
Abstract
Vitreous humor (VH) is a gelatinous substance contained in the posterior chamber of the eye, playing a mechanical role in the eyeball. It has been the subject of numerous studies in various forensic applications, primarily for the assessment of postmortem interval and for postmortem chemical analysis. Since most of the xenobiotics present in the bloodstream are detected in VH after crossing the selective blood-retinal barrier, VH is an alternative matrix useful for forensic toxicology. VH analysis offers particular advantages over other biological matrices: it is less prone to postmortem redistribution, is easy to collect, has relatively few interfering compounds for the analytical process, and shows sample stability over time after death. The present study is an overview of VH physiology, drug transport and elimination. Collection, storage, analytical techniques and interpretation of results from qualitative and quantitative points of view are dealt with. The distribution of xenobiotics in VH samples is thus discussed and illustrated by a table reporting the concentrations of 106 drugs from more than 300 case reports. For this purpose, a survey was conducted of publications found in the MEDLINE database from 1969 through April 30, 2015.
Similar content being viewed by others

Introduction
Vitreous humor (VH), also known as the vitreous body, is a gelatinous substance contained in the posterior chamber of the eye, between the crystalline lens and the retina. It plays a mechanical role, keeping the retina in place and maintaining the spherical shape and tonus of the eyeball. There have been numerous studies of VH in various forensic applications. The first was to the assay of VH potassium, released during postmortem membrane degradation, as a means of estimating time of death [1]. Studies have found variable and sometimes contradictory results, depending on the authors, experimental conditions, analytic methods and statistical models [2–7]. Other means of achieving the same objective have been proposed: hypoxanthine assay isolated [8, 9] or associated to potassium [10, 11], amino acids [12] or creatinine assay [13], or VH absorbance assessment [14]. More recently, in a study using proton nuclear magnetic resonance (1H NMR) multivariate analysis of goat VH samples, Rosa et al. [15] recommended analyzing the global metabolite profile rather just than one or a few metabolites. A combined potassium and chlorine assay was reported for determining the immersion time of a body in cold water [16]. The other main applications of VH in forensics relate to postmortem biochemistry for screening or confirming preexistent pathology and determining cause of death (Table 1) [17–50].
In forensic toxicology, VH has served as an alternative matrix for more than 50 years [51, 52]. Its lack of vascularization, anatomic remoteness from viscera, and relative protection by the eyeball render VH a useful alternative when blood cannot be sampled (exsanguinated or fragmentary cadaver) or in the case of suspected postmortem redistribution [53–56] or contamination by bacteria or chemicals (e.g., embalming) [57, 58]. As it is easy to sample, and because it can be used for immunological analysis of certain groups of chemical substances [59], VH has even been recommended for immunoenzymatic screening on the site where a victim was discovered [60]. While screening applications are acknowledged for a large number of compounds, the use of VH analysis for interpreting concentrations seems more limited.
The present literature review has two objectives. The first, by describing the physiology of VH and drug transport and elimination, is to suggest possible lines of research to improve our knowledge of forensic applications of this matrix. The second is to develop a practical tool for use at all levels of investigation using VH: sampling and sample storage, analytic techniques and interpretation of results. For this second objective, we conducted a MEDLINE search with “vitreous” as a keyword combined with “forensic sciences”, “toxicology”, “postmortem”, “post-mortem”, “autopsy” and/or “chromatography drug” (update, April 30, 2015). The research was restricted to the organic compounds most frequently encountered in forensic toxicology: medical drugs and narcotics. Ethanol was the first substance for which VH concentrations were interpreted [61]. The importance of VH quantification of ethanol has been widely studied and thoroughly reviewed by Kugelberg and Jones [62], and is therefore not dealt with in the present review.
Physiology and pharmacokinetics
Anatomy and composition of vitreous humor
The crystalline lens separates the anterior chamber of the eye, which contains a liquid (aqueous humor), from the posterior chamber, which contains VH (Fig. 1) [63]. The posterior chamber is bounded at the back, from inside to outside, by the retinal membrane, the choroid and the sclera, and at the front by the ciliary body and the crystalline lens. VH is highly hydrated tissue, with 98–99.7 % water content, and mean volume of 4 mL. Its gelatinous structure is due to fibrillar proteins: primarily collagen fibers associated with glycosaminoglycan carbohydrates (mainly hyaluronic acid). As many as 1205 proteins have been identified in the VH [64]. It also contains electrolytes (such as sodium, potassium, chlorine, lactate and ascorbate), carbohydrates such as glucose, and small amounts of proteins other than collagen, including opticin. VH is avascular and very poor in cells. Of the few cells, hyalocytes, or vitreous cells, are involved in synthesizing the constituents of VH and in the adaptive immune response that limits intraocular inflammation [65]. A change in composition with age leads to gradual liquefaction. In persons at the age of 4 years, liquid VH accounts for 20 % of the total vitreous volume, increasing to over 50 % by 80–90 years of age [66].
The blood-retinal barrier
The blood-retinal barrier (BRB) is a selective barrier, like the blood-brain barrier [67]. It ensures the input required for retinal function and restricts that of possible pathogens (e.g., enzymes, anaphylatoxins) [68]. It actually comprises two barriers (Fig. 1). The first, comprising the retinal pigment epithelium (RPE) separating the retina from the choroid, is the outer BRB. RPE cells have the particularity of being bound together by intercellular junctions (zonula adherens and zonula occludens), forcing the intracellular transit of compounds. The second, which constitutes the non-fenestrated epithelium of the retinal blood vessels, is the inner BRB. The two barriers are not successive; rather, they are associated with the two retinal penetration pathways: choroid capillaries for the outer BRB and retinal capillaries for the inner BRB. Selectivity may be impaired by various pathologies, the most frequent of which are diabetic retinopathy and age-related macular degeneration [69].

Anatomy of the eye and the blood-retinal barrier (adapted from [63])
Xenobiotic exchange between blood and vitreous humor
In certain inflammatory or infectious ophthalmic pathologies, the posterior chamber is a drug target. Eyewashes and systemic treatments generally fail to achieve effective doses in VH; periocular and intra-vitreous injection is increasingly used for administration, although with a risk of infection. Compounds of forensic interest derive mainly from the systemic circulation, penetrating the VH from the retina via the BRB [70]. Two elimination routes from the VH have been described: a posterior pathway through the BRB in the opposite direction, and an anterior pathway by diffusion into the aqueous humor via the zonular spaces (Fig. 1), with elimination by the renewal of aqueous humor and uveal blood flow [71].
Factors affecting xenobiotic penetration into the vitreous humor
Drug penetration into the retina depends on various factors, including plasma concentration, compound physicochemical and pharmacological properties, distribution volume, plasma protein binding and relative BRB permeability [70]. Drugs may diffuse passively or be actively transported across the barrier: in general, the higher the molecular weight and/or hydrophilicity, the more likely that passage across a membrane involves active transport [72]. Given that only non-bound drugs can cross biological membranes, the percentage of plasma protein binding is another factor determining diffusion. In a study of numerous compounds of forensic interest, Holmgren et al. [73] found significant correlation between blood/VH concentration ratios and percentage of plasma protein binding.
There are many transmembrane proteins expressed in the BRB that can act as transporters, playing a role in drug bioavailability in the posterior chamber. Two main types can be distinguished: efflux pumps, belonging to the ABC (ATP-binding cassette) transporter superfamily, and uptake pumps, belonging to the solute carrier (SLC) transporter superfamily. The main efflux transporters identified in the eye are multidrug resistance (MDR) transporters, including P-glycoprotein (P-gp or MDR1), multidrug resistance protein (MRP) and breast cancer resistance protein (BCRP). Unlike passive diffusion, active transport may be limited by saturation, if drug concentration exceeds transport capacity, and competition with other compounds or inhibition by certain specific substrates. Animal studies of concomitant administration of verapamil, a P-gp inhibitor, found longer VH elimination half-life for quinidine, whether administered intravitreously [74] or intravenously [75]. In forensic toxicology, such interactions may have a significant impact on the interpretation of VH concentration, especially as it affects the VH/blood concentration ratio.
There have been numerous studies of the VH pharmacokinetics of drugs used in ophthalmic therapy (e.g., antibiotics and anti-inflammatory agents), and of their transporters in particular. On the other hand, much less is known about compounds of general interest in forensic toxicology. The relative VH bioavailability of memantine was reported to be only 0.02 % after intravenous administration compared to intravitreous administration as reference; the concentration peaked at 29.68 ± 13.9 min, and the rapid elimination half-life (<2 h) argued for retinal elimination by active transport [76]. The research by Pitkänen et al. [77] into the effect of beta-blocker size and lipophilicity on both uptake and efflux permeation through the outer BRB is especially interesting. The most hydrophilic beta-blocker showed permeability coefficients that were seven- to eightfold lower than those for the most lipophilic beta-blockers (metoprolol, timolol and betaxolol). Atenolol uptake and efflux speeds were identical, whereas more lipophilic beta-blockers showed penetration faster than outflow. This permeation asymmetry in highly lipophilic beta-blockers may be due to an active transport component. Moreover, VH diffusion time was longer for lipophilic than for hydrophilic beta-blockers (permeation lag time for betaxolol = 107 min, versus 38.7 min for atenolol). Pitkänen et al. [77] suggested that this could be the consequence of drug binding to melanin: the outer BRB contains melanin, which is a molecular site for basic and lipophilic drug binding and interaction [78], influencing permeation.
The various transport mechanisms and the factors governing them are important for the understanding and description of the distribution of drugs from blood to VH. These factors seem to affect low-molecular-weight molecules: i.e., most compounds of forensic interest. Evidence of their exact impact on the interpretation of VH concentrations, however, is rarely documented in the forensic literature, except for plasma protein binding.
Postmortem evolution of vitreous humor
VH tends to liquefy according to postmortem interval and local conditions. Postmortem evolution involves dehydration, which some authors have assessed in terms of increased creatinine concentration [56]. To our knowledge, there have been no studies of VH bioavailability relative to postmortem time. In our own experience, VH was sampled in 80 % of autopsies performed in the Forensic Medicine Institute of Lyon (France) between 2010 and 2013.
Analysis of vitreous humour
Sampling and storage conditions
VH is sampled by syringe, and aspiration should be slow, from the center of the eyeball, to avoid epithelial cells of the retina or iris. For the same reason, volume must be limited to 2 mL per eye, even though the total volume of VH is greater [79]. The volume withdrawn may be replaced by water or physiological saline in order to maintain the aspect of the eyeball [80]. Total versus micro-aliquot (50 µL) sampling procedures were compared in a rabbit model [81]. Micro-sampling seemed more reproducible for ion assay (calcium, chloride, potassium, sodium and phosphorus), but is too limited at present for forensic toxicology investigation.
Harper et al. [79], in a study of 51 paired VH and femoral blood samples, found that VH samples were less subject to bacterial contamination, which is an advantage in terms of sample and xenobiotic stability during storage; to enhance this advantage, the authors recommended sampling under aseptic conditions (for syringe and container) to avoid bacterial contamination.
Electrolyte and glucose samples have been reported to be lateralization-sensitive [82–85]. Rather than reflecting differential concentration between the two eyeballs, this may be a question of reproducibility related to the sampling procedure [81], of analytical problems due to the gelatinous consistency of VH [86], and of hematic contamination [87]. Bévalot et al. [88], in a series of 92 human autopsies, found the left/right VH differential to be non-significant for meprobamate. Findings were similar for 3,4-methylenedioxymethamphetamine (MDMA) [89], phenytoin [90], barbiturates [90] and cocaine [91]. However, for compounds such as digitalis-glycoside, which accumulates dramatically in the retina [92], sampling problems such as choroid and retinal cell aspiration may affect observed concentrations, and we recommend separate sampling of the two VH specimens, without pooling.
The container should be suitable for the small-sample volume: 5-mL tubes are preferred to the classical autopsy vials used for most samples in order to limit headspace gas volume and, thus, evaporation of volatile substances such as ethanol [93].
Although VH is generally considered unaffected by postmortem enzymatic and bacterial phenomena (few cells, protected from bacterial contamination, etc.), there have been several reports using stabilizers such as sodium fluoride (NaF) or potassium fluoride (KF) to block enzyme activity, causing neoformation or degradation of certain xenobiotics. Holmgren et al. [73] assessed the effect of KF on blood and VH concentration stability in 46 drugs. VH samples were divided in two, with KF added to one aliquot; all aliquots were conserved for 1 year at −20 °C. Among the 46 drugs, only zopiclone (n = 13) showed a significant reduction in mean concentration without the stabilizer (KF), from 0.15 to 0.03 µg/g. Moreover, 6-monoacetylmorphine (6-MAM) was exclusively detected in samples with KF (number of samples and concentration unknown).
Melo et al. [94] studied temperature effects on VH stability for benzodiazepines (lorazepam, estazolam, ketazolam and chlordiazepoxide). There was no significant degradation over 6 months in sub-zero storage (−20, −80 °C). Some benzodiazepines were relatively stable for a few weeks at +4 °C and +25 °C, whereas ketazolam degraded completely within 12 weeks at these temperatures. The stability of cocaine in ovine VH was studied by Rees et al. [95], with and without stabilizer (NaF), for 84 days at three temperatures: room temperature, +4 °C and −18 °C. At −18 °C, concentrations were stable (loss <15 %) for 84 days, both with and without NaF, and they were unstable at +4 °C, with a loss of 25 and 50 % by day 14, with and without NaF, respectively. The same team also studied the stability of 6-MAM [96]. The addition of 1.5 % NaF had a much clearer effect, limiting degradation to <10 % at −18 °C for 84 days, compared to 42 % by day 14 and 95 % by day 84 without a stabilizer. At +4 °C, likewise, degradation was <10 % on day 35 with NaF, versus 52 % by day 14 without NaF.
On the basis of these experimental data, sampling of VH from each eyeball, without pooling, would seem to be a good compromise. One sample, dedicated to toxicology analysis, should be performed with a stabilizer (1.5 % NaF or KF) to prevent ethanol neoformation and degradation of xenobiotics such as benzodiazepines, 6-MAM or cocaine. The other sample, without stabilizer, serves for biochemistry analysis. Both samples should be stored at −20 °C.
Sample preparation
The composition of VH makes it relatively “clean” in comparison to other autopsy matrices, and analysis does not require complex preparation. Some assays do not include an extraction step. Davis et al. [97] described the analysis of four antibiotics (fluoroquinolones) by direct injection using high-performance liquid chromatography/ultraviolet (HPLC/UV) and HPLC/fluorescence detection. Logan and Stafford [90] developed an HPLC neuroleptic assay based on injection after dilution and filtration using a preconcentration column. A similar process was also proposed for cocaine and benzoylecgonine [98].
The extraction techniques generally used for blood and other fluids provide cleaner extracts of VH than other matrices. Solid-phase extraction (SPE) is widely used, notably for assaying benzodiazepines [99], drugs of abuse (e.g., opiates, methadone, cocaine,) [100–104] and paracetamol [103]. Bévalot et al. [105] described a generic technique, validated on six compounds and assessed for implementation in large-scale screening. Liquid/liquid extraction (LLE) was used to assay colchicine [106], clotiapine [107], benzodiazepines [108], memantine after 9-fluorenylmethyl-chloroformate-chloride derivation [109], beta-blockers using an Extrelut® column [110], narcotics [111], arylcyclohexylamines (methoxetamine, 3-methoxyeticyclidine and 3-methoxyphencyclidine) [112] and LSD [113]. For narcotics analysis, microwave-assisted LLE provided better recovery and precision than SPE [114]. Other, less widespread techniques have also been tested. Disposable pipette extraction showed recovery in a range of 72–91 % for opiates, with a volume of 100 μL required for the VH sample [115]. Supercritical-phase extraction, which limits the need for an organic solvent, has been used with success in opiate assay [115, 116].
Several authors have recommended liquefying VH samples before analysis by immunoassay analyzers in order to avoid the phenomenon of clogging due to viscosity. Liquefaction techniques include enzymatic hydrolysis by hyaluronidase, heating, microfiltration, dilution and centrifugation [86].
Analytical techniques
Instrumental techniques for analysis have greatly improved in sensitivity and selectivity. Unlike more complex postmortem matrices (e.g., blood, tissue), where enhanced selectivity serves to palliate interference, VH, which can be considered a “clean” matrix, benefits fully from the gain in sensitivity. Analysis of cocaine and cocaine derivatives is a good example illustrating the whole range of analytical techniques that have been implemented for VH analysis: gas chromatography/flame ionization detector (GC/FID) [104], high-performance liquid chromatography/diode array detector (HPLC/DAD) [100, 114], GC/mass spectrometry (MS) [101, 117, 118], GC/tandem mass spectrometry (MS–MS) [95], HPLC/MS–MS [119], capillary electrophoresis/DAD [111] and immunoanalysis (cloned enzyme donor immunoassay [120], enzyme multiplied immunoassay technique [59]). Few screening techniques seem to have been specifically dedicated to VH analysis. Pelander et al. [121] described an assay based on HPLC coupled with time-of-flight mass spectrometry that allowed the detection of 70 compounds, and more recently, a method for the simultaneous screening and quantification of 24 analytes of forensic interest was described by Arora et al. [122].
Interpretation of vitreous humor analysis results
In 1969, Felby and Olsen [51] published one of the first reports of postmortem medical drug analysis in VH. In this study, VH concentrations of barbiturates in a given individual were identical to blood ultrafiltrate levels but different from those in total blood, showing that barbiturates penetrate VH by passive diffusion. The authors also suggested an effect of plasma protein binding on VH penetration in certain barbiturates (phenobarbital, aprobarbital and barbital). No significant differences in concentration were found between left and right VH drug levels. The authors suggested that an ultrafiltrate/VH concentration ratio >1 indicated death earlier than the equilibrium phase, and thus shortly after intake. They considered analysis technically easier in VH than in blood, especially in the case of putrefaction. Thus, from its earliest applications in postmortem toxicology, various advantages of VH analysis have been highlighted, and most have since been studied in numerous medical and narcotic drug families.
Case reports
Table 2 [60, 101, 103, 106, 107, 113, 123–256] presents cause of death and blood and VH concentrations from more than 100 case reports. It is intended as a practical tool for toxicologists in analyzing and interpreting results for specific compounds, which were arranged in the alphabetical order.
Qualitative interpretation
Toxicologic analysis of VH is of undisputed qualitative interest, as seen from the large number of xenobiotics detected (Table 2). Its qualitative importance compared to blood and other matrices has been assessed for various groups of compounds.
Opiates and opioids
Interpretation of recent heroin intake via its tracer 6-MAM has been a particular focus of study. In a series of 29 deaths from opiates, Pragst et al. [257] reported that in two cases, 6-MAM was detected in VH but not in urine, despite generally higher 6-MAM concentrations in urine than in VH. Blood concentrations of 6-MAM were not reported in this study. Wyman and Bultman [258], Rees et al. [259], Antonides et al. [118] and Scott et al. [116] showed that in heroin-related deaths, if only blood were analyzed, 6-MAM would go undetected in 36 % (n = 25), 59 % (n = 70), 50 % (n = 12) and 25 % (n = 20) of cases, respectively. Two hypotheses have been put forward to explain why 6-MAM should be detected more often in VH than in blood: good membrane crossing due to lipophilicity (logP = 1.56) and absence of esterase in VH, thus limiting degradation. The second hypothesis, however, is to be taken with caution. There is, in fact, esterase activity in VH [260]; and acetylcholinesterase, causing heroin to hydrolyze into 6-MAM and 6-MAM into morphine, is present in VH in many animal species [261]. In the absence of hard evidence, the possibility that this activity is merely weaker or more saturable in VH than in blood or other organs cannot be excluded. Another hypothesis holds that the properties of esterases in VH are different from those in blood, as demonstrated for brain synapse acetylcholinesterase, which was unable to hydrolyze heroin, unlike erythrocyte acetylcholinesterase [262].
When 6-MAM is undetected in blood, the morphine/codeine ratio in blood or urine is sometimes used to determine whether the detected morphine resulted from codeine metabolism (morphine/codeine ratio <1) or from direct intake of morphine, and thus possibly of heroin (morphine/codeine ratio >1) [263]. Lin et al. [264] reported that the morphine/codeine ratios in 223 opiate-positive VH samples were systematically >1 when 6-MAM was also detected, and moreover, were close to these found in blood. Rees et al. [259] also found that the VH morphine/codeine ratio was useful in revealing heroin intake. The low codeine concentrations found in VH, however, close to the quantification limit, may hinder the use of VH for this purpose.
Benzodiazepines
In a postmortem analysis of 3 nitro-benzodiazepines (nitrazepam, flunitrazepam and clonazepam) and their 7-amino metabolites, Robertson and Drummer [265] reported that in 15 % of cases in which 7-amino metabolites were detected in blood, they were not detected in VH. Moreover, the parent drugs were detected in VH in only 10 % of cases versus 30 % in blood. This differential positivity may have been due to the fact that VH benzodiazepine levels were generally one-third of those in blood. In a series of 17 postmortem cases, Scott and Oliver [266] assayed three benzodiazepines (diazepam, nordazepam and temazepam) in blood and in VH; in seven cases, one or more were detected in blood but not in VH. These results may reflect the fact that benzodiazepines are highly bound to proteins; their neutral or weak acidic properties further decrease diffusion into VH, which may be alkaline, as observed in postmortem samples, with a mean pH value of 8.3 and range of 7.3–9.1 [267].
Other compounds
VH also appears useful, in the absence of blood, for revealing use of cocaine [104, 268]. Moreover, the detection window is wider than in blood, as seen from the cases where cocaine is detected in VH but not in blood [91]. Jenkins and Oblock [269] showed that phencyclidine (PCP) was systematically detected in VH when detected in blood and/or urine. Cox et al. [270] confirmed this qualitative interest for PCP in a series of 26 autopsies. Oxycodone [271] and phenytoin [90] were systematically detected in VH when detected in blood in a respective series of 30 and 12 cases.
These studies confirm the usefulness of VH in detecting xenobiotic consumption. Moreover, for certain compounds (6-MAM, cocaine, and PCP), the VH detection window is wider than that for blood. For other compounds such as benzodiazepines, the qualitative importance of VH seems more limited. However, this may be related to analytic techniques: the same analysis protocols as in blood are usually applied in VH, despite the fact that concentrations are generally lower. Dedicated techniques developed and validated for VH could lower detection thresholds and increased detection rates. This may be difficult to achieve with present-day analytic techniques for some compounds with very low VH concentrations, however, such as tetrahydrocannabinol (THC) or its metabolites (11-OH-THC, THC-COOH or THC-COOH glucuronide) [272].
Quantitative interpretation
Opiates
In 20 cases of death by heroin, Scott and Oliver [116] reported lower morphine concentrations in VH than in blood, with a significant correlation (r 2 = 0.697) between the two. In light of this correlation, the authors considered VH to be the “ideal matrix” for analyzing morphine in the absence of blood. 6-MAM concentrations in VH were higher than in blood, but without correlation. Rees et al. [259] confirmed these findings, but pointed out that the correlation between VH and blood morphine concentrations depends on the intake-to-death interval, and may also be affected by intake modalities. The authors concluded that blood morphine concentration could not be extrapolated from the VH level. In the same study, it was shown that codeine concentrations were higher in VH than in blood, with a correlation with femoral blood level (r 2 = 0.672). The authors suggested that codeine’s greater lipophilicity might account for higher VH concentrations than blood concentrations, unlike with morphine (logP codeine = 1.39; morphine = 0.87). Knittel et al. [271] reported that oxycodone showed a positive linear correlation between VH and blood concentrations, but with too great a scatter for extrapolation of levels from VH to blood.
Cocaine
More than in the case of other substances, postmortem blood concentrations of cocaine rarely correspond to those at time of death, largely due to strong in corpore and in vitro degradation. VH is one of the tissues in which xenobiotic composition is considered relatively stable over the early postmortem period, and its application as a matrix for cocaine quantification has naturally been widely studied. Results from several studies have, however, been divergent.
Antonides et al. [118], in a series of 40 autopsies, reported that VH cocaine concentrations were higher than in blood in 72 % of cases. This was confirmed by Logan and Stafford [98], who, moreover, found no correlation between the two matrices. The authors attributed these higher concentrations in VH to greater degradation of cocaine in blood. To circumvent the uncertainties of degradation, Duer et al. [119] investigated correlation for what they termed “total cocaine”, corresponding to the sum of the concentrations (in µmol/L) of cocaine and its metabolites (ecgonine, ecgonine methyl ester and benzoylecgonine). In this condition, correlations of 0.939 and 0.883 were obtained between VH and femoral blood levels, and between VH and cardiac blood levels, respectively. Thus the authors concluded that VH was as reliable as blood for cocaine analysis. Fernandez et al. [104] reported VH cocaine concentrations near to those of blood (mean ratio, 1.03; range, 0.36–2.94), with a significant correlation coefficient (r = 0.71). According to the authors, VH could confirm the presence of cocaine in absence of blood but by no means could it estimate the blood concentration accurately. Carvahlo et al. [273] reported excellent correlation between VH and blood levels for cocaine (r = 0.98) and benzoylecgonine (r = 0.95) in 7 deaths by cocaine overdose but not in the 11 cases of accidental death. Another study showed that mean concentrations (n = 53) of cocaine and cocaethylene did not significantly differ between blood and VH, unlike benzoylecgonine, and reported correlations between blood and VH levels for benzoylecgonine (r = 0.763) and cocaine (r = 0.854), but not for cocaethylene (r = 0.343) [268].
These divergences highlight the importance of parameters that cannot be controlled in postmortem cases (intake-to-death time, time from death, postmortem redistribution and stability). Thus, blood cocaine level at death may not be extrapolated with precision from VH level alone.
Benzodiazepines
In 52 postmortem cases in which nitro-benzodiazepines (nitrazepam, flunitrazepam and clonazepam) and their 7-amino metabolites were assayed in blood and in VH, Robertson and Drummer [265] reported a correlation of r = 0.626 for the parent drugs and r = 0.764 for the metabolites. According to the authors, such reasonable positive correlations, also found for the metabolites in urine, bile and liver, enabled more precise interpretation of the blood data. In a series of 17 autopsies, Scott and Oliver [266] reported coefficients of determination (r 2) of 0.788 for temazepam, 0.723 for diazepam and 0.068 for nordiazepam. In all cases, the VH levels were lower than in blood. Although correlations were identified, the authors reported a wide scatter in the results, probably related to variations in parameters such as intake modality, intake-to-death interval and time to autopsy. Finally, another autopsy study showed non-significant trends for nordazepam (n = 58, r 2 = 0.473), bromazepam (n = 31, r 2 = 0.345) and oxazepam (n = 28, r 2 = 0.588) between VH and blood [274]. It thus seems that quantitative interpretation of VH benzodiazepine concentrations cannot consist in straightforward extrapolation of blood levels, given the weak correlations and the scatter found in the results.
Gamma-hydroxybutyric acid
Gamma-hydroxybutyric acid (GHB) is naturally present in organisms. Moreover, potentially significant postmortem neoformation of an unclear origin has been reported [275]. GHB was also used in anesthesiology, and is misused as a recreational drug and to incapacitate a victim. The main objective in the interpretation of postmortem blood concentration is to determine whether the origin was purely endogenous or involved exogenous administration. Postmortem neoformation misleads us into interpreting an elevated blood level as indicative of an exogenous origin. VH is one of the alternative matrices proposed to confirm elevation in cardiac blood level [276, 277]. Kintz et al. [277] described an interpretation tree for determining exogenous origin, with a 50 mg/L threshold in cardiac blood, and confirmed, when positive, by the same threshold in femoral blood and VH. Moriya and Hashimoto [278] suggested a 10 mg/L threshold in urine and VH. Another study, in which cardiac and femoral blood, VH, urine and cerebrospinal fluid was analyzed, found that VH levels could exceed blood levels, and indeed, were sometimes the highest of any of the five matrices [275]. VH concentrations were systematically below 50 mg/mL but were often greater than 10 mg/mL. For interpretation of blood GHB, the authors concluded that VH should not be the sole alternative matrix. In a recent review, Castro et al. [279] stressed that the thresholds should be seen as interpretation aids on a case-by-case basis rather than as hard facts. Moreover, the authors observed that the thresholds for GHB reported in the various matrices showed a tendency to become lower with increasing expertise in sampling and storage.
Insulin
Insulin determination in postmortem blood is complex, especially in hemolyzed specimens, due to matrix interference and insulin degradation by insulin-degrading enzymes and a non-proteolytic process initiated by hemoglobin [280]. Thus, interpretation of blood insulin concentrations in postmortem investigation is often tricky. The determination of insulin in VH appears to hold promise, due to low analytical background, less pronounced postmortem changes [190] and the absence of hemoglobin. In four studies of populations with no history of diabetes or in subjects with type 1 diabetes where the cause of death was unrelated to insulin overdose, insulin was either undetectable in VH or shown in concentrations close to the limit of detection. These studies used various analytical methods: LC/MS–MS, limit of detection (LOD) = 2.4–4.8 µIU/mL, n = 10 [190]; LC/MS–MS, LOD = 4.5 µIU/mL, n = 46 [192]; chemiluminescence enzyme immunoassay, LOD = 0.2 µIU/mL, n = 40 [191]. The fourth study, by Nowicka et al. [193], was a comment article which reported insulin determination by immunoradiometric assay on 93 VH samples from a random autopsy population: insulin was not detected in 86 cases (LOD = 0.5 µIU/mL) and ranged from 1.42 to 24.42 µIU/mL in the seven remaining cases (the outlier with 24.42 µIU/mL was probably related to insulin administration, as the subject was not known to be diabetic, and an insulin syringe was found near the corpse). In some reported cases of death related to insulin overdose in which VH analysis has been used (cf. Table 2), VH insulin concentrations were notably higher (from 11.5 to 103 µIU/mL: five cases). These studies demonstrated that VH is an interesting matrix to sample and analyze as a complement to blood or serum in postmortem investigation of insulin intoxication. However, although Ojanperä et al. [281] demonstrated success in the detection of insulin or metabolites by HPLC coupled with high-resolution mass spectrometry in three cases of non-insulin-related death of diabetes mellitus subjects with postmortem intervals between 4 and 8 days, more data are needed on insulin stability over postmortem intervals.
MDMA
In a rabbit model, De Letter et al. [89] demonstrated a correlation between MDMA concentrations in VH and blood after concentrations reached equilibrium (i.e., about 1 h after administration). In this study, VH concentrations were more stable than those in blood in the case of long postmortem time (73 h), and thus more representative of antemortem blood levels.
Other compounds
Jenkins and Oblock [269] and Cox et al. [270] found no correlation between blood and VH PCP levels in 30 and 26 cases, respectively. Holmgren et al. [73] studied correlations between blood and VH concentrations in 46 compounds of various groups of drugs in samples stored with KF at −20 °C for 12 months. Correlations emerged for a half of the substances (n = 23), including amphetamine, diltiazem, tramadol and venlafaxine, while for compounds such as clomipramine, clozapine and sertraline, no correlation was observed. Given the lack of discussion of these results and the small sample sizes for certain drugs (e.g., tramadol: n = 4), these findings do not warrant extrapolation from VH to blood, but may serve as a basis for future studies.
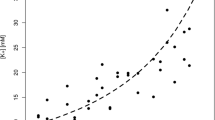
As described above, the main approach used for the extrapolation of blood levels at death from VH levels is based on correlations. However, Bévalot et al. [88] proposed the use of statistical data processing for interpreting meprobamate concentrations in VH, determining a VH concentration threshold to distinguish therapeutic from overdose levels. In a 117-case series (40 deemed therapeutic, 77 overdose), a threshold of 28 mg/L was determined for VH meprobamate, with sensitivity of 0.95 and absolute specificity of 1. In the same case series, an interpretation table was described for the probability of an association between a given VH level and a blood level in one of four concentration ranges: <30, 30–50, 50–100 and >100 mg/L. Using a similar approach, Parker and McIntyre [282] reported that VH quetiapine concentrations in non-toxic deaths (n = 8) ranged between 0.10 and 0.22 mg/L (95 % confidence interval [CI]), and in toxic deaths (n = 8) between 0.74 and 1.74 mg/L (95 % CI).
Survival time
Several authors have proposed the use of blood/VH concentration ratios for estimating survival time (intake-to-death interval) based on the time of distribution from blood to VH: soon after intake, the ratio is higher than when equilibrium between the two matrices is reached. Using inquest data providing an estimate of last intake time, Scott and Oliver [266] showed that the blood/VH concentration ratios for benzodiazepines were higher in rapid death. However, the authors stressed that data were lacking for various postmortem factors notably redistributions that were liable to impact the ratio. Teixeira et al. [283] demonstrated in a rabbit model that the blood/VH concentration ratio after intramuscular administration of diazepam was 20 up to 1 h, and then fell to 4.5 by 6 h. At equilibrium, 1–2 h post-administration, the ratio was 10. The authors suggested the blood/VH concentration ratio as a “complementary tool” for determining intake-to-death time, without giving further details of how it could be used. Antonides et al. [118], also investigating circumstances of death, reported that when blood concentrations of cocaine were higher than in VH, death had occurred sooner after intake. In these cases, blood benzoylecgonine levels were up to twofold higher than VH levels.
Postmortem redistribution
De Letter et al. [56] reported that the postmortem distribution of MDMA in a rabbit model showed VH concentrations to be more stable, and representative of antemortem rather than postmortem blood levels. VH MDMA levels, however, were especially elevated in the wall of the eyeball, so diffusion was a possibility, especially in the case of long postmortem time. The findings of a previous study of 73-h postmortem evolution of VH MDMA concentrations suggested that such accumulation contributed only moderately to VH concentration [89]. For digoxin, a digitalis derivative, Ritz et al. [92] reported very high concentrations in choroid and retinal tissue (63.9–485 ng/g), close to levels found in cardiac muscle and higher than those in VH (2.2–7.1 ng/mL), based on the results of an autopsy series (n = 19). A similar distribution was found for digitoxin [284]. The authors suggested that these differences could induce postmortem redistribution from choroid and retinal tissue to VH. In a study of postmortem redistribution of cocaine in a pig model, McKinney et al. [285] sacrificed the animals 5 min after intravenous administration, and performed sampling at sacrifice and 8 h later. While blood concentration had not changed, VH levels had risen considerably. The authors expected such a rise caused by redistribution from periorbital blood, but not to that degree: baseline concentrations were significantly lower in VH (mean = 939 ng/mL) than in blood (mean = 3245 ng/mL), whereas by 8 h, the two were close (VH, mean = 3067 ng/mL; blood, mean = 3568 ng/mL). They hypothesized that intraocular tissue such as the retina might be a region of accumulation, with postmortem release to VH. Teixeira et al. [283] found a twofold elevation of VH diazepam concentrations and a threefold elevation of nordazepam during a 24 h postmortem period in an animal model. Maskell et al. [286] investigated the postmortem redistribution of the heroin metabolites morphine and morphine-3-glucuronide (M3G) in nine biological matrices in a rabbit model. In VH collected at 24 h postmortem, a 181 % increase in free morphine concentration and a 425 % increase in total morphine concentration were observed. For M3G, the increase in concentration was 1.002 %, and among the nine matrices studied, VH was the only one in which the M3G concentration increased. The authors explained the increase in morphine and M3G concentrations in VH by diffusion from “tissue” without, however, specifying the tissue in question.
These studies suggest that VH is a matrix protected against the main sources of postmortem redistribution from the abdominal cavity, but that ocular tissue may be a region of accumulation of xenobiotics liable to diffuse into VH postmortem.
Conclusions
When blood is lacking or is modified by postmortem factors, alternative matrices may be useful. An ideal alternative should enable detection of the same xenobiotics as found in blood, at correlated concentrations, and without the postmortem effects. VH is the matrix that probably comes closest to this ideal. Moreover, from a practical point of view, VH is easy to sample, and compounds within it tend to be stable if certain storage conditions are warranted and analysis is straightforward, with no more preparation required than is necessary to ensure “cleanliness”. It is of particular screening interest in the absence of blood, as most compounds of forensic interest are detected from VH. For several compounds (6-MAM, PCP, cocaine), moreover, the detection window is wider than that in blood. This qualitative interest could be enhanced by dedicated techniques achieving lower detection thresholds than those of most other complex forensic matrices.
The limitations of VH for the purpose of forensic toxicology largely concern quantitative interpretation. Various controlled animal or autopsy studies have been conducted to determine the implication of a xenobiotic in a victim’s death by interpreting only VH concentrations. Their findings show that VH and blood concentrations do not correlate for all compounds, and that in others, the scatter of the autopsy data usually precludes extrapolation to blood concentrations without significant error. This scatter reflects various non-controllable and often unknown parameters such as survival time, postmortem time, ophthalmic pathology and drug interaction. To optimize quantitative interpretation, various possibilities must be considered, the first of which is improving our knowledge of xenobiotic distribution in VH. Although diffusion seems to be the preponderant mechanism taking place for most compounds, diffusion from blood is not merely passive, and an enhanced understanding is needed of the role and mechanisms of active transport in antemortem distribution of compounds of forensic interest. Second, it is important to explore distribution in ocular tissues, and particularly in the choroid and retina, which may act as accumulation regions, with possible postmortem redistribution toward VH. Third, statistical tools must be developed and implemented in order to assess the uncertainty of interpretation of VH concentrations to the greatest degree possible. Even more than for concentrations in blood, it is important to report and discuss the uncertainty of findings obtained from alternative matrices according to the specific data for forensic cases.
References
Adelson L, Sunshine I, Rushforth NB, Mankoff M (1963) Vitreous potassium concentration as an indicator of the postmortem interval. J Forensic Sci 8:503–514
Madea B, Rodig A (2006) Time of death dependent criteria in vitreous humor: accuracy of estimating the time since death. Forensic Sci Int 164:87–92
Coe JI (1989) Vitreous potassium as a measure of the postmortem interval: an historical review and critical evaluation. Forensic Sci Int 42:201–213
Lange N, Swearer S, Sturner WQ (1994) Human postmortem interval estimation from vitreous potassium: an analysis of original data from six different studies. Forensic Sci Int 66:159–174
Munoz Barus JI, Febrero-Bande M, Cadarso-Suarez C (2008) Flexible regression models for estimating postmortem interval (PMI) in forensic medicine. Stat Med 27:5026–5038
Bocaz-Beneventi G, Tagliaro F, Bortolotti F, Manetto G, Havel J (2002) Capillary zone electrophoresis and artificial neural networks for estimation of the post-mortem interval (PMI) using electrolytes measurements in human vitreous humour. Int J Legal Med 116:5–11
Chandrakanth HV, Kanchan T, Balaraj BM, Virupaksha HS, Chandrashekar TN (2013) Postmortem vitreous chemistry—an evaluation of sodium, potassium and chloride levels in estimation of time since death (during the first 36 h after death). J Forensic Leg Med 20:211–216
Munoz JI, Costas E, Rodriguez-Calvo MS, Suarez-Penaranda JM, Lopez-Rivadulla M, Concheiro L (2006) A high-performance liquid chromatography method for hypoxanthine determination in vitreous humour: application to estimation of post mortem interval. Hum Exp Toxicol 25:279–281
Camba A, Lendoiro E, Cordeiro C, Martinez-Silva I, Rodriguez-Calvo MS, Vieira DN, Munoz-Barus JI (2014) High variation in hypoxanthine determination after analytical treatment of vitreous humor samples. Forensic Sci Med Pathol 10:627–633
James RA, Hoadley PA, Sampson BG (1997) Determination of postmortem interval by sampling vitreous humour. Am J Forensic Med Pathol 18:158–162
Munoz Barus JI, Suarez-Penaranda JM, Otero XL, Rodriguez-Calvo MS, Costas E, Miguens X, Concheiro L (2002) Improved estimation of postmortem interval based on differential behaviour of vitreous potassium and hypoxantine in death by hanging. Forensic Sci Int 125:67–74
Girela E, Villanueva E, Irigoyen P, Girela V, Hernandez-Cueto C, Peinado JM (2008) Free amino acid concentrations in vitreous humor and cerebrospinal fluid in relation to the cause of death and postmortem interval. J Forensic Sci 53:730–733
Piette M (1989) The effect of the post-mortem interval on the level of creatine in vitreous humour. Med Sci Law 29:47–54
Gong ZQ, Xu XM, Sun YG, Wang DW, Zhou XM, Xu FY, Zhou JL (2001) Study on the relationship between postmortem interval and the change of absorbance in vitreous humor of rabbit after death (article in Chinese with English abstract). Fa Yi Xue Za Zhi 17:77–78
Rosa MF, Scano P, Noto A, Nioi M, Sanna R, Paribello F, De-Giorgio F, Locci E, d’Aloja E (2015) Monitoring the modifications of the vitreous humor metabolite profile after death: an animal model. Biomed Res Int 2015:627201
Bray M (1985) Chemical estimation of fresh water immersion intervals. Am J Forensic Med Pathol 6:133–139
Stolyszewski I, Niemcunowicz-Janica A, Pepinski W, Spolnicka M, Zbiec R, Janica J (2007) Vitreous humour as a potential DNA source for postmortem human identification. Folia Histochem Cytobiol 45:135–136
Tappero P, Merlino C, Cavallo R, Vai S, Negro Ponzi A (1989) Anti-HIV antibodies in postmortem vitreous humor. Panminerva Med 31:187–188
Grupenmacher F, Silva FM, Abib FC, Grupenmacher L, Silva AC, de Almeida PT (1991) Determination of cadaveric antibody against HIV in vitreous humor of HIV-positive patients: potential use in corneal transplantation. Ophthalmologica 203:12–16
Mietz H, Heimann K, Kuhn J, Wieland U, Eggers HJ (1993) Detection of HIV in human vitreous. Int Ophthalmol 17:101–104
Schor P, Sato EH, Kuroda A, Barros PS, Marcopito LA, Belfort R Jr (1997) HIV-antibody detection in vitreous humor and serum by enzyme immunosorbent assay and particle agglutination test. Cornea 16:541–544
Klatt EC, Shibata D, Strigle SM (1989) Postmortem enzyme immunoassay for human immunodeficiency virus. Arch Pathol Lab Med 113:485–487
Cowan WT Jr, Wahab S, Lucia HL (1988) Detection of human immunodeficiency virus antigen in vitreous humor. J Clin Microbiol 26:2421–2422
Karhunen PJ, Brummer-Korvenkontio H, Leinikki P, Nyberg M (1994) Stability of human immunodeficiency virus (HIV) antibodies in postmortem samples. J Forensic Sci 39:129–135
Gregora Z, Bruckova M (1986) Attempts to demonstrate specific antibody responses in the vitreous body of the eye. J Hyg Epidemiol Microbiol Immunol 30:195–198
Comment L, Reggiani Bonetti L, Mangin P, Palmiere C (2014) Measurement of beta-tryptase in postmortem serum, pericardial fluid, urine and vitreous humor in the forensic setting. Forensic Sci Int 240:29–34
Blumenfeld TA, Mantell CH, Catherman RL, Blanc WA (1979) Postmortem vitreous humor chemistry in sudden infant death syndrome and in other causes of death in childhood. Am J Clin Pathol 71:219–223
Richards RG, Fukumoto RI, Clardy DO (1983) Sudden infant death syndrome: a biochemical profile of postmortem vitreous humor. J Forensic Sci 28:404–414
Huser CJ, Smialek JE (1986) Diagnosis of sudden death in infants due to acute dehydration. Am J Forensic Med Pathol 7:278–282
Poulsen JP, Rognum TO, Hauge S, Oyasaeter S, Saugstad OD (1993) Post-mortem concentrations of hypoxanthine in the vitreous humor—a comparison between babies with severe respiratory failure, congenital abnormalities of the heart, and victims of sudden infant death syndrome. J Perinat Med 21:153–163
Lapinlampi TO, Hirvonen JI (1986) Catecholamines in the vitreous fluid and urine of guinea pigs dying of cold and the effect of postmortem freezing and autolysis. J Forensic Sci 31:1357–1365
Devos C, Piette M (1989) Hypothermia and combined post-mortem determination of amylase and isoamylase in the serum and the vitreous humour. Med Sci Law 29:218–228
Coe JI (1984) Hypothermia: autopsy findings and vitreous glucose. J Forensic Sci 29:389–395
Palmiere C, Bardy D, Letovanec I, Mangin P, Augsburger M, Ventura F, Iglesias K, Werner D (2013) Biochemical markers of fatal hypothermia. Forensic Sci Int 226:54–61
Rognum TO, Saugstad OD, Oyasaeter S, Olaisen B (1988) Elevated levels of hypoxanthine in vitreous humor indicate prolonged cerebral hypoxia in victims of sudden infant death syndrome. Pediatrics 82:615–618
McDonald L, Sullivan A, Sturner WQ (1981) Zinc concentrations in vitreous humor: a postmortem study comparing alcoholic and other patients. J Forensic Sci 26:476–479
Piette M, Timperman J, Vanheule A (1986) Is zinc a reliable biochemical marker of chronic alcoholism in the overall context of a medico-legal autopsy? Forensic Sci Int 31:213–223
Berkowicz A, Wallerstedt S, Wall K, Denison H (2003) Analysis of carbohydrate-deficient transferrin (CDT) in vitreous humour as a forensic tool for detection of alcohol misuse. Forensic Sci Int 137:119–124
Osuna E, Perez-Carceles MD, Moreno M, Bedate A, Conejero J, Abenza JM, Martinez P, Luna A (2000) Vitreous humor carbohydrate-deficient transferrin concentrations in the postmortem diagnosis of alcoholism. Forensic Sci Int 108:205–213
Pounder DJ, Stevenson RJ, Taylor KK (1998) Alcoholic ketoacidosis at autopsy. J Forensic Sci 43:812–816
Palmiere C, Augsburger M (2014) The postmortem diagnosis of alcoholic ketoacidosis. Alcohol Alcohol 49:271–281
Ross MP, Spiller HA (1999) Fatal ingestion of sodium hypochlorite bleach with associated hypernatremia and hyperchloremic metabolic acidosis. Vet Hum Toxicol 41:82–86
Luna A, Jimenez-Rios G, Villanueva E (1985) Aminopeptidase and cathepsin A activity in vitreous humor in relation to causes of death. Forensic Sci Int 29:171–178
Vieweg WV, David JJ, Rowe WT, Wampler GJ, Burns WJ, Spradlin WW (1985) Death from self-induced water intoxication among patients with schizophrenic disorders. J Nerv Ment Dis 173:161–165
Chong AP, Aw SE (1986) Postmortem endocrine levels in the vitreous humor. Ann Acad Med Singapore 15:606–609
Canfield DV, Chaturvedi AK, Boren HK, Veronneau SJ, White VL (2001) Abnormal glucose levels found in transportation accidents. Aviat Space Environ Med 72:813–815
Meatherall R, Younes J (2002) Fatality from olanzapine induced hyperglycemia. J Forensic Sci 47:893–896
Peclet C, Picotte P, Jobin F (1994) The use of vitreous humor levels of glucose, lactic acid and blood levels of acetone to establish antemortem hyperglycemia in diabetics. Forensic Sci Int 65:1–6
Sippel H, Mottonen M (1982) Combined glucose and lactate values in vitreous humour for postmortem diagnosis of diabetes mellitus. Forensic Sci Int 19:217–222
Fanton L, Bevalot F, Cartiser N, Palmiere C, Le Meur C, Malicier D (2010) Postmortem measurement of human chorionic gonadotropin in vitreous humor and bile. J Forensic Sci 55:792–794
Felby S, Olsen J (1969) Comparative studies of postmortem barbiturate and meprobamate in vitreous humor, blood and liver. J Forensic Sci 14:507–514
Sturner WQ, Garriott JC (1975) Comparative toxicology in vitreous humor and blood. Forensic Sci 6:31–39
Hilberg T, Ripel A, Slordal L, Bjorneboe A, Morland J (1999) The extent of postmortem drug redistribution in a rat model. J Forensic Sci 44:956–962
Pélissier-Alicot A-L, Gaulier J-M, Dupuis C, Feuerstein M, Léonetti G, Lachâtre G, Marquet P (2006) Post-mortem redistribution of three beta-blockers in the rabbit. Int J Legal Med 120:226–232
De Letter EA, Belpaire FM, Clauwaert KM, Lambert WE, Van Bocxlaer JF, Piette MH (2002) Post-mortem redistribution of 3,4-methylenedioxymethamphetamine (MDMA, “ecstasy”) in the rabbit. Part II: post-mortem infusion in trachea or stomach. Int J Legal Med 116:225–232
De Letter EA, Clauwaert KM, Belpaire FM, Lambert WE, Van Bocxlaer JF, Piette MH (2002) Post-mortem redistribution of 3,4-methylenedioxymethamphetamine (MDMA, “ecstasy”) in the rabbit. Part I: experimental approach after in vivo intravenous infusion. Int J Legal Med 116:216–224
Coe JI (1976) Comparative postmortem chemistries of vitreous humor before and after embalming. J Forensic Sci 21:583–586
Steinhauer JR, Volk A, Hardy R, Konrad R, Daly T, Robinson CA (2002) Detection of ketosis in vitreous at autopsy after embalming. J Forensic Sci 47:221–223
Vogel J, Hodnett CN (1981) Detection of drugs in vitreous humor with an enzyme immunoassay technique. J Anal Toxicol 5:307–309
Fucci N, De Giovanni N, De Giorgio F, Liddi R, Chiarotti M (2006) An evaluation of the Cozart RapiScan system as an on-site screening tool for drugs of abuse in a non-conventional biological matrix: vitreous humor. Forensic Sci Int 156:102–105
Sturner WQ, Coumbis RJ (1966) The quantitation of ethyl alcohol in vitreous humor and blood by gas chromatography. Am J Clin Pathol 46:349–351
Kugelberg FC, Jones AW (2007) Interpreting results of ethanol analysis in postmortem specimens: a review of the literature. Forensic Sci Int 165:10–29
Urtti A (2006) Challenges and obstacles of ocular pharmacokinetics and drug delivery. Adv Drug Deliv Rev 58:1131–1135
Murthy KR, Goel R, Subbannayya Y, Jacob HKC, Murthy PR, Manda SS, Patil AH, Sharma R, Sahasrabuddhe NA, Parashar A, Nair BG, Krishna V, Prasad TSK, Gowda H, Pandey A (2014) Proteomic analysis of human vitreous humor. Clin Proteomics 11:29
Sonoda K-H, Sakamoto T, Qiao H, Hisatomi T, Oshima T, Tsutsumi-Miyahara C, Exley M, Balk SP, Taniguchi M, Ishibashi T (2005) The analysis of systemic tolerance elicited by antigen inoculation into the vitreous cavity: vitreous cavity-associated immune deviation. Immunology 116:390–399
Le Goff MM, Bishop PN (2008) Adult vitreous structure and postnatal changes. Eye (Lond) 22:1214–1222
Steuer H, Jaworski A, Elger B, Kaussmann M, Keldenich J, Schneider H, Stoll D, Schlosshauer B (2005) Functional characterization and comparison of the outer blood-retina barrier and the blood-brain barrier. Invest Ophthalmol Vis Sci 46:1047–1053
Jordan J, Ruiz-Moreno JM (2013) Advances in the understanding of retinal drug disposition and the role of blood-ocular barrier transporters. Expert Opin Drug Metab Toxicol 9:1181–1192
Cummings M, Cunha-Vaz J (2008) Treatment of neovascular age-related macular degeneration in patients with diabetes. Clin Ophthalmol 2:369–375
Cunha-Vaz JG (2004) The blood-retinal barriers system. Basic concepts and clinical evaluation. Exp Eye Res 78:715–721
Gaudana R, Ananthula HK, Parenky A, Mitra AK (2010) Ocular drug delivery. AAPS J 12:348–360
Mannermaa E, Vellonen KS, Urtti A (2006) Drug transport in corneal epithelium and blood-retina barrier: emerging role of transporters in ocular pharmacokinetics. Adv Drug Deliv Rev 58:1136–1163
Holmgren P, Druid H, Holmgren A, Ahlner J (2004) Stability of drugs in stored postmortem femoral blood and vitreous humor. J Forensic Sci 49:820–825
Majumdar S, Hippalgaonkar K, Srirangam R (2009) Vitreal kinetics of quinidine in rabbits in the presence of topically coadministered P-glycoprotein substrates/modulators. Drug Metab Dispos 37:1718–1725
Hippalgaonkar K, Srirangam R, Avula B, Khan IA, Majumdar S (2010) Interaction between topically and systemically coadministered P-glycoprotein substrates/inhibitors: effect on vitreal kinetics. Drug Metab Dispos 38:1790–1797
Prieto E, Perez S, Pablo LE, Garcia MA, Bregante MA (2014) Vitreous pharmacokinetics and bioavailability of memantine after subtenon, intravenous, and intravitreal administration in rabbits. J Ocul Pharmacol Ther 30:392–399
Pitkanen L, Ranta VP, Moilanen H, Urtti A (2005) Permeability of retinal pigment epithelium: effects of permeant molecular weight and lipophilicity. Invest Ophthalmol Vis Sci 46:641–646
Leblanc B, Jezequel S, Davies T, Hanton G, Taradach C (1998) Binding of drugs to eye melanin is not predictive of ocular toxicity. Regul Toxicol Pharmacol 28:124–132
Harper DR (1989) A comparative study of the microbiological contamination of postmortem blood and vitreous humour samples taken for ethanol determination. Forensic Sci Int 43:37–44
Forrest AR (1993) ACP Broadsheet no 137: April 1993. Obtaining samples at post mortem examination for toxicological and biochemical analyses. J Clin Pathol 46:292–296
Wang L, Zhou W, Ren L, Liu Q, Liu L (2006) Comparative study of two vitreous humor sampling methods in rabbits. J Huazhong Univ Sci Technolog Med Sci 26:475–477
Mulla A, Massey KL, Kalra J (2005) Vitreous humor biochemical constituents: evaluation of between-eye differences. Am J Forensic Med Pathol 26:146–149
Pounder DJ, Carson DO, Johnston K, Orihara Y (1998) Electrolyte concentration differences between left and right vitreous humor samples. J Forensic Sci 43:604–607
Tagliaro F, Bortolotti F, Manetto G, Cittadini F, Pascali VL, Marigo M (2001) Potassium concentration differences in the vitreous humour from the two eyes revisited by microanalysis with capillary electrophoresis. J Chromatogr A 924:493–498
Gagajewski A, Murakami MM, Kloss J, Edstrom M, Hillyer M, Peterson GF, Amatuzio J, Apple FS (2004) Measurement of chemical analytes in vitreous humor: stability and precision studies. J Forensic Sci 49:371–374
Garg U, Althahabi R, Amirahmadi V, Brod M, Blanchard C, Young T (2004) Hyaluronidase as a liquefying agent for chemical analysis of vitreous fluid. J Forensic Sci 49:388–391
Cordeiro C, Seoane R, Camba A, Lendoiro E, Rodriguez-Calvo MS, Vieira DN, Munoz-Barus JI (2015) The application of flow cytometry as a rapid and sensitive screening method to detect contamination of vitreous humor samples and avoid miscalculation of the postmortem interval. J Forensic Sci 60:1346–1349
Bevalot F, Gustin MP, Cartiser N, Le Meur C, Malicier D, Fanton L (2011) Interpretation of drug concentrations in an alternative matrix: the case of meprobamate in vitreous humor. Int J Legal Med 125:463–468
De Letter EA, De Paepe P, Clauwaert KM, Belpaire FM, Lambert WE, Van Bocxlaer JF, Piette MHA (2000) Is vitreous humour useful for the interpretation of 3,4-methylenedioxymethamphetamine (MDMA) blood levels? Experimental approach with rabbits. Int J Legal Med 114:29–35
Logan BK, Stafford DT (1989) Direct analysis of anticonvulsant drugs in vitreous humour by HPLC using a column switching technique. Forensic Sci Int 41:125–134
Rees KA, Seulin S, Yonamine M, Leyton V, Munoz DR, Gianvecchio VA, Pounder DJ, Osselton MD (2013) Analysis of skeletal muscle has potential value in the assessment of cocaine-related deaths. Forensic Sci Int 226:46–53
Ritz S, Harding P, Martz W, Schutz HW, Kaatsch HJ (1992) Measurement of digitalis-glycoside levels in ocular tissues: a way to improve postmortem diagnosis of lethal digitalis-glycoside poisoning? I. Digoxin. Int J Legal Med 105:149–154
Olsen T, Hearn WL (2003) Stability of ethanol in postmortem blood and vitreous humor in long-term refrigerated storage. J Anal Toxicol 27:517–519
Melo P, Bastos ML, Teixeira HM (2012) Benzodiazepine stability in postmortem samples stored at different temperatures. J Anal Toxicol 36:52–60
Rees KA, Jones NS, McLaughlin PA, Seulin S, Leyton V, Yonamine M, Osselton MD (2012) The effect of sodium fluoride preservative and storage temperature on the stability of cocaine in horse blood, sheep vitreous and deer muscle. Forensic Sci Int 217:182–188
Rees KA, Jones NS, McLaughlin PA, Osselton MD (2012) The effect of sodium fluoride preservative and storage temperature on the stability of 6-acetylmorphine in horse blood, sheep vitreous and deer muscle. Forensic Sci Int 217:189–195
Davis LT, Kumar N, Nijm LM, Ulanski LJ 2nd, Tu EY, Fiscella RG, Peterson RJ, Glickman RD (2010) An adaptable HPLC method for the analysis of frequently used antibiotics in ocular samples. J Chromatogr B 878:2421–2426
Logan BK, Stafford DT (1990) High-performance liquid chromatography with column switching for the determination of cocaine and benzoylecgonine concentrations in vitreous humor. J Forensic Sci 35:1303–1309
Cabarcos P, Tabernero MJ, Alvarez I, Lopez P, Fernandez P, Bermejo AM (2010) Analysis of six benzodiazepines in vitreous humor by high-performance liquid chromatography-photodiode-array detection. J Anal Toxicol 34:539–542
Fernandez P, Seoane S, Vazquez C, Tabernero MJ, Carro AM, Lorenzo RA (2013) Chromatographic determination of drugs of abuse in vitreous humor using solid-phase extraction. J Appl Toxicol 33:740–745
Alvear E, von Baer D, Mardones C, Hitschfeld A (2014) Determination of cocaine and its major metabolite benzoylecgonine in several matrices obtained from deceased individuals with presumed drug consumption prior to death. J Forensic Leg Med 23:37–43
Bogusz MJ, Maier RD, Erkens M, Driessen S (1997) Determination of morphine and its 3- and 6-glucuronides, codeine, codeine-glucuronide and 6-monoacetylmorphine in body fluids by liquid chromatography atmospheric pressure chemical ionization mass spectrometry. J Chromatogr B 703:115–127
Sanches LR, Seulin SC, Leyton V, Paranhos BAPB, Pasqualucci CA, Munoz DR, Osselton MD, Yonamine M (2012) Determination of opiates in whole blood and vitreous humor: a study of the matrix effect and an experimental design to optimize conditions for the enzymatic hydrolysis of glucuronides. J Anal Toxicol 36:162–170
Fernandez P, Aldonza M, Bouzas A, Lema M, Bermejo AM, Tabernero MJ (2006) GC-FID determination of cocaine and its metabolites in human bile and vitreous humor. J Appl Toxicol 26:253–257
Bevalot F, Bottinelli C, Cartiser N, Fanton L, Guitton J (2014) Quantification of five compounds with heterogeneous physicochemical properties (morphine, 6-monoacetylmorphine, cyamemazine, meprobamate and caffeine) in 11 fluids and tissues, using automated solid-phase extraction and gas chromatography-tandem mass spectrometry. J Anal Toxicol 38:256–264
Jones GR, Singer PP, Bannach B (2002) Application of LC-MS analysis to a colchicine fatality. J Anal Toxicol 26:365–369
Sporkert F, Augsburger M, Giroud C, Brossard C, Eap CB, Mangin P (2007) Determination and distribution of clotiapine (Entumine) in human plasma, post-mortem blood and tissue samples from clotiapine-treated patients and from autopsy cases. Forensic Sci Int 170:193–199
Cartiser N, Bevalot F, Le Meur C, Gaillard Y, Malicier D, Hubert N, Guitton J (2011) Gas chromatography-tandem mass spectrometry assay for the quantification of four benzodiazepines and citalopram in eleven postmortem rabbit fluids and tissues, with application to animal and human samples. J Chromatogr B 879:2909–2918
Puente B, Hernandez E, Perez S, Pablo L, Prieto E, Garcia MA, Bregante MA (2011) Determination of memantine in plasma and vitreous humour by HPLC with precolumn derivatization and fluorescence detection. J Chromatogr Sci 49:745–752
Dupuis C, Gaulier JM, Pelissier-Alicot AL, Marquet P, Lachatre G (2004) Determination of three beta-blockers in biofluids and solid tissues by liquid chromatography-electrospray-mass spectrometry. J Anal Toxicol 28:674–679
Costa JL, Morrone AR, Resende RR, Chasin AA, Tavares MF (2014) Development of a method for the analysis of drugs of abuse in vitreous humor by capillary electrophoresis with diode array detection (CE-DAD). J Chromatogr B 945–946:84–91
De Paoli G, Brandt SD, Wallach J, Archer RP, Pounder DJ (2013) From the street to the laboratory: analytical profiles of methoxetamine, 3-methoxyeticyclidine and 3-methoxyphencyclidine and their determination in three biological matrices. J Anal Toxicol 37:277–283
Favretto D, Frison G, Maietti S, Ferrara SD (2007) LC-ESI-MS/MS on an ion trap for the determination of LSD, iso-LSD, nor-LSD and 2-oxo-3-hydroxy-LSD in blood, urine and vitreous humor. Int J Legal Med 121:259–265
Fernandez P, Seoane S, Vazquez C, Bermejo AM, Carro AM, Lorenzo RA (2011) A rapid analytical method based on microwave-assisted extraction for the determination of drugs of abuse in vitreous humor. Anal Bioanal Chem 401:2177–2186
Kovatsi L, Rentifis K, Giannakis D, Njau S, Samanidou V (2011) Disposable pipette extraction for gas chromatographic determination of codeine, morphine, and 6-monoacetylmorphine in vitreous humor. J Sep Sci 34:1716–1721
Scott KS, Oliver JS (1999) Vitreous humor as an alternative sample to blood for the supercritical fluid extraction of morphine and 6-monoacetylmorphine. Med Sci Law 39:77–81
Peres MD, Pelicao FS, Caleffi B, De Martinis BS (2014) Simultaneous quantification of cocaine, amphetamines, opiates and cannabinoids in vitreous humor. J Anal Toxicol 38:39–45
Antonides HM, Kiely ER, Marinetti LJ (2007) Vitreous fluid quantification of opiates, cocaine, and benzoylecgonine: comparison of calibration curves in both blood and vitreous matrices with corresponding concentrations in blood. J Anal Toxicol 31:469–476
Duer WC, Spitz DJ, McFarland S (2006) Relationships between concentrations of cocaine and its hydrolysates in peripheral blood, heart blood, vitreous humor and urine. J Forensic Sci 51:421–425
Chronister CW, Walrath JC, Goldberger BA (2001) Rapid detection of benzoylecgonine in vitreous humor by enzyme immunoassay. J Anal Toxicol 25:621–624
Pelander A, Ristimaa J, Ojanpera I (2010) Vitreous humor as an alternative matrix for comprehensive drug screening in postmortem toxicology by liquid chromatography-time-of-flight mass spectrometry. J Anal Toxicol 34:312–318
Arora B, Velpandian T, Saxena R, Lalwani S, Dogra TD, Ghose S (2015) Development and validation of an ESI-LC-MS/MS method for simultaneous identification and quantification of 24 analytes of forensic relevance in vitreous humour, whole blood and plasma. Drug Test Anal. doi:10.1002/dta.1797
Andreasen MF, Telving R, Rosendal I, Eg MB, Hasselstrom JB, Andersen LV (2015) A fatal poisoning involving 25C-NBOMe. Forensic Sci Int 251:e1–e8
Poklis JL, Devers KG, Arbefeville EF, Pearson JM, Houston E, Poklis A (2014) Postmortem detection of 25I-NBOMe [2-(4-iodo-2,5-dimethoxyphenyl)-N-[(2-methoxyphenyl)methyl]ethanamine] in fluids and tissues determined by high performance liquid chromatography with tandem mass spectrometry from a traumatic death. Forensic Sci Int 234:e14–e20
Decaestecker T, De Letter E, Clauwaert K, Bouche MP, Lambert W, Van Bocxlaer J, Piette M, Van den Eeckhout E, Van Peteghem C, De Leenheer A (2001) Fatal 4-MTA intoxication: development of a liquid chromatographic-tandem mass spectrometric assay for multiple matrices. J Anal Toxicol 25:705–710
Seetohul LN, Pounder DJ (2013) Four fatalities involving 5-IT. J Anal Toxicol 37:447–451
Barbera N, Busardo FP, Indorato F, Romano G (2013) The pathogenetic role of adulterants in 5 cases of drug addicts with a fatal outcome. Forensic Sci Int 227:74–76
Tracqui A, Kintz P, Wendling P, Ritter-Lohner S, Mangin P, Jaeger A (1992) Toxicological findings in a fatal case of acebutolol self-poisoning. J Anal Toxicol 16:398–400
Singer PP, Jones GR, Bannach BG, Denmark L (2007) Acute fatal acetaminophen overdose without liver necrosis. J Forensic Sci 52:992–994
Jones GR, Pounder DJ (1987) Site dependence of drug concentrations in postmortem blood—a case study. J Anal Toxicol 11:186–190
Singer PP, Jones GR (1997) Very unusual ethanol distribution in a fatality. J Anal Toxicol 21:506–508
Bicker W, Monticelli F, Bauer A, Roider G, Keller T (2013) Quantification of aconitine in post-mortem specimens by validated liquid chromatography-tandem mass spectrometry method: three case reports on fatal ‘monkshood’ poisoning. Drug Test Anal 5:753–762
Jenkins AJ, Levine B, Locke JL, Smialek JE (1997) A fatality due to alprazolam intoxication. J Anal Toxicol 21:218–220
Tracqui A, Kintz P, Ritter-Lohner S, Mangin P, Lugnier A, Chaumont A (1990) Toxicological findings after fatal amitriptyline self-poisoning. Hum Exp Toxicol 9:257–261
Hurst HE, Jarboe CH (1981) Clinical findings, elimination pharmacokinetics, and tissue drug concentrations following a fatal amitriptyline intoxication. Clin Toxicol 18:119–125
Langford AM, Pounder DJ (1997) Possible markers for postmortem drug redistribution. J Forensic Sci 42:88–92
Pounder DJ, Owen V, Quigley C (1994) Postmortem changes in blood amitriptyline concentration. Am J Forensic Med Pathol 15:224–230
Ziminski KR, Wemyss CT, Bidanset JH, Manning TJ, Lukash L (1984) Comparative study of postmortem barbiturates, methadone, and morphine in vitreous humor, blood, and tissue. J Forensic Sci 29:903–909
Winek CL, Wahba WW, Rozin L (1984) Amoxapine fatalities: three case studies. Forensic Sci Int 26:33–38
Kiely E, Lee CJ, Marinetti L (2009) A fatality from an oral ingestion of methamphetamine. J Anal Toxicol 33:557–560
Logan BK, Weiss EL, Harruff RC (1996) Case report: distribution of methamphetamine in a massive fatal ingestion. J Forensic Sci 41:322–323
Sayama H, Komura H, Kogayu M (2013) Application of hybrid approach based on empirical and physiological concept for predicting pharmacokinetics in humans—usefulness of exponent on prospective evaluation of predictability. Drug Metab Dispos 41:498–507
Quatrehomme G, Ricq O, Lapalus P, Jacomet Y, Ollier A (1992) Acute arsenic intoxication: forensic and toxicologic aspects (an observation). J Forensic Sci 37:1163–1171
Garside D, Ropero-Miller JD, Riemer EC (2006) Postmortem tissue distribution of atomoxetine following fatal and nonfatal doses—three case reports. J Forensic Sci 51:179–182
Brooks JP, Phillips M, Stafford DT, Bell JS (1982) A case of benzphetamine poisoning. Am J Forensic Med Pathol 3:245–247
Palmer RB, Alakija P, de Baca JE, Nolte KB (1999) Fatal brodifacoum rodenticide poisoning: autopsy and toxicologic findings. J Forensic Sci 44:851–855
Yazzie J, Kelly SC, Zumwalt RE, Kerrigan S (2004) Fatal bupivacaine intoxication following unusual erotic practices. J Forensic Sci 49:351–353
Hutchison JD Jr, Harris A, Rasmussen SA, Schlueter SA (1994) Concentrations of butriptyline in biological fluids following a fatal overdose. J Anal Toxicol 18:220–224
Garriott JC, Simmons LM, Poklis A, Mackell MA (1985) Five cases of fatal overdose from caffeine-containing “look-alike” drugs. J Anal Toxicol 9:141–143
Tombolini A, Cingolani M (1996) Fatal accidental ingestion of carbon tetrachloride: a postmortem distribution study. J Forensic Sci 41:166–168
Gerace E, Ciccotelli V, Rapetti P, Salomone A, Vincenti M (2012) Distribution of chloralose in a fatal intoxication. J Anal Toxicol 36:452–456
Wogoman H, Steinberg M, Jenkins AJ (1999) Acute intoxication with guaifenesin, diphenhydramine, and chlorpheniramine. Am J Forensic Med Pathol 20:199–202
Moriya F, Hashimoto Y, Kuo TL (1999) Pitfalls when determining tissue distributions of organophosphorus chemicals: sodium fluoride accelerates chemical degradation. J Anal Toxicol 23:210–215
Anastos N, McIntyre IM, Lynch MJ, Drummer OH (2002) Postmortem concentrations of citalopram. J Forensic Sci 47:882–884
Hargrove V, Molina DK (2009) A fatality due to cyproheptadine and citalopram. J Anal Toxicol 33:564–567
Magdalan J, Zawadzki M, Sloka T, Sozanski T (2013) Suicidal overdose with relapsing clomipramine concentrations due to a large gastric pharmacobezoar. Forensic Sci Int 229:e19–e22
Keller T, Miki A, Binda S, Dirnhofer R (1997) Fatal overdose of clozapine. Forensic Sci Int 86:119–125
Poklis A, Maginn D, Barr JL (1987) Tissue disposition of cocaine in man: a report of five fatal poisonings. Forensic Sci Int 33:83–88
Peretti FJ, Isenschmid DS, Levine B, Caplan YH, Smialek JE (1990) Cocaine fatality: an unexplained blood concentration in a fatal overdose. Forensic Sci Int 48:135–138
Furnari C, Ottaviano V, Sacchetti G, Mancini M (2002) A fatal case of cocaine poisoning in a body packer. J Forensic Sci 47:208–210
Amon CA, Tate LG, Wright RK, Matusiak W (1986) Sudden death due to ingestion of cocaine. J Anal Toxicol 10:217–218
Di Maio VJ, Garriott JC (1978) Four deaths due to intravenous injection of cocaine. Forensic Sci Int 12:119–125
Giroud C, Michaud K, Sporkert F, Eap C, Augsburger M, Cardinal P, Mangin P (2004) A fatal overdose of cocaine associated with coingestion of marijuana, buprenorphine, and fluoxetine. Body fluid and tissue distribution of cocaine and its metabolites determined by hydrophilic interaction chromatography-mass spectrometry (HILIC-MS). J Anal Toxicol 28:464–474
Frost J, Lokken TN, Brede WR, Hegstad S, Nordrum IS, Slordal L (2015) A validated method for simultaneous determination of codeine, codeine-6-glucuronide, norcodeine, morphine, morphine-3-glucuronide and morphine-6-glucuronide in post-mortem blood, vitreous fluid, muscle, fat and brain tissue by LC-MS. J Anal Toxicol 39:203–212
Cheze M, Deveaux M, Pepin G (2006) Liquid chromatography-tandem mass spectrometry for the determination of colchicine in postmortem body fluids. Case report of two fatalities and review of the literature. J Anal Toxicol 30:593–598
Lauer E, Widmer C, Versace F, Staub C, Mangin P, Sabatasso S, Augsburger M, Deglon J (2013) Body fluid and tissue analysis using filter paper sampling support prior to LC-MS/MS: application to fatal overdose with colchicine. Drug Test Anal 5:763–772
Blanco PJ, Rivero AG (2004) First case of illegal euthanasia in Spain: fatal oral potassium cyanide poisoning. Soud Lek 49:30–33
DiMaio VJ, Garriott JC, Putnam R (1975) Digoxin concentrations in postmortem specimens after overdose and therapeutic use. J Forensic Sci 20:340–347
Kaliciak HA, Huckin SN, Cave WS (1992) A death attributed solely to diltiazem. J Anal Toxicol 16:102–103
Baker AM, Johnson DG, Levisky JA, Hearn WL, Moore KA, Levine B, Nelson SJ (2003) Fatal diphenhydramine intoxication in infants. J Forensic Sci 48:425–428
Mozayani A, Schrode P, Carter J, Danielson TJ (2003) A multiple drug fatality involving MK-801 (dizocilpine), a mimic of phencyclidine. Forensic Sci Int 133:113–117
Anderson D, Reed S, Lintemoot J, Kegler S, DeQuintana S, Sandberg M, Muto J (2006) A first look at duloxetine (Cymbalta) in a postmortem laboratory. J Anal Toxicol 30:576–580
Morini L, Pozzi F, Risso E, Vignali C, Groppi A (2012) Distribution of embutramide and mebezonium iodide in a suicide after tanax injection. J Anal Toxicol 36:349–352
Broussard LA, Broussard AK, Pittman TS, Lirette DK (2000) Death due to inhalation of ethyl chloride. J Forensic Sci 45:223–225
Yacoub I, Robinson CA, Simmons GT, Hall M (1993) Death attributed to ethyl chloride. J Anal Toxicol 17:384–385
Morano RA, Spies C, Walker FB, Plank SM (1993) Fatal intoxication involving etryptamine. J Forensic Sci 38:721–725
Molina DK, Hargrove VM, Rodriguez RG (2008) Distribution of etomidate in a fatal intoxication. J Anal Toxicol 32:715–718
Anderson DT, Muto JJ (2000) Duragesic transdermal patch: postmortem tissue distribution of fentanyl in 25 cases. J Anal Toxicol 24:627–634
Coopman V, Cordonnier J, Pien K, Van Varenbergh D (2007) LC-MS/MS analysis of fentanyl and norfentanyl in a fatality due to application of multiple Durogesic transdermal therapeutic systems. Forensic Sci Int 169:223–227
Wiesbrock UO, Rochholz G, Franzelius C, Schwark T, Grellner W (2008) Excessive use of fentanyl patches as the only means of suicide. Arch Kriminol 222:23–30
Levine B, Chute D, Caplan YH (1990) Flecainide intoxication. J Anal Toxicol 14:335–336
Martinez MA, Ballesteros S, Piga FJ, Sanchez de la Torre C, Cubero CA (2007) The tissue distribution of fluoride in a fatal case of self-poisoning. J Anal Toxicol 31:526–533
Johnson RD, Lewis RJ, Angier MK (2007) The distribution of fluoxetine in human fluids and tissues. J Anal Toxicol 31:409–414
McIntyre IM, Syrjanen ML, Lawrence KL, Dow CA, Drummer OH (1994) A fatality due to flurazepam. J Forensic Sci 39:1571–1574
Kunsman GW, Rodriguez R, Rodriguez P (1999) Fluvoxamine distribution in postmortem cases. Am J Forensic Med Pathol 20:78–83
Ferrara SD, Tedeschi L, Frison G, Rossi A (1995) Fatality due to gamma-hydroxybutyric acid (GHB) and heroin intoxication. J Forensic Sci 40:501–504
Kintz P, Villain M, Pelissier AL, Cirimele V, Leonetti G (2005) Unusually high concentrations in a fatal GHB case. J Anal Toxicol 29:582–585
Mazarr-Proo S, Kerrigan S (2005) Distribution of GHB in tissues and fluids following a fatal overdose. J Anal Toxicol 29:398–400
Meatherall R, Lee C, Phillips S (2011) Accidental death from hydromorphone ingestion. J Forensic Sci 56(Suppl 1):S271–S274
Thevis M, Thomas A, Schanzer W, Ostman P, Ojanpera I (2012) Measuring insulin in human vitreous humour using LC-MS/MS. Drug Test Anal 4:53–56
Palmiere C, Sabatasso S, Torrent C, Rey F, Werner D, Bardy D (2015) Post-mortem determination of insulin using chemiluminescence enzyme immunoassay: preliminary results. Drug Test Anal 7:797–803
Hess C, Madea B, Daldrup T, Musshoff F (2013) Determination of hypoglycaemia induced by insulin or its synthetic analogues post mortem. Drug Test Anal 5:802–807
Nowicka J, Skowronek R, Czech E, Kulikowska J, Olszowy Z (2013) Comments on ‘Measuring insulin in human vitreous humour using LC-MS, MS’ by Thevis et al. Drug Test Anal 5:133–134
Winston DC (2000) Suicide via insulin overdose in nondiabetics: the New Mexico experience. Am J Forensic Med Pathol 21:237–240
Pricone MG, King CV, Drummer OH, Opeskin K, McIntyre IM (2000) Postmortem investigation of lamotrigine concentrations. J Forensic Sci 45:11–15
Yonemitsu K, Pounder DJ (1993) Postmortem changes in blood tranylcypromine concentration: competing redistribution and degradation effects. Forensic Sci Int 59:177–184
Mazzola CD, Miron S, Jenkins AJ (2000) Loxapine intoxication: case report and literature review. J Anal Toxicol 24:638–641
Gaillard YP, Cuquel AC, Boucher A, Romeuf L, Bevalot F, Prevosto JM, Menard JM (2013) A fatality following ingestion of the designer drug meta-chlorophenylpiperazine (mCPP) in an asthmatic–HPLC-MS/MS detection in biofluids and hair. J Forensic Sci 58:263–269
De Letter EA, Clauwaert KM, Lambert WE, Van Bocxlaer JF, De Leenheer AP, Piette MH (2002) Distribution study of 3,4-methylenedioxymethamphetamine and 3,4-methylenedioxyamphetamine in a fatal overdose. J Anal Toxicol 26:113–118
Moore KA, Mozayani A, Fierro MF, Poklis A (1996) Distribution of 3,4-methylenedioxymethamphetamine (MDMA) and 3,4-methylenedioxyamphetamine (MDA) stereoisomers in a fatal poisoning. Forensic Sci Int 83:111–119
De Letter EA, Bouche MP, Van Bocxlaer JF, Lambert WE, Piette MH (2004) Interpretation of a 3,4-methylenedioxymethamphetamine (MDMA) blood level: discussion by means of a distribution study in two fatalities. Forensic Sci Int 141:85–90
Adamowicz P, Tokarczyk B, Stanaszek R, Slopianka M (2013) Fatal mephedrone intoxication—a case report. J Anal Toxicol 37:37–42
Henry JL, Epley J, Rohrig TP (2003) The analysis and distribution of mescaline in postmortem tissues. J Anal Toxicol 27:381–382
Couper FJ, Chopra K, Pierre-Louis ML (2005) Fatal methadone intoxication in an infant. Forensic Sci Int 153:71–73
Wu Chen NB, Donoghue ER, Schaffer MI (1985) Methanol intoxication: distribution in postmortem tissues and fluids including vitreous humor. J Forensic Sci 30:213–216
Beno JM, Hartman R, Wallace C, Nemeth D, LaPoint S (2011) Homicidal methanol poisoning in a child. J Anal Toxicol 35:524–528
Pla A, Hernandez AF, Gil F, Garcia-Alonso M, Villanueva E (1991) A fatal case of oral ingestion of methanol. Distribution in postmortem tissues and fluids including pericardial fluid and vitreous humor. Forensic Sci Int 49:193–196
Andresen H, Schmoldt H, Matschke J, Flachskampf FA, Turk EE (2008) Fatal methanol intoxication with different survival times–morphological findings and postmortem methanol distribution. Forensic Sci Int 179:206–210
Avella J, Briglia E, Harleman G, Lehrer M (2005) Percutaneous absorption and distribution of methanol in a homicide. J Anal Toxicol 29:734–737
Akhgari M, Panahianpour MH, Bazmi E, Etemadi-Aleagha A, Mahdavi A, Nazari SH (2013) Fatal methanol poisoning: features of liver histopathology. Toxicol Ind Health 29:136–141
Hantson P, Haufroid V, Mahieu P (2000) Determination of formic acid tissue and fluid concentrations in three fatalities due to methanol poisoning. Am J Forensic Med Pathol 21:335–338
Moriya F, Hashimoto Y (2005) A fatal poisoning caused by methomyl and nicotine. Forensic Sci Int 149:167–170
McIntyre IM, Hamm CE, Aldridge L, Nelson CL (2013) Acute methylone intoxication in an accidental drowning–a case report. Forensic Sci Int 231:e1–e3
Cantrell FL, Ogera P, Mallett P, McIntyre IM (2014) Fatal oral methylphenidate intoxication with postmortem concentrations. J Forensic Sci 59:847–849
Rohrig TP, Rundle DA, Leifer WN (1987) Fatality resulting from metoprolol overdose. J Anal Toxicol 11:231–232
Stajic M, Granger RH, Beyer JC (1984) Fatal metoprolol overdose. J Anal Toxicol 8:228–230
Kempton J, Manoukian A, Levine B, Smialek J (1994) A mexiletine intoxication. J Anal Toxicol 18:346–347
Rohrig TP, Harty LE (1994) Postmortem distribution of mexiletine in a fatal overdose. J Anal Toxicol 18:354–356
Anderson DT, Fritz KL, Muto JJ (1999) Distribution of mirtazapine (Remeron) in thirteen postmortem cases. J Anal Toxicol 23:544–548
Kirkton C, McIntyre IM (2006) Therapeutic and toxic concentrations of mirtazapine. J Anal Toxicol 30:687–691
McIntyre IM, Trochta A, Stolberg S, Campman SC (2015) Mitragynine ‘Kratom’ related fatality: a case report with postmortem concentrations. J Anal Toxicol 39:152–155
McIntyre IM, King CV, Staikos V, Gall J, Drummer OH (1997) A fatality involving moclobemide, sertraline, and pimozide. J Forensic Sci 42:951–953
Winek CL, Wahba WW, Rozin L (1999) Heroin fatality due to penile injection. Am J Forensic Med Pathol 20:90–92
Gock SB, Wong SH, Stormo KA, Jentzen JM (1999) Self-intoxication with morphine obtained from an infusion pump. J Anal Toxicol 23:130–133
Kintz P, Mangin P, Lugnier AA, Chaumont AJ (1989) Toxicological data after heroin overdose. Hum Toxicol 8:487–489
Wetli CV, Rao A, Rao VJ (1997) Fatal heroin body packing. Am J Forensic Med Pathol 18:312–318
Seetohul LN, De Paoli G, Drummond G, Maskell PD (2015) Nefopam hydrochloride: a fatal overdose. J Anal Toxicol 39:486–489
Kemp PM, Sneed GS, George CE, Distefano RF (1997) Postmortem distribution of nicotine and cotinine from a case involving the simultaneous administration of multiple nicotine transdermal systems. J Anal Toxicol 21:310–313
Jenkins AJ, Sarconi KM, Raaf HN (1998) Determination of olanzapine in a postmortem case. J Anal Toxicol 22:605–609
Linnet K, Steentoft A, Simonsen KW, Sabers A, Hansen SH (2008) An oxcarbazepine-related fatality with an overview of 26 oxcarbazepine postmortem cases. Forensic Sci Int 177:248–251
Anderson DT, Fritz KL, Muto JJ (2002) Oxycontin: the concept of a “ghost pill” and the postmortem tissue distribution of oxycodone in 36 cases. J Anal Toxicol 26:448–459
Al-Asmari AI, Anderson RA, Cooper GA (2009) Oxycodone-related fatalities in the west of Scotland. J Anal Toxicol 33:423–432
Romain N, Giroud C, Michaud K, Mangin P (2003) Suicide by injection of a veterinarian barbiturate euthanasia agent: report of a case and toxicological analysis. Forensic Sci Int 131:103–107
Crichton ML, Shenton CF, Drummond G, Beer LJ, Seetohul LN, Maskell PD (2015) Analysis of phenazepam and 3-hydroxyphenazepam in post-mortem fluids and tissues. Drug Test Anal. doi:10.1002/dta.1790
Sturner WQ, Spruill FG, Garriott JC (1974) Two propylhexedrine-associated fatalities: benzedrine revisited. J Forensic Sci 19:572–574
Flammia DD, Valouch T, Venuti S (2006) Tissue distribution of quetiapine in 20 cases in Virginia. J Anal Toxicol 30:287–292
Langman LJ, Kaliciak HA, Carlyle S (2004) Fatal overdoses associated with quetiapine. J Anal Toxicol 28:520–525
Hopenwasser J, Mozayani A, Danielson TJ, Harbin J, Narula HS, Posey DH, Shrode PW, Wilson SK, Li R, Sanchez LA (2004) Postmortem distribution of the novel antipsychotic drug quetiapine. J Anal Toxicol 28:264–267
Coopman V, De Leeuw M, Cordonnier J, Jacobs W (2009) Suicidal death after injection of a castor bean extract (Ricinus communis L.). Forensic Sci Int 189:e13–e20
Duband S, Bidat C, Gaillard Y, Rochet M, Camdessanche JP, Peoc’h M (2012) A fatal intoxication case involving ropinirole. J Forensic Leg Med 19:422–425
Lewis RJ, Angier MK, Williamson KS, Johnson RD (2013) Analysis of sertraline in postmortem fluids and tissues in 11 aviation accident victims. J Anal Toxicol 37:208–216
Lindsey T, O’Hara J, Irvine R, Kerrigan S (2004) Strychnine overdose following ingestion of gopher bait. J Anal Toxicol 28:135–137
Ferslew KE, Hagardorn AN, McCormick WF (1989) Postmortem determination of the biological distribution of sufentanil and midazolam after an acute intoxication. J Forensic Sci 34:249–257
Lin DL, Lin RL (2005) Distribution of 11-nor-9-carboxy-Δ9-tetrahydrocannabinol in traffic fatality cases. J Anal Toxicol 29:58–61
Mozayani A, Carter J, Nix R (1999) Distribution of topiramate in a medical examiner’s case. J Anal Toxicol 23:556–558
Moriya F, Hashimoto Y (2003) A case of fatal triazolam overdose. Leg Med 5(Suppl 1):S91–S95
Bayley M, Walsh FM, Valske MJ (1975) Report of a fatal, acute tripelennamine intoxication. J Forensic Sci 20:539–543
Poklis A, Poklis JL, Trautman D, Treece C, Backer R, Harvey CM (1998) Disposition of valproic acid in a case of fatal intoxication. J Anal Toxicol 22:537–540
Stove CP, De Letter EA, Piette MH, Lambert WE (2013) Fatality following a suicidal overdose with varenicline. Int J Legal Med 127:85–91
Kunsman GW, Kunsman CM, Presses CL, Garavaglia JC, Farley NJ (2000) A mixed-drug intoxication involving venlafaxine and verapamil. J Forensic Sci 45:926–928
Goeringer KE, McIntyre IM, Drummer OH (2001) Postmortem tissue concentrations of venlafaxine. Forensic Sci Int 121:70–75
Parsons AT, Anthony RM, Meeker JE (1996) Two fatal cases of venlafaxine poisoning. J Anal Toxicol 20:266–268
Jaffe PD, Batziris HP, van der Hoeven P, DeSilva D, McIntyre IM (1999) A study involving venlafaxine overdoses: comparison of fatal and therapeutic concentrations in postmortem specimens. J Forensic Sci 44:193–196
Kintz P, Tracqui A, Potard D, Petit G, Mangin P (1994) An unusual death by zipeprol overdose. Forensic Sci Int 64:159–163
Gock SB, Wong SH, Nuwayhid N, Venuti SE, Kelley PD, Teggatz JR, Jentzen JM (1999) Acute zolpidem overdose—report of two cases. J Anal Toxicol 23:559–562
Meatherall RC (1997) Zopiclone fatality in a hospitalized patient. J Forensic Sci 42:340–343
Pragst F, Spiegel K, Leuschner U, Hager A (1999) Detection of 6-acetylmorphine in vitreous humor and cerebrospinal fluid—comparison with urinary analysis for proving heroin administration in opiate fatalities. J Anal Toxicol 23:168–172
Wyman J, Bultman S (2004) Postmortem distribution of heroin metabolites in femoral blood, liver, cerebrospinal fluid, and vitreous humor. J Anal Toxicol 28:260–263
Rees KA, Pounder DJ, Osselton MD (2013) Distribution of opiates in femoral blood and vitreous humour in heroin/morphine-related deaths. Forensic Sci Int 226:152–159
Behar-Cohen FF, Gauthier S, El Aouni A, Chapon P, Parel JM, Renard G, Chauvaud D (2001) Methylprednisolone concentrations in the vitreous and the serum after pulse therapy. Retina 21:48–53
Mains J, Tan LE, Zhang T, Young L, Shi R, Wilson C (2012) Species variation in small molecule components of animal vitreous. Invest Ophthalmol Vis Sci 53:4778–4786
Salmon AY, Goren Z, Avissar Y, Soreq H (1999) Human erythrocyte but not brain acetylcholinesterase hydrolyses heroin to morphine. Clin Exp Pharmacol Physiol 26:596–600
Konstantinova SV, Normann PT, Arnestad M, Karinen R, Christophersen AS, Morland J (2012) Morphine to codeine concentration ratio in blood and urine as a marker of illicit heroin use in forensic autopsy samples. Forensic Sci Int 217:216–221
Lin DL, Chen CY, Shaw KP, Havier R, Lin RL (1997) Distribution of codeine, morphine, and 6-acetylmorphine in vitreous humor. J Anal Toxicol 21:258–261
Robertson MD, Drummer OH (1998) Postmortem distribution and redistribution of nitrobenzodiazepines in man. J Forensic Sci 43:9–13
Scott KS, Oliver JS (2001) The use of vitreous humor as an alternative to whole blood for the analysis of benzodiazepines. J Forensic Sci 46:694–697
Devgun MS, Dunbar JA (1986) Biochemical investigation of vitreus: applications in forensic medicine, especially in relation to alcohol. Forensic Sci Int 31:27–34
Mackey-Bojack S, Kloss J, Apple F (2000) Cocaine, cocaine metabolite, and ethanol concentrations in postmortem blood and vitreous humor. J Anal Toxicol 24:59–65
Jenkins AJ, Oblock J (2008) Phencyclidine and cannabinoids in vitreous humor. Leg Med 10:201–203
Cox D, Jufer Phipps RA, Levine B, Jacobs A, Fowler D (2007) Distribution of phencyclidine into vitreous humor. J Anal Toxicol 31:537–539
Knittel JL, Clay DJ, Bailey KM, Gebhardt MA, Kraner JC (2009) Comparison of oxycodone in vitreous humor and blood using EMIT screening and gas chromatographic-mass spectrometric quantitation. J Anal Toxicol 33:433–438
Brunet B, Mura P (2012) L’humeur vitrée en toxicologie médico-légale: revue de la littérature et applications. Ann Toxicol Anal 24:9–15
Carvalho VM, Fukushima AR, Fontes LR, Fuzinato DV, Florio JC, Chasin AA (2013) Cocaine postmortem distribution in three brain structures: a comparison with whole blood and vitreous humour. J Forensic Leg Med 20:143–145
Bevalot F, Fanton L, Maujean G, Cartiser N, Chillon A, Mounier C, Prevosto J, Le Meur C, Malicier D (2008) Correlation study between blood concentrations and vitreous concentrations; case of meprobamate and some benzodiazepines (bromazepam, nordazepam, oxazepam). Proceedings of the 60th Anniversary Scientific Meeting of American Academy of Forensic Sciences, Washington, DC, p 418
Andresen-Streichert H, Jensen P, Kietzerow J, Schrot M, Wilke N, Vettorazzi E, Mueller A, Iversen-Bergmann S (2014) Endogenous gamma-hydroxybutyric acid (GHB) concentrations in post-mortem specimens and further recommendation for interpretative cut-offs. Int J Legal Med 129:57–68
Marinetti LJ, Isenschmid DS, Hepler BR, Kanluen S (2005) Analysis of GHB and 4-methyl-GHB in postmortem matrices after long-term storage. J Anal Toxicol 29:41–47
Kintz P, Villain M, Cirimele V, Ludes B (2004) GHB in postmortem toxicology. Discrimination between endogenous production from exposure using multiple specimens. Forensic Sci Int 143:177–181
Moriya F, Hashimoto Y (2005) Site-dependent production of gamma-hydroxybutyric acid in the early postmortem period. Forensic Sci Int 148:139–142
Castro AL, Dias M, Reis F, Teixeira HM (2014) Gamma-hydroxybutyric acid endogenous production and post-mortem behaviour—the importance of different biological matrices, cut-off reference values, sample collection and storage conditions. J Forensic Leg Med 27C:17–24
Wunder C, Kauert GF, Toennes SW (2014) Factors leading to the degradation/loss of insulin in postmortem blood samples. Forensic Sci Int 241:173–177
Ojanpera I, Sajantila A, Vinogradova L, Thomas A, Schanzer W, Thevis M (2013) Post-mortem vitreous humour as potential specimen for detection of insulin analogues by LC-MS/MS. Forensic Sci Int 233:328–332
Parker DR, McIntyre IM (2005) Case studies of postmortem quetiapine: therapeutic or toxic concentrations? J Anal Toxicol 29:407–412
Teixeira HM, Reis F, Proenca P, Ramos P, Quintela O, Lopez-Rivadulla M, Marques E, Vieira DN (2004) Vitreous humour as a complementary sample to blood for the detection/confirmation of diazepam: ante-mortem and post-mortem studies in an animal model. Hum Exp Toxicol 23:571–577
Ritz S, Harding P, Martz W, Schutz HW, Kaatsch HJ (1992) Measurement of digitalis-glycoside levels in ocular tissues: a way to improve postmortem diagnosis of lethal digitalis-glycoside poisoning? II. Digitoxin. Int J Legal Med 105:155–159
McKinney PE, Phillips S, Gomez HF, Brent J, MacIntyre M, Watson WA (1995) Vitreous humor cocaine and metabolite concentrations: do postmortem specimens reflect blood levels at the time of death? J Forensic Sci 40:102–107
Maskell PD, Albeishy M, De Paoli G, Wilson NE, Seetohul LN (2015) Postmortem redistribution of the heroin metabolites morphine and morphine-3-glucuronide in rabbits over 24 h. Int J Legal Med. doi:10.1007/s00414-015-1185-3
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
There are no financial or other relationships that could lead to a conflict of interest.
Ethical approval
This article does not contain any studies with human participants or animals performed by any of the authors.
Additional information
F. Bévalot and N. Cartiser contributed equally to this work.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Bévalot, F., Cartiser, N., Bottinelli, C. et al. Vitreous humor analysis for the detection of xenobiotics in forensic toxicology: a review. Forensic Toxicol 34, 12–40 (2016). https://doi.org/10.1007/s11419-015-0294-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11419-015-0294-5