice. Vector Borne Zoonotic Dis. 2015, 15, 759–764. [Google Scholar] [CrossRef] [Green Version]
- Bertolotti-Ciarlet, A.; Smith, J.; Strecker, K.; Paragas, J.; Altamura, L.A.; McFalls, J.M.; Frias-Stäheli, N.; García-Sastre, A.; Schmaljohn, C.S.; Doms, R.W. Cellular localization and antigenic characterization of Crimean-Congo hemorrhagic fever virus glycoproteins. J. Virol. 2005, 79, 6152–6161. [Google Scholar] [CrossRef] [Green Version]
- Golden, J.W.; Shoemaker, C.J.; Lindquist, M.E.; Zeng, X.; Daye, S.P.; Williams, J.A.; Liu, J.; Coffin, K.M.; Olschner, S.; Flusin, O.; et al. GP38-targeting monoclonal antibodies protect adult mice against lethal Crimean-Congo hemorrhagic fever virus infection. Sci. Adv. 2019, 5. [Google Scholar] [CrossRef] [Green Version]
- Mousavi-Jazi, M.; Karlberg, H.; Papa, A.; Christova, I.; Mirazimi, A. Healthy individuals’ immune response to the Bulgarian Crimean-Congo hemorrhagic fever virus vaccine. Vaccine 2012, 30, 6225–6229. [Google Scholar] [CrossRef] [PubMed]
- Haddock, E.; Feldmann, F.; Hawman, D.W.; Zivcec, M.; Hanley, P.W.; Saturday, G.; Scott, D.P.; Thomas, T.; Korva, M.; Avšič-Županc, T.; et al. A cynomolgus macaque model for Crimean-Congo haemorrhagic fever. Nat. Microbiol. 2018, 3, 556–562. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.R.; Shoemaker, C.J.; Zeng, X.; Garrison, A.R.; Golden, J.W.; Schellhase, C.W.; Pratt, W.; Rossi, F.; Fitzpatrick, C.J.; Shamblin, J.; et al. Persistent Crimean-Congo hemorrhagic fever virus infection in the testes and within granulomas of non-human primates with latent tuberculosis. PLoS Pathog. 2019, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gortazar, C.; Diez-Delgado, I.; Barasona, J.A.; Vicente, J.; de La Fuente, J.; Boadella, M. The wild side of disease control at the wildlife-livestock-human interface: A review. Front. Vet. Sci. 2014, 1, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Artois, M.; Blancou, J.; Dupeyroux, O.; Gilot-Fromont, E. Sustainable control of zoonotic pathogens in wildlife: How to be fair to wild animals? OIE Rev. Sci. Tech. 2011, 30, 733–743. [Google Scholar] [CrossRef] [Green Version]
- Monath, T.P. Vaccines against diseases transmitted from animals to humans: A one health paradigm. Vaccine 2013, 31, 5321–5338. [Google Scholar] [CrossRef]
- Cunningham, A.A.; Daszak, P.; Wood, J.L.N. One health, emerging infectious diseases and wildlife: Two decades of progress? Philos. Trans. R. Soc. B. Biol. Sci. 2017, 372, 20160167. [Google Scholar] [CrossRef] [Green Version]
- Brochier, B.; Blancou, J.; Thomas, I.; Languet, B.; Artois, M.; Kieny, M.-P.; Lecocq, J.-P.; Costy, F.; Desmettre, P.; Chappuis, G.; et al. Use of recombinant vaccinia-rabies glycoprotein virus for oral vaccination of wildlife against rabies: Innocuity to several non-target bait consuming species. J. Wildl. Dis. 2013, 25, 540–547. [Google Scholar] [CrossRef]
- Williams, S.C.; van Oosterwijk, J.G.; Linske, M.A.; Zatechka, S.; Richer, L.M.; Przybyszewski, C.; Wikel, S.K.; Stafford, K.C. Administration of an orally delivered substrate targeting a mammalian zoonotic pathogen reservoir population: Novel application and biomarker analysis. Vector Borne Zoonotic Dis. 2020. [Google Scholar] [CrossRef]
- Bhattacharya, D.; Bensaci, M.; Luker, K.E.; Luker, G.; Wisdom, S.; Telford, S.R.; Hu, L.T. Development of a baited oral vaccine for use in reservoir-targeted strategies against Lyme disease. Vaccine 2011, 29, 7818–7825. [Google Scholar] [CrossRef] [Green Version]
- Voordouw, M.J.; Tupper, H.; Önder, Ö.; Devevey, G.; Graves, C.J.; Kemps, B.D.; Brisson, D. Reductions in human Lyme disease risk due to the effects of oral vaccination on tick-to-mouse and mouse-to-tick transmission. Vector Borne Zoonotic Dis. 2013, 13, 203–214. [Google Scholar] [CrossRef] [Green Version]
- Bull, J.J.; Smithson, M.W.; Nuismer, S.L. Transmissible viral vaccines. Trends Microbiol. 2018, 26, 6–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polak, K.; Kommedal, A.T. (Eds.) Field Manual for Small Animal Medicine; Wiley: Hoboken, NJ, USA, 2018; ISBN 9781119243274. [Google Scholar]
- Betancur Hurtado, O.J.; Giraldo-Ríos, C. Economic and health impact of the ticks in production animals. In Ticks Tick-Borne Pathogens; IntechOpen: London, UK, 2019. [Google Scholar] [CrossRef] [Green Version]
- White, N.; Sutherst, R.W.; Hall, N.; Whish-Wilson, P. The vulnerability of the Australian beef industry to impacts of the cattle tick (Boophilus microplus) under climate change. Clim. Change 2003, 61, 157–190. [Google Scholar] [CrossRef]
- Hüe, T.; Hurlin, J.C.; Teurlai, M.; Naves, M. Comparison of tick resistance of crossbred Senepol × Limousin to purebred Limousin cattle. Trop. Anim. Health Prod. 2014, 46, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Uttenthal, Å.; Parida, S.; Rasmussen, T.B.; Paton, D.J.; Haas, B.; Dundon, W.G. Strategies for differentiating infection in vaccinated animals (DIVA) for foot-and mouth disease, classical swine fever and avian influenza. Expert Rev. Vaccines 2010, 9, 73–87. [Google Scholar] [CrossRef] [PubMed]
- Farzani, T.A.; Földes, K.; Hanifehnezhad, A.; Ilce, B.Y.; Dagalp, S.B.; Khiabani, N.A.; Ergünay, K.; Alkan, F.; Karaoglu, T.; Bodur, H.; et al. Bovine herpesvirus type 4 (BoHV-4) vector delivering nucleocapsid protein of Crimean-Congo hemorrhagic fever virus induces comparable protective immunity against lethal challenge in IFNα/βγγR-/- mice models. Viruses 2019, 11. [Google Scholar] [CrossRef] [Green Version]
- Ghiasi, S.M.; Salmanian, A.H.; Chinikar, S.; Zakeri, S. Mice orally immunized with a transgenic plant expressing the glycoprotein of Crimean-Congo hemorrhagic fever virus. Clin. Vaccine Immunol. 2011, 18, 2031–2037. [Google Scholar] [CrossRef] [Green Version]
- Donadeu, M.; Nwankpa, N.; Abela-Ridder, B.; Dungu, B. Strategies to increase adoption of animal vaccines by smallholder farmers with focus on neglected diseases and marginalized populations. PLoS Negl. Trop. Dis. 2019, 13, e0006989. [Google Scholar] [CrossRef]
- Abdela, N.; Ibrahim, N.; Begna, F. Prevalence, risk factors and vectors identification of bovine anaplasmosis and babesiosis in and around Jimma town, Southwestern Ethiopia. Acta Tropica 2018, 177, 9–18. [Google Scholar] [CrossRef]
- Latif, A.A.; Walker, A.R. An Introduction to the Biology and Control of Ticks in Africa. ICTTD-2 Project. 2004; 1–29. [Google Scholar]
- Sutherst, R.W.; Norton, G.A.; Barlow, N.D.; Conway, G.R.; Birley, M.; Comins, H.N. An analysis of management strategies for cattle tick (Boophilus microplus) control in Australia. J. Appl. Ecol. 1979, 16, 359. [Google Scholar] [CrossRef]
- Walker, A. Ticks of Domestic Animals in Africa: A Guide to Identification of Species; Bioscience Reports: Edinburgh, Scotland, 2003; ISBN 095451730X. [Google Scholar]
- Nejash, A.A. Review of economically important cattle tick and its control in Ethiopia. Vector Biol. J. 2016, 1. [Google Scholar] [CrossRef] [Green Version]
- Graf, J.F.; Gogolewski, R.; Leach-Bing, N.; Sabatini, G.A.; Molento, M.B.; Bordin, E.L.; Arantes, G.J. Tick control: An industry point of view. Parasitology 2004, 129, S427–S442. [Google Scholar] [CrossRef]
- Eiden, A.L.; Kaufman, P.E.; Oi, F.M.; Dark, M.J.; Bloomquist, J.R.; Miller, R.J. Determination of metabolic resistance mechanisms in pyrethroid-resistant and fipronil-tolerant brown dog ticks. Med. Vet. Entomol. 2017, 31, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Pavela, R.; Canale, A.; Mehlhorn, H.; Benelli, G. Application of ethnobotanical repellents and acaricides in prevention, control and management of livestock ticks: A review. Res. Vet. Sci. 2016, 109, 1–9. [Google Scholar] [CrossRef] [PubMed]
- De Meneghi, D.; Stachurski, F.; Adakal, H. Experiences in tick control by acaricide in the traditional cattle sector in Zambia and Burkina Faso: Possible environmental and public health implications. Front. Public Health 2016, 4, 239. [Google Scholar] [CrossRef] [Green Version]
- Abbas, R.Z.; Zaman, M.A.; Colwell, D.D.; Gilleard, J.; Iqbal, Z. Acaricide resistance in cattle ticks and approaches to its management: The state of play. Vet. Parasitol. 2014, 203, 6–20. [Google Scholar] [CrossRef]
- Nandi, A.; Sagar, S.V.; Chigure, G.; Fular, A.; Sharma, A.K.; Nagar, G.; Kumar, S.; Saravanan, B.C.; Ghosh, S. Determination and validation of discriminating concentration of ivermectin against Rhipicephalus microplus. Vet. Parasitol. 2018, 250, 30–34. [Google Scholar] [CrossRef]
- Pohl, P.C.; Carvalho, D.D.; Daffre, S.; da Silva Vaz, I.; Masuda, A. In vitro establishment of ivermectin-resistant Rhipicephalus microplus cell line and the contribution of ABC transporters on the resistance mechanism. Vet. Parasitol. 2014, 204, 316–322. [Google Scholar] [CrossRef] [Green Version]
- Reck, J.; Klafke, G.M.; Webster, A.; Dall’Agnol, B.; Scheffer, R.; Souza, U.A.; Corassini, V.B.; Vargas, R.; dos Santos, J.S.; de Souza Martins, J.R. First report of fluazuron resistance in Rhipicephalus microplus: A field tick population resistant to six classes of acaricides. Vet. Parasitol. 2014, 201, 128–136. [Google Scholar] [CrossRef]
- Li, A.Y.; Davey, R.B.; Miller, R.J.; George, J.E. Detection and characterization of amitraz resistance in the southern cattle tick, Boophilus microplus (Acari: Ixodidae). J. Med. Entomol. 2004, 41, 193–200. [Google Scholar] [CrossRef] [Green Version]
- Kunz, S.E.; Kemp, D.H. Insecticides and acaricides: Resistance and environmental impact. Rev. Sci. Tech. 1994, 13, 1249–1286. [Google Scholar] [CrossRef]
- Saramago, L.; Gomes, H.; Aguilera, E.; Cerecetto, H.; González, M.; Cabrera, M.; Alzugaray, M.F.; Da Silva Vaz, I., Jr.; Nunes da Fonseca, R.; Aguirre-López, B.; et al. Novel and selective Rhipicephalus microplus triosephosphate isomerase inhibitors with acaricidal activity. Vet. Sci. 2018, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adenubi, O.T.; McGaw, L.J.; Eloff, J.N.; Naidoo, V. In vitro bioassays used in evaluating plant extracts for tick repellent and acaricidal properties: A critical review. Vet. Parasitol. 2018, 254, 160–171. [Google Scholar] [CrossRef] [PubMed]
- Adenubi, O.T.; Fasina, F.O.; McGaw, L.J.; Eloff, J.N.; Naidoo, V. Plant extracts to control ticks of veterinary and medical importance: A review. S. Afr. J. Botany 2016, 105, 178–193. [Google Scholar] [CrossRef]
- Mulla, M.S.; Su, T. Activity and biological effects of neem products against arthropods of medical and veterinary importance. J. Am. Mosq. Control Assoc. 1999, 15, 133–152. [Google Scholar] [PubMed]
- Heimerdinger, A.; Olivo, C.J.; Molento, M.B.; Agnolin, C.A.; Ziech, M.F.; Scaravelli, L.F.B.; Skonieski, F.R.; Both, J.F.; Charão, P.S. Alcoholic extract of lemongrass (Cymbopogon citratus) on the control of Boophilus microplus in cattle. Rev. Bras. Parasitol. Vet. 2006, 15, 37–39. [Google Scholar]
- Samish, M.; Alekseev, E. Arthropods as predators of ticks (Ixodoidea). J. Med. Entomol. 2001, 38, 1–11. [Google Scholar] [CrossRef]
- Duffy, D.C.; Downer, R.; Brinkley, C. The effectiveness of Helmeted Guineafowl in the control of the deer tick, the vector of Lyme disease. Wilson Bull. 1992, 104, 342–345. [Google Scholar]
- Hasle, G. Transport of ixodid ticks and tick-borne pathogens by migratory birds. Front. Cell. Infect. Microbiol. 2013, 3, 48. [Google Scholar] [CrossRef] [Green Version]
- Sparagano, O.; George, D.; Giangaspero, A.; Špitalská, E. Arthropods and associated arthropod-borne diseases transmitted by migrating birds. The case of ticks and tick-borne pathogens. Vet. Parasitol. 2015, 213, 61–66. [Google Scholar] [CrossRef]
- Samish, M.; Ginsberg, H.; Glazer, I. Biological control of ticks. Parasitology 2004, 129, S389–S403. [Google Scholar] [CrossRef] [PubMed]
- Assenga, S.P.; You, M.; Shy, C.H.; Yamagishi, J.; Sakaguchi, T.; Zhou, J.; Kibe, M.K.; Xuan, X.; Fujisaki, K. The use of a recombinant baculovirus expressing a chitinase from the hard tick Haemaphysalis longicornis and its potential application as a bioacaricide for tick control. Parasitol. Res. 2006, 98, 111–118. [Google Scholar] [CrossRef]
- Fernandes, É.K.K.; Bittencourt, V.R.E.P.; Roberts, D.W. Perspectives on the potential of entomopathogenic fungi in biological control of ticks. Experimental Parasitol. 2012, 130, 300–305. [Google Scholar] [CrossRef] [PubMed]
- Samish, M.; Ginsberg, H.; Glazer, I. Anti-tick biological control agents: Assessment and future perspectives. In Ticks: Biology, Disease and Control; Cambridge University Press: Cambridge, UK, 2008; pp. 447–469. ISBN 9780511551802. [Google Scholar]
- Santi, L.; e Silva, L.A.D.; da Silva, W.O.B.; Corrêa, A.P.F.; Rangel, D.E.N.; Carlini, C.R.; Schrank, A.; Vainstein, M.H. Virulence of the entomopathogenic fungus Metarhizium anisopliae using soybean oil formulation for control of the cotton stainer bug, Dysdercus peruvianus. World J. Microbiol. Biotechnol. 2011, 27, 2297–2303. [Google Scholar] [CrossRef]
- Quesada-Moraga, E.; Santos-Quirós, R.; Valverde-García, P.; Santiago-Álvarez, C. Virulence, horizontal transmission, and sublethal reproductive effects of Metarhizium anisopliae (anamorphic fungi) on the German cockroach (Blattodea: Blattellidae). J. Invertebr. Pathol. 2004, 87, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Stafford, K.C.; Allan, S.A. Field applications of entomopathogenic fungi Beauveria bassiana and Metarhizium anisopliae F52 (Hypocreales: Clavicipitaceae) for the control of Ixodes scapularis (Acari: Ixodidae). J. Med. Entomol. 2010, 47, 1107–1115. [Google Scholar] [CrossRef] [Green Version]
- Samish, M.; Gindin, G.; Alekseev, E.; Glazer, I. Pathogenicity of entomopathogenuc fungi to different developmental states of Rhipicephalus sanguineus (Acari: Ixodidae). J. Parasitol. 2001, 87, 1355–1359. [Google Scholar] [CrossRef]
- Frazzon, A.P.G.; Vaz Junior, I.D.S.; Masuda, A.; Schrank, A.; Vainstein, M.H. In vitro assessment of Metarhizium anisopliae isolates to control the cattle tick Boophilus microplus. Vet. Parasitol. 2000, 94, 117–125. [Google Scholar] [CrossRef]
- Leemon, D.M.; Turner, L.B.; Jonsson, N.N. Pen studies on the control of cattle tick (Rhipicephalus (Boophilus) microplus) with Metarhizium anisopliae (Sorokin). Vet. Parasitol. 2008, 156, 248–260. [Google Scholar] [CrossRef]
- Suleiman, E.A.; Shigidi, M.T.; Hassan, S.M. Metarhizium anisopliae as a biological control agent against Hyalomma anatolicum (Acari: Ixodidae). Pak. J. Biolog. Sci. 2013, 16, 1943–1949. [Google Scholar] [CrossRef]
- Rodríguez-Mallon, A. Developing anti-tick vaccines. In Methods in Molecular Biology; Humana Press Inc.: New York, NY, USA, 2016; Volume 1404, pp. 243–259. [Google Scholar]
- De la Fuente, J.; Almazán, C.; Blas-Machado, U.; Naranjo, V.; Mangold, A.J.; Blouin, E.F.; Gortazar, C.; Kocan, K.M. The tick protective antigen, 4D8, is a conserved protein involved in modulation of tick blood ingestion and reproduction. Vaccine 2006, 24, 4082–4095. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Parameter | Key Considerations and Questions to Address Gaps in Knowledge of CCHFV |
|---|---|
| Tick distribution |
|
| Host distribution |
|
| Tick density |
|
| Host density |
|
| Virus prevalence in nature |
|
| Infection intensity * |
|
| Virus prevalence at slaughter |
|
| Biting rate (immatures) |
|
| Virus survival in blood and tissues of livestock |
|
| Tick survival in nature |
|
| Human behavior |
|
| Biting rate (adults) |
|
| Dose and route of exposure |
|
| Physical and intrinsic barriers (e.g., skin) |
|
| Host tissue susceptibility and permissiveness |
|
| Immune response |
|
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sorvillo, T.E.; Rodriguez, S.E.; Hudson, P.; Carey, M.; Rodriguez, L.L.; Spiropoulou, C.F.; Bird, B.H.; Spengler, J.R.; Bente, D.A. Towards a Sustainable One Health Approach to Crimean–Congo Hemorrhagic Fever Prevention: Focus Areas and Gaps in Knowledge. Trop. Med. Infect. Dis. 2020, 5, 113. https://doi.org/10.3390/tropicalmed5030113
Sorvillo TE, Rodriguez SE, Hudson P, Carey M, Rodriguez LL, Spiropoulou CF, Bird BH, Spengler JR, Bente DA. Towards a Sustainable One Health Approach to Crimean–Congo Hemorrhagic Fever Prevention: Focus Areas and Gaps in Knowledge. Tropical Medicine and Infectious Disease. 2020; 5(3):113. https://doi.org/10.3390/tropicalmed5030113
Chicago/Turabian StyleSorvillo, Teresa E., Sergio E. Rodriguez, Peter Hudson, Megan Carey, Luis L. Rodriguez, Christina F. Spiropoulou, Brian H. Bird, Jessica R. Spengler, and Dennis A. Bente. 2020. "Towards a Sustainable One Health Approach to Crimean–Congo Hemorrhagic Fever Prevention: Focus Areas and Gaps in Knowledge" Tropical Medicine and Infectious Disease 5, no. 3: 113. https://doi.org/10.3390/tropicalmed5030113