
Are There Benefits from Thermal Bacteria for Health? The Hydrogenome Role

 , ,
, ,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling Site
2.2. Sample Collection
2.3. Physicochemical Analysis
2.4. DNA Extraction and 16S Amplicon Sequencing
2.5. Bioinformatic Analysis
3. Results and Discussion
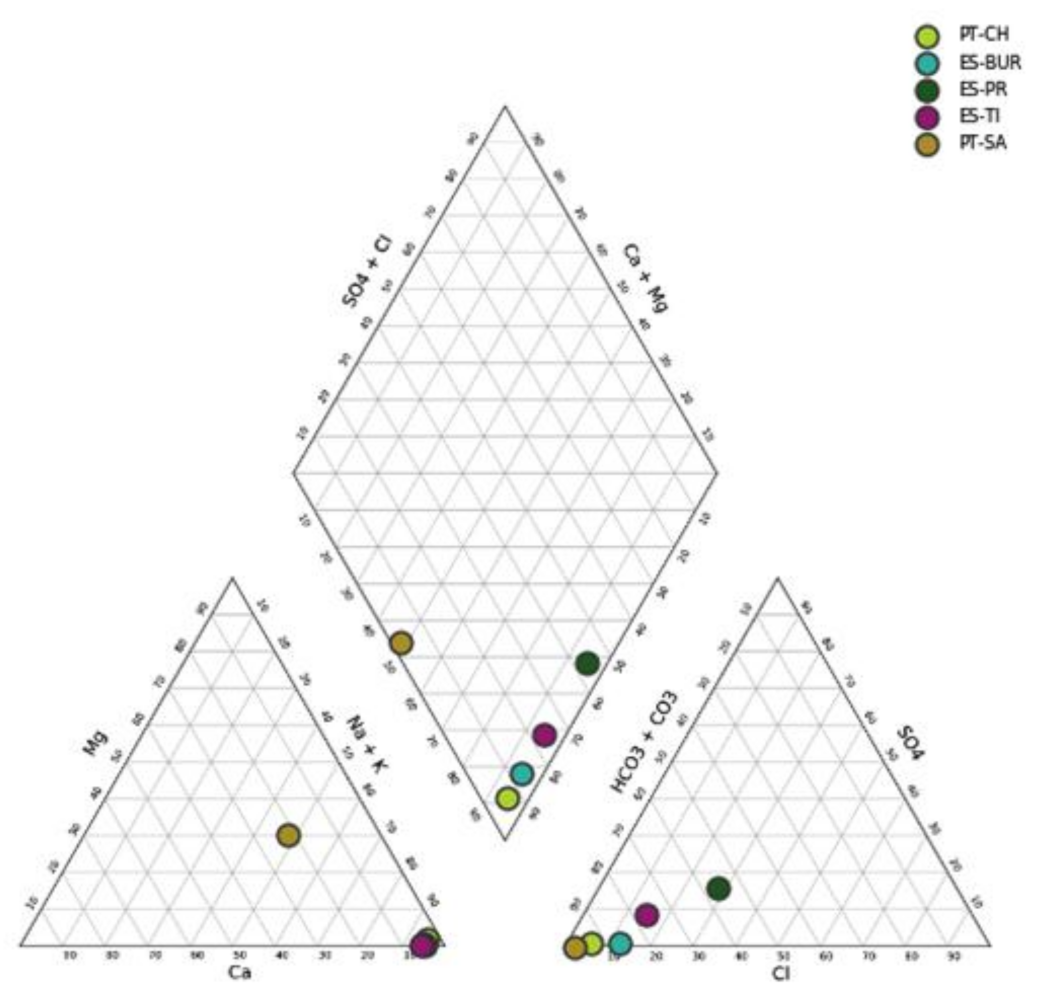
3.1. Physicochemical Classification of Natural Thermal Mineral Waters
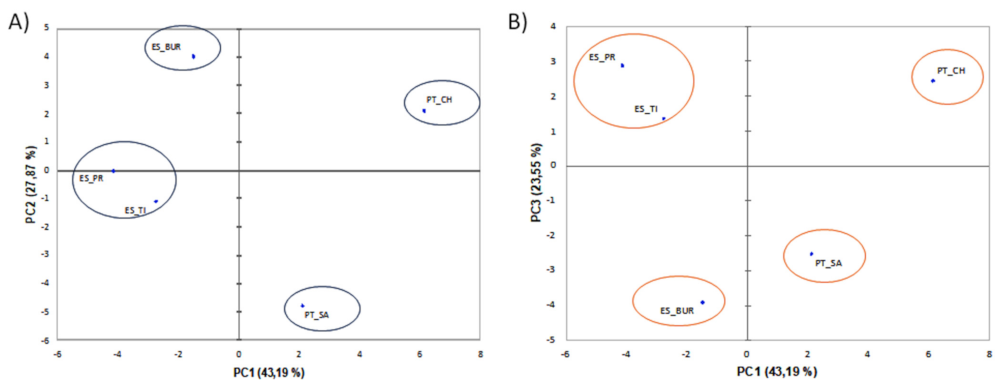
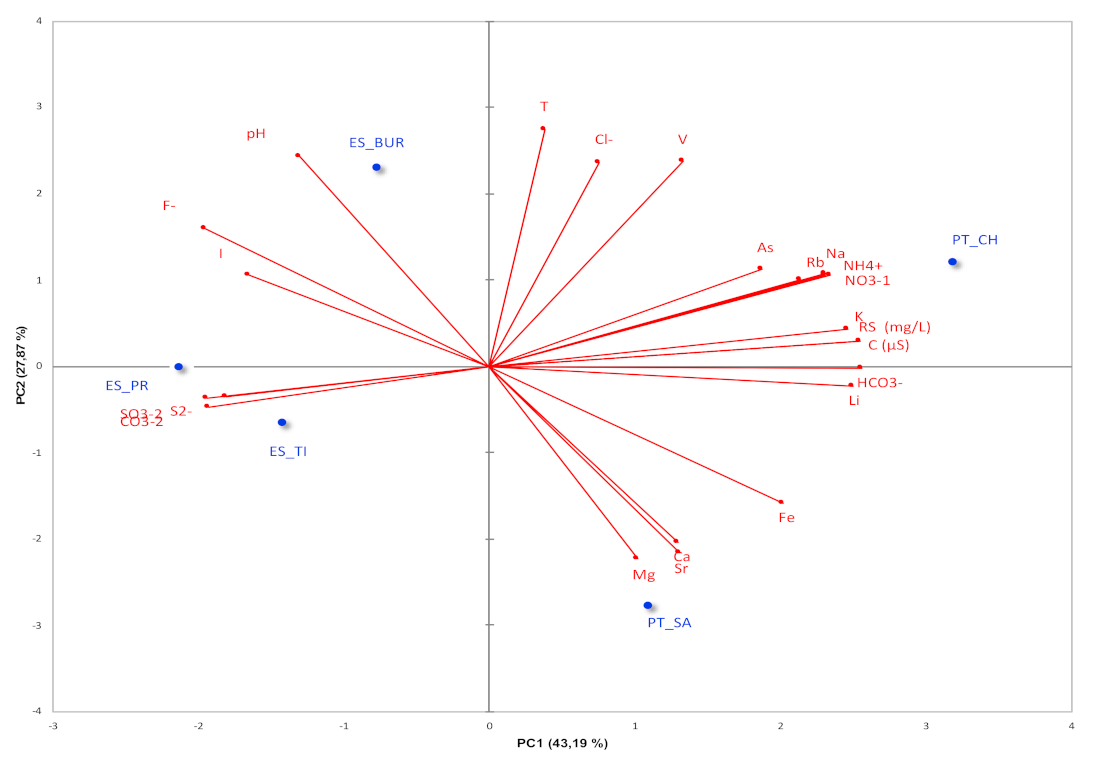
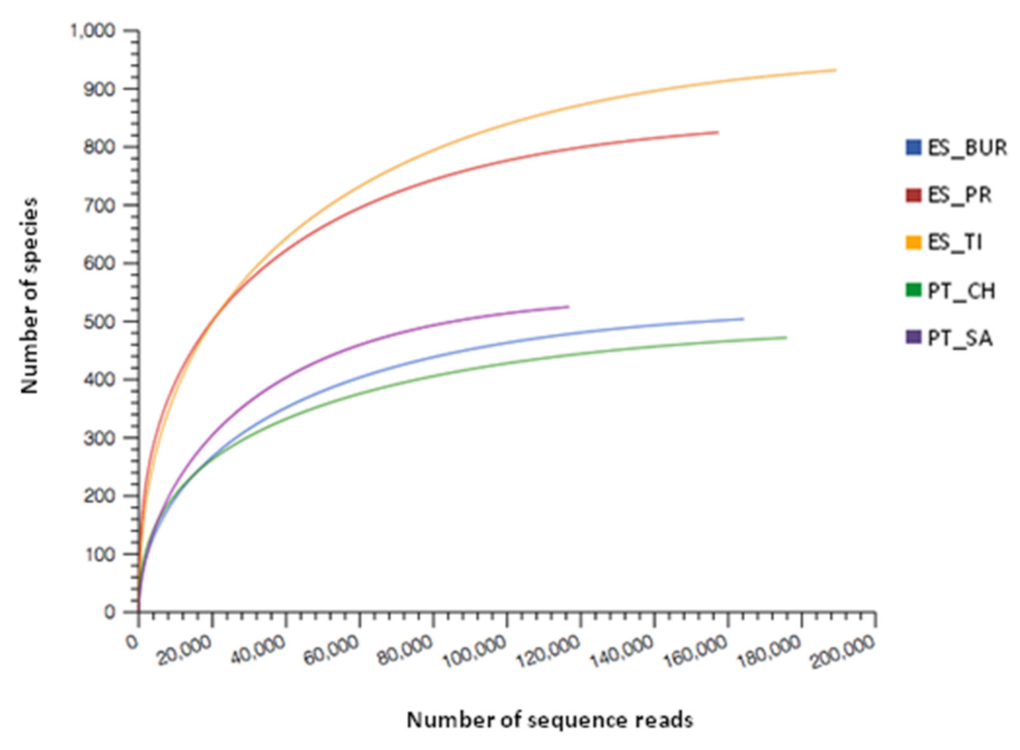
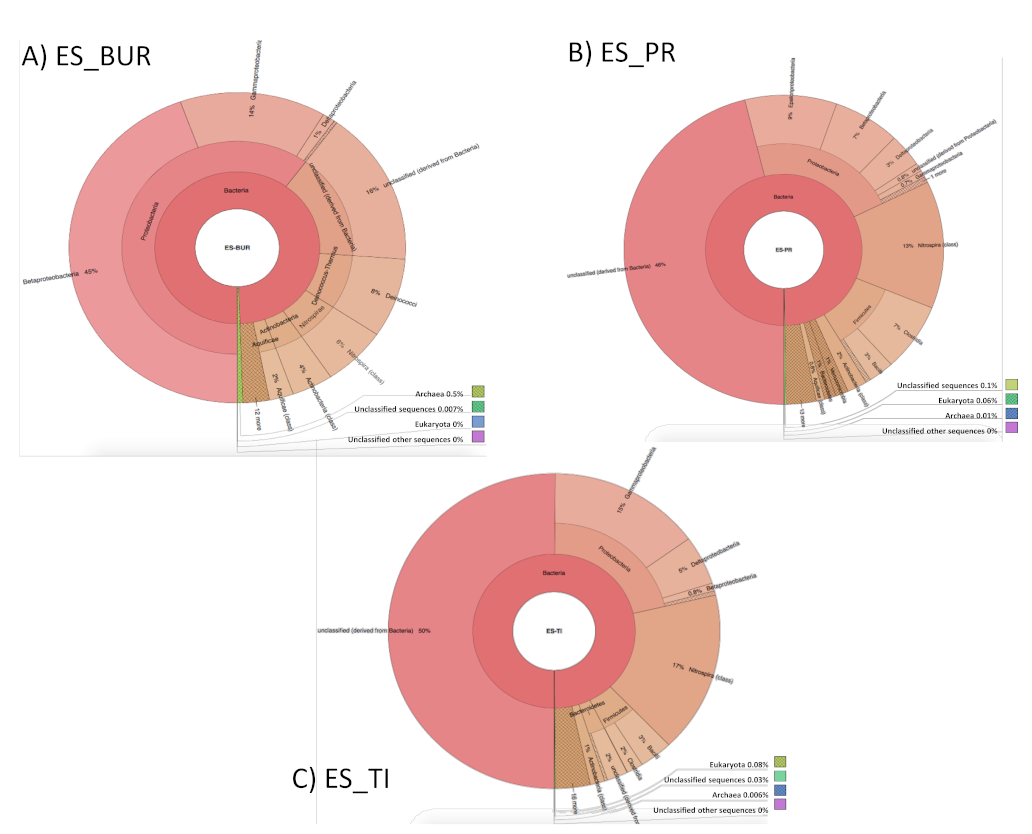
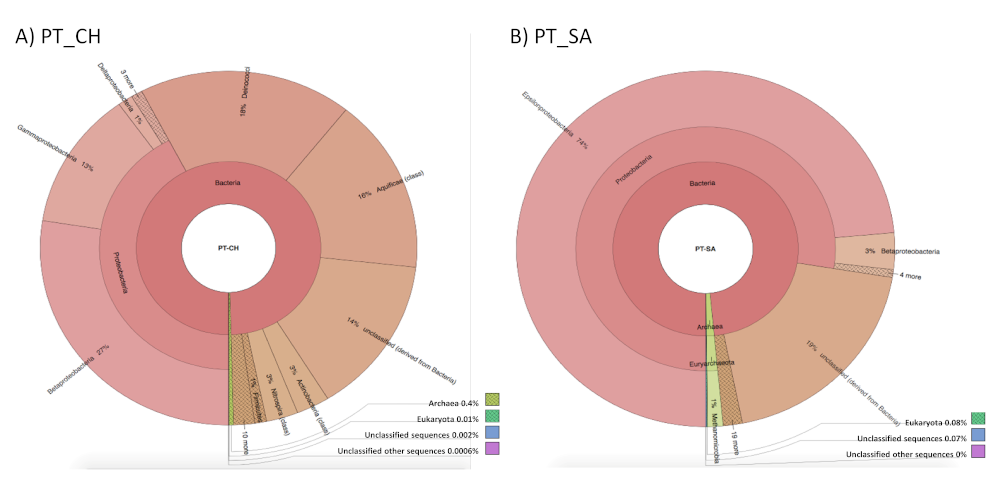
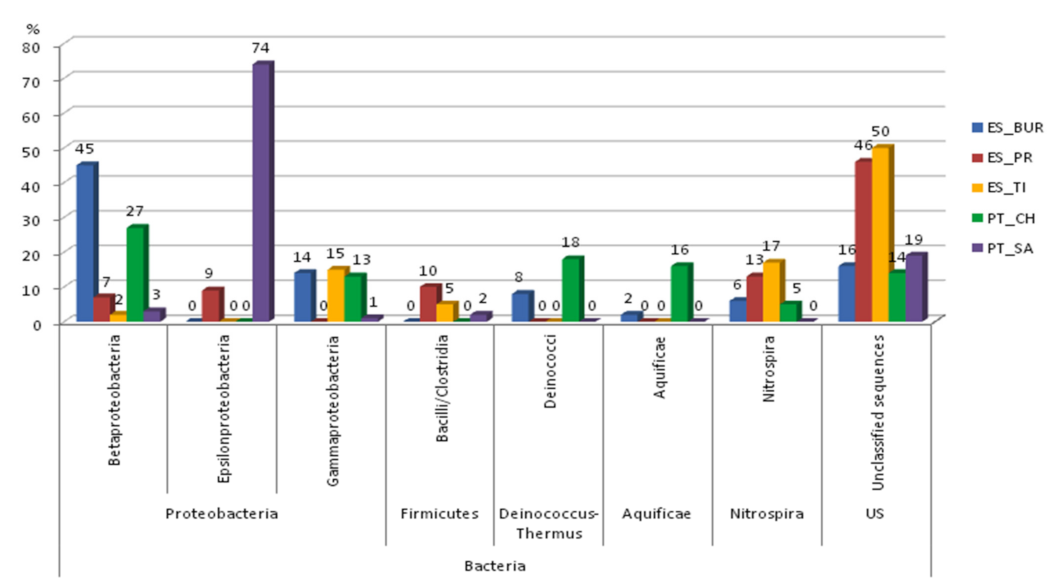
3.2. Characterization of Microbiota Communities of Natural Thermal Mineral Waters
Comparison of Microbial Taxonomic Profiles
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lozán, J.L.; Meyer, S.; Karbe, L. Water As the Basis of Life. In Global Change: Enough Water for All? Lozán, J.L., Grassl, H., Hupfer, L., Menzel, L., Schönwiese, C.-D., Eds.; Wissenschaftliche Auswertungen: Hamburg, Germany, 2007. [Google Scholar]
- Chaplin, M. Do we underestimate the importance of water in cell biology? Nat. Rev. Mol. Cell Biol. 2006, 7, 861–866. [Google Scholar] [CrossRef]
- Chaplin, M.F. Water: Its importance to life. Biochem. Mol. Biol. Educ. 2001, 29, 54–59. [Google Scholar] [CrossRef]
- Bacci, D.C.; Pataca, E.M. Educação para a água. Estud. Avançados 2008, 22, 211–226. Available online: http://www.scielo.br/pdf/ea/v22n63/v22n63a14.pdf (accessed on 15 March 2018). [CrossRef]
- Lund, J.W. Balneological Use of Thermal Waters. 1998. Available online: http://www.sld.cu/galerias/pdf/sitios/rehabilitacion-bal/lundwj2.pdf (accessed on 15 March 2018).
- Quintela, M.M. Saberes e práticas termais: Uma perspectiva comparada em Portugal (Termas de S. Pedro do Sul) e no Brasil (Caldas da Imperatriz). História Ciências Saúde-Manguinhos 2004, 11 (Suppl. 1), 239–260. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, A.S.; Almeida, S.M.S.; Reis, A.R.; Alencoão, A.M.P.; Sousa, L.M.O.; Lourenço, J.M.M. Águas Minerais de Trás-os-Montes e Alto Douro (NE de Portugal): Sistematização e Aproveitamentos . In Para Conhecer a Terra: Memórias e Notícias de Geociências no Espaço Lusófono; Lopes, F.C., Andrade, A.I., Henriques, M.H., Quinta-Ferreira, M., Barata, M.T., Pena dos Reis, R., Eds.; Imprensa daniversidade de Coimbra: Coimbra, Portugal, 2012; pp. 409–420. [Google Scholar]
- Ramos, A.R.C.C.V. O Termalismo em Portugal: Dos Factores de Obstrução à Revitalização pela Dimensão Turística. Ph.D. Thesis, Universidade de Aveiro, Aveiro, Portugal, 2005. [Google Scholar]
- Gianfaldoni, S.; Tchernev, G.; Wollina, U.; Roccia, M.G.; Fioranelli, M.; Gianfaldoni, R.; Lotti, T. History of the baths and thermal medicine. Open Access Maced. J. Med. Sci. 2017, 5, 566–568. [Google Scholar] [CrossRef] [Green Version]
- Silva, A.; Oliveira, A.S.; Vaz, C.V.; Correia, S.; Ferreira, R.; Breitenfeld, L.; Martinez-de-Oliveira, J.; Palmeira-de-Oliveira, R.; Pereira, C.M.F.; Palmeira-de-Oliveira, A.; et al. Anti-inflammatory potential of Portuguese thermal waters. Sci. Rep. 2020, 10, 22313. [Google Scholar] [CrossRef]
- Directive 2009/54/EC of the European Parliament and of the Council. Off. J. Eur. Union 2009, 164, 45–58.
- Pagourelias, E.D.; Zorou, P.G.; Tsaligopoulos, M.; Athyros, V.G.; Karagiannis, A.; Efthimiadis, G.K. Carbon dioxide balneotherapy and cardiovascular disease. Int. J. Biometeorol. 2011, 55, 657–663. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, H.E.; Bhatia, N.D.; Friedman, A.; Eng, R.M.; Seité, S. The role of cutaneous microbiota harmony in maintaining a functional skin barrier. J. Drugs Dermatol. 2017, 16, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Nunes, F.; Rodrigues, M.; Ribeiro, M.P.; Ugazio, E.; Cavalli, R.; Abollino, O.; Coutinho, P.; Araujo, A.R.T.S. Incorporation of Cró thermal water in a dermocosmetic formulation: Cytotoxicity effects, characterization and stability studies and efficacy evaluation. Int. J. Cosmet. Sci. 2019, 41, 604–612. [Google Scholar] [CrossRef]
- Oliveira, A.S.; Vaz, C.V.; Silva, A.; Correia, S.; Ferreira, R.; Breitenfeld, L.; Martinez-de-Oliveira, J.; Palmeira-de-Oliveira, R.; Pereira, C.; Cruz, M.T.; et al. In vitro evaluation of potential benefits of a silica-rich thermal water (Monfortinho Thermal Water) in hyperkeratotic skin conditions. Int. J. Biometeorol. 2020, 64, 1957–1968. [Google Scholar] [CrossRef]
- Viegas, J.; Esteves, A.F.; Cardoso, E.M.; Arosa, F.A.; Vitale, M.; Taborda-Barata, L. Biological Effects of Thermal Water-Associated Hydrogen Sulfide on Human Airways and Associated Immune Cells: Implications for Respiratory Diseases. Front. Public Health 2019, 7, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khaltaev, N.; Solimene, U.; Vitale, F.; Zanasi, A. Balneotherapy and hydrotherapy in chronic respiratory disease. J. Thorac. Dis. 2020, 12, 4459–4468. [Google Scholar] [CrossRef] [PubMed]
- Falagas, M.E.; Zarkadoulia, E.; Rafailidis, P.I. The therapeutic effect of balneotherapy: Evaluation of the evidence from randomised controlled trials. Int. J. Clin. Pract. 2009, 63, 1068–1084. [Google Scholar] [CrossRef]
- Karagülle, M.; Karagülle, M.Z. Effectiveness of balneotherapy and spa therapy for the treatment of chronic low back pain: A review on latest evidence. Clin. Rheumatol. 2015, 34, 207–214. [Google Scholar] [CrossRef]
- Varzaityte, L.; Kubilius, R.; Rapoliene, L.; Bartuseviciute, R.; Balcius, A.; Ramanauskas, K.; Nedzelskiene, I. The effect of balneotherapy and peloid therapy on changes in the functional state of patients with knee joint osteoarthritis: A randomized, controlled, single-blind pilot study. Int. J. Biometeorol. 2020, 64, 955–964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paduano, S.; Valeriani, F.; Romano-Spica, V.; Bargellini, A.; Borella, P.; Marchesi, I. Microbial biodiversity of thermal water and mud in an Italian spa by metagenomics: A pilot study. Water Sci. Technol. Water Supply 2018, 18, 1456–1465. [Google Scholar] [CrossRef]
- Valeriani, F.; Gianfranceschi, G.; Romano Spica, V. The microbiota as a candidate biomarker for SPA pools and SPA thermal spring stability after seismic events. Environ. Int. 2020, 137, 105595. [Google Scholar] [CrossRef] [PubMed]
- Rothschild, L.J.; Mancinelli, R.L. Life in extreme environments. Nature. 2001, 409, 1092–1101. [Google Scholar] [CrossRef]
- Rampelotto, P.H. Extremophiles and Extreme Environments. Life 2013, 3, 482–485. [Google Scholar] [CrossRef]
- Raddadi, N.; Cherif, A.; Daffonchio, D.; Neifar, M.; Fava, F. Biotechnological applications of extremophiles, extremozymes and extremolytes. Appl. Microbiol. Biotechnol. 2015, 99, 7907–7913. [Google Scholar] [CrossRef] [Green Version]
- Mirete, S.; Morgante, V.; González-Pastor, J.E. Functional metagenomics of extreme environments. Curr. Opin. Biotechnol. 2016, 38, 143–149. [Google Scholar] [CrossRef]
- Nicoletti, G.; Corbella, M.; Jaber, O.; Marone, P.; Scevola, D.; Faga, A. Non-pathogenic microflora of a spring water with regenerative properties. Biomed. Rep. 2015, 3, 758–762. [Google Scholar] [CrossRef]
- Seité, S.; Flores, G.E.; Henley, J.B.; Sarrazin, P.; Ainouche, R.; Fierer, N.; Martin, R. Skin microbiome in patients with psoriasis before and after balneotherapy at the thermal care center of La Roche-Posay. J. Am. Acad. Dermatol. 2016, 74, AB276. [Google Scholar]
- Aburto-Medina, A.; Shahsavari, E.; Cohen, M.; Mantri, N.; Ball, A.S. Analysis of the Microbiome (Bathing Biome) in Geothermal Waters from an Australian Balneotherapy Centre. Water 2020, 12, 1705. [Google Scholar] [CrossRef]
- Handelsman, J. Metagenomics: Application of genomics to uncultured microorganisms. Microbiol. Mol. Biol. Rev. 2004, 68, 669–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, A.S. Águas minerais naturais de Trás-os-Montes e Alto Douro: Património legado pela natureza. Bol. Cult. Esc. Secundária Camilo Castelo Branco 2011, 17, 18–29. [Google Scholar]
- Oliveira, A.S. Águas subterrâneas do setor norte de Portugal continental: Enquadramentos geográfico e geológico, tipologias e aproveitamentos. Bol. Soc. Española Hidrol. Med. 2015, 30, 57–71. [Google Scholar] [CrossRef] [Green Version]
- Meijide-Faílde, R.; Juncosa, R.; Delgado, J. Caracterización Hidrogeoquímica y Terapéutica de las Aguas Mineromedicinales y Minerales Naturales de Galicia. In Proceedings of the Libro de Actas del I Congreso Internacional del Agua “Termalismo y Calidad de Vida”, Ourense, Spain, 23–24 September 2015; pp. 61–72. [Google Scholar]
- Meijide-Faílde, R.; Leira, M.; Torres, E.; Rodríguez, M.C.L.; Carballeira, R. Estudio del componente biológico de las aguas mineromedicinales y termales de Ourense: Burgas y Outariz. In Proceedings of the Libro de Actas del I Congreso Internacional del Agua “Termalismo y Calidad de Vida”, Ourense, Spain, 23–24 September 2015; pp. 519–524. [Google Scholar]
- Michard, G.; Beaucaire, C. Les eaux thermales des granites de Galice (Espagne): Deseaux carbogazeuses aux eaux alcalines (Thermal waters from granites of Galicia (Spain): From CO2-rich to high-pH waters. Chem. Geol. 1993, 110, 345–360. [Google Scholar] [CrossRef]
- Ramírez Ortega, A. Revisión geoquímica de las aguas minero-medicinales de Galicia. An. Real Acad. Farm. 2001, 76, 1–45. [Google Scholar]
- Maraver, F. Vademecun de Aguas Mineromedicinales Españolas; Instituto de Salud Carlos III—Universidad Complutense de Madrid: Madrid, Spain, 2002. [Google Scholar]
- Pedro, A.; Cantista, P. O termalismo em Portugal. An. Hidrol. Med. 2010, 3, 79–107. [Google Scholar]
- Oliveira, A.S. Hidrogeologia dos Sistemas Gasocarbónicos da Província Hidromineral Transmontana: Ribeirinha (Mirandela), Sandim (Vinhais), Segirei e Salgadela (Chaves). Ph.D. Thesis, Universidade de Trás-os-Montes e Alto Douro, Vila Real, Spain, 2001. [Google Scholar]
- Machado, J.M.G.S. Ocorrências Hidrominerais do NE de Portugal Continental: Inventariação, Sistematização e Aproveitamentos didácticos. Master’s Thesis, Universidade de Trás-os-Montes e Alto Douro, Vila Real, Spain, 2009. [Google Scholar]
- Pacheco, F.A.L. Hidrogeologia em Maciços de Rochas Cristalinas (Morais-Chacim-Macedo de Cavaleiros). Bases Para a Gestão Integrada dos Recursos Hídricos da Região. Ph.D. Thesis, Universidade de Trás-os-Montes e Alto Douro, Vila Real, Spain, 2000. [Google Scholar]
- Piper, A.M. A graphic procedure in the geochemical interpretation of water analysis. Am. Geophys. Union Trans. 1944, 25, 914–923. [Google Scholar] [CrossRef]
- Back, W.; Hanshaw, B.B. Chemical geohydrology. Adv. Hydrosci. 1965, 2, 49–109. [Google Scholar]
- Malan, G. XLSTAT 2017: Data Analysis and Statistical Solution for Microsoft Excel; Addinsoft: Paris, France, 2017. [Google Scholar]
- Vieira, A.F.C.S. Aplicação de Métodos Quimiométricos aos Resultados de Técnicas Hifenadas Para Caracterização da Aguardente de Cana-de-Açúcar de Cabo Verde. Master’s Thesis, Universidade de Aveiro, Aveiro, Portugal, 2007. [Google Scholar]
- Thijs, S.; De Beeck, M.O.; Beckers, B.; Truyens, S.; Stevens, V.; Van Hamme, J.D.; Weyens, N.; Vangronsveld, J. Comparative evaluation of four bacteria-specific primer pairs for 16S rRNA gene surveys. Front. Microbiol. 2017, 8, 494. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Smith, D.P.; Peay, K.G. Sequence depth, not PCR replication, improves ecological inference from Next Generation DNA Sequencing. PLoS ONE 2014, 9, e90234. [Google Scholar] [CrossRef] [Green Version]
- Ison, S.A.; Delannoy, S.; Bugarel, M.; Nagaraja, T.G.; Renter, D.G.; den Bakker, H.C.; Nightingale, K.K.; Fach, P.; Loneragan, G.H. Targeted amplicon sequencing for SNP genotyping of attaching and effacing Escherichia coli O26: H11 cattle strains using a high-throughput library preparation technique. Appl. Environ. Microbiol. 2015, 82, 640–649. [Google Scholar] [CrossRef] [Green Version]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. 2010. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc (accessed on 12 September 2018).
- Wilke, A.; Gerlach, W.; Harrison, T.; Paczian, T.; Trimble, W.L.; Meyer, F. MG-RAST Manual for Version 4, Revision 3. 2017. Available online: http://ftp.metagenomics.anl.gov/data/manual/mg-rast-manual.pdf (accessed on 12 September 2018).
- Leggett, R.M.; Ramirez-Gonzalez, R.H.; Clavijo, B.J.; Waite, D.; Davey, R.P. Sequencing quality assessment tools to enable data-driven informatics for high throughput genomics. Front. Genet. 2013, 4, 288. [Google Scholar] [CrossRef]
- Marques, J.M.V.V.B. Geoquimica dos Fluidos e da Interacção Água-Rocha: Os Casos das Águas Mineralizadas Quentes e Frias de Chaves, Vilarelho da Raia, Vidago e Pedras Salgadas. Ph.D. Thesis, Universidade Técnica de Lisboa, Instituto Superior Técnico, Lisboa, Portugal, 1999. [Google Scholar]
- Branco, L.; Águas Minerais Naturais e Águas de Nascente. Associação Portuguesa dos Industriais de Águas Minerais Naturais e de Nascente. 2015. Available online: https://www.apiam.pt/images/newsconteudo/ficheiro1/158_LIVROBRANCO2015.pdf (accessed on 12 September 2018).
- Matz, H.; Orion, E.; Wolf, R. Balneotherapy in dermatology. Dermat. Ther. 2003, 16, 132–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Browne, P.D.; Nielsen, T.K.; Kot, W.; Aggerholm, A.; Gilbert, M.T.P.; Puetz, L.; Rasmussen, M.; Zervas, A.; Hansen, L.H. GC bias affects genomic and metagenomic reconstructions, underrepresenting GC-poor organisms. Gigascience 2020, 9, giaa008. [Google Scholar] [CrossRef] [PubMed]
- Wen, C.; Wu, L.; Qin, Y.; Van Nostrand, J.D.; Ning, D.; Sun, B.; Zhou, J. Evaluation of the reproducibility of amplicon sequencing with Illumina MiSeq platform. PLoS ONE 2017, 12, e0176716. [Google Scholar] [CrossRef]
- Fuciños González, P. Um Estúdio Integrado Das Lipasas de Thermus Thermophilus HB27: Condicións de Producción, Localización Celular, Purificación e Caracterización. Ph.D. Tesis, Universidad de Vigo, Vigo, Spain, 2007. [Google Scholar]
- Everroad, R.C.; Otaki, H.; Matsuura, K.; Haruta, S. Diversification of bacterial community composition along a temperature gradient at a thermal spring. Microbes Environ. 2012, 27, 374–381. [Google Scholar] [CrossRef] [Green Version]
- Lewin, A.; Wentzel, A.; Valla, S. Metagenomics of microbial life in extreme temperature environments. Curr. Opin. Biotechnol. 2013, 24, 516–525. [Google Scholar] [CrossRef]
- Stetter, K.O. Extremophiles and their adaptation to hot environments. FEBS Lett. 1999, 452, 22–25. [Google Scholar] [CrossRef]
- Wang, S.; Hou, W.; Dong, H.; Jiang, H.; Huang, L.; Wu, G.; Zhang, L. Control of temperature on microbial community structure in hot springs of the tibetan plateau. PLoS ONE 2013, 8, e62901. [Google Scholar] [CrossRef] [Green Version]
- Sharp, C.E.; Brady, A.L.; Sharp, G.H.; Grasby, S.E.; Stott, M.B.; Dunfield, P.F. Humboldt’s spa: Microbial diversity is controlled by temperature in geothermal environments. ISME J. 2014, 8, 1166–1174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ondov, B.D.; Bergman, N.H.; Phillippy, A.M. Interactive metagenomic visualization in a Web browser. BMC Bioinform. 2011, 12, 385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kersters, K.; De Vos, P.; Gillis, M.; Swings, J.; Vandamme, P.; Stackebrandt, E. Introduction to the Proteobacteria. In The Prokaryotes, 3rd ed.; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.-H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; pp. 3–37. [Google Scholar]
- Madigan, M.T.; Martinko, J.M. Brock Biology of Microorganisms, 11th ed.; Pearson Prentice Hall: Upper Saddle River, NJ, USA, 2006. [Google Scholar]
- HMPC. Human Microbiome Project Consortium. Structure, function and diversity of the healthy human microbiome. Nature 2012, 486, 207–214. [Google Scholar] [CrossRef] [Green Version]
- Zeeuwen, P.L.; Boekhorst, J.; van den Bogaard, E.H.; de Koning, H.D.; van de Kerkhof, P.M.; Saulnier, D.M.; van Swam, I.I.; van Hijum, S.A.; Kleerebezem, M.; Schalkwijk, J.; et al. Microbiome dynamics of huma nepidermis folowing skin barrier disruption. Genome Biol. 2012, 13, R101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cosseau, C.; Romano-Bertrand, S.; Duplan, H.; Lucas, O.; Ingrassia, I.; Ingrassia, I.; Roques, C.; Jumas-Bilak, E. Proteobacteria from the human skin microbiota: Species-level diversity and hypotheses. One Health 2016, 2, 33–41. [Google Scholar] [CrossRef] [Green Version]
- Mathieu, A.; Delmont, T.O.; Vogel, T.M.; Robe, P.; Nalin, R.; Simonet, P. Life on human sur-faces: Skin metagenomics. PLoS ONE 2013, 8, e65288. [Google Scholar] [CrossRef]
- Nakatsuji, T.; Gallo, R.L. Dermatological therapy by topical application of nonpathogenic bacteria. J. Investig. Dermatol. 2014, 134, 11–14. [Google Scholar] [CrossRef] [Green Version]
- Madigan, M.T.; Martinko, J.M.; Bender, K.S.; Buckley, D.H.; Stahl, D.A. Microbiologia de Brock, 14th ed.; Artmed: São Paulo, Brazil, 2016. [Google Scholar]
- Mangrola, A.; Dudhagara, P.; Koringa, P.; Joshi, C.G.; Parmar, M.; Patel, R. Deciphering the microbiota of Tuwa hot spring, India using shotgun metagenomic sequencing approach. Genom. Data 2015, 4, 153–155. [Google Scholar] [CrossRef] [Green Version]
- Panda, A.K.; Bisht, S.S.; De Mandal, S.; Kumar, N.S. Bacterial and archeal community composition in hot springs from Indo-Burma region, North-east India. AMB Express 2016, 6, 111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaudhuri, B.; Chowdhury, T.; Chattopadhyay, B. Comparative analysis of microbial diversity in two hot springs of Bakreshwar, West Bengal, India. Genom. Data 2017, 12, 122–129. [Google Scholar] [CrossRef]
- Garrity, G.M.; Holt, J.G. Phylum BIV. “Deinococcus–Thermus”. In Bergey’s Manual of Systematic Bacteriology, 2nd ed.; Boone, D., Castenholz, R., Eds.; Springer: New York, NY, USA, 2001; Volume 1, pp. 395–420. [Google Scholar]
- Zahradka, K.; Slade, D.; Bailone, A.; Sommer, S.; Averbeck, D.; Petranovic, M.; Lindner, A.B.; Radman, M. Reassembly of shattered chromosomes in Deinococcus radiodurans. Nature 2006, 443, 569–573. [Google Scholar] [CrossRef]
- Theodorakopoulos, N.; Bachar, D.; Christen, R.; Alain, K.; Chapon, V. Exploration of Deinococcus-Thermus molecular diversity by novel group-specific PCR primers. Microbiol. Open 2013, 2, 862–872. [Google Scholar]
- Sayeh, R.; Birrien, J.L.; Alain, K.; Barbier, G.; Hamdi, M.; Prieur, D. Microbial diversity in Tunisian geothermal springs as detected by molecular and culture-based approaches. Extremophiles 2010, 14, 501–514. [Google Scholar] [CrossRef]
- Huber, R.; Eder, W. Aquificales. In The Prokaryotes: An Evolving Electronic Resource for the Microbiological Community; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.-H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Griffiths, E.; Gupta, R.S. Molecular signatures in protein sequences that are characteristics of the phylum Aquificae. Int. J. Syst. Evol. Microbiol. 2006, 56, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Daims, H.; Nielsen, J.L.; Nielsen, P.H.; Schleifer, K.H.; Wagner, M. In situ characterization of Nitrospira-like nitrite-oxidizing bacteria active in wastewater treatment plants. Appl. Environ. Microbiol. 2001, 67, 5273–5284. [Google Scholar] [CrossRef] [Green Version]
- Freitag, T.E.; Chang, L.; Clegg, C.D.; Prosser, J.I. Influence of inorganic nitrogen management regime on the diversity of nitrite-oxidizing bacteria in agricultural grassland soils. Appl. Environ. Microbiol. 2005, 71, 8323–8334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watson, S.W.; Bock, E.; Valois, F.W.; Waterbury, J.B.; Schlosser, U. Nitrospira marina gen. nov. sp. nov.: A chemolithotrophic nitrite-oxidizing bacterium. Arch. Microbiol. 1986, 144, 1–7. [Google Scholar] [CrossRef]
- Lebedeva, E.V.; Alawi, M.; Fiencke, C.; Namsaraev, B.; Bock, E.; Spieck, E. Moderately thermophilic nitrifying bacteria from a hot spring of the Baikal rift zone. FEMS Microbiol. Ecol. 2005, 54, 297–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucker, S.; Wagner, M.; Maixner, F.; Pelletier, E.; Koch, H.; Vacherie, B.; Daims, H. A Nitrospira metagenome illuminates the physiology and evolution of globally important nitrite-oxidizing bacteria. Proc. Natl. Acad. Sci. USA 2010, 107, 13479–13484. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Physicochemical Parameters | Units | Water Samples | ||||
|---|---|---|---|---|---|---|
| ES_BUR | ES_PR | ES_TI | PT_CH | PT_SA | ||
| Condutivity (C) | µS | 956 | 461 | 447 | 2320 | 1455 |
| Dry residue (DR) at 110 °C | mg/L | 656 | 374 | 365 | 1626 | 998 |
| pH | 7.5 | 7.3 | 6.8 | 6.8 | 6.1 | |
| Emergency temperature | °C | 64 | 50.4 | 40 | 70 | 15 |
| Litium (Li) | mg/L | 0.98 | 0.74 | 0.54 | 2.33 | 1.84 |
| Sodium (Na) | mg/L | 222.8 | 82 | 87 | 703.7 | 205 |
| Potassium (K) | mg/L | 9.26 | 4.67 | 3.82 | 65.32 | 23.9 |
| Rubidium (Rb) | mg/L | 0.14 | 0.04 | 0.03 | 0.6 | 0.12 |
| Cesium (Cs) | mg/L | 0.18 | 0.11 | 0.06 | nd | 0.23 |
| Calcium (Ca) | mg/L | 8.90 | 3.1 | 3.78 | 20.46 | 72.8 |
| Magnesium (Mg) | mg/L | 0.64 | 0.12 | 0.16 | 4.54 | 37.1 |
| Strontium (Sr) | mg/L | <0.01 | 0.04 | 0.03 | 0.37 | 1.41 |
| Iron (Fe) | mg/L | 0.02 | 0.02 | <0.01 | 0.23 | 0.35 |
| Boron (B) | mg/L | 0.70 | 0.85 | 0.51 | 0.79 | 0.52 |
| Aluminum (Al) | mg/L | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
| Manganese (Mn) | mg/L | 0.04 | <0.01 | <0.01 | 0.02 | 0.24 |
| Lead (Pb) | μg/L | 0.30 | <0.2 | <0.2 | <0.2 | <0.2 |
| Chromium (Cr) | μg/L | <1 | <1 | <1 | <1 | <1 |
| Cadmium (Cd) | μg/L | <0.5 | <0.5 | <0.5 | <0.5 | <0.5 |
| Copper (Cu) | μg/L | 1.18 | <1 | <1 | <1 | <1 |
| Zinc (Zn) | μg/L | 10.17 | 1.6 | 0.9 | <1 | 2.2 |
| Total Arsenic | μg/L | 6 | 14 | 13 | 79.20 | <1 |
| Arsenic III | μg/L | nd | 13 | 10 | 20.30 | nd |
| Arsenic V | μg/L | nd | 1.79 | 3.25 | 58.90 | nd |
| Cobalt (Co) | μg/L | <1 | <1 | <1 | <1 | 1 |
| Molybdenum (Mo) | μg/L | 1.17 | 3.5 | 1.1 | 0.74 | <0.2 |
| Vanadium (V) | μg/L | 20.00 | <1 | <1 | 20.00 | <1 |
| Chloride (Cl−) | mg/L | 28.44 | 28.83 | 15.86 | 38.30 | 10.26 |
| Ammoniacal nitrogen (NH4+) | mg/L | 0.43 | 0.01 | 0.3 | 1.62 | 0.35 |
| Sulphite (SO32−) | mg/L | <1 | 4.3 | <1 | 1 | <1 |
| Sulphate (SO42−) | mg/L | 3.5 | 29.93 | 17.76 | 11.29 | 1.26 |
| Fluoride (F−) | mg/L | 16.63 | 21.97 | 14.76 | 8.01 | 0.54 |
| Sulphide (S2−) | mg/L | <0.05 | 9.05 | 3.38 | <0.05 | 0.46 |
| Carbonate (CO32−) | mg/L | 0 | 12 | 12 | 0 | 0 |
| Bicarbonate (HCO3−) | mg/L | 585.6 | 122 | 195.2 | 1769 | 1207.8 |
| Nitrite (NO2−) | mg/L | 0.64 | <0.05 | <0.05 | <0.05 | <0.05 |
| Nitrate (NO3−) | mg/L | <0.05 | <0.05 | <0.05 | 2.62 | <0.05 |
| Phosphate (PO42−) | mg/L | 1.53 | <0.05 | <0.05 | <0.05 | <0.05 |
| Iodide (I) | μg/L | 3.58 | 1.9 | 1.3 | nd | 1.3 |
| PC1 | PC2 | PC3 | PC4 | |
|---|---|---|---|---|
| Own value | 13.82 | 8.92 | 7.54 | 1.73 |
| Variability (%) | 43.19 | 27.87 | 23.55 | 5.393 |
| Cumulative variance (%) | 43.19 | 71.05 | 94.61 | 100.00 |
| Samples | Number of Reads before Quality Control | Number of Reads after Quality Control (%) | Average Reads Length (bp) | % of GC Content after Quality Control |
|---|---|---|---|---|
| ES_BUR | 164,464 | 164,377 (99.95) | 421 | 58 ± 5 |
| ES_PR | 157,364 | 157,320 (99.97) | 435 | 54 ± 5 |
| ES_TI | 189,567 | 189,439 (99.93) | 428 | 53 ± 5 |
| PT_CH | 175,886 | 175,741 (99.92) | 415 | 59 ± 4 |
| PT_SA | 116,877 | 116,838 (99.97) | 430 | 52 ± 5 |
| Total | 804,158 | 803,715 (99.95) | 426 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pascoal-Ferreira, P.; Glez-Peña, D.; Miranda, C.; Poeta, P.; Coutinho, J.; Fdez-Riverola, F.; Torrado-Agrasar, A.; Rúa, M.L.; Igrejas, G. Are There Benefits from Thermal Bacteria for Health? The Hydrogenome Role. Water 2021, 13, 1439. https://doi.org/10.3390/w13111439
Pascoal-Ferreira P, Glez-Peña D, Miranda C, Poeta P, Coutinho J, Fdez-Riverola F, Torrado-Agrasar A, Rúa ML, Igrejas G. Are There Benefits from Thermal Bacteria for Health? The Hydrogenome Role. Water. 2021; 13(11):1439. https://doi.org/10.3390/w13111439
Chicago/Turabian StylePascoal-Ferreira, Patrick, Daniel Glez-Peña, Carla Miranda, Patrícia Poeta, João Coutinho, Florentino Fdez-Riverola, Ana Torrado-Agrasar, María Luisa Rúa, and Gilberto Igrejas. 2021. "Are There Benefits from Thermal Bacteria for Health? The Hydrogenome Role" Water 13, no. 11: 1439. https://doi.org/10.3390/w13111439