Characterization of vB_Kpn_F48, a Newly Discovered Lytic Bacteriophage for Klebsiella pneumoniae of Sequence Type 101

 ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Host and Culture Conditions
2.2. Phage Isolation and Purification
2.3. Transmission Electron Microscopy (TEM)
2.4. Determination of Bacteriophage Host Range
2.5. Preparation of High Titer Bacteriophage Suspensions
2.6. Efficiency of Plating (EOP)
2.7. One-Step Growth Curve
2.8. Influence of Physical Agents on Phage Viability
2.9. Time-Kill Assay
2.10. Rate of Appearance of Phage-Resistant Mutants
2.11. Extraction of Bacteriophage vB_Kpn_F48 DNA
2.12. Genome Sequencing
2.13. Bioinformatics Analysis of Phage Genome
3. Results
3.1. Phage Isolation and Morphological Characterization
3.2. Phage Host Range
3.3. EOP of Phage vB_Kpn_F48
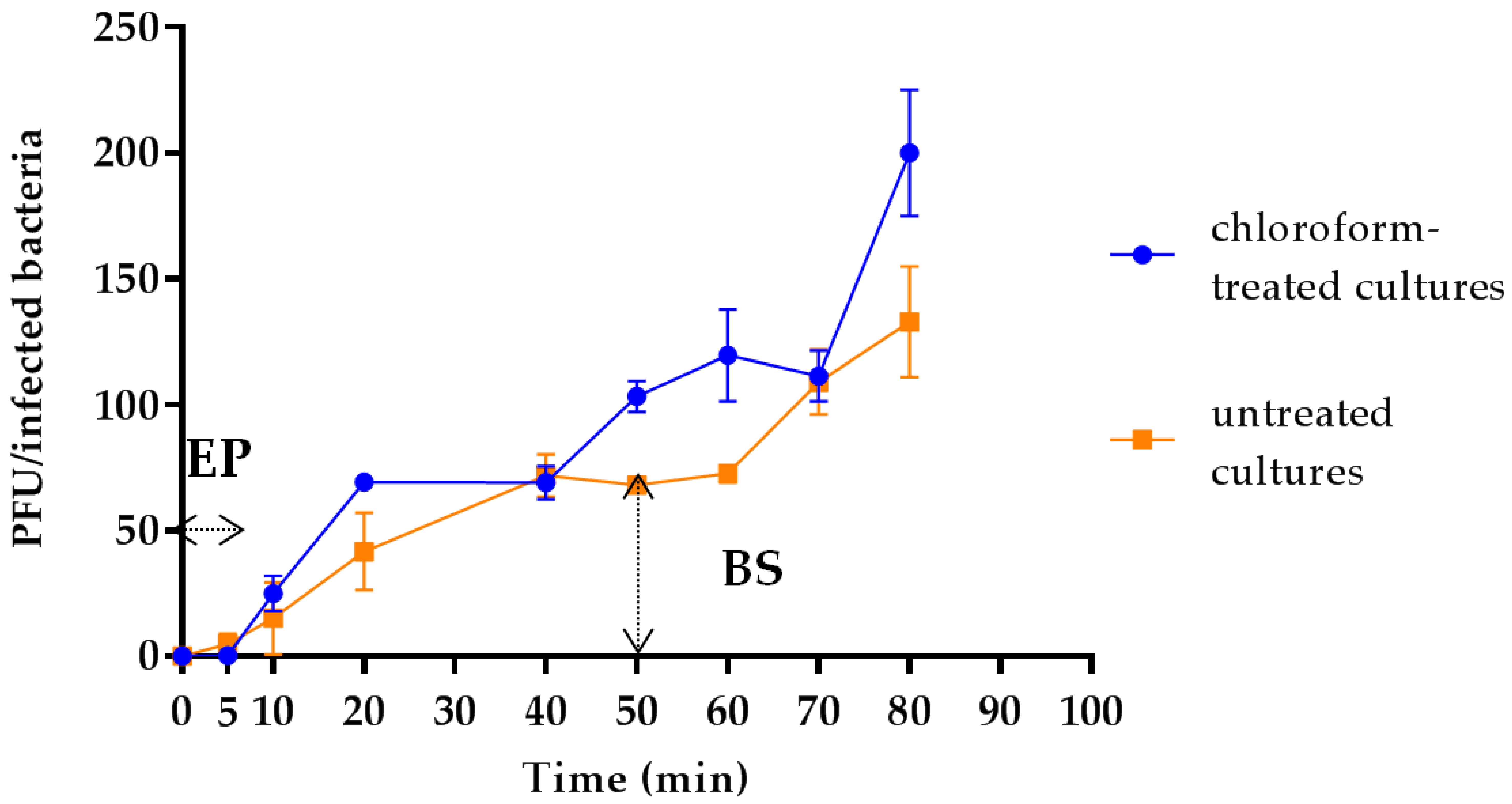
3.4. Latency Period, Eclipse Period and Burst Size Determination
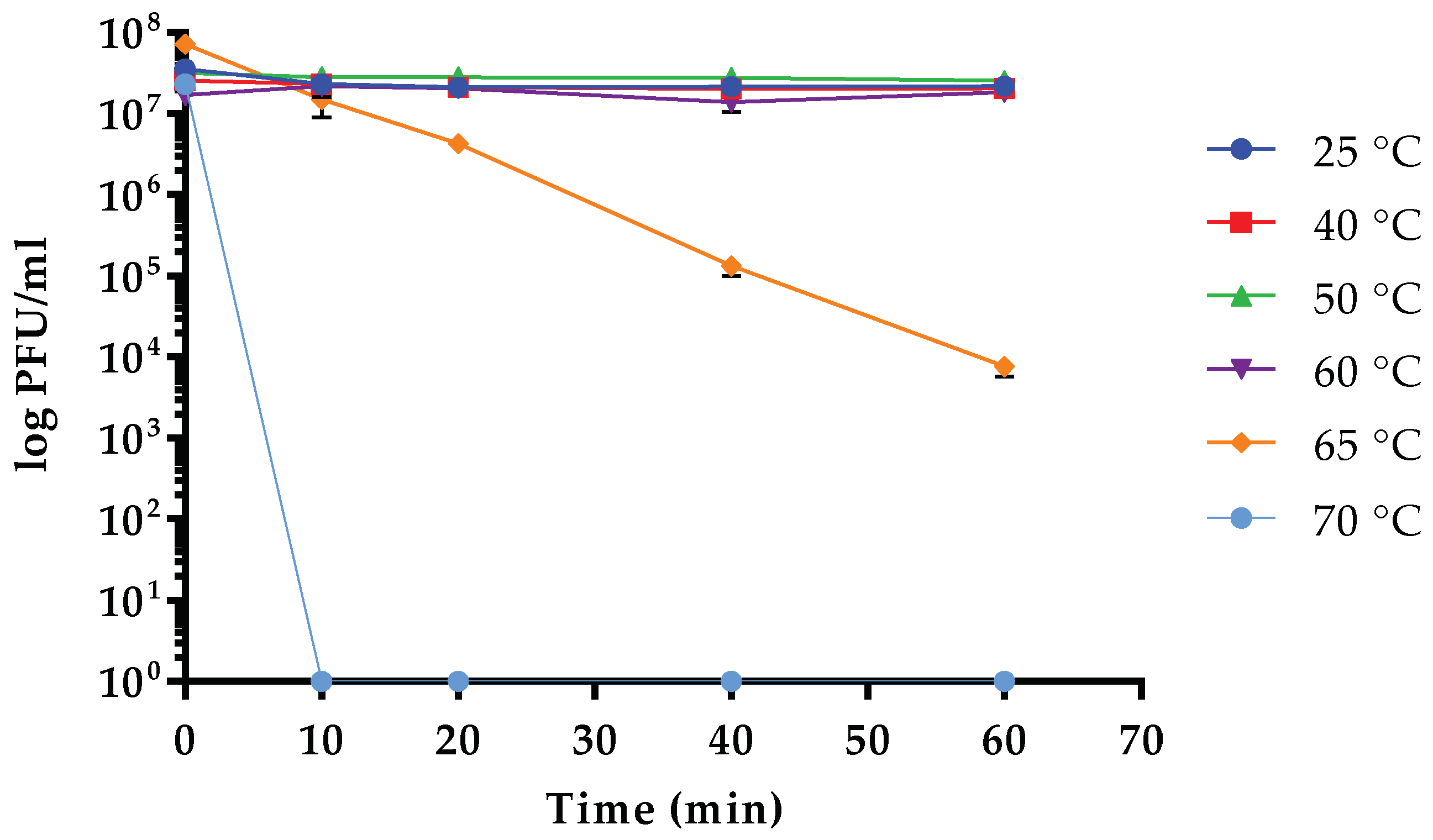
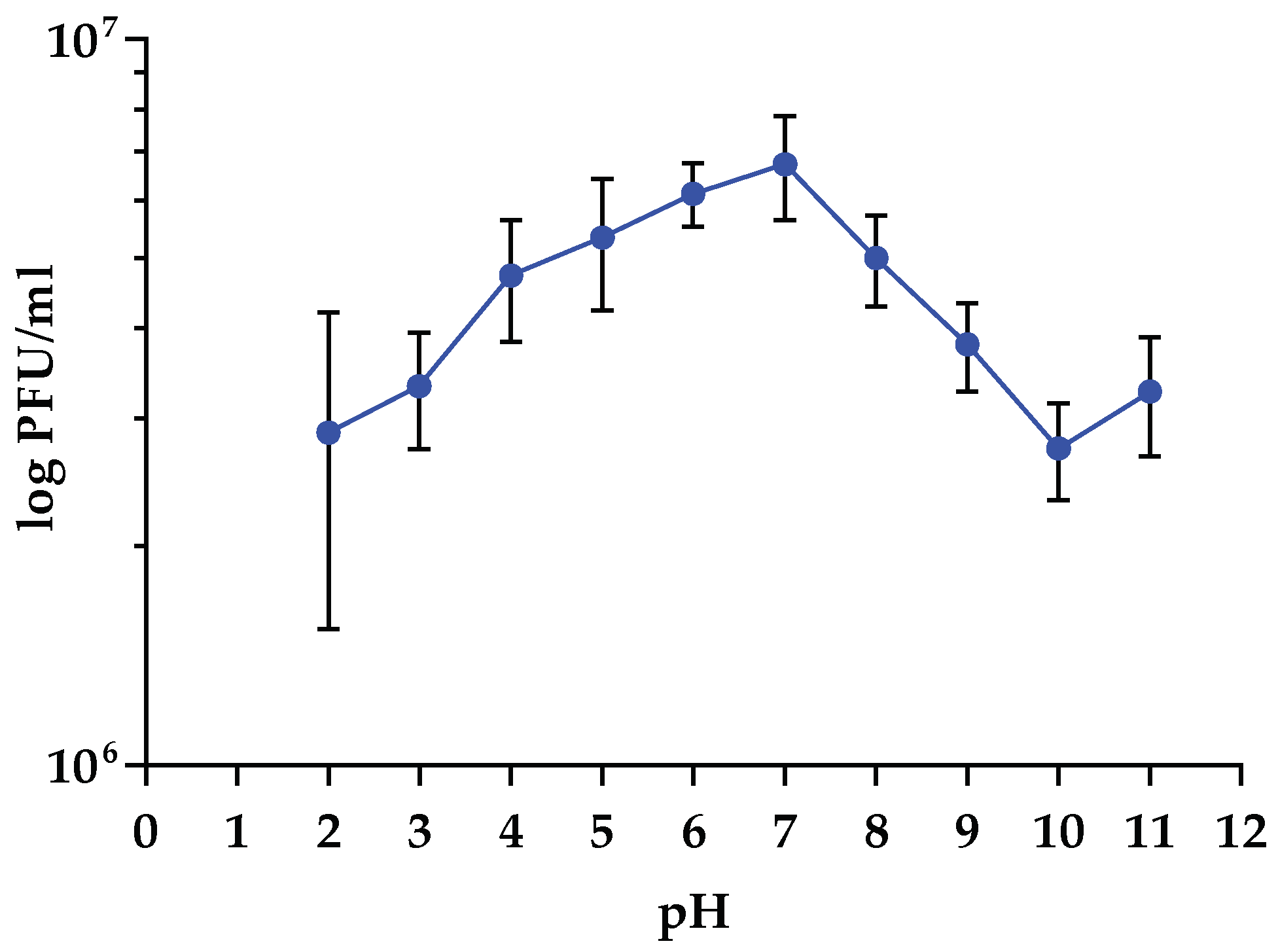
3.5. Sensitivity to Physical Parameters
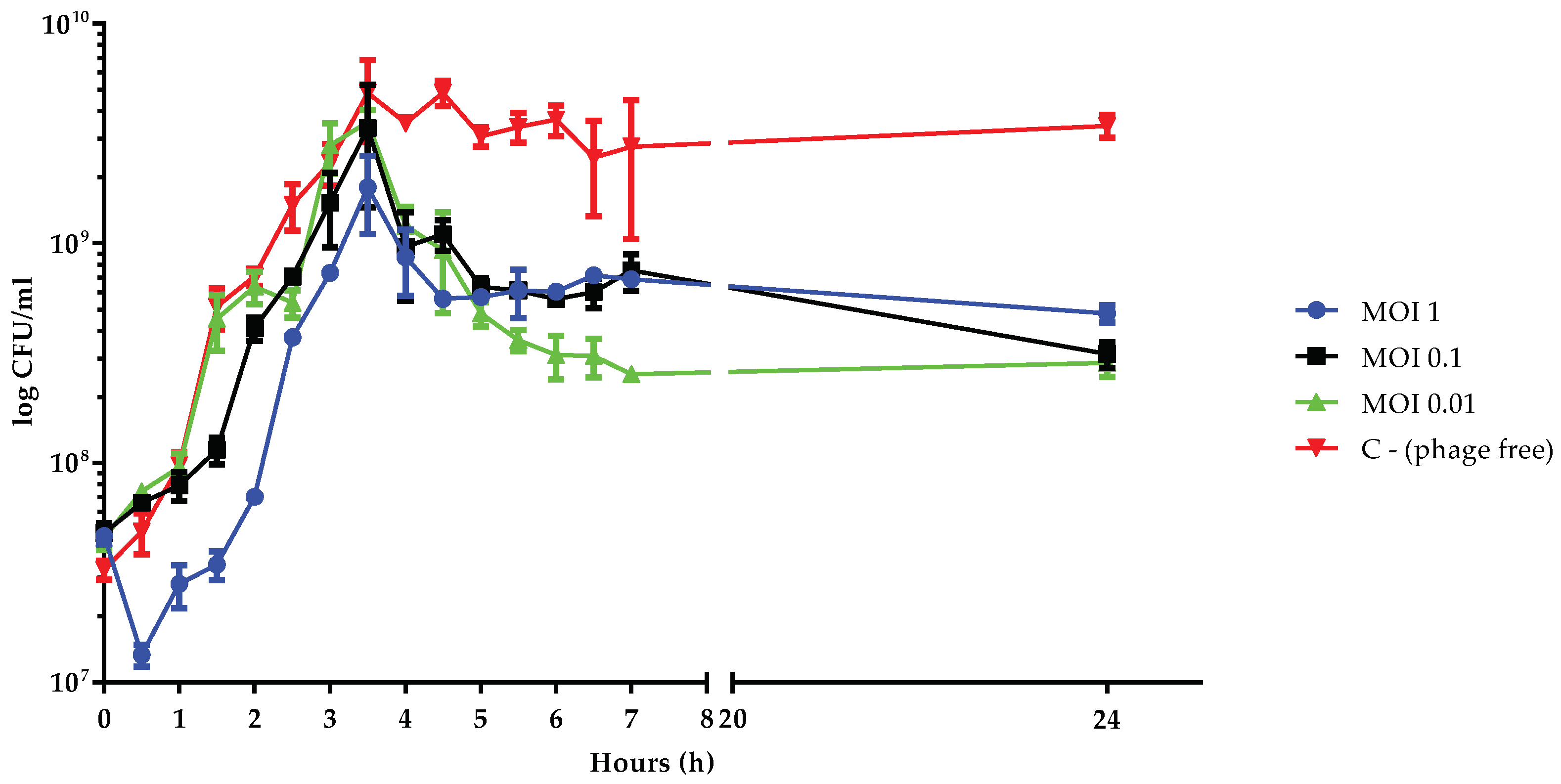
3.6. Killing Dynamic of vB_Kpn_F48 against the 12C47 Strain
3.7. Frequency of Emergence of Phage-Resistant Mutants
3.8. Genome Analysis
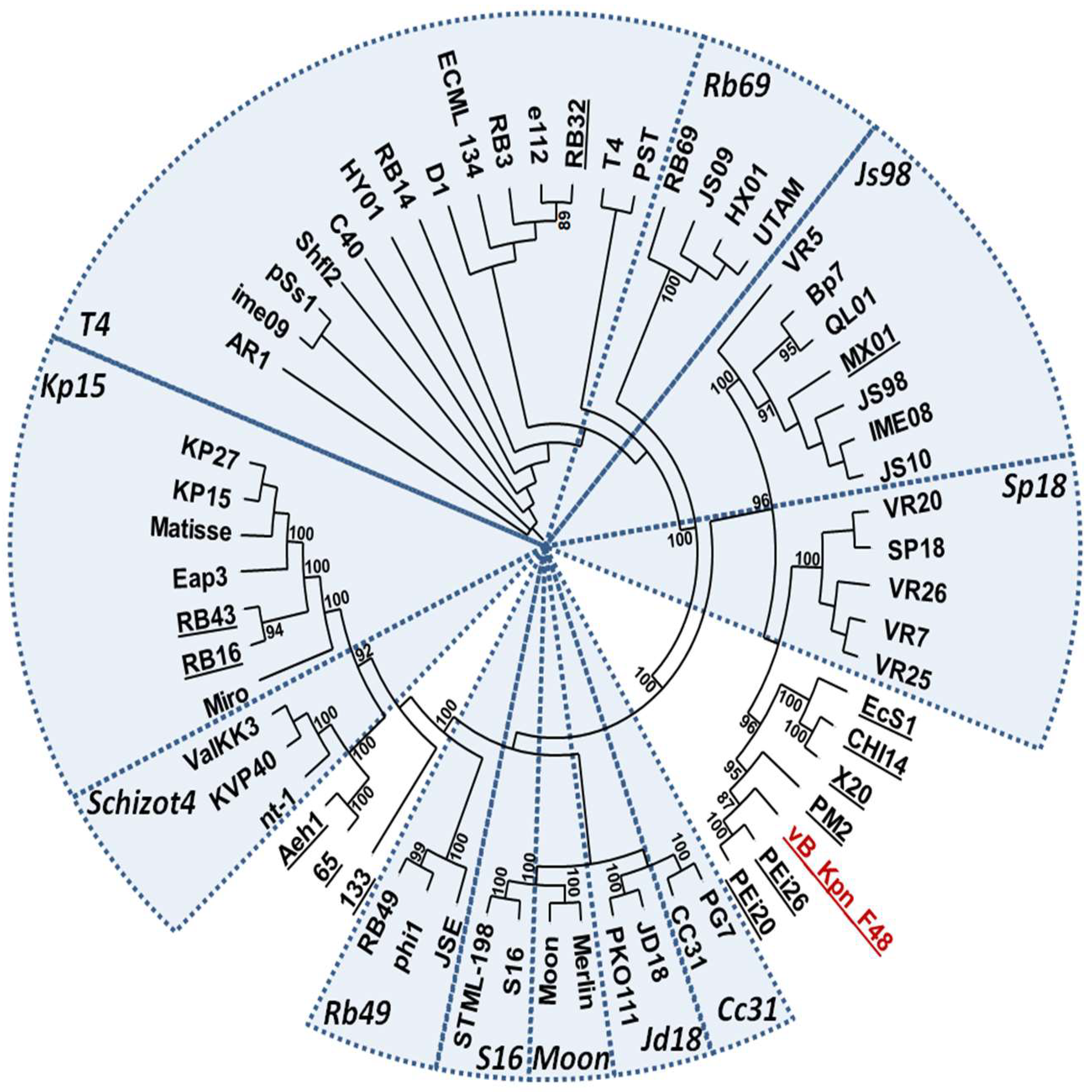
3.9. Phylogenetic Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rossolini, G.M.; Arena, F.; Pecile, P.; Pollini, S. Update on the antibiotic resistance crisis. Clin. Opin. Pharmacol. 2014, 18, 56–60. [Google Scholar] [CrossRef] [PubMed]
- Bush, K. Alarming β-lactamase-mediated resistance in multidrug-resistant Enterobacteriaceae. Curr. Opin. Microbiol. 2010, 13, 558–564. [Google Scholar] [CrossRef] [PubMed]
- Michael, C.A.; Dominey-Howes, D.; Labbate, M. The antimicrobial resistance crisis: Causes, consequences, and management. Front. Public Health 2014, 2, 145. [Google Scholar] [CrossRef] [PubMed]
- Prestinaci, F.; Pezzotti, P.; Pantosti, A. Antimicrobial resistance: A global multifaceted phenomenon. Pathog. Glob. Health. 2015, 109, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Navon-Venezia, S.; Kondratyeva, K.; Carattoli, A. Klebsiella pneumoniae: A major worldwide source and shuttle for antibiotic resistance. FEMS Microbiol. Rev. 2017, 41, 252–275. [Google Scholar] [CrossRef] [PubMed]
- Mammina, C.; Bonura, C.; Aleo, A.; Fasciana, T.; Brunelli, T.; Pesavento, G.; Degl’Innocenti, R.; Nastasi, A. Sequence type 101 (ST101) as the predominant carbapenem-non-susceptible Klebsiella pneumoniae clone in an acute general hospital in Italy. Int. J. Antimicrob. Agents. 2012, 39, 543–545. [Google Scholar] [CrossRef] [PubMed]
- Bonura, C.; Giuffrè, M.; Aleo, A.; Fasciana, T.; Di Bernardo, F.; Stampone, T.; Giammanco, A.; MDR-GN Working Group; Palma, D.M.; Mammina, C. An update of the evolving epidemic of blaKPC carrying Klebsiella pneumoniae in Sicily, Italy, 2014: Emergence of multiple non-ST258 clones. PLoS ONE 2015, 10, e0132936. [Google Scholar] [CrossRef] [PubMed]
- Conte, V.; Monaco, M.; Giani, T.; D’Ancona, F.; Moro, M.L.; Arena, F.; D’Andrea, M.M.; Rossolini, G.M.; Pantosti, A.; AR-ISS Study Group on Carbapenemase-Producing K. pneumoniae. Molecular epidemiology of KPC-producing Klebsiella pneumoniae from invasive infections in Italy: Increasing diversity with predominance of the ST512 clade II sublineage. J. Antimicrob. Chemother. 2016, 71, 3386–3391. [Google Scholar] [CrossRef] [PubMed]
- Mshana, S.E.; Fritzenwanker, M.; Falgenhauer, L.; Domann, E.; Hain, T.; Chakraborty, T.; Imirzalioglu, C. Molecular epidemiology and characterization of an outbreak causing Klebsiella pneumoniae clone carrying chromosomally located blaCTX-M-15 at a German University-Hospital. BMC Microbiol. 2015, 15, 122. [Google Scholar] [CrossRef] [PubMed]
- Skalova, A.; Chudejova, K.; Rotova, V.; Medvecky, M.; Studentova, V.; Chudackova, E.; Lavicka, P.; Bergerova, T.; Jakubu, V.; Zemlickova, H.; et al. Molecular Characterization of OXA-48-Like-Producing Enterobacteriaceae in the Czech Republic and Evidence for Horizontal Transfer of pOXA-48-Like Plasmids. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar]
- Oteo, J.; Pérez-Vázquez, M.; Bautista, V.; Ortega, A.; Zamarrón, P.; Saez, D.; Fernández-Romero, S.; Lara, N.; Ramiro, R.; Aracil, B.; et al. The spread of KPC-producing Enterobacteriaceae in Spain: WGS analysis of the emerging high-risk clones of Klebsiella pneumoniae ST11/KPC-2, ST101/KPC-2 and ST512/KPC-3. J. Antimicrob. Chemother. 2016, 71, 3392–3399. [Google Scholar] [CrossRef] [PubMed]
- Loucif, L.; Kassah-Laouar, A.; Saidi, M.; Messala, A.; Chelaghma, W.; Rolain, J.M. Outbreak of OXA-48-producing Klebsiella pneumoniae involving a Sequence Type 101 clone in Batna University Hospital, Algeria. Antimicrob. Agents Chemother. 2016, 60, 7494–7497. [Google Scholar] [PubMed]
- Low, Y.M.; Yap, P.S.; Abdul Jabar, K.; Ponnampalavanar, S.; Karunakaran, R.; Velayuthan, R.; Chong, C.W.; Abu Bakar, S.; Md Yusof, M.Y.; Teh, C.S. The emergence of carbapenem resistant Klebsiella pneumoniae in Malaysia: Correlation between microbiological trends with host characteristics and clinical factors. Antimicrob. Resist. Infect. Control 2017, 6, 5. [Google Scholar] [CrossRef] [PubMed]
- De Laveleye, M.; Huang, T.D.; Bogaerts, P.; Berhin, C.; Bauraing, C.; Sacré, P.; Noel, A.; Glupczynski, Y.; multicenter study group. Increasing incidence of carbapenemase-producing Escherichia coli and Klebsiella pneumoniae in Belgian hospitals. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Del Franco, M.; Paone, L.; Novati, R.; Giacomazzi, C.G.; Bagattini, M.; Galotto, C.; Montanera, P.G.; Triassi, M.; Zarrilli, R. Molecular epidemiology of carbapenem resistant Enterobacteriaceae in Valle d’Aosta region, Italy, shows the emergence of KPC-2 producing Klebsiella pneumoniae clonal complex 101 (ST101 and ST1789). BMC Microbiol. 2015, 15, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Poulou, A.; Voulgari, E.; Vrioni, G.; Koumaki, V.; Xidopoulos, G.; Chatzipantazi, V.; Markou, F.; Tsakris, A. Outbreak caused by an ertapenem-resistant, CTX-M-15-producing Klebsiella pneumoniae sequence type 101 clone carrying an OmpK36 porin variant. J. Clin. Microbiol. 2013, 51, 3176–3182. [Google Scholar] [CrossRef] [PubMed]
- Potron, A.; Poirel, L.; Rondinaud, E.; Nordmann, P. Intercontinental spread of OXA-48 β-lactamase-producing Enterobacteriaceae over a 11-year period, 2001 to 2011. Eurosurveillance 2013, 18, 20549. [Google Scholar] [CrossRef] [PubMed]
- Jayol, A.; Poirel, L.; Dortet, L.; Nordmann, P. National survey of colistin resistance among carbapenemase-producing Enterobacteriaceae and outbreak caused by colistin-resistant OXA-48-producing Klebsiella pneumoniae, France, 2014. Eurosurveillance 2016, 21, 30339. [Google Scholar] [CrossRef] [PubMed]
- Pitart, C.; Solé, M.; Roca, I.; Fàbrega, A.; Vila, J.; Marco, F. First outbreak of a plasmid-mediated carbapenem-hydrolyzing OXA-48 beta-lactamase in Klebsiella pneumoniae in Spain. Antimicrob. Agents Chemother. 2011, 55, 4398–4401. [Google Scholar] [CrossRef] [PubMed]
- Pires, D.P.; Cleto, S.; Sillankorva, S.; Azeredo, J.; Lu, T.K. Genetically engineered phages: A review of advances over the last decade. Microbiol. Mol. Biol. Rev. 2016, 80, 523–543. [Google Scholar] [CrossRef] [PubMed]
- Domingo-Calap, P.; Georgel, P.; Bahram, S. Back to the future: Bacteriophages as promising therapeutic tools. HLA 2016, 87, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Giani, T.; Pini, B.; Arena, F.; Conte, V.; Bracco, S.; Migliavacca, R.; AMCLI-CRE Survey Participants; Pantosti, A.; Pagani, L.; Luzzaro, F.; Rossolini, G.M. Epidemic diffusion of KPC carbapenemase-producing Klebsiella pneumoniae in Italy: Results of the first countrywide survey, 15 May to 30 June 2011. Eurosurveillance 2013, 18, 20489. [Google Scholar] [PubMed]
- Adams, M. Bacteriophage; Interscience Publishers: New York, NY, USA, 1959; pp. 137–159. [Google Scholar]
- Di Lallo, G.; Evangelisti, M.; Mancuso, F.; Ferrante, P.; Marcelletti, S.; Tinari, A.; Superti, F.; Migliore, L.; D’Addabbo, P.; Frezza, D.; et al. Isolation and partial characterization of bacteriophages infecting Pseudomonas syringae pv. actinidiae, causal agent of kiwifruit bacterial canker. J. Basic Microbiol. 2014, 54, 1210–1221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, C.R.; Lin, T.L.; Pan, Y.J.; Hsieh, P.F.; Wang, J.T. Isolation of a bacteriophage specific for a new capsular type of Klebsiella pneumoniae and characterization of its polysaccharide depolymerase. PLoS ONE 2013, 8, e70092. [Google Scholar] [CrossRef] [PubMed]
- Brisse, S.; Passet, V.; Haugaard, A.B.; Babosan, A.; Kassis-Chikhani, N.; Struve, C.; Decré, D. wzi Gene sequencing, a rapid method for determination of capsular type for Klebsiella strains. J. Clin. Microbiol. 2013, 51, 4073–4078. [Google Scholar] [CrossRef] [PubMed]
- Khan Mirzaei, M.; Nilsson, A.S. Isolation of phages for phage therapy: A comparison of spot tests and ffficiency of plating analyses for determination of host range and efficacy. PLoS ONE 2015, 10, e0118557. [Google Scholar] [CrossRef] [PubMed]
- D’Andrea, M.M.; Marmo, P.; Henrici De Angelis, L.; Palmieri, M.; Ciacci, N.; Di Lallo, G.; Demattè, E.; Vannuccini, E.; Lupetti, P.; Rossolini, G.M.; et al. φBO1E, a newly discovered lytic bacteriophage targeting carbapenemase-producing Klebsiella pneumoniae of the pandemic Clonal Group 258 clade II lineage. Sci. Rep. 2017, 7, 2614. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.H. Bacteriophage; Interscience Publishers: New York, NY, USA, 1959; pp. 450–456. [Google Scholar]
- Clinical and Laboratory Standards Institute. Methods for Determining Bactericidal Activity of Antimicrobial Agents: Approved Guidelines; M26-A; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 1999. [Google Scholar]
- Murray, P.; Baron, E.; Jorgensen, J.; Pfaller, M.; Yolken, R. Manual of Clinical Microbiology, 8th ed.; American Society for Microbiology: Washington, DC, USA, 2003. [Google Scholar]
- Merabishvili, M.; Vandenheuvel, D.; Kropinski, A.M.; Mast, J.; de Vos, D.; Verbeken, G.; Noben, J.P.; Lavigne, R.; Vaneechoutte, M.; Pirnay, J.P. Characterization of Newly Isolated Lytic Bacteriophages Active against Acinetobacter baumannii. PLoS ONE 2014, 9, e104853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gill, J.J. Phage Genomic DNA Extraction. OpenWetWare, 2015. Available online: https://openwetware.org/wiki/Gill:Phage_genomic_DNA_extraction (accessed on 31 May 2017).
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [PubMed]
- Lowe, T.M.; Chan, P.P. tRNAscan-SE On-line: Integrating search and context for analysis of transfer RNA genes. Nucleic Acids Res. 2016, 44, W54–W57. [Google Scholar] [CrossRef] [PubMed]
- Laslett, D.; Canback, B. ARAGORN, a program to detect tRNA genes and tmRNA genes in nucleotide sequences. Nucleic Acids Res. 2004, 32, 11–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchler-Bauer, A.; Derbyshire, M.K.; Gonzales, N.R.; Lu, S.; Chitsaz, F.; Geer, L.Y.; Geer, R.C.; He, J.; Gwadz, M.; Hurwitz, D.I.; et al. CDD: NCBI’s conserved domain database. Nucleic Acids Res. 2015, 43, D222–D226. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meier-Kolthoff, J.P.; Göker, M. VICTOR: Genome-based phylogeny and classification of prokaryotic viruses. Bioinformatics 2017, 33, 3396–3404. [Google Scholar] [CrossRef] [PubMed]
- McNair, K.; Bailey, B.A.; Edwards, R.A. PHACTS, a computational approach to classifying the lifestyle of phages. Bioinformatics 2012, 28, 614–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giani, T.; Arena, F.; Vaggelli, G.; Conte, V.; Chiarelli, A.; Henrici De Angelis, L.; Fornaini, R.; Grazzini, M.; Niccolini, F.; Pecile, P.; et al. Large Nosocomial Outbreak of Colistin-Resistant, Carbapenemase-Producing Klebsiella pneumoniae Traced to Clonal Expansion of an mgrB Deletion Mutant. J. Clin. Microbiol. 2015, 53, 3341–3344. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Mathema, B.; Pitout, J.D.D.; DeLeo, F.R.; Kreiswirth, B.N. Epidemic Klebsiella pneumoniae ST258 Is a Hybrid Strain. Mbio 2014, 5, e01355-14. [Google Scholar] [CrossRef] [PubMed]
- D’Andrea, M.M.; Amisano, F.; Giani, T.; Conte, V.; Ciacci, N.; Ambretti, S.; Santoriello, L.; Rossolini, G.M. Diversity of capsular polysaccharide gene clusters in KPC-producing Klebsiella pneumoniae clinical isolates of Sequence Type 258 involved in the Italian epidemic. PLoS ONE 2014, 9, e96827. [Google Scholar]
- Hyman, P.; Abedon, S.T. Bacteriophage host range and bacterial resistance. Adv. Appl. Microbiol. 2010, 70, 217–248. [Google Scholar] [PubMed]
- Viazis, S.; Akhtar, M.; Feirtag, J.; Brabban, A.D.; Diez-Gonzalez, F. Isolation and characterization of lytic bacteriophages against enterohaemorrhagic Escherichia coli. Appl. Microbiol. 2011, 110, 1323–1331. [Google Scholar] [CrossRef] [PubMed]
- Henry, M.; Biswas, B.; Vincent, L.; Mokashi, V.; Schuch, R.; Bishop-Lilly, K.A.; Sozhamannan, S. Development of a high throughput assay for indirectly measuring phage growth using the OmniLog™ system. Bacteriophage 2012, 2, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Luria, S.E.; Delbrück, M. Mutations of bacteria from virus sensitivity to virus resistance. Genetics 1943, 28, 491–511. [Google Scholar] [PubMed]
- Zhang, C.; Liu, W.; Ren, H. Complete Genome Sequence of Bp7, an Escherichia coli Bacteriophage with a Wide Host Range. J. Virol. 2012, 86, 13832. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Li, W.; Liu, W.; Zou, L.; Yan, C.; Lu, K.; Ren, H. T4-Like Phage Bp7, a Potential Antimicrobial Agent for Controlling Drug-Resistant Escherichia coli in Chickens. Appl. Environ. Microbiol. 2013, 79, 5559–5565. [Google Scholar] [CrossRef] [PubMed]
- Melegh, S.; Schneider, G.; Horváth, M.; Jakab, F.; Emődy, L.; Tigyi, Z. Identification and characterization of CTX-M-15 producing Klebsiella pneumoniae clone ST101 in a Hungarian university teaching hospital. Acta Microbiol. Immunol. Hung. 2015, 62, 233–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, J.D.C.; Moellering, R.C., Jr. Antimicrobial agents. In Manual of Clinical Microbiology, 7th ed.; Murray, P.R., Baron, E.J., Pfaller, M.A., Tenover, F.C., Yolken, R.H., Eds.; American Society for Microbiology: Washington, DC, USA, 1995; pp. 1474–1504. [Google Scholar]
- Pirnay, J.P.; Blasdel, B.G.; Bretaudeau, L.; Buckling, A.; Chanishvili, N.; Clark, J.R.; Corte-Real, S.; Debarbieux, L.; Dublanchet, A.; De Vos, D.; et al. Quality and Safety Requirements for Sustainable Phage Therapy Products. Pharm. Res. 2015, 32, 2173–2179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pirnay, J.P.; De Vos, D.; Verbeken, G.; Merabishvili, M.; Chanishvili, N.; Vaneechoutte, M.; Zizi, M.; Laire, G.; Lavigne, R.; Huys, I.; et al. The phage therapy paradigm: Prêt-à-porter or sur-mesure? Pharm. Res. 2011, 28, 934–937. [Google Scholar] [CrossRef] [PubMed]
- Montag, D.; Hashemolhosseini, S.; Henning, U. Receptor-recognizing proteins of T-even type bacteriophages. The receptor-recognizing area of proteins 37 of phages T4 TuIa and TuIb. J. Mol. Biol. 1990, 216, 327–334. [Google Scholar] [CrossRef]
- Chen, Z.; Schneider, T.D. Information theory based T7-like promoter models: Classification of bacteriophages and differential evolution of promoters and their polymerases. Nucleic Acids Res. 2005, 33, 6172–6187. [Google Scholar] [CrossRef] [PubMed]
- Adriaenssens, E.M.; Ceyssens, P.J.; Dunon, V.; Ackermann, H.W.; Van Vaerenbergh, J.; Maes, M.; de Proft, M.; Lavigne, R. Bacteriophages LIMElight and LIMEzero of Pantoea agglomerans, Belonging to the “phiKMV-Like Viruses”. Appl. Environ. Microbiol. 2011, 77, 3443–3450. [Google Scholar] [CrossRef] [PubMed]
- Ackermann, H.W.; Krisch, H.M.; Comeau, A.M. Morphology and genome sequence of phage ϕ1402: A dwarf myovirus of the predatory bacterium Bdellovibrio bacteriovorus. Bacteriophage 2011, 1, 138–142. [Google Scholar] [CrossRef] [PubMed]
- Cheepudom, J.; Lee, C.C.; Cai, B.; Meng, M. Isolation, characterization, and complete genome analysis of P1312, a thermostable bacteriophage that infects Thermobifida fusca. Front. Microbiol. 2015, 6, 959. [Google Scholar] [CrossRef] [PubMed]
- Kęsik-Szeloch, A.; Drulis-Kawa, Z.; Weber-Dąbrowska, B.; Kassner, J.; Majkowska-Skrobek, G.; Augustyniak, D.; Lusiak-Szelachowska, M.; Zaczek, M.; Górski, A.; Kropinski, A.M. Characterising the Biology of Novel Lytic Bacteriophages Infecting Multidrug Resistant Klebsiella pneumoniae. Virol. J. 2013, 10, 100. [Google Scholar] [CrossRef] [PubMed]
- Philipson, C.W.; Voegtly, L.J.; Lueder, M.R.; Long, K.A.; Rice, G.K.; Frey, K.G.; Biswas, B.; Cer, R.Z.; Hamilton, T.; Bishop-Lilly, K.A. Characterizing Phage Genomes for Therapeutic Applications. Viruses 2018, 10, 188. [Google Scholar] [CrossRef] [PubMed]
- Keen, E.C.; Bliskovsky, V.V.; Malagon, F.; Baker, J.D.; Prince, J.S.; Klaus, J.S.; Adhya, S.L. Novel “superspreader” bacteriophages promote horizontal gene transfer by transformation. MBio 2017, 8. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. of Strains a | Sequence Type | Capsular Genotype b | Deduced K-Type | Lysis |
|---|---|---|---|---|
| 11 | ST101 | wzi137 | K17 | + |
| 2 | ST2502 | wzi137 | K17 | + |
| 1 | ST11 | wzi75 | ND | - |
| 3 | ST14 | wzi2 | K2 | - |
| 2 | ST15 | wzi24 | K24 | - |
| 1 | ST1633 | wzi137 | K17 | + |
| 1 | ST208 | wzi35 | K35 | - |
| 3 | ST258 | wzi154 | ND | - |
| 1 | ST258 | wzi29 | K41 | - |
| 1 | ST1879 | wzi154 | ND | - |
| 6 | ST307 | wzi173 | ND | - |
| 1 | ST340 | wzi50 | K15/K17/K50/K51/K52 | - |
| 4 | ST37 | wzi96 | K38 | - |
| 1 | ST42 | wzi41 | ND | - |
| 1 | ST45 | wzi101 | ND | - |
| 2 | ST512 | wzi154 | ND | - |
| 1 | ST540 | wzi33 | ND | - |
| 1 | ST554 | wzi154 | ND | - |
| 2 | ST641 | wzi38 | K38 | - |
| 1 | ST745 | wzi154 | ND | - |
| 1 | ST23 | wzi1 | K1 | - |
| 1 | ST11 | wzi174 | ND | - |
| 1 | ST489 | wzi171 | ND | - |
| 1 | ST3 | wzi3 | K3 | - |
| 2 | ST16 | wzi50 | K15/K17/K50/K51/K52 | - |
| 2 | ST730 | wzi356 | ND | - |
| 1 | ST147 | wzi64 | K14/K64 | - |
| 1 | ST475 | wzi201-like | ND | - |
| 1 | ST25 | wzi72 | K2 | - |
| 1 | ST54 | wzi14 | K14 | - |
| 1 | ST859 | wzi2 | K2 | - |
| 1 | ST66 | wzi4 | K2 | - |
| 1 | ST340 | wzi19 | K19 | - |
| ID | Sequence Type | EOP | Production |
|---|---|---|---|
| 12C47 | ST101 | 1 | High |
| 12C73 | ST101 | 0.9 ± 0.2 | High |
| KPC174 | ST1633 | 0.44 ± 0.06 | Medium |
| 5559 | ST101 | 0.5 ± 0.2 | High |
| 5583 | ST2502 | 0 a | Inefficient |
| C002 | ST101 | 1.9 ± 0.7 | High |
| K13 | ST101 | 2.62 ± 0.01 | High |
| K18 | ST101 | <0.001 b | Inefficient |
| 6071 | ST2502 | 0 | Inefficient |
| 12C29 | ST101 | 2.2 ± 0.7 | High |
| 5546 | ST101 | 3.9 ± 0.3 | High |
| 12C72 | ST101 | 3.1 ± 0.3 | High |
| KPC220 | ST101 | 0 | Inefficient |
| 494647 | ST101 | 0.9 ± 0.2 | High |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ciacci, N.; D’Andrea, M.M.; Marmo, P.; Demattè, E.; Amisano, F.; Pilato, V.D.; Fraziano, M.; Lupetti, P.; Rossolini, G.M.; Thaller, M.C. Characterization of vB_Kpn_F48, a Newly Discovered Lytic Bacteriophage for Klebsiella pneumoniae of Sequence Type 101. Viruses 2018, 10, 482. https://doi.org/10.3390/v10090482
Ciacci N, D’Andrea MM, Marmo P, Demattè E, Amisano F, Pilato VD, Fraziano M, Lupetti P, Rossolini GM, Thaller MC. Characterization of vB_Kpn_F48, a Newly Discovered Lytic Bacteriophage for Klebsiella pneumoniae of Sequence Type 101. Viruses. 2018; 10(9):482. https://doi.org/10.3390/v10090482
Chicago/Turabian StyleCiacci, Nagaia, Marco Maria D’Andrea, Pasquale Marmo, Elisa Demattè, Francesco Amisano, Vincenzo Di Pilato, Maurizio Fraziano, Pietro Lupetti, Gian Maria Rossolini, and Maria Cristina Thaller. 2018. "Characterization of vB_Kpn_F48, a Newly Discovered Lytic Bacteriophage for Klebsiella pneumoniae of Sequence Type 101" Viruses 10, no. 9: 482. https://doi.org/10.3390/v10090482