
Summary
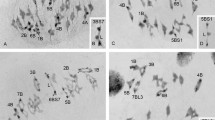
The seven chromosome pairs of the normal chromosome complement ofSecale céréale (A chromosomes) have been identified at pachytene. A detailed chromomere analysis of the A chromosomes and the “standard fragment” (a type of B chromosome previously described) including the number, size, and sequence of the chromomeres, permitted the construction of eight reference maps of these chromosomes.
-
(1)
There is a good agreement between the chromomere number and chromosome length of the different A chromosomes and the standard fragment.
-
(2)
Knob formations are seriations of large chromomeres and deeply stained fibrillae. The 11 knob formations of the A chromosomes plus the one of the standard fragment are all accompanied on one or both sides by seriations of particularly small chromomeres. The organization of the kinetochoric structure is of the same type in all the seven A chromosomes and the standard fragment.
-
(3)
Small conspicuous seriations composed generally of two consecutive pairs of larger chromomeres having between them a fibrilla pair particularly weakly stained are found at regular intervals in the chromosome arms. This results in a regional differentiation of the chromosome.
-
(4)
The length of the thick regions adjacent to the kinetochores is apparently not correlated with arm length. When the length of the arm varies it is mostly the median regions — with medium size chromomeres — that vary in length.
-
(5)
Chromomere pairs having chromomeres of unequal size or having one of the chromomere partners absent are found in all chromosomes. This evidence of the structural heterozygosity of the chromosome material of rye is in agreement with the cross-fertilizing breeding system of the species.
-
(6)
A structural rearrangement involving from one to seven chromomere pairs is present in chromosome VI. The number of chromomeres involved in this rearrangement varies from cell to cell, the rearrangement being absent in some P.M.C.'s.
-
(7)
Not all chromomeres have the usual round shape but some possess a hairy structure. Hairy chromomeres appear with a much greater frequency in the positively heteropycnotic regions of the chromosome where chromomeres are large than in the other regions of the chromosome where the chromomeres are of medium size or small.
-
(8)
It could be ascertained that the standard fragment of rye — like that in maize — is evolved to such an extent that there is no segment of the A chromosomes that can be directly homologized with it. B chromosomes may have arisen from A chromosomes through a hereditary variation in the properties of the kinetochore of an A chromosome, or their process of origin may have been accompanied by such a hereditary variation.
-
(9)
The standard fragment is not an exception to the general rule of the heterochromatic constitution of the B's since it also shows a certain degree of heteropycnosis when compared with the A chromosomes at pachytene. Both the number and extension of positively and negatively heteropycnotic regions is bigger in the standard fragment than in the A chromosomes.
-
(10)
The average distance in micra between the centres of two consecutive chromomeres decreases with increasing size of the chromomeres. At the same time the distance between the centres of large chromomeres is approximately of the same length as their diameter.
-
(11)
All A chromosomes possess the same general chromomeric pattern with minor variants. In all A chromosomes the chromomeres are particularly large on both sides of the kinetochore. There is a subsequent general decrease in chromomere size on both sides of the kinetochore towards the chromosome ends, where an abrupt increase in chromomere size takes place in most cases (knob formations). This gradient of chromomere size originates on both sides of the kinetochore where chromomeres of large size are found and finishes in chromomeres of very small size at the chromosome ends or just behind the knob formations when these are present. The regulation of the rate at which this decrease in chromomere size takes place is apparently controlled in the first place by the knob formations and, in their absence, by the arm length. The existence of this gradient and the variation of its shape in relation to knob formation position and arm length indicate that the size of a chromomere is not only determined by its genetical constitution and nuclear environment but is also dependent on its position within the chromosome.
-
(12)
Chromosomes III and VI are rather similar to chromosomes IV and VII respectively. The existence of a chromomere size gradient in the arms of the A chromosomes of rye furnishes the basis for an interpretation that conciliates at the same time the apparent genetical differentiation of these chromosomes with their morphological similarity.
-
(13)
In rye pachytene chromosomes there is no essential difference, at the mircoscopic level, between the structural organization of euchromatic and heterochromatic regions. Heteropycnosis appears as a phenomenon determined not only by the genetic constitution and nuclear environment of a certain chromosome region but also by the position of this same chromosome region within the chromosome body.
-
(14)
The structural comparative analysis of on one hand, the part of the chromosome arms not including the knob formations, and on the other hand kinetochores and knob formations, reveals that in rye pachytene chromosomes there is no essential morphological difference, at the microscopic level, between the components of the kinetochores, those of knob formations, and those of the remaining part of the chromosome arms, all these three chromosome regions being composed of chromomeres and fibrillae which are morphologically identical. This analysis also furnishes a series of data that permit an understanding of the apparently deviating morphological appearance of kinetochores and knob formations. The pachytene chromosome appears, at the microscopic level, as a structural unity.
Similar content being viewed by others


Literature cited
Belling, J.: The ultimate chromomeres ofLilium andAloë, with regard to the numbers of genes. Univ. California Publ. Bot.14, 307–318 (1928a).
—: Contraction of chromosomes during maturation divisions inLilium and other plants. Univ. California Publ. Bot.14, 335–343 (1928b).
Bosemark, N. O.: Accessory chromosomes inFestuca pratensis Huds. Hereditas36, 366–368 (1950).
Bridges, C. B.: Salivary chromosome maps. With a key to the banding of the chromosomes ofDrosophila melanogaster. J. Hered.26, 60–64 (1935).
Camara, A.: Efeitos do calor sobre a microsporogénese doSecale cereale. Sci. Genet.1, 86–102 (1939).
Darlington, C. D.: The origin and behaviour of chiasmata, VIII.Secale cereale (n, 8). Cytologia4, 444–452 (1933a).
—: Meiosis inAgapanthus andKniphofia. Cytologia4, 229–240 (1933b).
- Selection and transmission of B-chromosomes. John Innes Hort. Inst. Fortieth Ann. Rep. 1949, S. 17–18. 1950.
- Mendel and the determinants, S. 315–332. In Genetics in the 20th century, edited byL. C. Dunn. New York 1951.
Darlington, C. D., andP. T. Thomas: Morbid mitosis and the activity of inert chromosomes inSorghum. Proc. Roy. Soc. Lond., Ser. B No 859130, 127–150 (1941).
Fernandes, A.: Sur le comportement d'un chromosome surnuméraire pendant la mitose. Sci. Genet.1, 141–167 (1939).
—: Sur l'origine des chromosomes surnuméraires hétérochromatiques chezNarcissus Bulbocodium L. Bol. Soc. Brot.17 (2a), 251–256 (1943).
—: Sur le comportement des chromosomes surnuméraires hétérochromatiques pendant la meiose. I. Chromosomes longs hétérobrachiaux. Bol. Soc. Brot.20 (2a), 93–154 (1946).
Geitler, L.: Das Verhalten der Chromozentren vonAgapanthus während der Meiose. Österr. bot. Z.82, 277–282 (1933).
Håkansson, A.: Überzählige Chromosomen in einer Rasse vonGodetia nutans Hiorth. Bot. Not.1945, 1–19.
Heribert-Nilsson, N.: Rågförädlingine metodik och principer. Nord. Jordbrugsf.1919, 4–9.
—: Eine Prüfung der Wege und Theorien der Inzucht. Hereditas23, 236–256 (1937).
Huskins, C. L., andS. G. Smith: Meiotic chromosome structure inTrillium erectum L. Ann. of Bot.49, 119–150 (1935).
Kattermann, G.: Die Chromosomenverhältnisse bei Weizen-Roggenbastarden der zweiten Generation mit besonderer Berücksichtigung der Homologiebeziehungen. Z. Abstammgslehre70, 265–308 (1936).
—: Das Verhalten des Chromosoms für Behaarung roggenbehaarter Nachkommen aus Weizen-Roggenbastardierung in neuen Kreuzungen mit Roggen und Weizen. Z. Abstammgslehre74, 1–16 (1937). Ein neuer Karyotyp bei Roggen. Chromosoma1, 284–299 (1939).
Kaufmann, B. P.: Chromosome structure in relation to the chromosome cycle. II. Bot. Review14, 57–126 (1948).
Kostoff, D.: Chromosome behavior inTriticum hybrids and allied genera. II.Tr. Timopheevi (n=14) ×Secale cereale (n=7). Z. Zücht., Reihe A21, 378–379 (1937a).
—: Studies on polyploid plants. XIV. The behaviour ofHaynaldia genom in the trigeneric triple hybrid (Triticum, dicoccum × Haynaldiavillosa) ×Secale cereale. Bull. Inst. Genetics USSR11, 10–11 (1937b).
—: Interspecific hybrids inSecale (rye). I. C. R. [Doklady] Acad. Sci. URSS14, 213–214 (1937c).
Lamm, R.: Cytological studies on inbred rye. Hereditas22, 217–240 (1936).
Levan, A.: Studies on the meiotic mechanism of haploid rye. Hereditas28, 177–211 (1942).
Lewitsky, G. A.: The morphology of chromosomes. Bull. Appl. Bot., Genetics a. Plant Breeding27, 19–174 (1931).
Lima-de-Faria, A.: B chromosomes of rye at pachytene. Portug. Acta Biol. A2, 167–174 (1948).
—: The structure of the centromere of the chromosomes of rye. Hereditas35, 77–85 (1949a).
—: Genetics, origin and evolution of kinetochores. Hereditas35, 422–444 (1949b).
—: The Feulgen test applied to centromeric chromomeres. Hereditas36, 60–74 (1950a).
—: Meiosis and pollen mitosis in rye under controlled conditions. Hereditas36, 106–109 (1950b).
- The chromosomes ofSecale cereale at pachytene. Proc. Seventh Int. Bot. Congr.1950 c.
Ljung, E. W.: The rye breeding work of the Seed Association. “Svalöf 1886–1946” (Lund)1948, 127–134.
Longley, A. E.: Knob positions on corn chromosomes. J. Agricult. Res.59, 475–490 (1939).
Lundqvist, A.: On self-sterility and inbreeding effect in tetraploid rye. Hereditas33, 570–571 (1947).
Mather, K.: Statistical Analysis in Biology. London, 1946. 267 p.
Mather, K., andR. Lamm: The negative correlation of chiasma frequencies. Hereditas20, 65–70 (1935).
McClintock, B.: A method for making aceto-carmine smears permanent. Stain Technol.4, 53–56 (1929).
—: The association of non-homologous parts of chromosomes in the mid-prophase of meiosis inZea mays. Z. Zellforschg.19, 191–237 (1933).
—: The relation of a particular chromosomal element to the development of the nucleoli inZea mays. Z. Zellforschg.21, 294–328 (1934).
Melander, Y.: Accessory chromosomes in animals, especially inPolycelis tenuis. Hereditas36, 19–38 (1950).
Metz, C. W.: Observations on the nature and significance of minute chromosomal differences in species hybrids and in ordinary wild strains ofSciara. Genetics24, 105 (1939).
- Species hybrids, evolutionary chromosome changes, and the mechanism of chromosome rearrangement inSciara. Proc. VII. Internat. Genet. Congr. Edinburgh 1941, S. 215–218.
Morgan jr.,D. T.: The formation of chromocentres in interkinetic nuclei of maize. J. Hered.34, 194–198 (1943).
Müntzing, A.: Apomictic and sexual seed formation inPoa. Hereditas17, 131–154 (1933).
—: Chromosomenaberrationen bei Pflanzen und ihre genetische Wirkung. Z. Abstammgslehre76, 323–350 (1939).
—: Genetical effects of duplicated fragment chromosomes in rye. Hereditas29, 91–112 (1943a).
—: Double crosses of inbred rye. Bot. Not.1943, 333–345 (1943b).
—: Cytological studies of extra fragment chromosomes in rye. I. Iso-fragments produced by misdivision. Hereditas30, 231–248 (1944).
—: Experiences from work with induced polyploidy in cereals. “Svalöf 1886–1946” (Lund)1948, 324–337 (1948a).
—: Accessory chromosomes inPoa alpina. Heredity2, 49–61 (1948 b).
- Accessory chromosomes inSecale andPoa. Proc. Eighth Internat. Congr. of Gen. Hereditas Suppl.1949, 402–411.
—: Accessory chromosomes in rye populations from Turkey and Afghanistan. Hereditas36, 507–509 (1950).
—: Cyto-genetic properties and practical value of tetraploid rye. Hereditas37, 17–84 (1951).
Müntzing, A., andA. Lima-de-Faria: Pachytene analysis of standard fragments and large iso-fragments in rye. Hereditas35, 253–268 (1949).
Müntzing, A., andR. Prakken: Chromosomal aberrations in rye populations. Hereditas27, 273–308 (1941).
Nebel, B. R.: Chromosome structure. Bot. Review5, 563–626 (1939).
Östergren, G.: Heterochromatic B-chromosomes inAnthoxanthum. Hereditas33, 261–296 (1947).
—: Isopycnosis and isopycnotic, two new terms for use in chromosome studies. Hereditas36, 511–513 (1950).
Östergren, G., andR. Prakken: Behaviour on the spindle of the actively mobile chromosome ends of rye. Hereditas32, 473–494 (1946).
Pathak, G. N.: Studies in the cytology of cereals. J. Genet.39, 437–467 (1940).
Pesola, V. A.: Rye and wheat breeding in Finland. Heredity2, 141–143 (1948).
Prakken, R.: Studies of asynapsis in rye. Hereditas29, 475–495 (1943).
Prakken, R., andA. Müntzing: A meiotic peculiarity in rye, simulating a terminal centromere. Hereditas28, 441–482 (1942).
Randolph, L. F.: Genetic characteristics of the B chromosomes in maize. Genetics26, 608–631 (1941).
—: Inherited differences in pachytene configurations in maize. Genetics33, 121 (1948).
Rhoades, M. M.: Meiosis in maize. J. Hered.41, 58–67 (1950).
Ris, H.: The structure of meiotic chromosomes in the grasshopper and its bearing on the nature of “chromomeres” and “lamp-brush chromosomes”. Biol. Bull.89, 242–257 (1945).
Schultz, J.: The function of heterochromatin. Proc. VII. Internat. Genet. Congr. Edinburgh1941, S. 257–262.
Sears, E. R.: The cytology and genetics of the Wheats and their relatives. Adv. Genet.2, 239–270 (1948).
Tjio, J. H., andA. Levan: The use of oxiquinoline in chromosome analysis. Anales Est. Exper. Aula Dei2, 21–64 (1950).
White, M. J. D.: Animal cytology and evolution. Cambridge, 1945. 375 S.
Author information
Authors and Affiliations
Rights and permissions
About this article
Cite this article
Lima-de-Faria, A. Chromomere analysis of the chromosome complement of rye. Chromosoma 5, 1–68 (1953). https://doi.org/10.1007/BF01271482
Received:
Issue Date:
DOI: https://doi.org/10.1007/BF01271482