
Abstract
Ca2+-Calmodulin binding to the variable N-terminal region of the diacylglycerol/phorbol ester-binding UNC13/Munc13 family of proteins modulates the short-term synaptic plasticity characteristics in neurons. Here, we report the sequential backbone and side chain resonance assignment of the Ca2+-Calmodulin/Munc13-1458–492 peptide complex at pH 6.8 and 35°C (BMRB No. 15470).
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Biological context
UNC13/Munc13 family of proteins are diacylglycerol/phorbol ester-binding proteins specifically localized to presynaptic active zones, where the synaptic vesicles dock and release their neurotransmitters to the intersynaptic cleft for the chemical signal transmission among neurons (Junge et al. 2004; Brose et al. 1995). In Caenorhabditis elegans, unc-13 is essential for coordinated movement (Brenner 1974). At a molecular level, UNC13/Munc13 proteins are essential in the maturation process of synaptic vesicles, remodeling the SNARE complex of proteins leading to fusion competent molecular states. Upon deletion of UNC13/Munc13 proteins neurotransmitter release is completely impaired (Richmond et al. 2001).
Ca2+-Calmodulin (CaM) is a highly conserved ubiquitous eukaryotic signaling protein, regulating the activity of numerous proteins like protein kinases, phosphodiesterases, ion channels and pumps, and nitric-oxide synthases. Various studies have shown that CaM activates these proteins interacting with regulatory elements possessing an amphiphilic α-helix character (Crivici and Ikura 1995).
It has been shown (Junge et al. 2004) that some of the Munc13 protein isoforms contain a highly conserved CaM binding motif in their variable N-terminal region. However, this binding motif does not belong to any of the classical CaM binding motifs previously described (Rhoads and Friedberg 1997). Synthetic peptides of the Munc13-1 and ubMunc13-2 isoforms belonging to this CaM binding domain form high-affinity complexes with CaM in vitro. To unravel the CaM binding mechanism to Munc13 proteins, we carried out NMR experiments on the CaM/Munc13-1458–492 peptide complex.
Methods and experiments
NMR sample preparation
The costs of 15N, 13C-uniformly labeled synthetic peptides are prohibitive. Therefore, we chose a recombinant approach to obtain the Munc13-1458–492 peptide and will be described elsewhere. Briefly, the CaM binding domain (458–492) coding sequence of munc13-1 was cloned into the pGEX-2T expression vector (GE Healthcare). The uniformly labeled (15N, 13C) GST-Munc13-1458–492 fusion protein was over-expressed in M9 minimal medium containing 15NH4Cl and 13C-glucose as solely source of nitrogen and carbon, respectively. Due to solubility problems, the GST-Munc13-1458–492 fusion protein was co-expressed with CaM using resistance to two different antibiotics. The GST-Munc13-1458–492/CaM complex was co-purified via affinity chromatography using GST-Sepharose (GE-Healthcare) according to the vendor instructions. The GST fusion was cleaved with thrombin and the Munc13-1 peptide purified by reverse-phase HPLC. The eluted peptide was lyophilized, reconstituted with 250 μl of the NMR-sample aqueous buffer (20 mM Bis–Tris, 150 mM KCl, 10 mM CaCl2, pH 6.8) and mixed with a 1.2 M excess of unlabeled CaM dissolved in (Millipore) water to give a total final volume of 2 ml. The CaM/Munc13-1459–492 complex was concentrated by vacuum centrifugation in a Speedvac to a final volume of 250 μl giving a final concentration of approximately 0.5 mM. In order to get a lock signal in the NMR spectrometer, we added 10 μl of 99% D2O.
The (U-15N, 13C) labeled CaM was obtained as described (Guerini et al. 1984; Haberz et al. 2006). Briefly, CaM was over-expressed from a pET28a-CaM construct in E. coli BL21(DE3) in labeled M9 minimal medium. The cells were lysed by ultrasound sonication and the cell debris removed by centrifugation at 15,000 g for 45 min. The supernatant was precipitated with 2.84% trichloroacetic acid and subjected to hydrophobic interaction chromatography on a phenylsepharose column; CaM was eluted with an EGTA containing buffer. To prepare the complex sample, 8 mg of lyophilized (U-15N, 13C) labeled CaM were weighted and dissolved in 2 ml of (Millipore) water. Around 2.4 mg of a synthetic natural abundance Munc13-1459–492 peptide (1.2 equivalents) were weighted and dissolved in 250 μl of the NMR-sample aqueous buffer (20 mM Bis–Tris, 150 mM KCl, 10 mM CaCl2, pH 6.8) and added to the CaM solution; the mixture was concentrated by vacuum centrifugation in a Speedvac to give a final volume of 250 μl. 10 μl of 99% D2O were added for the NMR-lock signal. An additional (U-15N, 13C) labeled CaM/Munc13-1459–492 complex sample dissolved in 99% D2O was prepared in the same manner. The three samples were placed in 280 μl Shigemi microcells covered with their glass plunger and sealed with Parafilm.
NMR experiments
All NMR experiments were carried out at 35°C on the following Bruker spectrometers: Avance 600 MHz equipped with a z-gradient cryogenic HCN probe head; DRX 600 MHz with a z-gradient HCN probe head; Avance 700 MHz with a triple-axis gradient HCN probe head; DMX 800 MHz with a z-gradient cryogenic HCN probe head; and Avance 900 MHz with a z-gradient cryogenic HCN probe head.
The experiments made use of pulse field gradient coherence selection. Quadrature detection in the indirect dimensions was achieved either with the States-TPPI or Echo/antiecho methods. The chemical shift referencing was achieved internally by the addition of 0.4 mM DSS. The acquisition parameters for the experiments are given in Table 1. The spectra were processed with XWIN-NMR, Felix (Accelerys, Inc.), NMR-PIPE (Delaglio et al. 1995) and analyzed with SPARKY (Goddard and Kneller 1999).
Assignments and data deposition
The sequential backbone and side chain resonance assignment is nearly complete and had been deposited in the Biological Magnetic Resonance Data Bank (http://www.bmrb.wisc.edu) under the accession number 15,470. The 1H-15N-HSQC spectrum of the (U-15N, 13C) CaM/Munc13-1459–492 complex sample is shown in Fig. 1. Its general appearance is consistent with that one of Ca2+-CaM, however, many NH cross-peaks in the N-terminal domain (1–76) of CaM had large line-widths mainly in helices A and D. This gave rise to an assignment completeness of 95.7% for the backbone resonances (CA, CB, C, N, H, HA and HB). In contrast, the assignment of the C-terminal domain (83–148) of the Ca2+-CaM/peptide complex was nearly complete (98.8% Backbone resonances). This suggested that the N-terminal domain of Ca2+-CaM experiences conformational exchange, whereas the C-terminal domain consists of a single conformer under these conditions. In Fig. 2, the 1H-15N-HSQC spectrum of the (U-15N, 13C) Munc13-1458–492/CaM complex is shown. An overlay of this spectrum with that one of a (U-15N, 13C)-Munc13-1458–492 peptide in 8 M urea solution (supplementary material) clearly revealed an increase in the amide resonance dispersion of the peptide, characteristic of secondary structure formation in this amphiphilic α-helix motif upon complex formation with Ca2+-CaM.

1H-15N-HSQC spectrum of the (U-15N, 13C) CaM/Munc13-1459–492 complex measured at 35°C at 900 MHz. The highly overlapped central region is enlarged on the right side

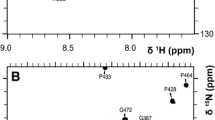
1H-15N-HSQC spectrum of the (U-15N, 13C) Munc13-1458–492/CaM complex recorded at 35°C at 600 MHz
The assignment of the labeled Munc13-1458–492 peptide in complex with Ca2+-CaM was completed to 99.2% of the backbone resonances. The chemical shifts of the ε-methyl protons in the methionines of Ca2+-CaM in complex with the Munc13-1458–492 peptide were compared with those of free Ca2+-CaM. Most of the ε-methyl protons of the eight methionines in Ca2+-CaM were shifted upfield upon binding to the peptide. Significant changes (>0.15 ppm) were found for M51 (−0.3 ppm), M71 (−0.31 ppm), M109 (−0.17 ppm), M124 (−1.17 ppm), M144 (−0.3 ppm) and M145 (−0.65 ppm). The effect on the chemical shift for the methionines is attributed to deshielding effects caused by the aromatic rings of the various aromatic amino acids present in the peptide complexed to Ca2+-CaM.
References
Brenner S (1974) The genetics of Caenorhabditis elegans. Genetics 77:71–94
Brose N, Hofmann K, Hata Y, Südhof TC (1995) Mammalian homologues of Caenorhabditis elegans unc-13 gene define novel family of C2-domain proteins. J Biol Chem 270:25273–25280
Crivici A, Ikura M (1995) Molecular and structural basis of target recognition by calmodulin. Annu Rev Biophys Biomol Struct 24:85–116
Delaglio F, Grzesiek S, Vuister GW, Zhu G, Pfeifer J, Bax A (1995) NMRPipe: a multidimensional spectral processing system based on UNIX pipes. J Biomol NMR 6:277–293
Goddard TD, Kneller DG (1999) SPARKY 3. University of California, San Francisco
Guerini D, Krebs J, Carafoli E (1984) Stimulation of the purified erythrocyte Ca2+-ATPase by tryptic fragments of calmodulin. J Biol Chem 259:15172–15177
Haberz P, Rodríguez-Castañeda F, Junker J, Becker S, Leonov A, Griesinger C (2006) Two new chiral EDTA-based metal chelates for weak alignment of proteins in solution. Org Lett 8:1275–1278
Junge HJ, Rhee J-S, Jahn O, Varoqueaux F, Spiess J, Waxham MN, Rosenmund C, Brose N (2004) Calmodulin and Munc13 form a Ca2+ sensor/effector complex that controls short-term synaptic plasticity. Cell 118:389–401
Rhoads AR, Friedberg F (1997) Sequence motifs for calmodulin recognition. FASEB J 11:331–340
Richmond JE, Weimer RM, Jorgensen EM (2001) An open form of syntaxin bypases the requirement for unc-13 in vesicle priming. Nature 412:338–341
Acknowledgments
The authors would like to thank Dr. Pierre Montaville for useful discussions in the protein preparation part; Volker Klaukien, Kerstin Overkamp and Gerhard Wolff for peptide synthesis and HPLC purification; and Dr. Vinesh Vijayan and Dr. Christophe Fàres for assistance in the NMR-measurements. This work has been financed by the Max-Planck Society.
Ethical standards
The experiments comply with the german current laws for ethical standards.
Conflict of interests
The authors declare that they have no conflict of interest.
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License (https://creativecommons.org/licenses/by-nc/2.0), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Rodríguez-Castañeda, F., Coudevylle, N., Becker, S. et al. 1H, 13C and 15N resonance assignments of the Calmodulin-Munc13-1 peptide complex. Biomol NMR Assign 4, 45–48 (2010). https://doi.org/10.1007/s12104-009-9204-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12104-009-9204-2