
Understanding Post Entry Sorting of Adenovirus Capsids; A Chance to Change Vaccine Vector Properties


Microbiologie Fondamentale et Pathogénicité, MFP CNRS UMR 5234, University of Bordeaux, 146 rue Leo Saignat, CEDEX, 33076 Bordeaux, France
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Viruses 2021, 13(7), 1221; https://doi.org/10.3390/v13071221
Submission received: 14 May 2021
/
Revised: 15 June 2021
/
Accepted: 17 June 2021
/
Published: 24 June 2021
(This article belongs to the Special Issue Adenovirus Cell and Immune Interactions)
Abstract
:Adenovirus vector-based genetic vaccines have emerged as a powerful strategy against the SARS-CoV-2 health crisis. This success is not unexpected because adenoviruses combine many desirable features of a genetic vaccine. They are highly immunogenic and have a low and well characterized pathogenic profile paired with technological approachability. Ongoing efforts to improve adenovirus-vaccine vectors include the use of rare serotypes and non-human adenoviruses. In this review, we focus on the viral capsid and how the choice of genotypes influences the uptake and subsequent subcellular sorting. We describe how understanding capsid properties, such as stability during the entry process, can change the fate of the entering particles and how this translates into differences in immunity outcomes. We discuss in detail how mutating the membrane lytic capsid protein VI affects species C viruses’ post-entry sorting and briefly discuss if such approaches could have a wider implication in vaccine and/or vector development.
1. Introduction
Viral infections, unlike bacterial infections that can be treated with antibiotics, remain a threat to health as very few antiviral remedies exist. Therefore, vaccination is the only widely applicable strategy to protect human (and animal) populations from viral infections. Until recently, vaccination against viral infections was mainly considered in the context of preventing childhood diseases or as the annual “flu” shot for influenza infection prevention. With the outbreak of the SARS-CoV-2 health crisis in 2020, vaccination has returned to the center of public attention as it is now seen as the only sustainable way out of this pandemic. In this context, adenovirus (Ad) vector-based vaccines have emerged as one of two major successful vaccination strategies. Following emergency approval, Ad vaccines are now being injected to a large proportion of the earth’s population in an unprecedented effort to install planetary immunity against SARS-CoV-2.
Adenovirus vector vaccines were not widely used before the COVID-19 outbreak, but their success is not unexpected. Viruses are in general very suitable vaccine platforms. They are biologically optimized and adapted for nucleic acid transport, and they elicit immune responses. Adenoviruses have many features that make them particularly ideal vaccine candidates. They have a low and well-characterized pathogenic profile paired with a high infectivity. They are technologically approachable and have been extensively used for vectorization. Their high production yield and stability allow industrial production under good manufacturing practice (GMP) conditions and provide practical features (i.e., storage) for regional distribution at a reduced cost [1]. Furthermore, Ad cell delivery into antigen-presenting cells is efficient, and they naturally provoke an innate immune response that acts as an adjuvant to boost vaccination success [2,3].
Understanding Ad biology has been crucial in developing Ad-based (vaccine) vectors. For example, the use of non-human or rare Ad serotypes in some of the currently marketed SARS-CoV-2 vaccines was deliberately chosen to prevent pre-existing vector immunity from reducing vaccine efficiency. The use of viral gene-deleted vectors provides improved vector safety. However, there are still many aspects of the viral life cycle that have not been considered when developing Ad-based vaccine vectors, and knowledge on rare or non-human genotypes is limited, including the ones currently used as SARS-CoV-2 vaccines. With this review, we would like to raise awareness for potential modifications that could be exploited when developing Ad vectors for medical purposes including vaccination. We focus on the viral capsid and how the choice of genotypes influences the uptake und subsequent subcellular sorting. We describe how understanding capsid properties such as stability during the entry process can change the fate of the entering particles and what different immunity outcomes are observed. As an example of how capsid stability and fate can be changed, we discuss in detail how mutating the membrane lytic capsid protein VI affects species C viruses. Finally, we briefly discuss if such approaches have the potential to be exploited for vaccine and/or vector development.
1.1. Adenoviruses Constitute a Diverse Family of Infectious Pathogens
Historically, Ad was first isolated in 1953 from adenoids of patients with respiratory infections [4], and soon after, the term "adenovirus" was universally adopted [5]. The family of Adenoviridae has more than 120 members, divided into five genera—depending on whether they infect mammals (e.g., Mastadenoviridae), birds (e.g., Aviadenoviridae), reptiles (e.g., Atadenoviridae), amphibians (Siadenoviridae), or fish (Ichtadenoviridae) [6,7]. The genus Mastadenoviridae also accounts for more than 70 types of human Ads [8,9], which are classified into seven species (A–G), according to their morphological, biological, and physicochemical properties, as listed in Table 1 [10,11]. Furthermore, Ads can also spontaneously recombine, thereby generating new species [12], as it was the case for AdE4, which is the only human species E virus and is likely to have resulted from the recombination between human species B and a simian Ad [13]. Although infections in humans with non-human Ads are naturally rare, they are not impossible [14,15]. The capacity of a variety of non-human Ads to infect (or transduce) human cells makes them attractive (vaccine) vector platforms. Among them are Ads derived from chimpanzees [16,17,18], gorilla [19], sheep [20], cow [21], dog [21,22], or the new world monkey [23].
Adenovirus infections can cause different pathologies depending on their cellular or tissue tropism including respiratory, ocular, urinary, or gastrointestinal diseases (see Table 1). Tissue tropism is often the result of distinct receptor usage by different Ads and may also be related to intrinsic particle stability [24]. Most infections with Ads are mild or asymptomatic in an immune-competent host. Primary Ad infections are mostly occurring during childhood and result in strong protective immunity. In contrast, in immune-suppressed individuals, Ads can cause uncontrolled, severe, and life-threatening systemic infections leading to serious cell toxicity, which can result in excessive inflammation and multiple organ failure, causing high mortality [12]. The best-studied human Ads are the species C viruses type 2 and 5, which are predominantly found in patients with upper respiratory tract and gastrointestinal infections [25] and which will be in the major focus of this review. Adenoviruses can be responsible for occasional outbreaks, such as species B viruses causing pneumonia (e.g., Ad3, 7, and 14) among immune-competent individuals [26] or Ad40/41 of species F and Ad12, 18, and 31 of species A, which infect the gastrointestinal tract (as reviewed in [26,27]). Species D viruses are a very diverse group and prone to recombination and are a leading cause of eye infections [28].
1.2. Adenovirus Capsids Are Metastable Structures
Despite their diversity, all Ads follow a similar structural and organizational framework. Adenoviruses are non-enveloped DNA viruses containing a relatively large, 26–45 kb linear double-stranded DNA genome [29,30,31]. The capsid is ~70–90 nm in diameter and has the form of an icosahedron with 20 facets and 12 vertices [32]. The capsid consists of three major structural proteins (hexon, penton, and fiber) and a variety of minor cement proteins (i.e., IIIa, VI, VIII, and IX in species C viruses) that stabilize the structure. The high-resolution structure of several Ad capsids has been obtained by cryo-electron microscopy and X-ray crystallography [24,33,34,35,36]. The icosahedral capsid has a pseudo T25 structure with 720 hexons assembled in different trimers depending on their location in the capsid. The 20 facets of the capsid are composed of nine assembled hexons (so called GON for Group of Nine), whereas at the 12 vertices, hexons are assembled in GOS (for Group of Six) (see [37,38] for more details). The pentameric penton base is inserted into the 12 vertices and serves as the basis for the protruding trimeric fiber [39,40]. The interaction between major and minor capsid proteins stabilizes the capsid, mainly through the interaction with hexons. For example, GON are stabilized by IX [41]; IX and IIIa are themselves stabilized by their interaction with VIII, which links them with GOS and GON, respectively [37]. VIII molecules outline the GON and help to stabilize hexons on the inside of the capsid. Protein VI also stabilizes hexon by binding the hexon trimer central cavity exposed to the inner viral lumen [42,43,44].
The capsid surrounds and protects the viral core composed of the viral genome, the viral core proteins (i.e., V, VII, Mu in species C viruses), and the terminal protein (TP), which is covalently bound to each 5’ end of the Ad genome. Minor capsid proteins interact with core proteins establishing a connection between capsid and genome; however, their spatial organization in relation to the core proteins remains poorly understood. It has been shown that V maintains DNA inside the capsid by interacting with VI (capsid side) and VII (genome side) [45,46,47] and probably contributes to genome release at the nuclear pore [48]. VII is the main capsid protein associated with viral DNA and promotes its condensation inside the capsid [49,50,51]. VII also protects the viral genome from immune detection through cellular factors [50,52,53,54]. Interestingly, capsid protein IX and protein V are only present in Mastadenoviridae [55,56], and viral capsids can be produced without IX [57] or protein VII [58], showing some flexibility in the capsid structure.
The genome of human Ads encodes ~45 proteins (including the structural proteins), organized into temporally regulated transcription units [59]. The transcription factor E1A is the first expressed protein and is necessary to initiate viral gene expression [60]. The genome also harbors a 5’ encapsidation sequence and inverted terminal repeats (ITR) at each extremity which are required for replication [59,61,62,63,64]. Adenoviruses encode an adenoviral protease (AVP) which is packed into the particle and processes several proteins (i.e., IIIa, VI, VII, VIII, µ, TP, and 52.55K in species C viruses) [65,66,67]. This step is required for the formation of mature infectious particles and induces the formation of metastable (i.e., less stable) capsids primed for capsid disassembly upon cell attachment and entry [68,69].
There are also species-specific differences in the structure of human Ads, e.g., in the fiber length and flexibility, which directly affects receptor binding [70,71,72]. Differences can also be observed in penton assembly. Penton of species B viruses can form inter-penton contacts, resulting in alternative and less stable virus-like particles that are devoid of genomes and occasionally without fibers. Such particles are called dodecahedron and are studied for the cell transfer of plasmids or peptides as an alternative to Ad vectors. Dodecahedrons better expose a RGD short peptide motif present in penton of most Ad species, which is required to bind target cells [73,74]. Differences in capsid stability, as discussed below, may be an important property that determines the entry fate of Ad particles.
1.3. Adenoviruses Follow a Lytic Life Cycle
The Ad infection cycle has been extensively reviewed elsewhere [75,76,77]. Here, we briefly focus on the entry part of the life cycle, and, unless stated otherwise, we will refer to species C viruses, which are the best studied (Figure 1). Adenoviruses use the capsid fiber as the primary cell attachment molecule. Most Ads, including species C viruses, use the coxsackievirus and Ad receptor (CAR) to bind target cells [78,79,80,81]. In contrast, some species B viruses preferentially bind the CD46 receptor [82] or desmoglein 2 [83], and some species D viruses were shown to interact with sialic acids [84,85] or directly with αvβ integrins (see Table 1) [86]. Thus, fiber switching between species is an attractive strategy to change tropism [87,88,89]. The primary role of the fiber molecule is to mediate the physical association with target cells, which is followed by a second interaction between penton and integrins, such as αvβ5 [90]. This interaction is mediated by the RGD peptide motif in the penton sequence [91]. Absence of this motif, as in species F viruses, is associated with less efficient cell entry [92]. Integrin binding results in integrin clustering at the cell surface [93] that triggers a signaling cascade, leading to the reorganization of the actin cytoskeleton [94,95] and the endocytic uptake via clathrin-mediated endocytosis or micropinocytosis [96,97]. RGD binding may also destabilize the capsid [98] by decreasing contacts between penton and hexon, helping the capsid to uncoat [99]. As a consequence, Ad particles lose their fiber, and one can find dissociated hexon and penton inside the endosome [100,101,102]. The endosome is a dynamic organelle primed to mature into degradative lysosomes, and Ads have to escape rapidly into the cytosol. Endosome acidification may accelerate the escape process by further weakening the capsid [100,103]. The endosome penetration process relies on the release of the internal capsid protein VI [103,104]. The released VI is characterized by a N-terminal amphipathic helix that binds to the inner leaflet of the endosome and induces positive membrane curvature, resulting in locally confined membrane rupture [75,103,105,106]. Cells respond to the virus-induced membrane rupture and activate a localized autophagy response to clear the damaged organelle [107,108,109]. To avoid degradation, Ads stall autophagy via a short PPxY peptide motif in protein VI until the particle has reached the safety of the cytosol [104,108]. Species C viruses were shown to escape from early endosomes [110]. Other species including A, B, and D first traffic to the lysosome before escaping into the cytosol [111,112,113], which may affect the immune activation triggered by those viruses [114].
Once Ads have reached the cytosol, they engage with dynein motors and use retrograde transport along microtubules to reach the microtubule organizing center [115,116,117]. Motor binding is probably mediated by hexon [115] and may require acidic priming [118]. In the vicinity of the nucleus, capsids switch transport directionality, probably by binding to kinesin motor proteins, and accumulate at the nuclear envelope [115,117,119,120]. They dock at the nuclear pore complex, then capsids are completely disassembled, and genomes are released and imported into the nucleus [51,121,122,123,124]. Once inside the nucleus, the cycle continues by transcription activation of the genome and expression of the immediate early E1A gene, which serves as a transcription factor for all other early transcription units [60,125,126,127,128,129,130]. Following expression of the replication enzymes, viral genomes are replicated and accumulate at the periphery of replication centers [131]. Then, structural proteins are expressed from the late expression unit under the control of the major late promoter [132,133]. Expression of late genes culminates in production and nuclear accumulation of all structural proteins where they assemble into progeny and package newly synthesized genomes. Nuclear assembly of the next generation of Ads then results in particle egress and cell lysis [42,134,135,136]. As mentioned above, assembly of the capsids is followed by maturation, i.e., the proteolytic cleavage of several capsid proteins by the AVP to produce infectious particles [66]. The importance of this process is exemplified by the temperature-sensitive mutant, Ts1, initially identified in a mutagenesis screen in Ad2. At the non-permissive temperature, this mutant fails to package the AVP, and assembled particles do not undergo maturation, resulting in hyperstable particles [137]. The responsible AVP mutation was genetically introduced into Ad5 retaining this phenotype. Ts1 capsids are still able to attach to cell receptors and to be endocytosed, but then remain trapped in the endosome due to their failure to liberate protein VI [103,138,139].
2. Immune Detection of Adenovirus
Adenoviruses are widespread, and a large majority of the population has been confronted with this virus, mainly during childhood. Adenoviruses are highly immunogenic, and every encounter provokes the development of specific anti-Ad immunity, both innate and adaptive, that can last a lifetime. This is important in the fight against the virus and renders Ads less harmful to the immune-competent host, but it can be an obstacle when Ads are to be used as a vector in vaccinology or gene therapy [140,141]. Understanding how Ad (and its vector derivatives) activates the immune system will allow us to improve the effectiveness and safety of vaccines. Observations in vivo, from patients participating in clinical trials, clearly showed that Ad-based vectors have the ability to induce potent immune activation [142,143], sometimes with fatal consequences [144]. In addition, various studies carried out in vitro or in animal models [145] suggest a key role of the viral particle itself (i.e., capsid and incorporated DNA) in activating the initial immune response. Here, we briefly focus on immune responses associated with the entering capsid; for a more in-depth overview, see previously published reviews [146,147,148,149].
2.1. Adenoviruses Trigger Cell Intrinsic Immunity
With the development of Ads as vectors, it became apparent that cell intrinsic or innate immunity induced by Ads is a response to the invading virion. Interestingly, both replicative and non-replicative viruses (i.e., UV-inactivated virus, non-replicative vectors, or empty capsids) activate immune responses in the infected cells, showing that the capsid has a key role in this response [145,150,151,152,153,154,155,156].
In general, Ads are sensed by cells at several steps throughout the entire viral life cycle and activate innate immunity pathways (Figure 1). This activation starts with the Ad fiber binding to the cell surface receptor CAR, which activates the mitogen-activated protein kinase (MAPK) signaling that leads to the activation of the transcription factor NF-κB. This trigger results in the production of pro-inflammatory cytokines (IL-6, IL-8) [157,158]. However, depending on Ad species, other receptors usage may modulate the strength of the signaling cascade involved in cytokine expression [159]. The interaction between viral penton and cellular integrin is a further trigger of immune activation. A study in murine macrophages showed that this binding activates the expression of IL-1α via the RGD motif [160]. This might be restricted to immune cells because in other cell models (e.g., HeLa, epithelial cells line), the role of the integrin/RGD interaction in chemokines and pro-inflammatory cytokines production was not confirmed [151,161,162]. This suggests a more important role of the internalization process itself in immune activation than the interaction with integrins. Indeed, penton binding to integrin also activates the phosphoinositide 3-kinase (PI3K) signaling to increase viral internalization [163]. Moreover, PI3K activation has been shown upon Ad infection to trigger the production of pro-inflammatory cytokines, such as TNFα [164], and the internalization process has been described as required for immune activation [165]. These studies suggest that immune activation mainly occurs at a post-internalization step, highlighting the importance of post-entry sorting (reviewed in [166]).
Once in the endosome, Ads are exposed to intraluminal pathogen-recognition receptors (PRR) such as TLR9, which is a double-stranded DNA sensor restricted to immune cells. Murine macrophages can sense Ad vectors via TLR9 to induce the production of pro-inflammatory cytokines [167]. TLR9 was also strictly required for IFNα/β production when murine plasmacytoid dentritic cells (pDC) were challenged with Ad [168]. However, in TLR9 knock-out mouse models, IFNα/β was still produced, suggesting that, in cells other than pDC, IFN production occurs independently from TLR9 signaling [168,169].
Viral genomes (or vector genomes) become accessible to sensing during or after the endosomal escape of the virus. Once they reach the cytosol, they are sensed by the cytosolic double-stranded DNA sensor cGAS [168,170]. Upon viral genome recognition, cGAS promotes phosphorylation and nuclear translocation of the transcription factor IRF3 to drive IFNα/β expression, inducing an antiviral state. Endosomal passage requires virus-induced endosomal membrane damage to facilitate endosomal escape of viral particles. Membrane rupture thus considerably contributes to immune activation, highlighted by the fact that the escape defective Ts1 mutant fails to activate a complete and efficient immune response [161,162,170,171]. A pivotal role in the inflammatory response upon Ad membrane penetration is played by the tank-binding kinase, TBK1, suggested to be part of a down-stream cytosolic sensing of the viral genome via the cGAS/STING pathway [168,170].
Adenovirus membrane damage also results in the release of cathepsin B from the endo-lysosomal compartment, causing oxidative stress, which activates the NLRP3 inflammasome, resulting in IL-1β maturation [114]. Species C viruses escape from early endosomes, while species B viruses traffic until late endosomes, presumably due to differences in receptor usage. Acidification in late endosome/lysosome is more likely to activate lysosomal acid hydrolases. Both species induce inflammasome activation [172], but, probably owed to the residing time in the endosome, the extent of their responses differ. Globally, species B viruses induce a stronger innate and adaptive immune activation than species C viruses [113,173,174]. This may explain why species B viruses are more pathogenic, causing outbreaks in immune competent hosts [26]. It further suggests that cells are able to discriminate the penetration compartment (i.e., endosome vs. lysosome, [109]) and adapt the efficiency of immunity. Endosomal membrane damage during the Ad escape process can also be a danger signal in itself. As discussed in detail below, membrane damage results in the cytosolic exposure of intraluminal glycans, which are detected by the cell as danger signals (as reviewed in [109]). The detection of exposed glycans by galectins subsequently activates autophagy, triggering a second branch of antiviral immunity [108].
2.2. Adenoviruses Provoke an Aadaptive Immune Response and Subvert Antiviral Autophagy upon Cell Entry
Innate immune responses provoked by Ads or their vectors subsequently translates into adaptive immunity through chemokine secretion that attracts immune cells (neutrophils, natural killer cells, and macrophages) and reinforces potent antigen presentation [153,154]. Adenoviruses are highly immunogenic, and their main immunogens are the major capsid proteins (hexon, penton, and fiber) that trigger the species-specific production of neutralizing antibodies by B cells [175,176,177]. Neutralizing antibodies are mostly directed against the hyper-variable loops of the hexon protein [178,179,180] and represent a major limitation for the use of Ads as vectors, if derived from the same serotype. This problem has been circumvented by the use of less seroprevalent or non-human Ads [181,182,183,184,185]. Despite this anti-vector immunity, when Ads are used as vaccine platforms (e.g., to elicit immune protection against Ebola in clinical trials), they still induce a strong B cell response, allowing the production of antibodies against the desired vaccine immunogen that can last for up to 6 months [186]. In addition, pre-existing vector immunity could be overcome with higher vaccine vector doses, as in the case of the SARS-CoV-2 vaccine vector based on the highly seroprevalant AdC5 [187].
As with neutralizing antibodies, Ad infections trigger a persistent T-cell immunity, mainly through CD4+ activation. This protection can still be found in adults, suggesting a long-lasting immunity from childhood [188,189,190]. In addition, activation of CD8+ is also described to maintain a cytotoxic response [191,192,193]. This T-cell activation is not necessarily serotype-specific and allows cross-protection against different Ad subgroups and serotypes [192,193,194]. Moreover, in stem-cell-transplanted patients infected by Ad, a strong activation of CD4+ and CD8+ has been described [195]. This activation is directed against hexon peptides and leads to a clearance of Ad viremia, emphasizing the major role of both CD4+ and CD8+ T-cell activation in immune protection. While this T-cell-based immunity is directed against the Ad particle, Ads also trigger strong cellular immunity based on the T-cell response against the vaccine antigen, which is an advantage for long-lasting immunity when using recombinant Ad vectors as vaccine platforms [196,197]. Importantly, data from clinical trials have confirmed that when Ad is used as a vaccine platform (with the desired antigen as transgene), it induces both cytotoxic CD8+ T-cell and CD4+ T-cell activation, leading to memory immunity against the expressed transgene [184,186,198,199,200,201,202].
T-cell activation relies on the presentation of viral peptides through the major histocompatibility complex (MHC). Briefly, antigen presentation by MHC class I results from proteasomal degradation, and the peptides obtained will induce CD8+ T-cell activation, allowing priming and proliferation of a cytotoxic T-cell response. MHC class II antigen presentation results from lysosomal proteolysis, and the resulting peptides will induce CD4+ T-cell activation upon presentation, triggering the subsequent activation of B cells [203]. It is now well characterized that autophagy promotes peptide presentation to the MHC class II molecules [204]. Not surprisingly, autophagy is one of the oldest defense mechanisms against infection and an important part of the cellular repertoire against invading pathogens, participating in both innate and adaptive immune responses. It can directly sequester a pathogen for degradation (xenophagy) and participates in their presentation to the immune system [205,206,207]. Macroautophagy (more commonly called autophagy) is a conserved lysosomal degradation pathway that participates in many fundamental physiological processes such as homeostasis and innate and adaptive immunity [208,209,210]. Autophagy is characterized by the formation of a double membrane vesicle called “autophagosome” that engulfs cytoplasmic cargoes (i.e., organelles, aggregates, and pathogens) destined for degradation (Figure 2). During this process, cargo-containing autophagosomes fuse with lysosomes in order to be degraded and recycled.
Antigen presentation via autophagy has been described for both MHC class I and class II [204]. After induction of autophagy, intracellular antigens are engulfed in autophagosomes, fusing with MHC class II containing compartments to provide them with external pathogen material (Figure 2) [211]. The role of autophagy in this particular MHC class II antigen presentation was first described for Epstein-Barr virus infection, where inhibition of autophagy led to decreased CD4+ T-cell activation [212,213]. To date, examples include several viruses including herpes simplex virus [214], influenza virus [215], and human immunodeficiency virus type 1 (HIV-1) [216], and probably extends to Ads (see below). MHC class II presentation is consistent with the process of autophagy as it is a cytosolic defense mechanism.Autophagy is also involved in MHC class II cross-presentation of extracellular antigens [214]. Indeed, the MHC presentation of extracellular ovalbumin is more efficient when the autophagy machinery (and especially ATG5) is functional, and the uptake of extracellular material was shown to involve a non-canonical form of autophagy called LC3-associated phagocytosis (LAP) [217]. Once present in the LAP-osome, extracellular antigens can be exposed through MHC class II presentation [218]. Autophagy is also involved to some degree in MHC class I presentation, although the vast majority of the epitopes are presented via a mechanism strictly dependent on the transporter associated with the antigen processing (TAP) complex [219]. TAP-independent presentation mediated by autophagy was so far shown only under TAP depletion conditions [219,220]. If this mode of presentation truly exists remains controversial [221] and is discussed elsewhere [204,222,223].
To prevent MHC presentations, Ads dedicate a great deal of their genome, namely the E3 region, to undermine the MHC presentation system in cells [224,225,226,227,228]. In most (vaccine) vectors, these genes are absent, and recent work suggests that increasing antigen presentation by MHC could improve Ad vaccine efficiency [229,230]. Autophagy plays a crucial role in processing Ad antigens for MHC class II presentation in Ad-induced immunity [108,229].
Montespan et al. showed that Ads induce autophagy upon endosome penetration and that entering particles cleared by autophagy in this process are efficiently presented to the immune system [108]. This observation is consistent with the study published by Klein et al. showing that in the context of oncolytic Ads, the viral antigens are matured by a JNK-dependent form of autophagy [229]. However, autophagy in the context of cancer plays a dual role. As a homeostasis keeper, autophagy limits tumor initiation [231] while in advanced cancer autophagy acts as a survival process to favor tumor growth [232,233]. Thus, in the context of oncolytic vectors, tumor-specific autophagy features should be carefully evaluated.
3. Adenoviruses as (Vaccine) Vectors
Vectorization of a virus involves the exploitation of its natural properties, while simultaneously minimizing the associated biological risk. In these terms, Ads are excellent vector candidates for vaccine development [234]. Adenovirus vectors are stable, are easy to produce to a high titer, and have large cloning capacities. This allows insertion of a transgene, i.e., to express an antigen of choice. They have a broad infectivity spectrum and transduce non-dividing cells, and their genome is non-integrative [1]. As described above, they also provoke an inflammatory response in antigen-presenting cells (APCs) that serves an adjuvant function to amplify the immune response necessary to vaccinate efficiently [2,3,142,235,236]. Moreover, Ad-derived vectors have been extensively used in preclinical and clinical trials to prove their safety [237,238]. To eliminate the unwanted part of the replication cycle, viral replication needs to be suppressed to prevent cell lysis and viral propagation. Adenovirus vector development started during the early 1990s as a platform for a large range of therapeutic approaches, such as gene therapy, oncolytic vectors, and vaccine development. Most of the early vector developments were modified versions of species C viruses.
3.1. Adenovirus Vector Development, a Generational Approach
The first generation of an Ad vector was made by deleting the E1 and E3 region from the viral genome, notably encoding the immediate early transcription factor, preventing these vectors from replicating. These vectors had a cloning capacity of around 8 kb to insert a transgene with regulatory sequences [239,240,241]. The first-generation vectors were produced in human embryonic kidney (HEK) cells stably transformed with a part of the Ad5 viral genome that complements the E1 region in trans [242,243,244,245,246,247]. One of the major drawbacks of this system is that it allows the reconstitution of replication-competent genomes after spontaneous recombination [248]. The additional deletion of E3, which is not necessary for vector production, increases the size of the transgene cassette [243], and promotes a better immune response [240]. Unfortunately, first-generation Ad vectors were not completely transcriptionally inert and still elicited a strong immune response against viral capsid proteins that were expressed at baseline levels [141,249,250]. This anti-vector immunity contributed to vector toxicity and short-term elimination of transduced cells [145]. The second generation of Ad vectors additionally had the E2 and E4 regions removed from the vector genome. E2 encodes the replication enzymes and E4 additional regulatory proteins [251,252,253]. This allowed space for 10.5 kb of transgene sequences, including up to four expression cassettes [254]. Likewise, the supplemental deletions strongly reduced unwanted recombination and prolonged transgene expression [255]. This was probably the result of a reduced aberrant viral gene expression that further reduces anti-vector immunity [141,250,251]. Adenoviral vectors of the third generation are conceptually different. They consist of a packaged vector genome and a non-packaged viral genome (helper virus) that produces vector particles. Such vectors are also known as “helper-dependent”, “gutless”, or “high-capacity” (HC-Ad) vectors. They have the highest possible cloning capacity and can carry sequences of up to 36 kb [256,257,258]. High-capacity Ads lack any viral coding sequence except ITRs and a short encapsidation signal that permits genome packaging and which is absent from the helper virus [259,260,261,262]. This strategy has allowed us to finally overcome most anti-vector immunity, permitting the long-term expression of transgenes [263,264,265,266,267]. However, HC-Ads are not part of the current vaccine vector repertoire.
3.2. Adenovirus as a Vaccine Vector
Before exploring how Ad vectors have been exploited, it is noteworthy to mention that vaccination against Ad species B viruses type 4 and type 7 themselves was developed in the early 1970s [268]. This oral live vaccine is mandatory since 2011 for military recruits in the USA and dramatically decreased infection rates, reaching virtually zero [269,270,271]. This highlighted the intrinsic efficiency and safety of Ad as a vaccine and opened the field to use Ad as a vaccination platform. To date, over 200 Ad-based vaccines entered clinical trials [272]. Many of them are directed against infectious pathogens such as HIV-1 and Ebola [230,273,274]. Adenovirus-based vaccines are not without drawbacks. A recently developed anti-HIV vaccine, based on the species C virus Ad5, used a mixture of vectors expressing the HIV-1 gag, pol, and nef genes [275,276]. The vectors were safe, immunogenic, and well-tolerated. Unfortunately, the resulting STEP clinical trial had to be prematurely stopped [277,278,279]. Not only has this vaccine been shown not to protect against HIV infection, but epidemiological data suggested that vaccination with Ad in this case increased the HIV infection risk [280,281,282]. Having a pre-existing immunity against Ad5 was identified as a potential post-vaccination risk factor [283,284]. However, this observation was not confirmed in similar studies [285,286] and remains mechanistically unclear [287,288]. Next to HIV, Ad-based vector vaccines have been developed in the fight against Ebola (Ad-EBOV), a recurrent threat in Western Africa [289]. All Ad-EBOV vaccines use the Ebola glycoprotein (GP) as the vaccine antigen. Initial preclinical trials in rodents and primates used Ad5 and showed protection over 3 months with production of neutralizing antibodies and CD8+ activation [273,290,291,292]. This vaccine went through phase II clinical trial; however, pre-existing immunity to Ad5 in the vaccinated population has led to variation in the level of elicited neutralizing antibodies [293,294,295,296,297]. To overcome this issue, vaccines based on the less prevalent species D Ad26 have been developed [186,298,299]. However, Ad26 is less immunogenic in humans, resulting in decreased vaccination efficiency, and required a prime boost using a measles virus-based vaccine (MVA). This vaccination regime has reached phase III clinical trials [289,300,301]. Use of non-human Ad vectors has also been under development [184]. Use of chimpanzee ChAd3 as a vaccine platform against EBOV showed protection in rodents and in chimpanzees [302,303], and clinical trials employing those vectors are at different steps of progress [17,300,301,304,305,306]. Until recently, HIV-1 and EBOV vaccines were the two main examples for the application of Ad vector-based vaccines, but they are not the only ones. Different attempts have been made to employ Ad vaccine vectors against various pathogens such as Mycobacterium tuberculosis [307,308,309,310], dengue virus [311,312,313], hepatitis B and C [314,315,316,317,318,319,320], rabies [321,322,323,324], or influenza [325,326,327,328]. However, their efficacy was always below expectations (for more details, see [18,142,300,329]). The EBOV vaccine candidate has remained the only practically applied Ad vaccine and is used with some restrictions in the Democratic Republic of Congo. This vaccine is based on the Ad26 type (Ad26-ZEBOV/MVA-BN-FILO), and, as of May 2020, 20,339 people received the first dose of vaccination, and 9560 were fully vaccinated [230,301,330].
Adenovirus-vectorized vaccines have received a major public boost in response to the SARS-CoV-2 pandemic. The urgency of the situation has made it possible to capitalize on their development, including experience gained with vaccines raised against coronaviruses that caused previous regionally confined SARS-CoV-1 outbreaks [331]. The coronavirus spike (S) glycoprotein was identified as the best vaccine antigen for immunity to coronavirus infections [332,333]. To date, four Ad vector vaccines, all encoding the spike antigen, are in an advanced state and have been granted for emergency use in large parts of the world. The applied vector strategies have been slightly different, but essentially involve the use of first-generation vectors. They range from using species C Ad5-based vectors (Ad5-nCoV), overcoming pre-existing immunity with a high vector dose (CanSino) [187,334], to the use of less prevalent species D virus Ad26, using a stabilized spike glycoprotein (JNJ-78436735/Ad26.COV2.S) [335]. An alternative approach uses ChAdOx1-S-(AZD1222), an Ad vector from chimpanzees [336,337]. The only vaccine in use based on a heterologous prime-boost strategy to overcome vector immunity (Gam-COVID-Vac/Sputnik V) applies first an Ad26-based vector and then in the booster injection uses an Ad5 vector [338,339]. In all cases, clinical trial data have been very promising, showing high levels of induced protective immunity, and emergency use has been granted in several countries for the use of Ad-based vaccines. Furthermore, epidemiological surveys in the ongoing vaccination campaigns in different countries suggest an overwhelming success in the use of Ad-based SARS-CoV-2 vaccines.
4. Modulating Adenovirus (Vaccine) Vector Efficacy, the Capsid Leads the Way
Adenovirus (vaccine) vector design has to take into account a number of factors. These include maintaining a long and sustained expression of the encoded antigen and the activation of an immunological context (adjuvant effect) that permits efficient translation of the antigen expression into a persistent adaptive immunity directed against the immunogen but not against the vector itself. The immunological context is provided by the innate immune activation that the vaccine vector elicits during administration. In this context, the efficiency with which innate immune sensors detect the vector is important, and will in turn determine the amplitude of the adaptive response. As described in the sections above, innate immune sensing is mainly taking place at the post-entry level, concomitant with Ad endosome penetration. The efficient expression of the (vaccine) antigen depends then on the balance between the level of vector clearance vs. successful delivery and expression of vector genomes to the nucleus. With the development of capsid display vectors, expression of the transgene is not required to elicit immune activation. This strategy is based on the insertion of an epitope of interest into Ad capsid proteins, allowing its direct exposure to the immune system. Even if this approach has some advantages, most vaccine vectors rely on genetically encoded genes. For further reading on capsid display vectors, please see these reviews, [340,341].
Pre-existing immunity against a viral vector (e.g., 30–90% against the species C virus Ad5, depending on the geographic location) can reduce vector uptake and decrease the antigen expression and the resulting immune response [342,343,344]. Several strategies have been developed to decrease anti-vector immunity and have been recently reviewed [249]. Briefly, the neutralizing antibodies mainly recognize the hypervariable loop of hexons and as a consequence are serotype-specific [178,179,180]. These hypervariable loops can therefore be replaced by those of another, less prevalent serotype [178,345]. This prevents the vector from being neutralized and inactivated. As Ad5 is the most seroprevalent type, using rare serotypes such as Ad26, Ad35, or non-human Ads would also avoid this issue [177,346,347,348,349,350]. However, a major drawback with the use of rare types is their lower immunogenicity that makes them less efficient regarding vaccine development [298]. Therefore, strategies to decrease anti-vector immunity should be developed in parallel to strategies to enhance vector immunogenicity in order to obtain the most efficient vector. Even if the initial level of neutralizing antibodies against the vector is low, vaccination with a given vector type may trigger a response that prevents its future use. Priming of the anti-vector response also takes place at the post-entry level, when vector particles are sorted and processed as antigens themselves. Eliciting an immunologically favorable context for vaccination success may come at the expense of increased priming for vector immunity. Understanding the parameters that determine the fate of Ad vectors after uptake may provide an opportunity for vector design.
4.1. Stability Lies in the Species
Receptor-mediated uptake and escape from the endo-lysosomal compartment are hallmarks of Ad infection and vector transduction. The initial uptake is determined by the interaction of the virus fiber and penton molecules with cell receptors. Fiber and penton are common to all Ads, but, according to their species, they engage with different cellular receptors (Table 1). Species C viruses use CAR, and entering particles escape from an early endosomal compartment [79]. If species C viruses (Ad5) are engineered to contain a fiber molecule from a species B virus (Ad16), the resulting particle (Ad5F16) then escapes from a lysosomal compartment (as shown by costaining with the lysosomal marker LAMP1) instead of the early endosome [351]. This delayed escape from lysosomes is a feature shared with all tested species B viruses [111,352,353]. Furthermore, the hybrid Ad5F16 as well as species B viruses elicit a stronger pro-inflammatory response than species C viruses [159,351]. This is attributed to increased detection via intraluminal TLR9 in immune cells [354]. In addition, lysosomal membrane damage, unlike early endosome damage, causes strong oxidative stress. Lysosome rupture co-releases lysosomal hydrolases such as cathepsin B that can damage mitochondria, thus amplifying the damage response through inflammasome activation [355,356,357]. The Ad uptake process destabilizes the capsid, allowing the release of the viral protein VI [103,106]. Released protein VI then penetrates the membrane from within the endo-lysosomal compartment to allow the virus to gain access to the cytosol. Comparing species C and B shows that both viruses are equally efficient in cytosol entry and genome delivery, but differ in their escape kinetics [111]. The fact that a simple fiber swap can transfer the entry and immunogenic properties from species B to a species C virus is remarkable. It provides a proof of concept that hybrid vectors with specific properties can be designed. The underlying difference seems to be that protein VI release is delayed in species B viruses (or the hybrid virus) and may require either acidification or another, unknown disassembly trigger. Since protein VI release is delayed in this case, this could reflect an increased capsid stability. Indeed, species C Ad5 viruses were shown to release the fiber molecule upon cell binding, a step assumed to weaken the capsid and prime the disassembly process for protein VI release. In this context, fiberless Ad5 were shown to be less stable than native particles [358]. In contrast, non-matured, hyperstable Ts1 Ad2/5 do not release fiber or protein VI and are poor inducers of inflammation, since they are neither triggering TLR9 nor causing membrane damage [114,138,359,360]. Whether fiber swapping with species B prevents initial fiber release and renders the species C capsid more stable has not been investigated but is probable. Capsid stability maybe an essential determinant of post-entry capsid fate and, by extension, a feature that could be exploited for vector design. Species B viruses may be more stable due to the use of a different receptor or through a unique fiber-penton interaction. In contrast, species F viruses Ad40/41 infect the gastrointestinal tract and have increased capsid stability due to their tropism and their adaptation to the harsh gut environment [24,361]. The gut has an intrinsic temperature that is a few degrees above the upper respiratory tract targeted by species C viruses (37 °C vs. 33 °C) [362]. Thus, Ad40/41 are intrinsically more thermostable, supporting the infection process [24], making them also difficult to propagate [24,361,363]. In addition, they lack a RGD motif in the penton molecule, resulting in a further reduced cell entry [92]. Whether these unique capsid properties translate into escape from a lysosomal compartment or partial disassembly/protein VI release defects upon entry is not known. In contrast, these properties may provide a context for the development of oral vaccine applications. Unlike species B fibers, species F fiber-harboring vectors have a decreased immune activation [364] when applied at standard environmental conditions. If a given human Ad species is now taken out of context and employed as a vaccine vector, it could be worthwhile to look at the natural tropism of this vector to look for clues as to predict its behavior. The currently used vaccine vector based on Ad26 is a species D virus, and its natural tropism is the eyes and the gastrointestinal tract [365]. Species D viruses including Ad26 are less immunogenic than Ad5 [366] and were shown to traffic to the late endosomal compartment, probably involving delayed protein VI release. Consequently, blocking acidification of the late endosomal compartment further reduced its immunogenicity [113]. In contrast to human Ads, the knowledge on non-human Ads also used in vaccination and vectorization is very limited. Chimpanzee-derived Ads including the ChAd3 are closely related to species D viruses [367,368]. Interestingly, Ad5 vectors are more immunogenic than chimpanzee-derived Ad vectors and have a higher transduction potential. This observation was not linked to differences in the penton RGD motif or the fiber, and its mechanistic reason remains unknown [369]. Potential differences in capsid stability have not been investigated. Another non-human Ad exploited as a vector is the canine Ad type 2 (CAV-2). This vector has the remarkable property to transduce neurons selectively, but not surrounding glia cells, unlike an Ad5 control vector, despite both vectors using the same receptor [370,371,372]. This ability may be linked to an increased capsid stability, allowing uncoating only in neuronal cells. Dogs have a body temperature that is ~2 °C higher than humans, which potentially results in elevated CAV-2 capsid stability [373], analogous to the species F viruses. CAV-2 poorly elicits an innate immune response, consistent with a lack of efficient disassembly in target cells [372]. Thus, CAV-2 vectors might be another example where a biophysical property linked to its natural tropism or host has the potential to provide novel vector properties.
4.2. Fine-Tuning the Capsid Structure, the Example of Protein VI in Species C Ad2/5
If capsid stability and protein VI release determine the downstream fate of the viral capsid, the question arises whether this can be modified in Ad vectors, i.e., to attribute novel properties to viral vectors (Figure 3). Most of our understanding of how Ads penetrate the endo-lysosomal compartment comes from the species C viruses Ad2/5. The discovery of protein VI as a membrane lytic factor was essential to understand this process [103,106]. Protein VI encodes a N-terminal amphipathic helix that is the actual membrane lytic part of the protein. During virus production, protein VI is expressed as a precursor protein that associates with hexon trimers via the amphipathic helix [42,374]. The complex is imported into the nucleus via transport signals encoded in the N- and C-terminus of protein VI, which are removed during virus assembly and maturation by the viral protease. The C-terminal 11 amino acid peptide also activates the protease, providing a smart way to link virus assembly with shielding of the amphipathic helix. The Ts1 Ad2/5 mutant virus has a packaging defect for the protease, and none of the capsid proteins, including protein VI, is processed. Resulting particles are hyperstable, retain fibers, and do not deploy protein VI. As consequence, Ts1 particles are sorted into lysosomes upon receptor-mediated endocytosis. The exact location of protein VI in the mature virus is not clear, but the cleaved N-terminus localizes to the inner hexon surface of peripentonal hexons [37,104,375]. Two point mutations in protein VI, G33A and S28C, were shown to increase the Ad5 capsid stability (Figure 3A) [376,377]. The first mutant precedes the N-terminal protease cleavage site in protein VI and impairs the processing. Resulting particles are partially immature and are characterized by an endosome lysis defect, suggesting that the N-terminus of protein VI stabilizes intra-virion protein-protein interactions [376]. The second mutant was chosen for its ability to form intra-molecular disulfide bridges [377]. The virus contained partially unprocessed proteins, potentially including protein VII, but surprisingly did not show any infectivity defects, despite its increased stability. Whether either of the viruses has an altered immune activation profile is not known. Another point mutation in protein VI that affects the virus fate is the protein VI L40Q mutation, directly situated in the amphipathic helix (Figure 3A) [378,379]. Viruses with this mutation mature normally but are partially defective for the endosome lysis step. In contrast to the above mentioned mutants, L40Q virions are less stable than their wild type counterparts. Instead of a delay, protein VI as well as penton are released prematurely from the L40Q capsid. L40Q virions are less infectious and subject to partial lysosome sorting. However, the L40Q mutation does not completely abolish the membrane lysis, and some viruses escape to the cytosol, resulting in an overall ~four-fold reduced infectivity of a L40Q vector [379,380]. The observations suggest that the amphipathic helix contributes with its interaction to the particle stability. It is not known if and how the L40Q mutation alters the immunogenic property of Ad5 vectors. Recent observations suggest that release of protein VI from the capsid is triggered by internal competition with protein VII for the same or overlapping binding sites in hexons [375]. Consistently, a mutant virus devoid of protein VII fails to escape from the endosome and has a defect in endosome lysis [58]. Further understanding the structural and dynamic organization of the internal capsid (i.e., protein VI) and core proteins (i.e., protein V and VII) in the particle and during cell entry will greatly help to understand what determines (reversible) capsid stability.
Controlling the efficiency of immune activation of Ad vectors is a major interest for the development of vaccines, and expanding MHC presentation of antigens would improve vaccine development [381]. As mentioned previously, autophagy regulates immune activation and feeds peptides to the MHC. Autophagy is one of the oldest cellular defense mechanisms against infection, yet Ads, as adapted human pathogens, have not evolved mechanisms to avoid autophagy upon entry. Instead, they seem to embrace autophagy activation and to divert the response to promote infection. Protein VI plays a central role in controlling the autophagy response of the cell. Adenovirus membrane lysis by protein VI leads to the recruitment of galectin 3 and galectin 8, which are recognized by the cell as danger signals (Figure 3B) [108,109,382]. Detection of Ad penetration sites by galectins activates selective autophagy in the infected cell, as illustrated by the recruitment of autophagy receptor (e.g., NDP52 andp62) and LC3 punctae formation [108]. Interestingly, Ad5 vector particles manage to escape from the ruptured endosome and are able to traffic to the nucleus. Inhibiting autophagy does not affect viral infectivity, but delays their genome delivery to the nucleus, showing that Ads hijack the cellular machinery for their own profit [108]. Escape from the endosome therefore allows the virus to avoid degradation by autophagy via an active process. This process was discovered by yet another protein VI mutant, the M1 mutant. All human Ads (and several non-human Ads) encode a highly conserved PPxY peptide motif in protein VI. In the M1 mutant, this motif was changed to PGAA (Figure 3) [104]. Both wild type and the M1 mutant efficiently release protein VI upon entry and trigger autophagy after rupture of the endosomal membrane. However, the M1 mutant virus is unable to escape efficiently into the cytosol, and EM images have shown that M1 mutant viruses are trapped in ruptured endosomes, targeted by autophagy. Consequently, the M1 mutant virus was shown to have a strong infectivity defect, which is restored upon the pharmacological and genetic inhibition of autophagy. The study showed that wild type viruses use their PPxY motif to recruit the ubiquitin ligase NEDD4.2 and limit the maturation of autophagosomes long enough to escape to the cytosol. Furthermore, this property allows the virus to limit its antigenic presentation [108]. Importantly, the M1 mutant vector injected into mice resulted in much stronger anti-vector immunity and a reduced CD8+ response to the encoded transgene, showing that a simple capsid alteration can have a profound impact on the immunogenicity of the Ad5 capsid. Whether any of the other protein VI mutations affects immunogenicity is not known. However, the presence of the PPxY motif in the L40Q virus may explain why the vector retains a relatively high infectivity despite the membrane lysis defect [379,380]. Because the endosome lysis is less efficient, and L40Q vectors end up in part in the lysosomal compartment, it is conceivable that the elicited immune response is yet different from either wild type, Ts1, or M1 Ad5 capsids.
5. Conclusions
As demonstrated in this review, development of Ad-based (vaccine) vectors with specific and desired properties is in part driven by a comparison between available Ad species without necessarily understanding the mechanistics behind existing differences. The lack of data concerning the underlying biology for most vectors, other than species C, means that relying on rare or non-human serotypes bears a risk of missing out on properties or overlooking problems that can arise. We strongly advocate to ameliorate this lack of knowledge by pairing vector development and application with investigating the underlying vector and virus biology. In the meantime, we believe that the existing knowledge base on Ad biology, especially for species C viruses, could help with the design of new Ad vectors, including rare or non-human species, using rational design. In this review, we highlighted the link between Ad post-entry sorting and the ensuing innate immune response and provided examples of how Ad post-entry sorting differs between species and has been modulated. The non-exhaustive list of possible capsid modifications exemplified on capsid protein VI and restricted to Ad2/5 shows the great potential that still lies buried in the Ad capsid, waiting to be exploited. Lysosomal membrane damage, such as early endosomal membrane damage, also triggers autophagy, but in addition results in oxidative stress that alters the innate immune response of the cell [383]. Altering the endo-lysosomal escape process of a species B, species D, or non-human Ad by introducing desirable mutations into the respective protein VI gene is only one of the possible options that come to mind. Other capsid proteins may hold additional functions related to transport or immune modulation. Extending rational capsid modifications to last-generation HC-Ad vectors and optimized antigen expression cassettes hold great promise for the next generation of Ad-based vaccine vectors.
Author Contributions
C.F.D., N.P. and H.W. wrote, reviewed, and edited the paper. C.F.D. and N.P. contributed equally to this manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
C.F.D. received support from the Agence Nationale de la recherche (project VirMEDACO grant number ANR-19-CE15-0013-01) and N.P. received support from the Fondation pour la Recherche Médicale en France FRM DEQ20180339229. H.W. is an INSERM fellow.
Acknowledgments
We acknowledge the help of Sarah Cookson for manuscript preparation.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ghosh, S.S.; Gopinath, P.; Ramesh, A. Adenoviral vectors: A promising tool for gene therapy. Appl. Biochem. Biotechnol. 2006, 133, 9–29. [Google Scholar] [CrossRef]
- Adams, W.; Loré, K. Recombinant adenovirus vector infection of human dendritic cells. Viral Gene Ther. 2011. [Google Scholar] [CrossRef]
- Mercier, S.; Gahéry-Segard, H.; Monteil, M.; Lengagne, R.; Guillet, J.-G.; Eloit, M.; Denesvre, C. Distinct roles of adenovirus vector-transduced dendritic cells, myoblasts, and endothelial cells in mediating an immune response against a transgene product. J. Virol. 2002, 76, 2899–2911. [Google Scholar] [CrossRef] [Green Version]
- Rowe, W.P.; Huebner, R.J.; Gilmore, L.K.; Parrott, R.H.; Ward, T.G. Isolation of a cytopathogenic agent from human adenoids undergoing spontaneous degeneration in tissue culture. Proc. Soc. Exp. Biol. Med. 1953, 84, 570–573. [Google Scholar] [CrossRef]
- Enders, J.F.; Bell, J.A.; Dingle, J.H.; Francis, T.; Hilleman, M.R.; Huebner, R.J.; Payne, A.M.M. “Adenoviruses”: Group name proposed for new respiratory-tract viruses. Science 1956, 124, 119–120. [Google Scholar] [CrossRef]
- Lefkowitz, E.J.; Dempsey, D.M.; Hendrickson, R.C.; Orton, R.J.; Siddell, S.G.; Smith, D.B. Virus taxonomy: The database of the International Committee on Taxonomy of Viruses (ICTV). Nucleic Acids Res. 2018, 46, D708–D717. [Google Scholar] [CrossRef] [Green Version]
- Harrach, B.; Tarján, Z.L.; Benkő, M. Adenoviruses across the animal kingdom: A walk in the zoo. FEBS Lett. 2019, 593, 3660–3673. [Google Scholar] [CrossRef] [PubMed]
- Hage, E.; Liebert, U.G.; Bergs, S.; Ganzenmueller, T.; Heim, A. Human mastadenovirus type 70: A novel, multiple recombinant species D mastadenovirus isolated from diarrhoeal faeces of a haematopoietic stem cell transplantation recipient. J. Gen. Virol. 2015, 96, 2734–2742. [Google Scholar] [CrossRef]
- Sarantis, H.; Johnson, G.; Brown, M.; Petric, M.; Tellier, R. Comprehensive detection and serotyping of human adenoviruses by PCR and sequencing. J. Clin. Microbiol. 2004, 42, 3963–3969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crenshaw, B.J.; Jones, L.B.; Bell, C.R.; Kumar, S.; Matthews, Q.L. Perspective on adenoviruses: Epidemiology, pathogenicity, and gene therapy. Biomedicines 2019, 7, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghebremedhin, B. Human adenovirus: Viral pathogen with increasing importance. Eur. J. Microbiol. Immunol. 2014, 4, 26–33. [Google Scholar] [CrossRef] [Green Version]
- Lion, T. Adenovirus infections in immunocompetent and immunocompromised patients. Clin. Microbiol. Rev. 2014, 27, 441–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dehghan, S.; Seto, J.; Liu, E.B.; Walsh, M.P.; Dyer, D.W.; Chodosh, J.; Seto, D. Computational analysis of four human adenovirus type 4 genomes reveals molecular evolution through two interspecies recombination events. Virology 2013, 443, 197–207. [Google Scholar] [CrossRef] [Green Version]
- Dehghan, S.; Seto, J.; Liu, E.B.; Ismail, A.M.; Madupu, R.; Heim, A.; Jones, M.S.; Dyer, D.W.; Chodosh, J.; Seto, D. A zoonotic adenoviral human pathogen emerged through genomic recombination among human and nonhuman simian hosts. J. Virol. 2019, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medkour, H.; Amona, I.; Akiana, J.; Davoust, B.; Bitam, I.; Levasseur, A.; Tall, M.L.; Diatta, G.; Sokhna, C.; Hernandez-Aguilar, R.A.; et al. Adenovirus infections in African humans and wild non-human primates: Great diversity and cross-species transmission. Viruses 2020, 12, 657. [Google Scholar] [CrossRef]
- Hodgson, S.H.; Ewer, K.J.; Bliss, C.M.; Edwards, N.J.; Rampling, T.; Anagnostou, N.A.; de Barra, E.; Havelock, T.; Bowyer, G.; Poulton, I.D.; et al. Evaluation of the efficacy of ChAd63-MVA vectored vaccines expressing circumsporozoite protein and ME-TRAP against controlled human malaria infection in malaria-naive individuals. J. Infect. Dis. 2015, 211, 1076–1086. [Google Scholar] [CrossRef]
- Tapia, M.D.; Sow, S.O.; Lyke, K.E.; Haidara, F.C.; Diallo, F.; Doumbia, M.; Traore, A.; Coulibaly, F.; Kodio, M.; Onwuchekwa, U.; et al. Use of ChAd3-EBO-Z Ebola virus vaccine in Malian and US adults, and boosting of Malian adults with MVA-BN-Filo: A phase 1, single-blind, randomised trial, a phase 1b, open-label and double-blind, dose-escalation trial, and a nested, randomised, double-bli. Lancet Infect. Dis. 2016, 16, 31–42. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Mondal, M.; Zhou, D. Development of novel vaccine vectors: Chimpanzee adenoviral vectors. Hum. Vaccines Immunother. 2018, 14, 1679–1685. [Google Scholar] [CrossRef] [Green Version]
- Hollingdale, M.R.; Sedegah, M.; Limbach, K. Development of replication-deficient adenovirus malaria vaccines. Expert Rev. Vaccines 2017, 16, 261–271. [Google Scholar] [CrossRef]
- Kümin, D.; Hofmann, C.; Rudolph, M.; Both, G.W.; Löser, P. Biology of ovine adenovirus infection of nonpermissive cells. J. Virol. 2002, 76, 10882–10893. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, U.B.; Benchaibi, M.; Meyer, V.; Schlesinger, Y.; Schughart, K. Novel human gene transfer vectors: Evaluation of wild-type and recombinant animal adenoviruses in human-derived cells. Hum. Gene Ther. 1999, 10, 2587–2599. [Google Scholar] [CrossRef]
- De Vleeschauwer, A.R.; Zhou, X.; Lefebvre, D.J.; Garnier, A.; Watier, F.; Pignon, C.; Lacour, S.A.; Zientara, S.; Bakkali-Kassimi, L.; de Clercq, K.; et al. A canine adenovirus type 2 vaccine vector confers protection against foot-and-mouth disease in guinea pigs. Vaccine 2018, 36, 2193–2198. [Google Scholar] [CrossRef]
- Chen, E.C.; Yagi, S.; Kelly, K.R.; Mendoza, S.P.; Maninger, N.; Rosenthal, A.; Spinner, A.; Bales, K.L.; Schnurr, D.P.; Lerche, N.W.; et al. Cross-species transmission of a novel adenovirus associated with a fulminant pneumonia outbreak in a new world monkey colony. PLoS Pathog. 2011, 7, e1002155. [Google Scholar] [CrossRef]
- Pérez-Illana, M.; Martínez, M.; Condezo, G.N.; Hernando-Pérez, M.; Mangroo, C.; Brown, M.; Marabini, R.; Martín, C.S. Cryo-EM structure of enteric adenovirus HAdV-F41 highlights structural variations among human adenoviruses. Sci. Adv. 2021, 7, 1–15. [Google Scholar] [CrossRef]
- Berciaud, S.; Rayne, F.; Kassab, S.; Jubert, C.; Faure-Della Corte, M.; Salin, F.; Wodrich, H.; Lafon, M.E.; Barat, P.; Fayon, M.; et al. Adenovirus infections in Bordeaux University Hospital 2008–2010: Clinical and virological features. J. Clin. Virol. 2012, 54, 302–307. [Google Scholar] [CrossRef] [PubMed]
- Khanal, S.; Ghimire, P.; Dhamoon, A.S. The repertoire of adenovirus in human disease: The innocuous to the deadly. Biomedicines 2018, 6, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynch, J.P.; Kajon, A.E. Adenovirus: Epidemiology, global spread of novel serotypes, and advances in treatment and prevention. Semin. Respir. Crit. Care Med. 2016, 37, 586–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, C.M.; Seto, D.; Jones, M.S.; Dyer, D.W.; Chodosh, J. Molecular evolution of human species D adenoviruses. Infect. Genet. Evol. 2011, 11, 1208–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greber, U.F. Disassembly: The adenovirus. Rev. Med. Virol. 1998, 222, 213–222. [Google Scholar] [CrossRef]
- Chroboczek, J.; Bieber, F.; Jacrot, B. The sequence of the genome of adenovirus type 5 and its comparison with the genome of adenovirus type 2. Virology 1992, 186, 280–285. [Google Scholar] [CrossRef]
- El Bakkouri, M.; Seiradake, E.; Cusack, S.; Ruigrok, R.W.H.; Schoehn, G. Structure of the C-terminal head domain of the fowl adenovirus type 1 short fibre. Virology 2008, 378, 169–176. [Google Scholar] [CrossRef] [Green Version]
- Nemerow, G.R.; Stewart, P.L.; Reddy, V.S. Structure of human adenovirus. Curr. Opin. Virol. 2012, 2, 115–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, P.L.; Burnett, R.M.; Cyrklaff, M.; Fuller, S.D. Image reconstruction reveals the complex molecular organization of adenovirus. Cell 1991, 67, 145–154. [Google Scholar] [CrossRef]
- Fabry, C.M.S.; Rosa-Calatrava, M.; Conway, J.F.; Zubieta, C.; Cusack, S.; Ruigrok, R.W.H.; Schoehn, G. A quasi-atomic model of human adenovirus type 5 capsid. EMBO J. 2005, 24, 1645–1654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Jin, L.; Koh, S.B.S.; Atanasov, I.; Schein, S.; Wu, L.; Zhou, Z.H. Atomic structure of human adenovirus by Cryo-EM reveals interactions among protein networks. Science 2010, 329, 1038–1043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vassal-Stermann, E.; Effantin, G.; Zubieta, C.; Burmeister, W.; Iseni, F.; Wang, H.; Lieber, A.; Schoehn, G.; Fender, P. CryoEM structure of adenovirus type 3 fibre with desmoglein 2 shows an unusual mode of receptor engagement. Nat. Commun. 2019, 10, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Martín, C.S. Latest insights on adenovirus structure and assembly. Viruses 2012, 4, 847–877. [Google Scholar] [CrossRef] [Green Version]
- Reddy, V.S.; Barry, M.A. Structural organization and protein-protein interactions in human adenovirus capsid. In Subcellular Biochemistry; Springer Science and Business Media: Berlin, Germany, 2021; Volume 96, pp. 503–518. [Google Scholar]
- Cao, C.; Dong, X.; Wu, X.; Wen, B.; Ji, G.; Cheng, L.; Liu, H. Conserved fiber-penton base interaction revealed by nearly atomic resolution cryo-electron microscopy of the structure of adenovirus provides insight into receptor interaction. J. Virol. 2012, 86, 12322–12329. [Google Scholar] [CrossRef] [Green Version]
- Zubieta, C.; Schoehn, G.; Chroboczek, J.; Cusack, S. The structure of the human adenovirus 2 penton. Mol. Cell 2005, 17, 121–135. [Google Scholar] [CrossRef] [PubMed]
- Furcinitti, P.S.; van Oostrum, J.; Burnett, R.M. Adenovirus polypeptide IX revealed as capsid cement by difference images from electron microscopy and crystallography. EMBO J. 1989, 8, 3563–3570. [Google Scholar] [CrossRef] [PubMed]
- Wodrich, H.; Guan, T.; Cingolani, G.; von Seggern, D.; Nemerow, G.; Gerace, L. Switch from capsid protein import to adenovirus assembly by cleavage of nuclear transport signals. EMBO J. 2003, 22, 6245–6255. [Google Scholar] [CrossRef] [Green Version]
- San Martín, C. Transmission electron microscopy and the molecular structure of icosahedral viruses. Arch. Biochem. Biophys. 2015, 581, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Wu, L.; Sun, R.; Zhou, Z.H. Atomic structures of minor proteins VI and VII in human adenovirus. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [Green Version]
- Russell, W.C. Adenoviruses: Update on structure and function. J. Gen. Virol. 2009, 90, 1–20. [Google Scholar] [CrossRef]
- Chatterjee, P.K.; Vayda, M.E.; Flint, S.J. Interactions among the three adenovirus core proteins. J. Virol. 1985, 55, 379–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez-Vargas, J.; Vaughan, R.C.; Houser, C.; Hastie, K.M.; Kao, C.C.; Nemerow, G.R. Isolation and characterization of the DNA and protein binding activities of adenovirus core protein V. J. Virol. 2014, 88, 9287–9296. [Google Scholar] [CrossRef] [Green Version]
- Bauer, M.; Gomez-Gonzalez, A.; Suomalainen, M.; Hemmi, S.; Greber, U.F. The E3 ubiquitin ligase Mind bomb 1 enhances nuclear import of viral DNA by inactivating a virion linchpin protein that suppresses exposure of virion pathogen-associated molecular patterns. bioRxiv 2020. [Google Scholar] [CrossRef]
- Wang, I.H.; Suomalainen, M.; Andriasyan, V.; Kilcher, S.; Mercer, J.; Neef, A.; Luedtke, N.W.; Greber, U.F. Tracking viral genomes in host cells at single-molecule resolution. Cell Host Microbe 2013, 14, 468–480. [Google Scholar] [CrossRef] [Green Version]
- Komatsu, T.; Dacheux, D.; Kreppel, F.; Nagata, K.; Wodrich, H. A method for visualization of incoming adenovirus chromatin complexes in fixed and living cells. PLoS ONE 2015, 10, e0137102. [Google Scholar] [CrossRef]
- Greber, U.F.; Suomalainen, M.; Stidwill, R.P.; Boucke, K.; Ebersold, M.W.; Helenius, A. The role of the nuclear pore complex in adenovirus DNA entry. EMBO J. 1997, 16, 5998–6007. [Google Scholar] [CrossRef] [Green Version]
- Karen, K.A.; Hearing, P. Adenovirus core protein VII protects the viral genome from a DNA damage response at early times after infection. J. Virol. 2011, 85, 4135–4142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avgousti, D.C.; Herrmann, C.; Kulej, K.; Pancholi, N.J.; Sekulic, N.; Petrescu, J.; Molden, R.C.; Blumenthal, D.; Paris, A.J.; Reyes, E.D.; et al. A core viral protein binds host nucleosomes to sequester immune danger signals. Nature 2016, 535, 173–177. [Google Scholar] [CrossRef] [Green Version]
- Avgousti, D.C.; della Fera, A.N.; Otter, C.J.; Herrmann, C.; Pancholi, N.J.; Weitzman, M.D. Adenovirus core protein VII downregulates the DNA damage response on the host genome. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parks, R.J. Adenovirus protein IX: A new look at an old protein. Mol. Ther. 2005, 11, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Gorman, J.J.; Wallis, T.P.; Whelan, D.A.; Shaw, J.; Both, G.W. LH3, a “homologue” of the mastadenoviral E1B 55-kDa protein is a structural protein of atadenoviruses. Virology 2005, 342, 159–166. [Google Scholar] [CrossRef] [Green Version]
- Colby, W.W.; Shenk, T. Adenovirus type 5 virions can be assembled in vivo in the absence of detectable polypeptide IX. J. Virol. 1981, 39, 977–980. [Google Scholar] [CrossRef] [Green Version]
- Ostapchuk, P.; Suomalainen, M.; Zheng, Y.; Boucke, K.; Greber, U.F.; Hearing, P. The adenovirus major core protein VII is dispensable for virion assembly but is essential for lytic infection. PLoS Pathog. 2017, 13, e1006455. [Google Scholar] [CrossRef] [Green Version]
- Cusack, S. Adenovirus complex structures. Curr. Opin. Struct. Biol. 2005, 15, 237–243. [Google Scholar] [CrossRef]
- Berk, A.J.; Lee, F.; Harrison, T.; Williams, J.; Sharp, P.A. Pre-early adenovirus 5 gene product regulates synthesis of early viral messenger RNAs. Cell 1979, 17, 935–944. [Google Scholar] [CrossRef]
- Zhang, W.; Imperiale, M.J. Requirement of the adenovirus IVa2 Protein for virus assembly. J. Virol. 2003, 77, 3586–3594. [Google Scholar] [CrossRef] [Green Version]
- Ahi, Y.S.; Mittal, S.K. Components of adenovirus genome packaging. Front. Microbiol. 2016, 7, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Lehmberg, E.; Traina, J.A.; Chakel, J.A.; Chang, R.J.; Parkman, M.; McCaman, M.T.; Murakami, P.K.; Lahidji, V.; Nelson, J.W.; Hancock, W.S.; et al. Reversed-phase high-performance liquid chromatographic assay for the adenovirus type 5 proteome. J. Chromatogr. B Biomed. Sci. Appl. 1999, 732, 411–423. [Google Scholar] [CrossRef]
- Stewart, P.L.; Fuller, S.D.; Burnett, R.M. Difference imaging of adenovirus: Bridging the resolution gap between X-ray crystallography and electron microscopy. EMBO J. 1993, 12, 2589–2599. [Google Scholar] [CrossRef] [PubMed]
- Rancourt, C.; Tihanyi, K.; Bourbonniere, M.; Weber, J.M. Identification of active-site residues of the adenovirus endopeptidase. Proc. Natl. Acad. Sci. USA 1994, 91, 844–847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mangel, W.F.; Martín, C.S. Structure, function and dynamics in adenovirus maturation. Viruses 2014, 6, 4536–4570. [Google Scholar] [CrossRef] [Green Version]
- Perez-Berna, A.J.; Mangel, W.F.; McGrath, W.J.; Graziano, V.; Flint, J.; San Martin, C. Processing of the L1 52/55k protein by the adenovirus protease: A new substrate and new insights into virion maturation. J. Virol. 2014, 88, 1513–1524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cotten, M.; Weber, J.M. The adenovirus protease is required for virus entry into host cells. Virology 1995, 213, 494–502. [Google Scholar] [CrossRef]
- Pérez-Berná, A.J.; Marabini, R.; Scheres, S.H.W.; Menéndez-Conejero, R.; Dmitriev, I.P.; Curiel, D.T.; Mangel, W.F.; Flint, S.J.; Martín, C.S. Structure and uncoating of immature adenovirus. J. Mol. Biol. 2009, 392, 547–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, E.; Pache, L.; von Seggern, D.J.; Mullen, T.-M.; Mikyas, Y.; Stewart, P.L.; Nemerow, G.R. Flexibility of the adenovirus fiber is required for efficient receptor interaction. J. Virol. 2003, 77, 7225–7235. [Google Scholar] [CrossRef] [Green Version]
- Seki, T.; Dmitriev, I.; Kashentseva, E.; Takayama, K.; Rots, M.; Suzuki, K.; Curiel, D.T. Artificial extension of the adenovirus fiber shaft inhibits infectivity in coxsackievirus and adenovirus receptor-positive cell lines. J. Virol. 2002, 76, 1100–1108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shayakhmetov, D.M.; Lieber, A. Dependence of adenovirus infectivity on length of the fiber shaft domain. J. Virol. 2000, 74, 10274–10286. [Google Scholar] [CrossRef] [Green Version]
- Fender, P.; Ruigrok, R.W.H.; Gout, E.; Buffet, S.; Chroboczek, J. Adenovirus dodecahedron, a new vector for human gene transfer. Nat. Biotechnol. 1997, 15, 52–56. [Google Scholar] [CrossRef]
- Fender, P.; Schoehn, G.; Foucaud-Gamen, J.; Gout, E.; Garcel, A.; Drouet, E.; Chroboczek, J. Adenovirus dodecahedron allows large multimeric protein transduction in human cells. J. Virol. 2003, 77, 4960–4964. [Google Scholar] [CrossRef] [Green Version]
- Pied, N.; Wodrich, H. Imaging the adenovirus infection cycle. FEBS Lett. 2019, 593, 3419–3448. [Google Scholar] [CrossRef] [Green Version]
- Nemerow, G.R.; Pache, L.; Reddy, V.; Stewart, P.L. Insights into adenovirus host cell interactions from structural studies. Virology 2009, 384, 380–388. [Google Scholar] [CrossRef] [Green Version]
- Kremer, E.J.; Nemerow, G.R. Adenovirus tales: From the cell surface to the nuclear pore complex. PLoS Pathog. 2015, 11, e1004821. [Google Scholar] [CrossRef] [PubMed]
- Philipson, L.; Lonberg-holm, K.; Pettersson, U.L.F. Virus-receptor interaction in an adenovirus system. J. Virol. 1968, 2, 1064–1075. [Google Scholar] [CrossRef] [Green Version]
- Bergelson, J.M.; Cunningham, J.A.; Droguett, G.; Kurt-Jones, E.A.; Krithivas, A.; Hong, J.S.; Horwitz, M.S.; Crowell, R.L.; Finberg, R.W. Isolation of a common receptor for coxsackie B viruses and adenoviruses 2 and 5. Science 1997, 275, 1320–1323. [Google Scholar] [CrossRef] [PubMed]
- Roelvink, P.W.; Lizonova, A.; Lee, J.G.M.; Li, Y.; Bergelson, J.M.; Finberg, R.W.; Brough, D.E.; Kovesdi, I.; Wickham, T.J. The coxsackievirus-adenovirus receptor protein can function as a cellular attachment protein for adenovirus serotypes from subgroups A, C, D, E, and F. J. Virol. 1998, 72, 7909–7915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mese, K.; Bunz, O.; Schellhorn, S.; Volkwein, W.; Jung, D.; Gao, J.; Zhang, W.; Baiker, A.; Ehrhardt, A. Identification of novel human adenovirus candidates using the coxsackievirus and adenovirus receptor for cell entry. Virol. J. 2020, 17, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Gaggar, A.; Shayakhmetov, D.M.; Lieber, A. CD46 is a cellular receptor for group B adenoviruses. Nat. Med. 2003, 9, 1408–1412. [Google Scholar] [CrossRef]
- Wang, H.; Li, Z.Y.; Liu, Y.; Persson, J.; Beyer, I.; Möller, T.; Koyuncu, D.; Drescher, M.R.; Strauss, R.; Zhang, X.B.; et al. Desmoglein 2 is a receptor for adenovirus serotypes 3, 7, 11 and 14. Nat. Med. 2011, 17, 96–104. [Google Scholar] [CrossRef]
- Arnberg, N.; Pring-Åkerblom, P.; Wadell, G. Adenovirus type 37 uses sialic acid as a cellular receptor on chang C cells. J. Virol. 2002, 76, 8834–8841. [Google Scholar] [CrossRef] [Green Version]
- Arnberg, N.; Kidd, A.H.; Edlund, K.; Nilsson, J.; Pring-Åkerblom, P.; Wadell, G. Adenovirus type 37 binds to cell surface sialic acid through a charge-dependent interaction. Virology 2002, 302, 33–43. [Google Scholar] [CrossRef] [Green Version]
- Arnberg, N.; Kidd, A.H.; Edlund, K.; Olfat, F.; Wadell, G. Initial interactions of subgenus D adenoviruses with A549 cellular receptors: Sialic acid versus αVintegrins. J. Virol. 2000, 74, 7691–7693. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Curiel, D.T. Fiber-modified adenoviruses for targeted gene therapy. Methods Mol. Biol. 2008, 434, 113–132. [Google Scholar] [CrossRef]
- Noureddini, S.C.; Curiel, D.T. Genetic targeting strategies for adenovirus. Mol. Pharm. 2005, 2, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, M.; Both, G.W.; Banizs, B.; Tsuruta, Y.; Yamamoto, S.; Kawakami, Y.; Douglas, J.T.; Tani, K.; Curiel, D.T.; Glasgow, J.N. An adenovirus serotype 5 vector with fibers derived from ovine atadenovirus demonstrates CAR-independent tropism and unique biodistribution in mice. Virology 2006, 350, 103–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wickham, T.J.; Mathias, P.; Cheresh, D.A.; Nemerow, G.R. Integrins aJS and U& promote adenovirus internalization but not virus attachment. Cell 1993, 73, 309–319. [Google Scholar] [PubMed]
- Bai, M.; Harfe, B.; Freimuth, P. Mutations that alter an Arg-Gly-Asp (RGD) sequence in the adenovirus type 2 penton base protein abolish its cell-rounding activity and delay virus reproduction in flat cells. J. Virol. 1993, 67, 5198–5205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albinsson, B.; Kidd, A.H. Adenovirus type 41 lacks an RGD α(v)-integrin binding motif on the penton base and undergoes delayed uptake in A549 cells. Virus Res. 1999, 64, 125–136. [Google Scholar] [CrossRef]
- Chiu, C.Y.; Mathias, P.; Nemerow, G.R.; Stewart, P.L. Structure of adenovirus complexed with its internalization receptor, αvβ5 integrin. J. Virol. 1999, 73, 6759–6768. [Google Scholar] [CrossRef] [Green Version]
- Li, E.; Stupack, D.; Bokoch, G.M.; Nemerow, G.R. Adenovirus endocytosis requires actin cytoskeleton reorganization mediated by rho family GTPases. J. Virol. 1998, 72, 8806–8812. [Google Scholar] [CrossRef] [Green Version]
- Li, E.; Stupack, D.G.; Brown, S.L.; Klemke, R.; Schlaepfer, D.D.; Nemerow, G.R. Association of p130(CAS) with phosphatidylinositol-3-OH kinase mediates adenovirus cell entry. J. Biol. Chem. 2000, 275, 14729–14735. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Huang, S.; Kapoor-Munshi, A.; Nemerow, G. Adenovirus internalization and infection require dynamin. J. Virol. 1998, 72, 3455–3458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meier, O.; Boucke, K.; Hammer, S.V.; Keller, S.; Stidwill, R.P.; Hemmi, S.; Greber, U.F. Adenovirus triggers macropinocytosis and endosomal leakage together with its clathrin-mediated uptake. J. Cell Biol. 2002, 158, 1119–1131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindert, S.; Silvestry, M.; Mullen, T.-M.; Nemerow, G.R.; Stewart, P.L. Cryo-electron microscopy structure of an adenovirus-integrin complex indicates conformational changes in both penton base and integrin. J. Virol. 2009, 83, 11491–11501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snijder, J.; Reddy, V.S.; May, E.R.; Roos, W.H.; Nemerow, G.R.; Wuite, G.J.L. Integrin and defensin modulate the mechanical properties of adenovirus. J. Virol. 2013, 87, 2756–2766. [Google Scholar] [CrossRef] [Green Version]
- Greber, U.F.; Willetts, M.; Webster, P.; Helenius, A. Stepwise dismantling of adenovirus 2 during entry into cells. Cell 1993, 75, 477–486. [Google Scholar] [CrossRef] [Green Version]
- Martin-Fernandez, M.; Longshaw, S.V.; Kirby, I.; Santis, G.; Tobin, M.J.; Clarke, D.T.; Jones, G.R. Adenovirus type-5 entry and disassembly followed in living cells by FRET, fluorescence anisotropy, and FLIM. Biophys. J. 2004, 87, 1316–1327. [Google Scholar] [CrossRef] [Green Version]
- Burckhardt, C.J.; Suomalainen, M.; Schoenenberger, P.; Boucke, K.; Hemmi, S.; Greber, U.F. Drifting motions of the adenovirus receptor CAR and immobile integrins initiate virus uncoating and membrane lytic protein exposure. Cell Host Microbe 2011, 10, 105–117. [Google Scholar] [CrossRef] [Green Version]
- Wiethoff, C.M.; Wodrich, H.; Gerace, L.; Nemerow, G.R. Adenovirus protein VI mediates membrane disruption following capsid disassembly. J. Virol. 2005, 79, 1992–2000. [Google Scholar] [CrossRef] [Green Version]
- Wodrich, H.; Henaff, D.; Jammart, B.; Segura-Morales, C.; Seelmeir, S.; Coux, O.; Ruzsics, Z.; Wiethoff, C.M.; Kremer, E.J. A capsid-encoded PPxY-motif facilitates adenovirus entry. PLoS Pathog. 2010, 6, e1000808. [Google Scholar] [CrossRef] [Green Version]
- Maier, O.; Galan, D.L.; Wodrich, H.; Wiethoff, C.M. An N-terminal domain of adenovirus protein VI fragments membranes by inducing positive membrane curvature. Virology 2010, 402, 11–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maier, O.; Wiethoff, C.M. N-terminal α-helix-independent membrane interactions facilitate adenovirus protein VI induction of membrane tubule formation. Virology 2010, 408, 31–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montespan, C.; Wiethoff, C.M.; Wodrich, H. A small viral PPxY peptide motif to control antiviral autophagy. J. Virol. 2017, 91, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montespan, C.; Marvin, S.A.; Austin, S.; Burrage, A.M.; Roger, B.; Rayne, F.; Faure, M.; Campell, E.M.; Schneider, C.; Reimer, R.; et al. Multi-layered control of Galectin-8 mediated autophagy during adenovirus cell entry through a conserved PPxY motif in the viral capsid. PLoS Pathog. 2017, 13, e1006217. [Google Scholar] [CrossRef]
- Daussy, C.F.; Wodrich, H. “Repair me if you can”: Membrane damage, response, and control from the viral perspective. Cells 2020, 9, 2042. [Google Scholar] [CrossRef]
- Gastaldelli, M.; Imelli, N.; Boucke, K.; Amstutz, B.; Meier, O.; Greber, U.F. Infectious adenovirus type 2 transport through early but not late endosomes. Traffic 2008, 9, 2265–2278. [Google Scholar] [CrossRef] [PubMed]
- Miyazawa, N.; Leopold, P.L.; Hackett, N.R.; Ferris, B.; Worgall, S.; Falck-Pedersen, E.; Crystal, R.G. Fiber swap between adenovirus subgroups B and C alters intracellular trafficking of adenovirus gene transfer vectors. J. Virol. 1999, 73, 6056–6065. [Google Scholar] [CrossRef] [Green Version]
- Shayakhmetov, D.M.; Li, Z.-Y.; Ternovoi, V.; Gaggar, A.; Gharwan, H.; Lieber, A. The interaction between the fiber knob domain and the cellular attachment receptor determines the intracellular trafficking route of adenoviruses. J. Virol. 2003, 77, 3712–3723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teigler, J.E.; Kagan, J.C.; Barouch, D.H. Late endosomal trafficking of alternative serotype adenovirus vaccine vectors augments antiviral innate immunity. J. Virol. 2014, 88, 10354–10363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barlan, A.U.; Griffin, T.M.; Mcguire, K.A.; Wiethoff, C.M. Adenovirus membrane penetration activates the NLRP3 inflammasome. J. Virol. 2011, 85, 146–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bremner, K.; Scherer, J.; Yi, J. Adenovirus transport through a direct cytoplasmic dynein-hexon interaction. Cell Host 2009, 6, 523–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leopold, P.L.; Kreitzer, G.; Miyazawa, N.; Rempel, S.; Pfister, K.K.; Rodriguez-Boulan, E.; Crystal, R.G. Dynein- and microtubule-mediated translocation of adenovirus serotype 5 occurs after endosomal lysis. Hum. Gene Ther. 2000, 11, 151–165. [Google Scholar] [CrossRef]
- Suomalainen, M.; Nakano, M.Y.; Keller, S.; Boucke, K.; Stidwill, R.P.; Greber, U.F. Microtubule-dependent plus- and minus end-directed motilities are competing processes for nuclear targeting of adenovirus. J. Cell Biol. 1999, 144, 657–672. [Google Scholar] [CrossRef] [Green Version]
- Scherer, J.; Vallee, R.B. Conformational changes in the adenovirus hexon subunit responsible for regulating cytoplasmic dynein recruitment. J. Virol. 2015, 89, 1013–1023. [Google Scholar] [CrossRef] [Green Version]
- Kelkar, S.A.; Pfister, K.K.; Crystal, R.G.; Leopold, P.L. Cytoplasmic dynein mediates adenovirus binding to microtubules. J. Virol. 2004, 78, 10122–10132. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Scherer, J.; Yi, J.; Vallee, R.B. Role of kinesins in directed adenovirus transport and cytoplasmic exploration. PLoS Pathog. 2018, 14, e1007055. [Google Scholar] [CrossRef]
- Trotman, L.C.; Mosberger, N.; Fornerod, M.; Stidwill, R.P.; Greber, U.F. Import of adenovirus DNA involves the nuclear pore complex receptor CAN/Nup214 and histone H1. Nat. Cell Biol. 2001, 3, 1092–1100. [Google Scholar] [CrossRef]
- Cassany, A.; Ragues, J.; Guan, T.; Bégu, D.; Wodrich, H.; Kann, M.; Nemerow, G.R.; Gerace, L. Nuclear import of adenovirus DNA involves direct interaction of hexon with an N-terminal domain of the nucleoporin Nup214. J. Virol. 2015, 89, 1719–1730. [Google Scholar] [CrossRef] [Green Version]
- Strunze, S.; Engelke, M.F.; Wang, I.H.; Puntener, D.; Boucke, K.; Schleich, S.; Way, M.; Schoenenberger, P.; Burckhardt, C.J.; Greber, U.F. Kinesin-1-mediated capsid disassembly and disruption of the nuclear pore complex promote virus infection. Cell Host Microbe 2011, 10, 210–223. [Google Scholar] [CrossRef] [Green Version]
- Puntener, D.; Engelke, M.F.; Ruzsics, Z.; Strunze, S.; Wilhelm, C.; Greber, U.F. Stepwise loss of fluorescent core protein V from human adenovirus during entry into cells. J. Virol. 2011, 85, 481–496. [Google Scholar] [CrossRef] [Green Version]
- Crisostomo, L.; Soriano, A.M.; Mendez, M.; Graves, D.; Pelka, P. Temporal dynamics of adenovirus 5 gene expression in normal human cells. PLoS ONE 2019, 14, e0211192. [Google Scholar] [CrossRef] [PubMed]
- Thomas, G.P.; Mathews, M.B. DNA replication and the early to late transition in adenovirus infection. Cell 1980, 22, 523–533. [Google Scholar] [CrossRef]
- Nevins, J.R. Mechanism of activation of early viral transcription by the adenovirus E1A gene product. Cell 1981, 26, 213–220. [Google Scholar] [CrossRef]
- Flint, S.J. Regulation of adenovirus mrna formation. Adv. Virus Res. 1986, 31, 169–228. [Google Scholar] [CrossRef]
- Nevins, J.R. Regulation of early adenovirus gene expression. Microbiol. Rev. 1987, 51, 419–430. [Google Scholar] [CrossRef] [PubMed]
- Farley, D.C.; Brown, J.L.; Leppard, K.N. Activation of the early-late switch in adenovirus type 5 major late transcription unit expression by L4 gene products. J. Virol. 2004, 78, 1782–1791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hidalgo, P.; Gonzalez, R.A. Formation of adenovirus DNA replication compartments. FEBS Lett. 2019, 593, 3518–3530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mondersert, G.; Tribouley, C.; Kedinger, C. Identification of a novel downstream bingind protein implicated in late-phase-specific activation of the adenovirus major late promoter. Nucleic Acids Res. 1992, 20, 3881–3889. [Google Scholar] [CrossRef] [PubMed]
- Shaw, A.R.; Ziff, E.B. Transcripts from the adenovirus-2 major late promoter yield a single early family of 3′ coterminal mRNAs and five late families. Cell 1980, 22, 905–916. [Google Scholar] [CrossRef]
- Weber, J.M.; Déry, C.V.; Amin Mirza, M.; Horvath, J. Adenovirus DNA synthesis is coupled to virus assembly. Virology 1985, 140, 351–359. [Google Scholar] [CrossRef]
- Tollefson, A.E.; Ryerse, J.S.; Scaria, A.; Hermiston, T.W.; Wold, W.S.M. The E3-11.6-kDa adenovirus death protein (ADP) is required for efficient cell death: Characterization of cells infected with adp mutants. Virology 1996, 220, 152–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tollefson, A.E.; Scaria, A.; Hermiston, T.W.; Ryerse, J.S.; Wold, L.J.; Wold, W.S. The adenovirus death protein (E3-11.6K) is required at very late stages of infection for efficient cell lysis and release of adenovirus from infected cells. J. Virol. 1996, 70, 2296–2306. [Google Scholar] [CrossRef] [Green Version]
- Weber, J. Genetic analysis of adenovirus type 2 III. Temperature sensitivity of processing viral proteins. J. Virol. 1976, 17, 462–471. [Google Scholar] [CrossRef] [Green Version]
- Greber, U.F.; Webster, P.; Weber, J.; Helenius, A. The role of the adenovirus protease in virus entry into cells. EMBO J. 1996, 15, 1766–1777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, E.K.; Nemerow, G.R.; Smith, J.G. Direct evidence from single-cell analysis that human α-defensins block adenovirus uncoating to neutralize infection. J. Virol. 2010, 84, 4041–4049. [Google Scholar] [CrossRef] [Green Version]
- Worgall, S.; Wolff, G.; Falck-Pedersen, E.; Crystal, R.G. Innate immune mechanisms dominate elimination of adenoviral vectors following in vivo administration. Hum. Gene Ther. 1997, 8, 37–44. [Google Scholar] [CrossRef]
- Yang, Y.; Nunes, F.A.; Berencsi, K.; Furth, E.E.; Gönczöl, E.; Wilson, J.M. Cellular immunity to viral antigens limits E1-deleted adenoviruses for gene therapy. Proc. Natl. Acad. Sci. USA 1994, 91, 4407–4411. [Google Scholar] [CrossRef] [Green Version]
- Tatsis, N.; Ertl, H.C.J. Adenoviruses as vaccine vectors. Mol. Ther. 2004, 10, 616–629. [Google Scholar] [CrossRef] [PubMed]
- Li, J.X.; Hou, L.H.; Meng, F.Y.; Wu, S.P.; Hu, Y.M.; Liang, Q.; Chu, K.; Zhang, Z.; Xu, J.J.; Tang, R.; et al. Immunity duration of a recombinant adenovirus type-5 vector-based Ebola vaccine and a homologous prime-boost immunisation in healthy adults in China: Final report of a randomised, double-blind, placebo-controlled, phase 1 trial. Lancet Glob. Health 2017, 5, e324–e334. [Google Scholar] [CrossRef] [Green Version]
- Raper, S.E.; Chirmule, N.; Lee, F.S.; Wivel, N.A.; Bagg, A.; Gao, G.P.; Wilson, J.M.; Batshaw, M.L. Fatal systemic inflammatory response syndrome in a ornithine transcarbamylase deficient patient following adenoviral gene transfer. Mol. Genet. Metab. 2003, 80, 148–158. [Google Scholar] [CrossRef] [PubMed]
- Schnell, M.A.; Zhang, Y.; Tazelaar, J.; Gao, G.P.; Yu, Q.C.; Qian, R.; Chen, S.J.; Varnavski, A.N.; LeClair, C.; Raper, S.E.; et al. Activation of innate immunity in nonhuman primates following intraportal administration of adenoviral vectors. Mol. Ther. 2001, 3, 708–722. [Google Scholar] [CrossRef] [PubMed]
- Hendrickx, R.; Stichling, N.; Koelen, J.; Kuryk, L.; Lipiec, A.; Greber, U.F. Innate immunity to adenovirus. Hum. Gene Ther. 2014, 25, 265–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atasheva, S.; Shayakhmetov, D.M. Innate Immune Response to Adenovirus Vector Administration in Vivo, 2nd ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2016; ISBN 9780128002766. [Google Scholar]
- Atasheva, S.; Yao, J.; Shayakhmetov, D.M. Innate immunity to adenovirus: Lessons from mice. FEBS Lett. 2019, 593, 3461–3483. [Google Scholar] [CrossRef] [PubMed]
- Gregory, S.M.; Nazir, S.A.; Metcalf, J.P. Implications of the innate immune response to adenovirus and adenoviral vectors. Future Virol. 2011, 6, 357–374. [Google Scholar] [CrossRef] [Green Version]
- Borgland, S.L.; Bowen, G.P.; Wong, N.C.W.; Libermann, T.A.; Muruve, D.A. Adenovirus vector-induced expression of the C-X-C chemokine IP-10 is mediated through capsid-dependent activation of NF-κB. J. Virol. 2000, 74, 3941–3947. [Google Scholar] [CrossRef] [Green Version]
- Bowen, G.P.; Borgland, S.L.; Lam, M.; Libermann, T.A.; Wong, N.C.W.; Muruve, D.A. Adenovirus vector-induced inflammation: Capsid-dependent induction of the C-C chemokine RANTES requires NF-κB. Hum. Gene Ther. 2002, 13, 367–379. [Google Scholar] [CrossRef]
- Higginbotham, J.N.; Seth, P.; Blaese, R.M.; Ramsey, W.J. The release of inflammatory cytokines from human peripheral blood mononuclear cells in vitro following exposure to adenovirus variants and capsid. Hum. Gene Ther. 2002, 13, 129–141. [Google Scholar] [CrossRef] [PubMed]
- Mccoy, R.D.; Roessler, B.J.; Huffnagle, G.A.R.Y.B.; Janich, S.L.; Laing, T.J.; Sevion, R.H. Pulmonary inflammation induced by incomplete or inactivated adenoviral particles. Hum. Gene Ther. 1995, 6, 1553–1560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muruve, D.A.; Barnes, M.J.; Stillman, I.E.; Libermann, T.A. Adenoviral gene therapy leads to rapid induction of multiple chemokines and acute neutrophil-dependent hepatic injury in vivo. Hum. Gene Ther. 1999, 10, 965–976. [Google Scholar] [CrossRef] [PubMed]
- Muruve, D.A. The innate immune response to adenovirus vectors. Hum. Gene Ther. 2004, 15, 1157–1166. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chirmule, N.; Gao, G.P.; Qian, R.; Croyle, M.; Joshi, B.; Tazelaar, J.; Wilson, J.M. Acute cytokine response to systemic adenoviral vectors in mice is mediated by dendritic cells and macrophages. Mol. Ther. 2001, 3, 697–707. [Google Scholar] [CrossRef] [PubMed]
- Tamanini, A.; Nicolis, E.; Bonizzato, A.; Bezzerri, V.; Melotti, P.; Assael, B.M.; Cabrini, G. Interaction of adenovirus type 5 fiber with the coxsackievirus and adenovirus receptor activates inflammatory response in human respiratory cells. J. Virol. 2006, 80, 11241–11254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farmer, C.; Morton, P.E.; Snippe, M.; Santis, G.; Parsons, M. Coxsackie adenovirus receptor (CAR) regulates integrin function through activation of p44/42 MAPK. Exp. Cell Res. 2009, 315, 2637–2647. [Google Scholar] [CrossRef]
- Iacobelli-Martinez, M.; Nepomuceno, R.R.; Connolly, J.; Nemerow, G.R. CD46-utilizing adenoviruses inhibit C/EBPβ-dependent expression of proinflammatory cytokines. J. Virol. 2005, 79, 11259–11268. [Google Scholar] [CrossRef] [Green Version]
- Di Paolo, N.C.; Miao, E.A.; Iwakura, Y.; Kaja, M.; Flavell, R.A.; Papayannopoulou, T.; Shayakhmetov, D.M. Virus sensing at the plasma membrane triggers interleukin-1α-mediated pro-inflammatory macrophage response in vivo. Thalia Stud. Lit. Humor 2010, 31, 110–121. [Google Scholar] [CrossRef]
- Suomalainen, M.; Nakano, M.Y.; Boucke, K.; Keller, S.; Greber, U.F. Adenovirus-activated PKA and p38/MAPK pathways boost microtubule-mediated nuclear targeting of virus. EMBO J. 2001, 20, 1310–1319. [Google Scholar] [CrossRef]
- Tibbles, L.A.; Spurrell, J.C.L.; Bowen, G.P.; Liu, Q.; Lam, M.; Zaiss, A.K.; Robbins, S.M.; Hollenberg, M.D.; Wickham, T.J.; Muruve, D.A. Activation of p38 and ERK signaling during adenovirus vector cell entry lead to expression of the C-X-C chemokine IP-10. J. Virol. 2002, 76, 1559–1568. [Google Scholar] [CrossRef] [Green Version]
- Li, E.; Stupack, D.; Klemke, R.; Cheresh, D.A.; Nemerow, G.R. Adenovirus endocytosis via αvintegrins requires phosphoinositide-3-OH kinase. J. Virol. 1998, 72, 2055–2061. [Google Scholar] [CrossRef] [Green Version]
- Philpott, N.J.; Nociari, M.; Elkon, K.B.; Falck-Pedersen, E. Adenovirus-induced maturation of dendritic cells through a PI3 kinase-mediated TNF-α induction pathway. Proc. Natl. Acad. Sci. USA 2004, 101, 6200–6205. [Google Scholar] [CrossRef] [Green Version]
- Zsengellér, Z.; Otake, K.; Hossain, S.-A.; Berclaz, P.-Y.; Trapnell, B.C. Internalization of adenovirus by alveolar macrophages initiates early proinflammatory signaling during acute respiratory tract infection. J. Virol. 2000, 74, 9655–9667. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Muruve, D.A. Molecular basis of the inflammatory response to adenovirus vectors. Gene Ther. 2003, 10, 935–940. [Google Scholar] [CrossRef] [Green Version]
- Cerullo, V.; Seiler, M.P.; Mane, V.; Brunetti-Pierri, N.; Clarke, C.; Bertin, T.K.; Rodgers, J.R.; Lee, B. Toll-like receptor 9 triggers an innate immune response to helper-dependent adenoviral vectors. Mol. Ther. 2007, 15, 378–385. [Google Scholar] [CrossRef]
- Fejer, G.; Drechsel, L.; Liese, J.; Schleicher, U.; Ruzsics, Z.; Imelli, N.; Greber, U.F.; Keck, S.; Hildenbrand, B.; Krug, A.; et al. Key role of splenic myeloid DCs in the IFN-αβ response to adenoviruses in vivo. PLoS Pathog. 2008, 4, e1000208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.; Huang, X.; Yang, Y. Innate immune response to adenoviral vectors is mediated by both toll-like receptor-dependent and -independent pathways. J. Virol. 2007, 81, 3170–3180. [Google Scholar] [CrossRef] [Green Version]
- Stein, S.C.; Falck-Pedersen, E. Sensing adenovirus infection: Activation of interferon regulatory factor 3 in RAW 264.7 cells. J. Virol. 2012, 86, 4527–4537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nociari, M.; Ocheretina, O.; Murphy, M.; Falck-Pedersen, E. Adenovirus induction of IRF3 occurs through a binary trigger targeting jun N-terminal kinase and TBK1 kinase cascades and type I interferon autocrine signaling. J. Virol. 2009, 83, 4081–4091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muruve, D.A.; Pétrilli, V.; Zaiss, A.K.; White, L.R.; Clark, S.A.; Ross, P.J.; Parks, R.J.; Tschopp, J. The inflammasome recognizes cytosolic microbial and host DNA and triggers an innate immune response. Nature 2008, 452, 103–107. [Google Scholar] [CrossRef]
- Lam, E.; Falck-Pedersen, E. Unabated adenovirus replication following activation of the cGAS/STING-dependent antiviral response in human cells. J. Virol. 2014, 88, 14426–14439. [Google Scholar] [CrossRef] [Green Version]
- Pahl, J.H.W.; Verhoeven, D.H.J.; Kwappenberg, K.M.C.; Vellinga, J.; Lankester, A.C.; van Tol, M.J.D.; Schilham, M.W. Adenovirus type 35, but not type 5, stimulates NK cell activation via plasmacytoid dendritic cells and TLR9 signaling. Mol. Immunol. 2012, 51, 91–100. [Google Scholar] [CrossRef]
- Wohlfart, C.E.; Svensson, U.K.; Everitt, E. Interaction between HeLa cells and adenovirus type 2 virions neutralized by different antisera. J. Virol. 1985, 56, 896–903. [Google Scholar] [CrossRef] [Green Version]
- Watson, G.; Burdon, M.G.; Russell, W.C. An antigenic analysis of the adenovirus type 2 fibre polypeptide. J. Gen. Virol. 1988, 69, 525–535. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Xiang, Z.Q.; Li, Y.; Kurupati, R.K.; Jia, B.; Bian, A.; Zhou, D.M.; Hutnick, N.; Yuan, S.; Gray, C.; et al. Adenovirus-based vaccines: Comparison of vectors from three species of adenoviridae. J. Virol. 2010, 84, 10522–10532. [Google Scholar] [CrossRef] [Green Version]
- Roberts, D.M.; Nanda, A.; Havenga, M.J.E.; Abbink, P.; Lynch, D.M.; Ewald, B.A.; Liu, J.; Thorner, A.R.; Swanson, P.E.; Gorgone, D.A.; et al. Hexon-chimaeric adenovirus serotype 5 vectors circumvent pre-existing anti-vector immunity. Nature 2006, 441, 239–243. [Google Scholar] [CrossRef]
- Sumida, S.M.; Truitt, D.M.; Lemckert, A.A.C.; Vogels, R.; Custers, J.H.H.V.; Addo, M.M.; Lockman, S.; Peter, T.; Peyerl, F.W.; Kishko, M.G.; et al. Neutralizing antibodies to adenovirus serotype 5 vaccine vectors are directed primarily against the adenovirus hexon protein. J. Immunol. 2005, 174, 7179–7185. [Google Scholar] [CrossRef] [Green Version]
- Bradley, R.R.; Lynch, D.M.; Iampietro, M.J.; Borducchi, E.N.; Barouch, D.H. Adenovirus serotype 5 neutralizing antibodies target both hexon and fiber following vaccination and natural infection. J. Virol. 2012, 86, 625–629. [Google Scholar] [CrossRef] [Green Version]
- Ahi, S.Y.; Bangari, D.S.; Mittal, S.K. Adenoviral vector immunity: Its implications and circumvention strategies. Curr. Gene Ther. 2011, 11, 307–320. [Google Scholar] [CrossRef]
- Mast, T.C.; Kierstead, L.; Gupta, S.B.; Nikas, A.A.; Kallas, E.G.; Novitsky, V.; Mbewe, B.; Pitisuttithum, P.; Schechter, M.; Vardas, E.; et al. International epidemiology of human pre-existing adenovirus (Ad) type-5, type-6, type-26 and type-36 neutralizing antibodies: Correlates of high Ad5 titers and implications for potential HIV vaccine trials. Vaccine 2010, 28, 950–957. [Google Scholar] [CrossRef]
- Zaiss, A.K.; Machado, H.B.; Herschman, H.R. The influence of innate and pre-existing immunity on adenovirus therapy. J. Cell. Biochem. 2009, 108, 778–790. [Google Scholar] [CrossRef] [Green Version]
- Colloca, S.; Barnes, E.; Folgori, A.; Ammendola, V.; Capone, S.; Cirillo, A.; Siani, L.; Naddeo, M.; Grazioli, F.; Esposito, M.L.; et al. Generation and screening of a large collection of novel simian Adenovirus allows the identification of vaccine vectors inducing potent cellular immunity in humans. Sci. Transl. Med. 2012, 4, 115ra2. [Google Scholar] [CrossRef] [Green Version]
- Nébié, I.; Edwards, N.J.; Tiono, A.B.; Ewer, K.J.; Sanou, G.S.; Soulama, I.; Sanon, S.; Diarra, A.; Yaro, J.B.; Kangoye, D.; et al. Assessment of chimpanzee adenovirus serotype 63 neutralizing antibodies prior to evaluation of a candidate malaria vaccine regimen based on viral vectors. Clin. Vaccine Immunol. 2014, 21, 901–903. [Google Scholar] [CrossRef] [Green Version]
- De Santis, O.; Audran, R.; Pothin, E.; Warpelin-Decrausaz, L.; Vallotton, L.; Wuerzner, G.; Cochet, C.; Estoppey, D.; Steiner-Monard, V.; Lonchampt, S.; et al. Safety and immunogenicity of a chimpanzee adenovirus-vectored Ebola vaccine in healthy adults: A randomised, double-blind, placebo-controlled, dose-finding, phase 1/2a study. Lancet Infect. Dis. 2016, 16, 311–320. [Google Scholar] [CrossRef] [Green Version]
- Zhu, F.C.; Guan, X.H.; Li, Y.H.; Huang, J.Y.; Jiang, T.; Hou, L.H.; Li, J.X.; Yang, B.F.; Wang, L.; Wang, W.J.; et al. Immunogenicity and safety of a recombinant adenovirus type-5-vectored COVID-19 vaccine in healthy adults aged 18 years or older: A randomised, double-blind, placebo-controlled, phase 2 trial. Lancet 2020, 396, 479–488. [Google Scholar] [CrossRef]
- Olive, M.; Eisenlohr, L.C.; Flomenberg, P. T-cell responses from healthy adults. Viral Immunol. 2001, 14, 403–413. [Google Scholar] [CrossRef]
- Flomenberg, P.; Piaskowski, V.; Truitt, R.L.; Casper, J.T. Characterization of human proliferative t cell responses to adenovirus. J. Infect. Dis. 1995, 171, 1090–1096. [Google Scholar] [CrossRef]
- Pédron, B.; Guérin, V.; Cordeiro, D.J.; Masmoudi, S.; Dalle, J.H.; Sterkers, G. Development of cytomegalovirus and adenovirus-specific memory CD4 T-cell functions from birth to adulthood. Pediatr. Res. 2011, 69, 106–111. [Google Scholar] [CrossRef] [Green Version]
- Flomenberg, P.; Piaskowski, V.; Truitt, R.L.; Casper, J.T. Human adenovirus-specific CD8+ T-cell responses are not inhibited by E3-19K in the presence of gamma interferon. J. Virol. 1996, 70, 6314–6322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leen, A.M.; Sili, U.; Savoldo, B.; Jewell, A.M.; Piedra, P.A.; Brenner, M.K.; Rooney, C.M. Fiber-modified adenoviruses generate subgroup cross-reactive, adenovirus-specific cytotoxic T lymphocytes for therapeutic applications. Blood 2004, 103, 1011–1019. [Google Scholar] [CrossRef] [Green Version]
- Smith, C.A.; Woodruff, L.S.; Rooney, C.; Kitchingman, G.R. Extensive cross-reactivity of cytotoxic T cells overview summary introduction. Hum. Gene Ther. 1998, 1427, 1419–1427. [Google Scholar] [CrossRef]
- Veltrop-Duits, L.A.; Heemskerk, B.; Sombroek, C.C.; van Vreeswijk, T.; Gubbels, S.; Toes, R.E.M.; Melief, C.J.M.; Franken, K.L.M.C.; Havenga, M.; van Tol, M.J.D.; et al. Human CD4+ T cells stimulated by conserved adenovirus 5 hexon peptides recognize cells infected with different species of human adenovirus. Eur. J. Immunol. 2006, 36, 2410–2423. [Google Scholar] [CrossRef]
- Zandvliet, M.L.; Falkenburg, J.h.F.; van Liempt, E.V.; Veltrop-Duits, L.A.; Lankester, A.C.; Kalpoe, J.S.; Kester, M.G.D.; van der Steen, D.M.; van Tol, M.J.; Willemze, R.; et al. Combined cd8+ and cd4+ adenovirus hexon-specific T cells associated with viral clearance after stem cell transplantation as treatment for adenovirus infection. Haematologica 2010, 95, 1943–1951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, T.C.; Dayball, K.; Wan, Y.H.; Bramson, J. Detailedanalysis of the CD8+ T-cell response followingadenovirusvaccination. J. Virol. 2003, 77, 13407–13411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teng, C.Y.; Millar, J.B.; Grinshtein, N.; Bassett, J.; Finn, J.; Bramson, J.L. T-cell immunity generated by recombinant adenovirus vaccines. Expert Rev. Vaccines 2007, 6, 347–356. [Google Scholar]
- Ouédraogo, A.; Tiono, A.B.; Kargougou, D.; Yaro, J.B.; Ouédraogo, E.; Kaboré, Y.; Kangoye, D.; Bougouma, E.C.; Gansane, A.; Henri, N.; et al. A phase 1b randomized, controlled, double-blinded dosage-escalation trial to evaluate the safety, reactogenicity and immunogenicity of an adenovirus type 35 based circumsporozoite malaria vaccine in burkinabe healthy adults 18 to 45 years of age. PLoS ONE 2013, 8, e78679. [Google Scholar] [CrossRef] [Green Version]
- Xiang, Z.Q.; Yang, Y.; Wilson, J.M.; Ertl, H.C.J. A replication-defective human adenovirus recombinant serves as a highly efficacious vaccine carrier. Virology 1996, 219, 220–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bassett, J.D.; Swift, S.L.; Bramson, J.L. Optimizing vaccine-induced CD8 + T-cell immunity: Focus on recombinant adenovirus vectors. Expert Rev. Vaccines 2011, 10, 1307–1319. [Google Scholar] [CrossRef] [PubMed]
- Tatsis, N.; Fitzgerald, J.C.; Reyes-Sandoval, A.; Harris-McCoy, K.C.; Hensley, S.E.; Zhou, D.; Lin, S.W.; Bian, A.; Zhi, Q.X.; Iparraguirre, A.; et al. Adenoviral vectors persist in vivo and maintain activated CD8+ T cells: Implications for their use as vaccines. Blood 2007, 110, 1916–1923. [Google Scholar] [CrossRef] [Green Version]
- Heemskerk, B.; Veltrop-Duits, L.A.; van Vreeswijk, T.; ten Dam, M.M.; Heidt, S.; Toes, R.E.M.; van Tol, M.J.D.; Schilham, M.W. extensive cross-reactivity of CD4+ adenovirus-specific T cells: Implications for immunotherapy and gene therapy. J. Virol. 2003, 77, 6562–6566. [Google Scholar] [CrossRef] [Green Version]
- Rock, K.L.; Reits, E.; Neefjes, J. Present yourself! By MHC class I and MHC class II molecules. Trends Immunol. 2016, 37, 724–737. [Google Scholar] [CrossRef] [Green Version]
- Münz, C. Autophagy beyond intracellular MHC class II antigen presentation. Trends Immunol. 2016, 37, 755–763. [Google Scholar] [CrossRef] [PubMed]
- Cadwell, K. Crosstalk between autophagy and inflammatory signalling pathways: Balancing defence and homeostasis. Nat. Rev. Immunol. 2016, 16, 661–675. [Google Scholar] [CrossRef] [PubMed]
- Weidberg, H.; Elazar, Z. TBK1 mediates crosstalk between the innate immune response and autophagy. Sci. Signal. 2011, 4, 1–4. [Google Scholar] [CrossRef]
- Deretic, V.; Levine, B. Autophagy balances inflammation in innate immunity. Autophagy 2018, 14, 243–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parzych, K.R.; Klionsky, D.J. An overview of autophagy: Morphology, mechanism, and regulation. Antioxid. Redox Signal. 2014, 20, 460–473. [Google Scholar] [CrossRef] [Green Version]
- Deretic, V. Autophagy as an immune defense mechanism. Curr. Opin. Immunol. 2006, 18, 375–382. [Google Scholar] [CrossRef] [PubMed]
- Münz, C. Autophagy proteins influence endocytosis for MHC restricted antigen presentation. Semin. Cancer Biol. 2020, 66, 110–115. [Google Scholar] [CrossRef]
- Nimmerjahn, F.; Milosevic, S.; Behrends, U.; Jaffee, E.M.; Pardoll, D.M.; Bornkamm, G.W.; Mautner, J. Major histocompatibility complex class II-restricted presentation of a cytosolic antigen by autophagy. Eur. J. Immunol. 2003, 33, 1250–1259. [Google Scholar] [CrossRef] [PubMed]
- Paludan, C.; Schmid, D.; Landthaler, M.; Vockerodt, M.; Kube, D.; Tuschl, T.; Münz, C. Endogenous MHC class II processing of a viral nuclear antigen after autophagy. Science 2005, 307, 593–596. [Google Scholar] [CrossRef]
- Leung, C.S.; Haigh, T.A.; Mackay, L.K.; Rickinson, A.B.; Taylor, G.S. Nuclear location of an endogenously expressed antigen, EBNA1, restricts access to macroautophagy and the range of CD4 epitope display. Proc. Natl. Acad. Sci. USA 2010, 107, 2165–2170. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.K.; Mattei, L.M.; Steinberg, B.E.; Alberts, P.; Lee, Y.H.; Chervonsky, A.; Mizushima, N.; Grinstein, S.; Iwasaki, A. In vivo requirement for Atg5 in antigen presentation by dendritic cells. Immunity 2010, 32, 227–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comber, J.D.; Robinson, T.M.; Siciliano, N.A.; Snook, A.E.; Eisenlohr, L.C. Functional macroautophagy induction by influenza a virus without a contribution to major histocompatibility complex class II-restricted presentation. J. Virol. 2011, 85, 6453–6463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanchet, F.P.; Moris, A.; Nikolic, D.S.; Lehmann, M.; Cardinaud, S.; Stalder, R.; Garcia, E.; Dinkins, C.; Leuba, F.; Wu, L.; et al. Human immunodeficiency virus-1 inhibition of immunoamphisomes in dendritic cells impairs early innate and adaptive immune responses. Immunity 2010, 32, 654–669. [Google Scholar] [CrossRef] [Green Version]
- Heckmann, B.L.; Green, D.R. LC3-associated phagocytosis at a glance. J. Cell Sci. 2019, 132, 1–2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romao, S.; Gasser, N.; Becker, A.C.; Guhl, B.; Bajagic, M.; Vanoaica, D.; Ziegler, U.; Roesler, J.; Dengjel, J.; Reichenbach, J.; et al. Autophagy proteins stabilize pathogen-containing phagosomes for prolonged MHC II antigen processing. J. Cell Biol. 2013, 203, 757–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tey, S.K.; Khanna, R. Autophagy mediates transporter associated with antigen processing-independent presentation of viral epitopes through MHC class I pathway. Blood 2012, 120, 994–1004. [Google Scholar] [CrossRef] [Green Version]
- Das, M.; Kaveri, S.V.; Bayry, J. Cross-presentation of antigens by dendritic cells: Role of autophagy. Oncotarget 2015, 6, 28527–28528. [Google Scholar] [CrossRef]
- Uhl, M.; Kepp, O.; Jusforgues-Saklani, H.; Vicencio, J.M.; Kroemer, G.; Albert, M.L. Autophagy within the antigen donor cell facilitates efficient antigen cross-priming of virus-specific CD8+ T cells. Cell Death Differ. 2009, 16, 991–1005. [Google Scholar] [CrossRef]
- Yi, Y.; Zhou, Z.; Shu, S.; Fang, Y.; Twitty, C.; Hilton, T.L.; Aung, S.; Urba, W.J.; Fox, B.A.; Hu, H.M.; et al. Autophagy-assisted antigen cross-presentation: Autophagosome as the argo of shared tumor-specifc antigens and DAMPs. Oncoimmunology 2012, 1, 976–978. [Google Scholar] [CrossRef] [Green Version]
- Crotzer, V.L.; Blum, J.S. Autophagy and its role in MHC-mediated antigen presentation. J. Immunol. 2009, 182, 3335–3341. [Google Scholar] [CrossRef] [Green Version]
- Windheim, M.; Hilgendorf, A.; Burgert, H.G. Immune evasion by adenovirus E3 proteins: Exploitation of intracellular trafficking pathways. Curr. Top. Microbiol. Immunol. 2004, 273, 29–85. [Google Scholar]
- Ginsberg, H.S. The ups and downs of adenovirus vectors. Bull. N. Y. Acad. Med. J. Urban. Health 1996, 73, 53–58. [Google Scholar]
- Bennett, E.M.; Bennink, J.R.; Yewdell, J.W.; Brodsky, F.M. Cutting edge: Adenovirus E19 has two mechanisms for affecting class I MHC expression. J. Immunol. 1999, 162, 5049–5052. [Google Scholar]
- Oliveira, E.R.A.; Bouvier, M. Immune evasion by adenoviruses: A window into host-virus adaptation. FEBS Lett. 2019, 593, 3496–3503. [Google Scholar] [CrossRef] [Green Version]
- Burgert, H.G. Subversion of the MHC class I antigen-presentation pathway by adenoviruses and herpes simplex viruses. Trends Microbiol. 1996, 4, 107–112. [Google Scholar] [CrossRef]
- Klein, S.R.; Jiang, H.; Hossain, M.B.; Fan, X.; Gumin, J.; Dong, A.; Alonso, M.M.; Gomez-Manzano, C.; Fueyo, J. Critical role of autophagy in the processing of adenovirus capsid-incorporated cancer-specific antigens. PLoS ONE 2016, 11, e0153814. [Google Scholar] [CrossRef] [PubMed]
- Neukirch, L.; Fougeroux, C.; Andersson, A.M.C.; Holst, P.J. The Potential of Adenoviral Vaccine Vectors with Altered Antigen Presentation Capabilities; Taylor & Francis: Milton Park, UK, 2020; Volume 19, ISBN 0000000275668. [Google Scholar]
- Tang, D.; Kang, R.; Livesey, K.M.; Cheh, C.W.; Farkas, A.; Loughran, P.; Hoppe, G.; Bianchi, M.E.; Tracey, K.J.; Zeh, H.J.; et al. Endogenous HMGB1 regulates autophagy. J. Cell Biol. 2010, 190, 881–892. [Google Scholar] [CrossRef] [PubMed]
- Degenhardt, K.; Mathew, R.; Beaudoin, B.; Bray, K.; Anderson, D.; Chen, G.; Mukherjee, C.; Shi, Y.; Gélinas, C.; Fan, Y.; et al. Autophagy promotes tumor cell survival and restricts necrosis, inflammation, and tumorigenesis. Cancer Cell 2006, 10, 51–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Jiang, L.; Fu, X.; Wang, W.; Ma, J.; Tian, T.; Nan, K.; Liang, X. Cytoplasmic liver kinase B1 promotes the growth of human lung adenocarcinoma by enhancing autophagy. Cancer Sci. 2018, 109, 3055–3067. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Martis, P.C.; Excoffon, K.J.D.A. Adenovirus transduction: More complicated than receptor expression. Virology 2017, 502, 144–151. [Google Scholar] [CrossRef]
- Tritel, M.; Stoddard, A.M.; Flynn, B.J.; Darrah, P.A.; Wu, C.; Wille, U.; Shah, J.A.; Huang, Y.; Xu, L.; Betts, M.R.; et al. Prime-boost vaccination with HIV-1 gag protein and cytosine phosphate guanosine oligodeoxynucleotide, followed by adenovirus, induces sustained and robust humoral and cellular immune responses. J. Immunol. 2003, 171, 2538–2547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ertl, H.C. Viral vectors as vaccine carriers. Curr. Opin. Virol. 2016, 21, 1–8. [Google Scholar] [CrossRef]
- Ginn, S.L.; Amaya, A.K.; Alexander, I.E.; Edelstein, M.; Abedi, M.R. Gene therapy clinical trials worldwide to 2017: An update. J. Gene Med. 2018, 20, e3015. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Mese, K.; Bunz, O.; Ehrhardt, A. State-of-the-art human adenovirus vectorology for therapeutic approaches. FEBS Lett. 2019, 593, 3609–3622. [Google Scholar] [CrossRef] [Green Version]
- McGrory, W.J.; Bautista, D.S.; Graham, F.L. A simple technique for the rescue of early region I mutations into infectious human adenovirus type 5. Virology 1988, 163, 614–617. [Google Scholar] [CrossRef]
- Wold, W.S.M.; Doronin, K.; Toth, K.; Kuppuswamy, M.; Lichtenstein, D.L.; Tollefson, A.E. Immune responses to adenoviruses: Viral evasion mechanisms and their implications for the clinic. Curr. Opin. Immunol. 1999, 11, 380–386. [Google Scholar] [CrossRef]
- Crystal, R.G. Adenovirus: The first effective in vivo gene delivery vector. Hum. Gene Ther. 2014, 25, 3–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bett, A.J.; Prevec, L.; Graham, F.L. Packaging capacity and stability of human adenovirus type 5 vectors. J. Virol. 1993, 67, 5911–5921. [Google Scholar] [CrossRef] [Green Version]
- Bett, A.J.; Haddara, W.; Prevec, L.; Graham, F.L. An efficient and flexible system for construction of adenovirus vectors with insertions or deletions in early regions 1 and 3. Proc. Natl. Acad. Sci. USA 1994, 91, 8802–8806. [Google Scholar] [CrossRef] [Green Version]
- Akusjärvi, G. Proteins with transcription regulatory properties encoded by human adenoviruses. Trends Microbiol. 1993, 1, 163–170. [Google Scholar] [CrossRef]
- Graham, F.L.; Smiley, J.; Russell, W.C.; Nairn, R. Characteristics of a human cell line transformed by DNA from human adenovirus type 5. J. Gen. Virol. 1977, 36, 59–72. [Google Scholar] [CrossRef] [PubMed]
- Shaw, G.; Morse, S.; Ararat, M.; Graham, F.L. Preferential transformation of human neuronal cells by human adenoviruses and the origin of HEK 293 cells. FASEB J. 2002, 16, 869–871. [Google Scholar] [CrossRef] [PubMed]
- Benihoud, K.; Yeh, P.; Perricaudet, M. Adenovirus vectors for gene delivery. Curr. Opin. Biotechnol. 1999, 10, 440–447. [Google Scholar] [CrossRef]
- Hehir, K.M.; Armentano, D.; Cardoza, L.M.; Choquette, T.L.; Berthelette, P.B.; White, G.A.; Couture, L.A.; Everton, M.B.; Keegan, J.; Martin, J.M.; et al. Molecular characterization of replication-competent variants of adenovirus vectors and genome modifications to prevent their occurrence. J. Virol. 1996, 70, 8459–8467. [Google Scholar] [CrossRef] [Green Version]
- Kreppel, F.; Hagedorn, C. Capsid and genome modification strategies to reduce the immunogenicity of adenoviral vectors. Int. J. Mol. Sci. 2021, 22, 2417. [Google Scholar] [CrossRef]
- Yang, Y.; Ertl, H.C.J.; Wilson, J.M. MHC class I-cestricted cytotoxic T lymphocytes to viral antigens destroy hepatocytes in mice infected with E1-deleted recombinant adenoviruses. Immunity 1994, 1, 433–442. [Google Scholar] [CrossRef]
- Engelhardt, J.F.; Litzky, L.; Wilson, J.M. Prolonged transgene expression in cotton rat lung with recombinant adenoviruses defective in E2a. Hum. Gene Ther. 1994, 5, 1217–1229. [Google Scholar] [CrossRef] [Green Version]
- Lusky, M.; Christ, M.; Rittner, K.; Dieterle, A.; Dreyer, D.; Mourot, B.; Schultz, H.; Stoeckel, F.; Pavirani, A.; Mehtali, M. In vitro and in vivo biology of recombinant adenovirus with E1, E1/E2A, or E1/E4 deleted. J. Virol. 1998, 72, 11. [Google Scholar] [CrossRef] [Green Version]
- Schaack, J. Induction and inhibition of innate inflammatory responses by adenovirus early region proteins. Viral Immunol. 2005, 18, 79–88. [Google Scholar] [CrossRef]
- Janssen, J.M.; Liu, J.; Skokan, J.; Gonçalves, M.A.F.V.; de Vries, A.A.F. Development of an AdEasy-based system to produce first- and second-generation adenoviral vectors with tropism for CAR- or CD46-positive cells. J. Gene Med. 2013, 15, 1–11. [Google Scholar] [CrossRef]
- Dedieu, J.F.; Vigne, E.; Torrent, C.; Jullien, C.; Mahfouz, I.; Caillaud, J.M.; Aubailly, N.; Orsini, C.; Guillaume, J.M.; Opolon, P.; et al. Long-term gene delivery into the livers of immunocompetent mice with E1/E4-defective adenoviruses. J. Virol. 1997, 71, 4626–4637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kochanek, S. High-capacity adenoviral vectors for gene transfer and somatic gene therapy. Hum. Gene Ther. 1999, 10, 2451–2459. [Google Scholar] [CrossRef] [PubMed]
- Segura, M.; Alba, R.; Bosch, A.; Chillon, M. Advances in helper-dependent adenoviral vector research. Curr. Gene Ther. 2008, 8, 222–235. [Google Scholar] [CrossRef]
- Brunetti-Pierri, N.; Ng, T.; Iannitti, D.; Cioffi, W.; Stapleton, G.; Law, M.; Breinholt, J.; Palmer, D.; Grove, N.; Rice, K.; et al. Transgene expression up to 7 years in nonhuman primates following hepatic transduction with helper-dependent adenoviral vectors. Hum. Gene Ther. 2013, 24, 761–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandig, V.; Youil, R.; Bett, A.J.; Franlin, L.L.; Oshima, M.; Maione, D.; Wang, F.; Metzker, M.L.; Savino, R.; Caskey, C.T. Optimization of the helper-dependent adenovirus system for production and potency in vivo. Proc. Natl. Acad. Sci. USA 2000, 97, 1002–1007. [Google Scholar] [CrossRef] [Green Version]
- Fisher, K.J.; Kelley, W.M.; Burda, J.F.; Wilson, J.M. A novel adenovirus-adeno-associated virus hybrid vector that displays efficient rescue and delivery of the AAV genome. Hum. Gene Ther. 1996, 7, 2079–2087. [Google Scholar] [CrossRef]
- Hardy, S.; Kitamura, M.; Harris-Stansil, T.; Dai, Y.; Phipps, M.L. Construction of adenovirus vectors through Cre-lox recombination. J. Virol. 1997, 71, 1842–1849. [Google Scholar] [CrossRef] [Green Version]
- Hartigan-O’Connor, D.; Amalfitano, A.; Chamberlain, J.S. Improved production of gutted adenovirus in cells expressing adenovirus preterminal protein and DNA polymerase. J. Virol. 1999, 73, 7835–7841. [Google Scholar] [CrossRef] [Green Version]
- Morral, N.; Parks, R.J.; Zhou, H.; Langston, C.; Schiedner, G.; Quinones, J.; Graham, F.L.; Kochanek, S.; Beaudet, A.L. High doses of a helper-dependent adenoviral vector yield supraphysiological levels of α1-antitrypsin with negligible toxicity. Hum. Gene Ther. 1998, 9, 2709–2716. [Google Scholar] [CrossRef]
- Schiedner, G.; Morral, N.; Parks, R.J.; Wu, Y.; Koopmans, S.C.; Langston, C.; Graham, F.L.; Beaudet, A.L.; Kochanek, S. Genomic DNA transfer with a high-capacity adenovirus vector results in improved in vivo gene expression and decreased toxicity. Nat. Genet. 1998, 18, 180–183. [Google Scholar] [CrossRef] [PubMed]
- Brunetti-Pierri, N.; Palmer, D.J.; Mane, V.; Finegold, M.; Beaudet, A.L.; Ng, P. Increased hepatic transduction with reduced systemic dissemination and proinflammatory cytokines following hydrodynamic injection of helper-dependent adenoviral vectors. Mol. Ther. 2005, 12, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Brunetti-Pierri, N.; Liou, A.; Patel, P.; Palmer, D.; Grove, N.; Finegold, M.; Piccolo, P.; Donnachie, E.; Rice, K.; Beaudet, A.; et al. Balloon catheter delivery of helper-dependent adenoviral vector results in sustained, therapeutic hFIX expression in rhesus macaques. Mol. Ther. 2012, 20, 1863–1870. [Google Scholar] [CrossRef] [Green Version]
- Ehrhardt, A.; Xu, H.; Dillow, A.M.; Bellinger, D.A.; Nichols, T.C.; Kay, M.A. A gene-deleted adenoviral vector results in phenotypic correction of canine hemophilia B without liver toxicity or thrombocytopenia. Blood 2003, 102, 2403–2411. [Google Scholar] [CrossRef] [Green Version]
- Top, F.H.; Buescher, E.L.; Bancroft, W.H.; Russell, P.K. Immunization with live types 7 and 4 adenovirus vaccines.Ii. antibody response and protective effect against acute respiratory disease due to adenovirus type 7. J. Infect. Dis. 1971, 124, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Deal, C.; Pekosz, A.; Ketner, G. Prospects for oral replicating adenovirus-vectored vaccines. Vaccine 2013, 31, 3236–3243. [Google Scholar] [CrossRef] [Green Version]
- Radin, J.M.; Hawksworth, A.W.; Blair, P.J.; Faix, D.J.; Raman, R.; Russell, K.L.; Gray, G.C. Dramatic decline of respiratory illness among us military recruits after the renewed use of adenovirus vaccines. Clin. Infect. Dis. 2014, 59, 962–968. [Google Scholar] [CrossRef] [PubMed]
- Broderick, M.; Myers, C.; Balansay, M.; Vo, S.; Osuna, A.; Russell, K. Adenovirus 4/7 vaccine’s effect on disease rates is associated with disappearance of adenovirus on building surfaces at a military recruit base. Mil. Med. 2017, 182, e2069–e2072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Home—ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ (accessed on 11 May 2021).
- Sullivan, N.J.; Sanchez, A.; Rollin, P.E.; Yang, Z.Y.; Nabel, G.J. Development of a preventive vaccine for Ebola virus infection in primates. Nature 2000, 408, 605–609. [Google Scholar] [CrossRef]
- Ewer, K.; Sebastian, S.; Spencer, A.J.; Gilbert, S.; Hill, A.V.S.; Lambe, T. Chimpanzee adenoviral vectors as vaccines for outbreak pathogens. Hum. Vaccines Immunother. 2017, 13, 3020–3032. [Google Scholar] [CrossRef]
- Priddy, F.H.; Brown, D.; Kublin, J.; Monahan, K.; Wright, D.P.; Lalezari, J.; Santiago, S.; Marmor, M.; Lally, M.; Novak, R.M.; et al. Safety and immunogenicity of a replication-incompetent adenovirus type 5 HIV-1 clade B gag/pol/nef vaccine in healthy adults. Clin. Infect. Dis. 2008, 46, 1769–1781. [Google Scholar] [CrossRef] [Green Version]
- Shiver, J.W.; Emini, E.A. Recent advances in the development of HIV-1 vaccines using replication-incompetent adenovirus vectors. Annu. Rev. Med. 2004, 55, 355–372. [Google Scholar] [CrossRef] [Green Version]
- Buchbinder, S.P.; Mehrotra, D.V.; Duerr, A.; Fitzgerald, D.W.; Mogg, R.; Li, D.; Gilbert, P.B.; Lama, J.R.; Marmor, M.; del Rio, C.; et al. Efficacy assessment of a cell-mediated immunity HIV-1 vaccine (the Step Study): A double-blind, randomised, placebo-controlled, test-of-concept trial. Lancet 2008, 372, 1881–1893. [Google Scholar] [CrossRef] [Green Version]
- Cohen, J. AIDS research: Promising AIDS vaccine’s failure leaves field reeling. Science 2007, 318, 28–29. [Google Scholar] [CrossRef] [PubMed]
- Press Release: Merck Halts Study of “Ineffective” HIV Vaccine|FierceBiotech. Available online: https://www.fiercebiotech.com/biotech/press-release-merck-halts-study-of-ineffective-hiv-vaccine (accessed on 11 May 2021).
- Harro, C.; Sun, X.; Stek, J.E.; Leavitt, R.Y.; Mehrotra, D.V.; Wang, F.; Bett, A.J.; Casimiro, D.R.; Shiver, J.W.; DiNubile, M.J.; et al. Safety and immunogenicity of the merck adenovirus serotype 5 (MRKAd5) and MRKAd6 human immunodeficiency virus type 1 trigene vaccines alone and in combination in healthy adults. Clin. Vaccine Immunol. 2009, 16, 1285–1292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harro, C.D.; Robertson, M.N.; Lally, M.A.; O’Neill, L.D.; Edupuganti, S.; Goepfert, P.A.; Mulligan, M.J.; Priddy, F.H.; Dubey, S.A.; Kierstead, L.S.; et al. Safety and immunogenicity of adenovirus-vectored near-consensus HIV type 1 clade B gag vaccines in healthy adults. AIDS Res. Hum. Retroviruses 2009, 25, 103–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patterson, L.J. The “STEP-Wise” Future of adenovirus-based HIV vaccines. Curr. Med. Chem. 2011, 18, 3981–3986. [Google Scholar] [CrossRef]
- Duerr, A.; Huang, Y.; Buchbinder, S.; Coombs, R.W.; Sanchez, J.; del Rio, C.; Casapia, M.; Santiago, S.; Gilbert, P.; Corey, L.; et al. Extended follow-up confirms early vaccine-enhanced risk of HIV acquisition and demonstrates waning effect over time among participants in a randomized trial of recombinant adenovirus HIV vaccine (Step Study). J. Infect. Dis. 2012, 206, 258–266. [Google Scholar] [CrossRef] [Green Version]
- Koblin, B.A.; Mayer, K.H.; Noonan, E.; Wang, C.Y.; Marmor, M.; Sanchez, J.; Brown, S.J.; Robertson, M.N.; Buchbinder, S.P. Sexual risk behaviors, circumcision status, and preexisting immunity to adenovirus type 5 among men who have sex with men participating in a randomized HIV-1 vaccine efficacy trial: Step study. J. Acquir. Immune Defic. Syndr. 2012, 60, 405–413. [Google Scholar] [CrossRef] [Green Version]
- Curlin, M.E.; Cassis-Ghavami, F.; Magaret, A.S.; Spies, G.A.; Duerr, A.; Celum, C.L.; Sanchez, J.L.; Margolick, J.B.; Detels, R.; McElrath, M.J.; et al. Serological immunity to adenovirus serotype 5 is not associated with risk of HIV infection: A case-control study. AIDS 2011, 25, 153–158. [Google Scholar] [CrossRef] [Green Version]
- Stephenson, K.E.; Hural, J.; Buchbinder, S.P.; Sinangil, F.; Barouch, D.H. Preexisting adenovirus seropositivity is not associated with increased HIV-1 acquisition in three HIV-1 vaccine efficacy trials. J. Infect. Dis. 2012, 205, 1806–1810. [Google Scholar] [CrossRef] [Green Version]
- Zak, D.E.; Andersen-Nissen, E.; Peterson, E.R.; Sato, A.; Hamilton, M.K.; Borgerding, J.; Krishnamurty, A.T.; Chang, J.T.; Adams, D.J.; Hensley, T.R.; et al. Merck Ad5/HIV induces broad innate immune activation that predicts CD8+ T-cell responses but is attenuated by preexisting Ad5 immunity. Proc. Natl. Acad. Sci. USA 2012, 109, E3503–E3512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Duerr, A.; Frahm, N.; Zhang, L.; Moodie, Z.; de Rosa, S.; Mcelrath, M.J.; Gilbert, P.B. Immune-correlates analysis of an HIV-1 vaccine efficacy trial reveals an association of nonspecific interferon-C secretion with increased HIV-1 infection risk: A cohort- Based modeling study. PLoS ONE 2014, 9, e108631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilbert, S.C. Adenovirus-vectored Ebola vaccines. Expert Rev. Vaccines 2015, 14, 1347–1357. [Google Scholar] [CrossRef] [PubMed]
- Vanderzanden, L.; Bray, M.; Fuller, D.; Roberts, T.; Custer, D.; Spik, K.; Jahrling, P.; Huggins, J.; Schmaljohn, A.; Schmaljohn, C. DNA vaccines expressing either the GP or NP genes of Ebola virus protect mice from lethal challenge. Virology 1998, 246, 134–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sullivan, N.J.; Geisbert, T.W.; Gelsbert, J.B.; Xu, L.; Yang, Z.Y.; Roederer, M.; Koup, R.A.; Jahrling, P.B.; Nabel, G.J. Accelerated vaccination for Ebola virus haemorrhagic fever in non-human primates. Nature 2003, 424, 681–684. [Google Scholar] [CrossRef]
- Sullivan, N.J.; Geisbert, T.W.; Geisbert, J.B.; Shedlock, D.J.; Xu, L.; Lamoreaux, L.; Custers, J.H.H.V.; Popernack, P.M.; Yang, Z.Y.; Pau, M.G.; et al. Immune protection of nonhuman primates against Ebola virus with single low-dose adenovirus vectors encoding modified GPs. PLoS Med. 2006, 3, e177. [Google Scholar] [CrossRef]
- Ledgerwood, J.E.; Costner, P.; Desai, N.; Holman, L.; Enama, M.E.; Yamshchikov, G.; Mulangu, S.; Hu, Z.; Andrews, C.A.; Sheets, R.A.; et al. A replication defective recombinant Ad5 vaccine expressing Ebola virus GP is safe and immunogenic in healthy adults. Vaccine 2010, 29, 304–313. [Google Scholar] [CrossRef]
- Quinn, K.M.; da Costa, A.; Yamamoto, A.; Berry, D.; Lindsay, R.W.B.; Darrah, P.A.; Wang, L.; Cheng, C.; Kong, W.-P.; Gall, J.G.D.; et al. Comparative analysis of the magnitude, quality, phenotype, and protective capacity of simian immunodeficiency virus gag-specific CD8 + T cells following human-, simian-, and chimpanzee-derived recombinant adenoviral vector immunization. J. Immunol. 2013, 190, 2720–2735. [Google Scholar] [CrossRef] [Green Version]
- Zhu, F.C.; Hou, L.H.; Li, J.X.; Wu, S.P.; Liu, P.; Zhang, G.R.; Hu, Y.M.; Meng, F.Y.; Xu, J.J.; Tang, R.; et al. Safety and immunogenicity of a novel recombinant adenovirus type-5 vector-based Ebola vaccine in healthy adults in China: Preliminary report of a randomised, double-blind, placebo-controlled, phase 1 trial. Lancet 2015, 385, 2272–2279. [Google Scholar] [CrossRef]
- Wu, S.; Kroeker, A.; Wong, G.; He, S.; Hou, L.; Audet, J.; Wei, H.; Zhang, Z.; Fernando, L.; Soule, G.; et al. An adenovirus vaccine expressing Ebola virus variant makona glycoprotein is efficacious in Guinea pigs and nonhuman primates. J. Infect. Dis. 2016, 214, S326–S332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, L.; Zhang, Z.; Gao, H.; Li, Y.; Hou, L.; Yao, H.; Wu, S.; Liu, J.; Wang, L.; Zhai, Y.; et al. Open-label phase I clinical trial of Ad5-EBOV in Africans in China. Hum. Vaccines Immunother. 2017, 13, 2078–2085. [Google Scholar] [CrossRef]
- Geisbert, T.W.; Bailey, M.; Hensley, L.; Asiedu, C.; Geisbert, J.; Stanley, D.; Honko, A.; Johnson, J.; Mulangu, S.; Pau, M.G.; et al. Recombinant adenovirus serotype 26 (Ad26) and Ad35 vaccine vectors bypass immunity to Ad5 and protect nonhuman primates against ebolavirus challenge. J. Virol. 2011, 85, 4222–4233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milligan, I.D.; Gibani, M.M.; Sewell, R.; Clutterbuck, E.A.; Campbell, D.; Plested, E.; Nuthall, E.; Voysey, M.; Silva-Reyes, L.; McElrath, M.J.; et al. Safety and immunogenicity of novel adenovirus type 26-and modified vaccinia Ankara-vectored Ebola vaccines: A randomized clinical trial. JAMA J. Am. Med. Assoc. 2016, 315, 1610–1623. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Kumar, R.; Agrawal, B. Adenoviral vector-based vaccines and gene therapies: Current status and future prospects. In Adenoviruses; IntechOpen: London, UK, 2019. [Google Scholar]
- Matz, K.M.; Marzi, A.; Feldmann, H. Ebola vaccine trials: Progress in vaccine safety and immunogenicity. Expert Rev. Vaccines 2019, 18, 1229–1242. [Google Scholar] [CrossRef] [PubMed]
- Kobinger, G.P.; Feldmann, H.; Zhi, Y.; Schumer, G.; Gao, G.; Feldmann, F.; Jones, S.; Wilson, J.M. Chimpanzee adenovirus vaccine protects against Zaire Ebola virus. Virology 2006, 346, 394–401. [Google Scholar] [CrossRef] [Green Version]
- Stanley, D.A.; Honko, A.N.; Asiedu, C.; Trefry, J.C.; Lau-Kilby, A.W.; Johnson, J.C.; Hensley, L.; Ammendola, V.; Abbate, A.; Grazioli, F.; et al. Chimpanzee adenovirus vaccine generates acute and durable protective immunity against ebolavirus challenge. Nat. Med. 2014, 20, 1126–1129. [Google Scholar] [CrossRef]
- Ledgerwood, J.E.; DeZure, A.D.; Stanley, D.A.; Coates, E.E.; Novik, L.; Enama, M.E.; Berkowitz, N.M.; Hu, Z.; Joshi, G.; Ploquin, A.; et al. Chimpanzee adenovirus vector ebola vaccine. N. Engl. J. Med. 2017, 376, 928–938. [Google Scholar] [CrossRef] [PubMed]
- Ewer, K.; Rampling, T.; Venkatraman, N.; Bowyer, G.; Wright, D.; Lambe, T.; Imoukhuede, E.B.; Payne, R.; Fehling, S.K.; Strecker, T.; et al. A monovalent chimpanzee adenovirus Ebola vaccine boosted with MVA. N. Engl. J. Med. 2016, 374, 1635–1646. [Google Scholar] [CrossRef]
- Kennedy, S.B.; Bolay, F.; Kieh, M.; Grandits, G.; Badio, M.; Ballou, R.; Eckes, R.; Feinberg, M.; Follmann, D.; Grund, B.; et al. Phase 2 placebo-controlled trial of two vaccines to prevent Ebola in Liberia. N. Engl. J. Med. 2017, 377, 1438–1447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mu, J.; Jeyanathan, M.; Shaler, C.R.; Horvath, C.; Damjanovic, D.; Zganiacz, A.; Kugathasan, K.; McCormick, S.; Xing, Z. Respiratory mucosal immunization with adenovirus gene transfer vector induces helper CD4 T cell-independent protective immunity. J. Gene Med. 2010, 12, 693–704. [Google Scholar] [CrossRef] [PubMed]
- Forbes, E.K.; Sander, C.; Ronan, E.O.; McShane, H.; Hill, A.V.S.; Beverley, P.C.L.; Tchilian, E.Z. Multifunctional, high-level cytokine-producing Th1 cells in the lung, but not spleen, correlate with protection against mycobacterium tuberculosis aerosol challenge in mice. J. Immunol. 2008, 181, 4955–4964. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Thorson, L.; Stokes, R.W.; Santosuosso, M.; Huygen, K.; Zganiacz, A.; Hitt, M.; Xing, Z. Single mucosal, but not parenteral, immunization with recombinant adenoviral-based vaccine provides potent protection from pulmonary tuberculosis. J. Immunol. 2004, 173, 6357–6365. [Google Scholar] [CrossRef]
- Smaill, F.; Jeyanathan, M.; Smieja, M.; Medina, M.F.; Thanthrige-Don, N.; Zganiacz, A.; Yin, C.; Heriazon, A.; Damjanovic, D.; Puri, L.; et al. A human type 5 adenovirus-based tuberculosis vaccine induces robust T cell responses in humans despite preexisting anti-adenovirus immunity. Sci. Transl. Med. 2013, 5. [Google Scholar] [CrossRef] [PubMed]
- Raviprakash, K.; Wang, D.; Ewing, D.; Holman, D.H.; Block, K.; Woraratanadharm, J.; Chen, L.; Hayes, C.; Dong, J.Y.; Porter, K. A tetravalent dengue vaccine based on a complex adenovirus vector provides significant protection in rhesus monkeys against all four serotypes of dengue virus. J. Virol. 2008, 82, 6927–6934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaiswal, S.; Khanna, N.; Swaminathan, S. Replication-defective adenoviral vaccine vector for the induction of immune responses to dengue virus type 2. J. Virol. 2003, 77, 12907–12913. [Google Scholar] [CrossRef] [Green Version]
- Khanam, S.; Pilankatta, R.; Khanna, N.; Swaminathan, S. An adenovirus type 5 (AdV5) vector encoding an envelope domain III-based tetravalent antigen elicits immune responses against all four dengue viruses in the presence of prior AdV5 immunity. Vaccine 2009, 27, 6011–6021. [Google Scholar] [CrossRef]
- Hung, P.P.; Chanda, P.K.; Natuk, R.J.; Mason, B.B.; Chengalvala, M.; Bhat, B.M.; Molnar-Kimber, K.L.; Dheer, S.K.; Morin, J.E.; Mizutani, S.; et al. Adenovirus vaccine strains genetically engineered to express HIV-1 or HBV antigens for use as live recombinant vaccines. Nat. Immun. Cell Growth Regul. 1990, 9, 160–164. [Google Scholar]
- Lubeck, M.D.; Davis, A.R.; Chengalvala, M.; Natuk, R.J.; Morin, J.E.; Molnar-Kimber, K.; Mason, B.B.; Bhat, B.M.; Mizutani, S.; Hung, P.P.; et al. Immunogenicity and efficacy testing in chimpanzees of an oral hepatitis B vaccine based on live recombinant adenovirus. Proc. Natl. Acad. Sci. USA 1989, 86, 6763–6767. [Google Scholar] [CrossRef] [Green Version]
- Makimura, M.; Miyake, S.; Akino, N.; Takamori, K.; Matsuura, Y.; Miyamura, T.; Saito, I. Induction of antibodies against structural proteins of hepatitis C virus in mice using recombinant adenovirus. Vaccine 1996, 14, 28–34. [Google Scholar] [CrossRef]
- Park, S.H.; Yang, S.H.; Lee, C.G.; Youn, J.W.; Chang, J.; Sung, Y.C. Efficient induction of T helper 1 CD4+ T-cell responses to hepatitis C virus core and E2 by a DNA prime-adenovirus boost. Vaccine 2003, 21, 4555–4564. [Google Scholar] [CrossRef]
- Matsui, M.; Moriya, O.; Akatsuka, T. Enhanced induction of hepatitis C virus-specific cytotoxic T lymphocytes and protective efficacy in mice by DNA vaccination followed by adenovirus boosting in combination with the interleukin-12 expression plasmid. Vaccine 2003, 21, 1629–1639. [Google Scholar] [CrossRef]
- Arribillaga, L.; de Cerio, A.L.D.; Sarobe, P.; Casares, N.; Gorraiz, M.; Vales, A.; Bruna-Romero, O.; Borrás-Cuesta, F.; Paranhos-Baccala, G.; Prieto, J.; et al. Vaccination with an adenoviral vector encoding hepatitis C virus (HCV) NS3 protein protects against infection with HCV-recombinant vaccinia virus. Vaccine 2002, 21, 202–210. [Google Scholar] [CrossRef]
- Seong, Y.R.; Choi, S.; Lim, J.S.; Lee, C.H.; Lee, C.K.; Im, D.S. Immunogenicity of the E1E2 proteins of hepatitis C virus expressed by recombinant adenoviruses. Vaccine 2001, 19, 2955–2964. [Google Scholar] [CrossRef]
- Prevec, L.; Campbell, J.B.; Christie, B.S.; Belbeck, L.; Graham, F.L. A recombinant human adenovirus vaccine against rabies. J. Infect. Dis. 1990, 161, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Vos, A.; Neubert, A.; Pommerening, E.; Müller, T.; Döhner, L.; Neubert, L.; Hughes, K. Immunogenicity of an E1-deleted recombinant human adenovirus against rabies by different routes of administration. J. Gen. Virol. 2001, 82, 2191–2197. [Google Scholar] [CrossRef] [PubMed]
- Bouet-Cararo, C.; Contreras, V.; Fournier, A.; Jallet, C.; Guibert, J.M.; Dubois, E.; Thiery, R.; Bréard, E.; Tordo, N.; Richardson, J.; et al. Canine adenoviruses elicit both humoral and cell-mediated immune responses against rabies following immunisation of sheep. Vaccine 2011, 29, 1304–1310. [Google Scholar] [CrossRef]
- Xiang, Z.Q.; Greenberg, L.; Ertl, H.C.; Rupprecht, C.E. Protection of non-human primates against rabies with an adenovirus recombinant vaccine. Virology 2014, 450–451, 243–249. [Google Scholar] [CrossRef] [Green Version]
- Van Kampen, K.R.; Shi, Z.; Gao, P.; Zhang, J.; Foster, K.W.; Chen, D.T.; Marks, D.; Elmets, C.A.; Tang, D.C.C. Safety and immunogenicity of adenovirus-vectored nasal and epicutaneous influenza vaccines in humans. Vaccine 2005, 23, 1029–1036. [Google Scholar] [CrossRef]
- Peters, W.; Brandl, J.R.; Lindbloom, J.D.; Martinez, C.J.; Scallan, C.D.; Trager, G.R.; Tingley, D.W.; Kabongo, M.L.; Tucker, S.N. Oral administration of an adenovirus vector encoding both an avian influenza A hemagglutinin and a TLR3 ligand induces antigen specific granzyme B and IFN-γ T cell responses in humans. Vaccine 2013, 31, 1752–1758. [Google Scholar] [CrossRef]
- Gurwith, M.; Lock, M.; Taylor, E.M.; Ishioka, G.; Alexander, J.; Mayall, T.; Ervin, J.E.; Greenberg, R.N.; Strout, C.; Treanor, J.J.; et al. Safety and immunogenicity of an oral, replicating adenovirus serotype 4 vector vaccine for H5N1 influenza: A randomised, double-blind, placebo-controlled, phase 1 study. Lancet Infect. Dis. 2013, 13, 238–250. [Google Scholar] [CrossRef] [Green Version]
- Gao, W.; Soloff, A.C.; Lu, X.; Montecalvo, A.; Nguyen, D.C.; Matsuoka, Y.; Robbins, P.D.; Swayne, D.E.; Donis, R.O.; Katz, J.M.; et al. Protection of mice and poultry from lethal H5N1 avian influenza virus through adenovirus-based immunization. J. Virol. 2006, 80, 1959–1964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Zhou, D. Adenoviral vector-based strategies against infectious disease and cancer. Hum. Vaccines Immunother. 2016, 12, 2064–2074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebola Virus Disease Democratic Republic of Congo: External Situation Report 94/2019. Available online: https://www.who.int/publications/i/item/10665-332654 (accessed on 12 May 2021).
- Der Li, Y.; Chi, W.Y.; Su, J.H.; Ferrall, L.; Hung, C.F.; Wu, T.C. Coronavirus vaccine development: From SARS and MERS to COVID-19. J. Biomed. Sci. 2020, 27, 104. [Google Scholar]
- Dai, L.; Gao, G.F. Viral targets for vaccines against COVID-19. Nat. Rev. Immunol. 2021, 21, 73–82. [Google Scholar] [CrossRef]
- Ou, X.; Liu, Y.; Lei, X.; Li, P.; Mi, D.; Ren, L.; Guo, L.; Guo, R.; Chen, T.; Hu, J.; et al. Characterization of spike glycoprotein of SARS-CoV-2 on virus entry and its immune cross-reactivity with SARS-CoV. Nat. Commun. 2020, 11, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Zhu, F.C.; Li, Y.H.; Guan, X.H.; Hou, L.H.; Wang, W.J.; Li, J.X.; Wu, S.P.; Wang, B.S.; Wang, Z.; Wang, L.; et al. Safety, tolerability, and immunogenicity of a recombinant adenovirus type-5 vectored COVID-19 vaccine: A dose-escalation, open-label, non-randomised, first-in-human trial. Lancet 2020, 395, 1845–1854. [Google Scholar] [CrossRef]
- Mercado, N.B.; Zahn, R.; Wegmann, F.; Loos, C.; Chandrashekar, A.; Yu, J.; Liu, J.; Peter, L.; McMahan, K.; Tostanoski, L.H.; et al. Single-shot Ad26 vaccine protects against SARS-CoV-2 in rhesus macaques. Nature 2020, 586, 583–588. [Google Scholar] [CrossRef]
- Van Doremalen, N.; Lambe, T.; Spencer, A.; Belij-Rammerstorfer, S.; Purushotham, J.N.; Port, J.R.; Avanzato, V.A.; Bushmaker, T.; Flaxman, A.; Ulaszewska, M.; et al. ChAdOx1 nCoV-19 vaccine prevents SARS-CoV-2 pneumonia in rhesus macaques. Nature 2020, 586, 578–582. [Google Scholar] [CrossRef] [PubMed]
- Graham, S.P.; McLean, R.K.; Spencer, A.J.; Belij-Rammerstorfer, S.; Wright, D.; Ulaszewska, M.; Edwards, J.C.; Hayes, J.W.P.; Martini, V.; Thakur, N.; et al. Evaluation of the immunogenicity of prime-boost vaccination with the replication-deficient viral vectored COVID-19 vaccine candidate ChAdOx1 nCoV-19. Vaccines 2020, 5. [Google Scholar] [CrossRef]
- Logunov, D.Y.; Dolzhikova, I.V.; Zubkova, O.V.; Tukhvatullin, A.I.; Shcheblyakov, D.V.; Dzharullaeva, A.S.; Grousova, D.M.; Erokhova, A.S.; Kovyrshina, A.V.; Botikov, A.G.; et al. Safety and immunogenicity of an rAd26 and rAd5 vector-based heterologous prime-boost COVID-19 vaccine in two formulations: Two open, non-randomised phase 1/2 studies from Russia. Lancet 2020, 396, 887–897. [Google Scholar] [CrossRef]
- Logunov, D.Y.; Dolzhikova, I.V.; Shcheblyakov, D.V.; Tukhvatulin, A.I.; Zubkova, O.V.; Dzharullaeva, A.S.; Kovyrshina, A.V.; Lubenets, N.L.; Grousova, D.M.; Erokhova, A.S.; et al. Safety and efficacy of an rAd26 and rAd5 vector-based heterologous prime-boost COVID-19 vaccine: An interim analysis of a randomised controlled phase 3 trial in Russia. Lancet 2021, 397, 671–681. [Google Scholar] [CrossRef]
- Lanzi, A.; Youssef, G.B.; Perricaudet, M.; Benihoud, K. Anti-adenovirus humoral responses influence on the efficacy of vaccines based on epitope display on adenovirus capsid. Vaccine 2011, 29, 1463–1471. [Google Scholar] [CrossRef] [PubMed]
- Vujadinovic, M.; Vellinga, J. Progress in adenoviral capsid-display vaccines. Biomedicines 2018, 6, 81. [Google Scholar] [CrossRef] [Green Version]
- Barouch, D.H.; Kik, S.V.; Weverling, G.J.; Dilan, R.; King, S.L.; Maxfield, L.F.; Clark, S.; Ng’ang’a, D.; Brandariz, K.L.; Abbink, P.; et al. International seroepidemiology of adenovirus serotypes 5, 26, 35, and 48 in pediatric and adult populations. Vaccine 2011, 29, 5203–5209. [Google Scholar] [CrossRef] [Green Version]
- Thorner, A.R.; Lemckert, A.A.C.; Goudsmit, J.; Lynch, D.M.; Ewald, B.A.; Denholtz, M.; Havenga, M.J.E.; Barouch, D.H. Immunogenicity of heterologous recombinant adenovirus prime-boost vaccine regimens is enhanced by circumventing vector cross-reactivity. J. Virol. 2006, 80, 12009–12016. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Chen, L.; Ding, Y.; Zhang, J.; Hua, J.; Geng, Q.; Ya, X.; Zeng, S.; Wu, J.; Jiang, Y.; et al. Viral etiology of medically attended influenza-like illnesses in children less than five years old in Suzhou, China, 2011–2014. J. Med. Virol. 2016, 88, 1334–1340. [Google Scholar] [CrossRef]
- Ma, J.; Duffy, M.R.; Deng, L.; Dakin, R.S.; Uil, T.; Custers, J.; Kelly, S.M.; McVey, J.H.; Nicklin, S.A.; Baker, A.H. Manipulating adenovirus hexon hypervariable loops dictates immune neutralisation and coagulation factor X-dependent cell interaction in vitro and in vivo. PLoS Pathog. 2015, 11, e1004673. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Huang, W.; Zhou, X.; Zhao, Q.; Wang, Q.; Jia, B. Seroprevalence of neutralizing antibodies to human adenoviruses type-5 and type-26 and chimpanzee adenovirus type-68 in healthy Chinese adults. J. Med. Virol. 2013, 85, 1077–1084. [Google Scholar] [CrossRef]
- McCoy, K.; Tatsis, N.; Korioth-Schmitz, B.; Lasaro, M.O.; Hensley, S.E.; Lin, S.-W.; Li, Y.; Giles-Davis, W.; Cun, A.; Zhou, D.; et al. Effect of preexisting immunity to adenovirus human serotype 5 antigens on the immune responses of nonhuman primates to vaccine regimens based on human- or chimpanzee-derived adenovirus vectors. J. Virol. 2007, 81, 6594–6604. [Google Scholar] [CrossRef] [Green Version]
- Moffatt, S.; Hays, J.; Hogenesch, H.; Mittal, S.K. Circumvention of vector-specific neutralizing antibody response by alternating use of human and non-human adenoviruses: Implications in gene therapy. Virology 2000, 272, 159–167. [Google Scholar] [CrossRef] [Green Version]
- Hammond, J.M.; McCoy, R.J.; Jansen, E.S.; Morrissy, C.J.; Hodgson, A.L.M.; Johnson, M.A. Vaccination with a single dose of a recombinant porcine adenovirus expressing the classical swine fever virus gp55 (E2) gene protects pigs against classical swine fever. Vaccine 2000, 18, 1040–1050. [Google Scholar] [CrossRef]
- Wüest, T.; Both, G.W.; Prince, A.M.; Hofmann, C.; Löser, P. Recombinant ovine atadenovirus induces a strong and sustained T cell response against the hepatitis C virus NS3 antigen in mice. Vaccine 2004, 22, 2717–2721. [Google Scholar] [CrossRef]
- Barlan, A.U.; Danthi, P.; Wiethoff, C.M. Lysosomal localization and mechanism of membrane penetration influence nonenveloped virus activation of the NLRP3 inflammasome. Virology 2011, 412, 306–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Defer, C.; Belin, M.T.; Caillet-Boudin, M.L.; Boulanger, P. Human adenovirus-host cell interactions: Comparative study with members of subgroups B and C. J. Virol. 1990, 64, 3661–3673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyazawa, N.; Crystal, R.G.; Leopold, P.L. Adenovirus serotype 7 retention in a late endosomal compartment prior to cytosol escape is modulated by fiber protein. J. Virol. 2001, 75, 1387–1400. [Google Scholar] [CrossRef] [Green Version]
- Iacobelli-Martinez, M.; Nemerow, G.R. Preferential activation of toll-like receptor nine by CD46-utilizing adenoviruses. J. Virol. 2007, 81, 1305–1312. [Google Scholar] [CrossRef] [Green Version]
- Campden, R.I.; Zhang, Y. The role of lysosomal cysteine cathepsins in NLRP3 inflammasome activation. Arch. Biochem. Biophys. 2019, 670, 32–42. [Google Scholar] [CrossRef]
- Kelley, N.; Jeltema, D.; Duan, Y.; He, Y. The NLRP3 inflammasome: An overview of mechanisms of activation and regulation. Int. J. Mol. Sci. 2019, 20, 3328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bossy-Wetzel, E.; Newmeyer, D.D.; Green, D.R. Mitochondrial cytochrome c release in apoptosis occurs upstream of DEVD-specific caspase activation and independently of mitochondrial transmembrane depolarization. EMBO J. 1998, 17, 37–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kupgan, G.; Hentges, D.C.; Muschinske, N.J.; Picking, W.D.; Picking, W.L.; Ramsey, J.D. The effect of fiber truncations on the stability of adenovirus type 5. Mol. Biotechnol. 2014, 56, 979–991. [Google Scholar] [CrossRef] [PubMed]
- Wiethoff, C.M.; Nemerow, G.R. Adenovirus membrane penetration: Tickling the tail of a sleeping dragon. Virology 2015, 479–480, 591–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nociari, M.; Ocheretina, O.; Schoggins, J.W.; Falck-Pedersen, E. Sensing infection by adenovirus: Toll-like receptor-independent viral DNA recognition signals activation of the interferon regulatory factor 3 master regulator. J. Virol. 2007, 81, 4145–4157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Favier, A.L.; Burmeister, W.P.; Chroboczek, J. Unique physicochemical properties of human enteric Ad41 responsible for its survival and replication in the gastrointestinal tract. Virology 2004, 322, 93–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- V’kovski, P.; Gultom, M.; Kelly, J.N.; Steiner, S.; Russeil, J.; Mangeat, B.; Cora, E.; Pezoldt, J.; Holwerda, M.; Kratzel, A.; et al. Disparate temperature-dependent virus-host dynamics for SARS-CoV-2 and SARS-CoV in the human respiratory epithelium. PLoS Biol. 2021, 19, e3001158. [Google Scholar] [CrossRef]
- Brown, M.; Wilson-Friesen, H.L.; Doane, F. A block in release of progeny virus and a high particle-to-infectious unit ratio contribute to poor growth of enteric adenovirus types 40 and 41 in cell culture. J. Virol. 1992, 66, 3198–3205. [Google Scholar] [CrossRef] [Green Version]
- Schoggins, J.W.; Nociari, M.; Philpott, N.; Falck-Pedersen, E. Influence of fiber detargeting on adenovirus-mediated innate and adaptive immune activation. J. Virol. 2005, 79, 11627–11637. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Veesler, D.; Campbell, M.G.; Barry, M.E.; Asturias, F.J.; Barry, M.A.; Reddy, V.S. Cryo-EM structure of human adenovirus D26 reveals the conservation of structural organization among human adenoviruses. Sci. Adv. 2017, 3, e1602670. [Google Scholar] [CrossRef] [Green Version]
- Abbink, P.; Lemckert, A.A.C.; Ewald, B.A.; Lynch, D.M.; Denholtz, M.; Smits, S.; Holterman, L.; Damen, I.; Vogels, R.; Thorner, A.R.; et al. Comparative seroprevalence and immunogenicity of six rare serotype recombinant adenovirus vaccine vectors from subgroups B and D. J. Virol. 2007, 81, 4654–4663. [Google Scholar] [CrossRef] [Green Version]
- Peruzzi, D.; Dharmapuri, S.; Cirillo, A.; Bruni, B.E.; Nicosia, A.; Cortese, R.; Colloca, S.; Ciliberto, G.; la Monica, N.; Aurisicchio, L. A novel Chimpanzee serotype-based adenoviral vector as delivery tool for cancer vaccines. Vaccine 2009, 27, 1293–1300. [Google Scholar] [CrossRef]
- Roy, S.; Gao, G.; Clawson, D.S.; Vandenberghe, L.H.; Farina, S.F.; Wilson, J.M. Complete nucleotide sequences and genome organization of four chimpanzee adenoviruses. Virology 2004, 324, 361–372. [Google Scholar] [CrossRef] [Green Version]
- Dicks, M.D.J.; Spencer, A.J.; Coughlan, L.; Bauza, K.; Gilbert, S.C.; Hill, A.V.S.; Cottingham, M.G. Differential immunogenicity between HAdV-5 and chimpanzee adenovirus vector ChAdOx1 is independent of fiber and penton RGD loop sequences in mice. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef]
- Kremer, E.J. CAR chasing: Canine adenovirus vectors—All bite and no bark? J. Gene Med. 2004, 6, S139–S151. [Google Scholar] [CrossRef] [PubMed]
- Soudais, C.; Laplace-Builhe, C.; Kissa, K.; Kremer, E.J. Preferential transduction of neurons by canine adenovirus vectors and their efficient retrograde transport in vivo. FASEB J. 2001, 15, 2283–2285. [Google Scholar] [CrossRef] [PubMed]
- Perreau, M.; Mennechet, F.; Serratrice, N.; Glasgow, J.N.; Curiel, D.T.; Wodrich, H.; Kremer, E.J. Contrasting effects of human, canine, and hybrid adenovirus vectors on the phenotypical and functional maturation of human dendritic cells: Implications for clinical efficacy. J. Virol. 2007, 81, 3272–3284. [Google Scholar] [CrossRef] [Green Version]
- Soudais, C.; Boutin, S.; Kremer, E.J. Characterization of cis-acting sequences involved in canine adenovirus packaging. Mol. Ther. 2001, 3, 631–640. [Google Scholar] [CrossRef] [PubMed]
- Matthews, D.A.; Russell, W.C. Adenovirus protein-protein interactions: Molecular parameters governing the binding of protein VI to hexon and the activation of the adenovirus 23K protease. J. Gen. Virol. 1995, 76, 1959–1969. [Google Scholar] [CrossRef] [PubMed]
- Hernando-Pérez, M.; Martín-González, N.; Pérez-Illana, M.; Suomalainen, M.; Condezo, G.N.; Ostapchuk, P.; Gallardo, J.; Menéndez, M.; Greber, U.F.; Hearing, P.; et al. Dynamic competition for hexon binding between core protein VII and lytic protein VI promotes adenovirus maturation and entry. Proc. Natl. Acad. Sci. USA 2020, 117, 13699–13707. [Google Scholar] [CrossRef]
- Moyer, C.L.; Besser, E.S.; Nemerow, G.R. A single maturation cleavage site in adenovirus impacts cell entry and capsid assembly. J. Virol. 2016, 90, 521–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Rosmalen, M.G.M.; Nemerow, G.R.; Wuite, G.J.L.; Roos, W.H. A single point mutation in precursor protein VI doubles the mechanical strength of human adenovirus. J. Biol. Phys. 2018, 44, 119–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moyer, C.L.; Nemerow, G.R. Disulfide-bond formation by a single cysteine mutation in adenovirus protein VI impairs capsid release and membrane lysis. Virology 2012, 428, 41–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, R.; Schellenberger, P.; Vasishtan, D.; Aknin, C.; Austin, S.; Dacheux, D.; Rayne, F.; Siebert, A.; Ruzsics, Z.; Gruenewald, K.; et al. The amphipathic helix of adenovirus capsid protein VI contributes to penton release and postentry sorting. J. Virol. 2015, 89, 2121–2135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moyer, C.L.; Wiethoff, C.M.; Maier, O.; Smith, J.G.; Nemerow, G.R. Functional genetic and biophysical analyses of membrane disruption by human adenovirus. J. Virol. 2011, 85, 2631–2641. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, H. Antigen presentation in vaccine development. Comp. Immunol. Microbiol. Infect. Dis. 2003, 26, 309–328. [Google Scholar] [CrossRef]
- Maier, O.; Marvin, S.A.; Wodrich, H.; Campbell, E.M.; Wiethoff, C.M. Spatiotemporal dynamics of adenovirus membrane rupture and endosomal escape. J. Virol. 2012, 86, 10821–10828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, F.; Gómez-Sintes, R.; Boya, P. Lysosomal membrane permeabilization and cell death. Traffic 2018, 19, 918–931. [Google Scholar] [CrossRef]
Figure 1.
Adenovirus cycle and associated immune response. Along its journey to the nucleus, Ad faces several cellular immune restrictions. Interaction of fiber and penton are required for initial attachment of Ad to cellular receptors; this interaction also induces a signaling cascade leading to the production of pro-inflammatory cytokines. Following entry, Ad is trapped inside an endosome where viral genomes can be detected via TLR9. The membrane lytic function of the viral protein VI allows viral penetration of the endosomal membrane, permitting viral escape into the cytosol. Endosomal escape occurs from within early or late endosome, depending on the Ad species. Membrane penetration of the viral capsid allows exposure of viral DNA in the cytosol that can be sensed by cGAS/STING. Membrane damage is also detected as a danger signal and induces both inflammasome and autophagy activation.
Figure 1.
Adenovirus cycle and associated immune response. Along its journey to the nucleus, Ad faces several cellular immune restrictions. Interaction of fiber and penton are required for initial attachment of Ad to cellular receptors; this interaction also induces a signaling cascade leading to the production of pro-inflammatory cytokines. Following entry, Ad is trapped inside an endosome where viral genomes can be detected via TLR9. The membrane lytic function of the viral protein VI allows viral penetration of the endosomal membrane, permitting viral escape into the cytosol. Endosomal escape occurs from within early or late endosome, depending on the Ad species. Membrane penetration of the viral capsid allows exposure of viral DNA in the cytosol that can be sensed by cGAS/STING. Membrane damage is also detected as a danger signal and induces both inflammasome and autophagy activation.

Figure 2.
MHC class II presentation by autophagy. Autophagy starts with the formation of a double membrane structure called phagophore that will elongate around the cargo to form a double-membrane vesicle called autophagosome. After fusion with lysosome, the cargo will be degraded in a so-called autolysosome. Further fusion of this vesicle with MHC class II compartment allows the loading of peptides for the ensuing MHC class II antigen presentation.
Figure 2.
MHC class II presentation by autophagy. Autophagy starts with the formation of a double membrane structure called phagophore that will elongate around the cargo to form a double-membrane vesicle called autophagosome. After fusion with lysosome, the cargo will be degraded in a so-called autolysosome. Further fusion of this vesicle with MHC class II compartment allows the loading of peptides for the ensuing MHC class II antigen presentation.

Figure 3.
Protein VI, a multifunctional viral protein determining the post-entry fate of the particle. (A) Domain organization of the membrane lytic protein VI. Protein VI is produced as a precursor form (pre_VI) and processed by the viral protease (AVP) at two sites. A role for the N-terminal part in capsid stability has been shown by two-point mutation (S28C and G33A). An amphipathic helix is responsible for the membrane lytic activity of protein VI, and a point mutation in this domain (L40Q) reduces the lytic activity. Protein VI encodes a conserved PPxY motif required for viral control of the cellular autophagy response against membrane penetration. Mutating the motif (PGAA) reduces viral infectivity. (B) Adenovirus membrane penetration via protein VI is recognized by the cell through glycan binding galectin 8 followed by autophagy induction. WT virus uses the PPxY motif in protein VI to recruit the ubiquitin ligase NEDD4.2 to suppress or halt autophagy until endosomal escape is completed. A mutation in this motif (PGAA) abolishes viral autophagy control and renders the mutant virus (M1) particle susceptible to autophagic degradation and enhanced antigenic presentation.
Figure 3.
Protein VI, a multifunctional viral protein determining the post-entry fate of the particle. (A) Domain organization of the membrane lytic protein VI. Protein VI is produced as a precursor form (pre_VI) and processed by the viral protease (AVP) at two sites. A role for the N-terminal part in capsid stability has been shown by two-point mutation (S28C and G33A). An amphipathic helix is responsible for the membrane lytic activity of protein VI, and a point mutation in this domain (L40Q) reduces the lytic activity. Protein VI encodes a conserved PPxY motif required for viral control of the cellular autophagy response against membrane penetration. Mutating the motif (PGAA) reduces viral infectivity. (B) Adenovirus membrane penetration via protein VI is recognized by the cell through glycan binding galectin 8 followed by autophagy induction. WT virus uses the PPxY motif in protein VI to recruit the ubiquitin ligase NEDD4.2 to suppress or halt autophagy until endosomal escape is completed. A mutation in this motif (PGAA) abolishes viral autophagy control and renders the mutant virus (M1) particle susceptible to autophagic degradation and enhanced antigenic presentation.


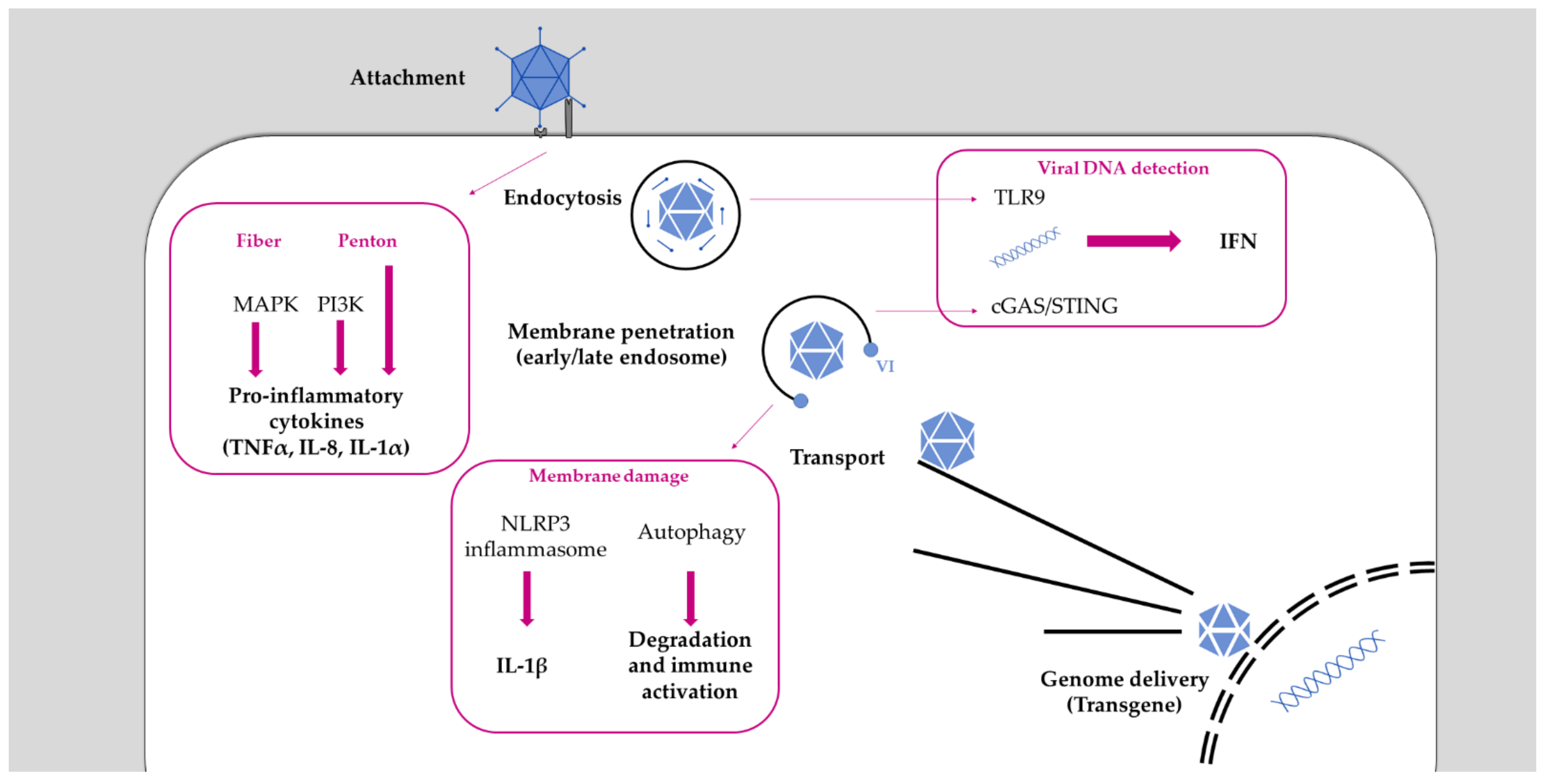{kind=link}
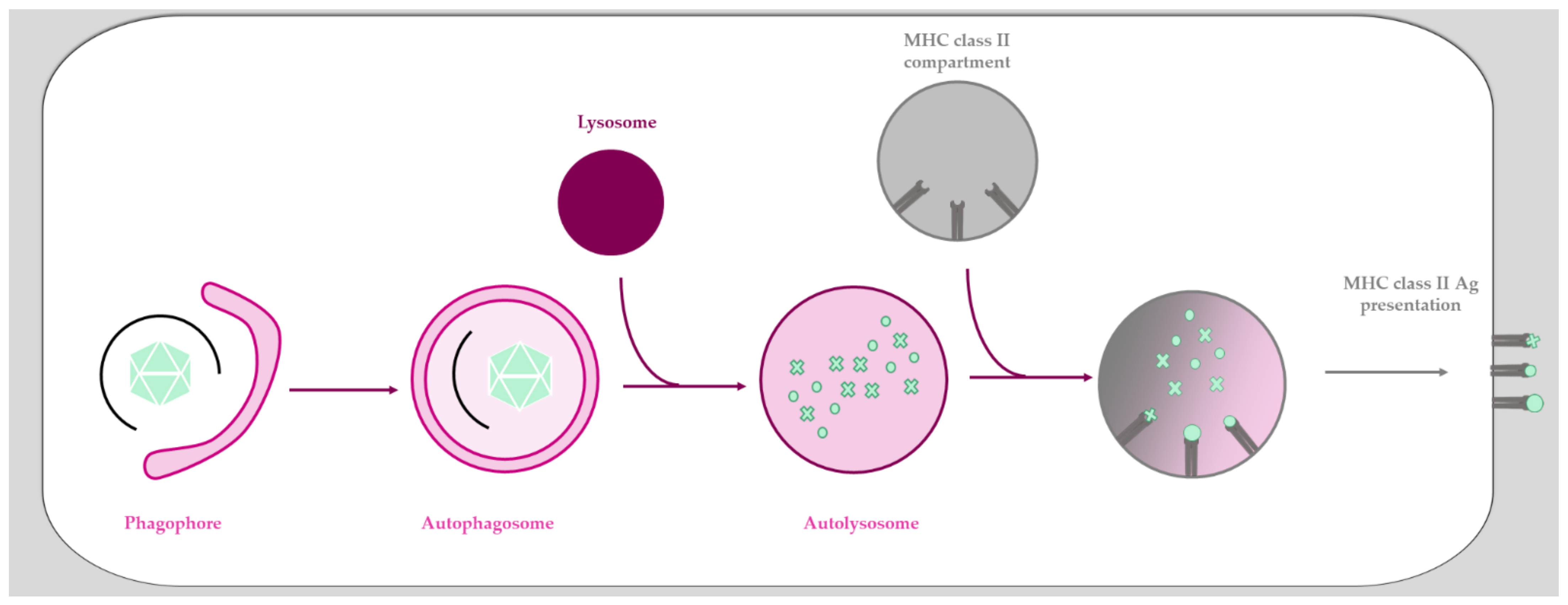{kind=link}
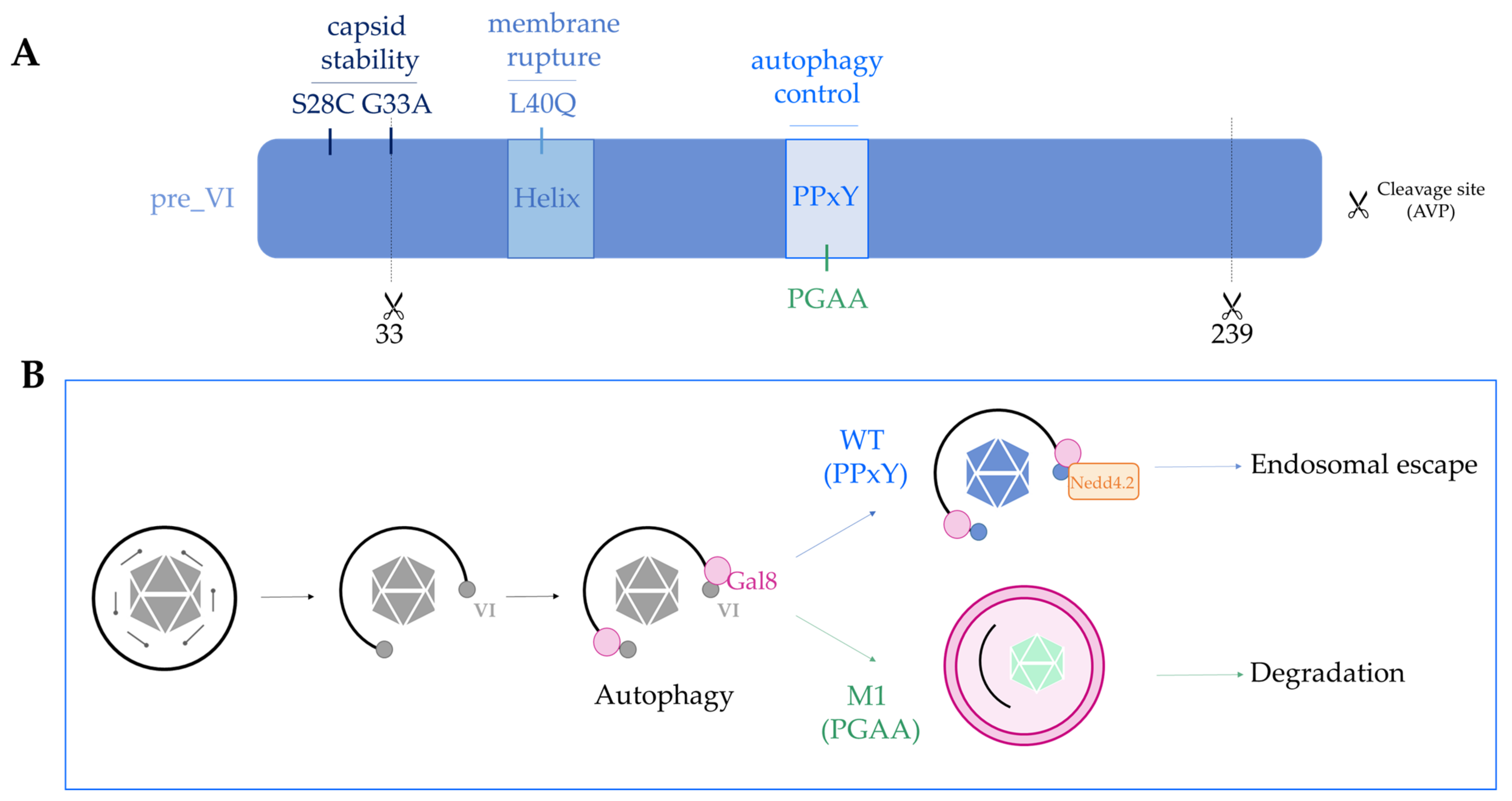{kind=link}
Table 1.
A simplified overview of human Ads and non-human Ads and their uses for vaccination purposes. Species of human Ads, their receptor, and main tropism.
Table 1.
A simplified overview of human Ads and non-human Ads and their uses for vaccination purposes. Species of human Ads, their receptor, and main tropism.
| Species | Type | Tropism | Receptor | Remarks |
|---|---|---|---|---|
| A | 12, 18 and 31 | Intestinal | CAR | |
| B | B1 : 3, 7, 16, 21 and 50 B2 : 11, 14, 34 and 35 | Respiratory and ocular | CD46/DSG-2 | |
| C | 1, 2, 5 and 6 | Respiratory | CAR | Ad5 : used as vaccine against SARS-CoV-2 and in clinical trial against Ebola and HIV-1 (but prematurely stopped) |
| D | 8-10, 13, 15, 17, 19, 20, 22-30, 32, 33, 36-39, 42-49 | Ocular and intestinal | CAR | Ad26 : used vaccine against Ebola and SARS-CoV-2 |
| E | 4 | Respiratory and ocular | CAR | |
| F | 40,41 | Intestinal | CAR | Oral vaccine |
| G | 52 | Intestinal | CAR/sialic acid | |
| Non-human (Chimpanzee) (Canine) | ChAd3 ChAdOx1 CAV-2 | — | CAR | ChAd3 : used as a vaccine in a clinical trial against Ebola ChAdOx1 : used as a vaccine against SARS-CoV-2 CAV-2 : can target neuronal cells |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Daussy, C.F.; Pied, N.; Wodrich, H. Understanding Post Entry Sorting of Adenovirus Capsids; A Chance to Change Vaccine Vector Properties. Viruses 2021, 13, 1221. https://doi.org/10.3390/v13071221
AMA Style
Daussy CF, Pied N, Wodrich H. Understanding Post Entry Sorting of Adenovirus Capsids; A Chance to Change Vaccine Vector Properties. Viruses. 2021; 13(7):1221. https://doi.org/10.3390/v13071221
Chicago/Turabian StyleDaussy, Coralie F., Noémie Pied, and Harald Wodrich. 2021. "Understanding Post Entry Sorting of Adenovirus Capsids; A Chance to Change Vaccine Vector Properties" Viruses 13, no. 7: 1221. https://doi.org/10.3390/v13071221
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.