
Abstract
Unusually high losses of honey bee colonies are reported in many regions of the world, but little data is available concerning the status of honey bee stocks in Africa. However, the situation on this continent, where beekeeping is weakly developed and where the wild population of the pollinator remains large, can give us an insight on the causes of increased mortalities elsewhere. In this study, we evaluate the health status of Apis mellifera intermissa and A. m. sahariensis populations inhabiting the north and the south of Algeria, respectively. We report few colony losses associated with an atypical pattern of prevalence for common honey bee parasites and pathogens. The presence or absence of these risk factors is discussed in relation to the occurrence of local and global colony losses to contribute to our understanding of how honey bee pathogens and parasite impact this pollinator’s health.
Similar content being viewed by others


1 Introduction
Honey bees represent a well-studied group of social insects with both ecological and agricultural importance (Winston 1987; Breeze et al. 2014). It was estimated that one third of the food crop benefits for the service of pollinators, of which honey bees represent a large proportion (Delaplane and Mayer 2000; Gallai et al. 2008; Aizen and Harder 2009). Worldwide elevated mortality of honey bee colonies is therefore a worrying problem for beekeeping and agriculture, but also for ecosystem functioning. Although several factors may play a role in these declines (vanEngelsdorp et al. 2010; Cornman et al. 2012), most mortalities have been attributed to high loads of parasites and pathogens, such as high infestations by the ectoparasitic mite Varroa destructor, together with associated viruses (Cox-Foster et al. 2007; Berthoud et al. 2010; Dainat et al. 2012a, b; Dietemann et al. 2012; Nazzi et al. 2012; Francis et al. 2013). The microsporidian Nosema ceranae has also been incriminated in colony mortality in southern Europe (Higes et al. 2008). The detection, biology, epidemiology of various parasites and pathogens and their relationships with mortality of bees are therefore studied in Europe, USA and Asia (e.g. Tentcheva et al. 2004; Chen and Siede 2007; Genersch and Aubert 2010; Cornman et al. 2012; Li et al. 2012; Francis et al. 2013; Yang et al. 2013). However little data on honeybee pathogens and health status is available from Africa (Dietemann et al. 2009; Adjlane et al. 2012a, b, c; Mumoki et al. 2014; Amakpe et al. 2015). Defining the relationship between honeybees and their pests and pathogens in various contexts, including those in which honey bee colonies are not declining, improves our understanding, to date still patchy, of the causes of the honey bee colony losses occurring globally. To contribute to the filling of this gap, we investigated the health status and pathogens occurring in the two honey bee subspecies endemic to Algeria, Apis mellifera intermissa and A.m. sahariensis. A. m. sahariensis predominates in the south of the country due to its adaptation to drought conditions, whereas A. m. intermissa occurs in the North. The presence of N. ceranae, SBV and DWV has already been reported (Matheson 1993; Higes et al. 2009a; Loucif-Ayad et al. 2013) for some localities in north and west Algeria in A. m. intermissa. To expand on these results, we measured the prevalence and infestation rates of the parasitic mite V. destructor, as well as the prevalence of deformed wing virus (DWV), Varroa destructor virus (VDV-1), sacbrood virus (SBV), black queen cell virus (BQCV), acute bee paralysis virus (ABPV), Kashmir bee virus (KBV), Israeli acute paralysis virus (IAPV), chronic bee paralysis virus (CBPV), as well as of N. apis and N. ceranae in the major beekeeping districts. In addition, we determined the percentage of colony losses experienced by beekeepers with a questionnaire (van der Zee et al. 2012) and investigated the relation of parasite or pathogen occurrence to these losses. We compare the patterns observed in Algeria to the reports from other countries in Africa and worldwide in order to contribute to a more comprehensive understanding of the role of pathogens in honey bee health.
2 Materials and methods
2.1 Study areas
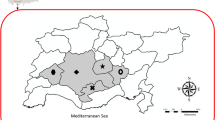
A. m. intermissa inhabits the north of Algeria from the coast, over the Tellian Atlas to the Saharian Atlas (Figure 1). The coast as well as the plains and high altitude plateaus between the mountains are rich in cultures, forests and low plants providing forage to the honeybees. The climate is of Mediterranean type, and brood production is interrupted shortly in December during winter when the temperature drop, as well as during the warmest month in summer. The oases south of the Saharan Atlas are home to A. m. saharensis (Figure 1). Given the more extreme temperatures in this desertic region, brood interruption in both winter and summer is longer. The subspecies can be differentiated by their distinct distribution areas on each side of the Saharan Atlas and by their differences in coloration. A. m. intermissa is black, whereas A. m. sahariensis is yellow. A. m. intermissa colonies in the south of Algeria were identified based on the beekeepers’ migration records and worker body colour.

Location of sampling areas for Nosema spp. and virus screening (circles) as well as V. destructor (squared area). Apiaries of A. m. intermissa are numbered 1–6 and apiaries of A. m. sahariensis are numbered 7–10. Distribution ranges of the endemic honey bee subspecies are indicated in shaded areas.
2.2 Colony losses
Colony losses for the winter 2012–2013 were quantified with a questionnaire sent to beekeepers (Van der Zee et al. 2012). They were asked how many production colonies they had before winter and how many of these colonies were lost during winter (December to March). From these values, we calculated the percentage of colonies lost over winter. Out of the total 165 beekeepers who answered the questionnaire, 136 of the participating beekeepers kept A. m. intermissa in the north of Algeria, while 20 beekeepers were known to migrate colonies of this subspecies to the South: In the South, nine beekeepers using the endemic A. m. sahariensis answered the questionnaire. The participating individuals represent 35 % of the beekeepers in their administrative districts.
2.3 Colonies used for parasite and pathogen screening
We collected samples from 130 apiaries within the administrative districts where beekeeping is practised (Figure 1) and belonging to beekeepers volunteering to take part to the study. Ninety apiaries located in northern Algeria and 28 in the South of the country hosted A. m. intermissa. In addition, 12 apiaries hosting A. m. sahariensis in southern Algeria were used. From each apiary, we randomly selected three apparently healthy colonies for sampling. These colonies showed no obvious clinical signs of varroasis, nosemosis, American or European foulbrood. Workers collected from brood frames were fixed in RNA later solution and placed in a refrigerated box for transportation to the laboratory. After a maximum of 2-h transport, the samples were frozen at −80 °C until shipping to Switzerland for molecular diagnosis (Dainat et al. 2011). After transport at room temperature between the countries, samples were frozen at −20 °C for a few days until analysis.
2.4 Varroa destructor
To determine the average V. destructor infestation rate of an adult honey bees per apiary (N = 130), 300 workers were collected from the brood frames of each colony sampled (N =360, 1-3 colonies per apiary). This was done between January and April 2012, a period between winter and beginning of spring in both north and south Algeria. The samples were stored at −18 °C until infestation rate measurement. For this measurement, we separated the mites from their host by placing the workers in jars filled with 70° ethanol and shaking for 3 min (Shimanuki and Knox 2000). To measure brood infestation rates, we obtained two frames with capped brood from each colony. A total of 250 cells were opened per colony to calculate the percentage of infested cells (see Dietemann et al. 2013 for methodology).
2.5 Virus and Nosema spp. screening
Six of the 90 A. m. intermissa and four of the 12 A. m. sahariensis apiaries were selected in the North and South regions, respectively (Figure 1) to be representative for both subspecies. Samples of 50 worker bees were collected once in spring, between May and July 2012. Virus screening and Nosema spp. species identification were performed on pools of these 50 workers. V. destructor functions as a vector and host for honeybee viruses (Rosenkranz et al. 2010). In order to determine whether these parasites might play a role in spreading viruses in Algerian honey bee populations, we also screened one to nine mites obtained from one to three colonies in five A. m. intermissa and three A. m. sahariensis apiaries (totalling 12 and six colonies, respectively). See supplementary material for technical details on molecular diagnosis of the pathogens.
2.6 Statistical analysis
A χ 2 test with Yates correction was performed to compare virus prevalence between A. m. intermissa and A. m. sahariensis colonies. The same test was used to compare Nosema spp. prevalence between the subspecies. An ANOVA was used to compare the proportions of colony losses between the subspecies and the V. destructor infestation rates between the three population studied (N = 130 colonies). Percentage data were transformed with arcsine. Pairwise comparisons were performed with the post-hoc Scheffé test. Normal distribution was verified with the K-S (Lilliefors) test and homogeneity of variance with the Levene test. When one of these assumptions was not met, the results of the ANOVA were confirmed with a Kruskal-Wallis non-parametric test. Statistical analyses were performed with SYSTAT (version13.0).
3 Results
3.1 Colony losses
The proportion of colony losses was significantly higher for A. m. intermissa in the North than for both subspecies in the South (ANOVA, F (2162) = 45.02, P < 0.01, Figure 2), where there was no significant difference between A. m. intermissa and A. m. sahariensis (P = 0.98).

Percentage of colony losses over the winter 2012/2013 in the Algerian honey bee populations studied.
3.2 Varroa destructor
Varroa mites were found in 100 % of the 118 A. m. intermissa colonies (both from the north and south of Algeria) and in 100 % out of the 12 A. m. sahariensis colonies. Average infestation rates of adult bee populations varied in a narrow range (13.2 and 14.2 %) between the three populations (Figure 3). The same trend could be observed for average brood infestation rates, which ranged between 4.3 and 5.6 % between the three populations. Adult infestation rates were not significantly different between North and South populations of A. m. intermissa and between A. m. intermissa and A. m. sahariensis (ANOVA, F (2127) = 1.22, P = 0.30). The same was not true for the brood infestation rates, which were significantly different (ANOVA, F (2127) = 4.35, P = 0.02, Figure 3). Brood infestation rate of A. m. intermissa in the North was significantly higher than A. m. sahariensis (P = 0.024), but not than A. m. intermissa colonies kept in the South (P = 0.35).

V. destructor infestation rates (±SD) of adult bees and brood in three populations of A. m. intermissa and A. m. sahariensis in Algeria. Different letters above the bars indicate P values below 0.05 with the Scheffé test.
3.3 Viruses
Six of the seven viruses screened were detected at prevalence ranging from 6 to 100 %. There was no significant difference in prevalence at colony level of most viruses between the two subspecies (χ2 CBPV(1, N = 29) = 0.24, P CBPV < 0.62; χ2 BQCV(1, N = 29) = 0.32, P BQCV < 0.57; χ2 DWV(1, N = 29) = 0.26, P DWV < 0.61; χ2 APBV(1, N = 29) = 0.0, P ABPV = 1.0; χ2 SBV(1, N = 11) = 0.85, P SBV = 0.36). Of those, CBPV was the most prevalent, followed by BQCV, DWV, APBV and SBV (Figure 4). IAPV showed the largest difference between subspecies (Figure 4), but the difference was not significant (χ2 IAPV (1, N = 29) = 3.56; P IAPV = 0.06). IAPV came third in the decreasing order of prevalence in A. m. sahariensis and fifth in A. m. intermissa. VDV-1 was not found in this survey. The proportion of dual infections in the honeybee samples was 27 %, which of triple infections 16 % and 12 % for quadruple infections (Supplementary material Table S2).

Average apiary pathogen prevalence in percent (±SD) for A. m. intermissa in north Algeria (N = 17 colonies in six apiaries) and A. m. sahariensis in south Algeria (N = 12 colonies in four apiaries).
In contrast to honey bees, ABPV was the most prevalent virus in V. destructor found on A. m. intermissa with 33 % of samples infected, followed by CBPV (20 %) and DWV (10 %). In A. m. sahariensis, only DWV was detected and found in 33 % of the mite samples. BQCV, SBV and IAPV were not detected in mites.
3.4 Nosema spp.
In both subspecies, most colonies (Figure 4) and all apiaries were found infected with N. ceranae. There was no significant difference in prevalence between the subspecies (χ 2 (1, N = 29) = 0.03, P = 0.86). N. apis was not detected.
4 Discussion
Colony losses were low in Algeria during winter 2012–2013, in comparison to most regions of Europe and North America at this time (van der Zee et al. 2012, 2014; vanEngelsdorp et al. 2012; Spleen et al. 2013; Steinhauer et al. 2014). In the south of Algeria, losses were approximately three times lower than apiaries located in the North, with no effect of honey bee subspecies or obvious link to pathogen or parasite prevalence. Colony management in Algeria is similar in the North and the South, irrespective of the subspecies used, and this parameter can a priori be excluded as an explanation for the differential mortality. Losses are therefore possibly affected by other factors linked with geographical or climatic factors, or use of agrochemicals (Adjlane et al. 2012a) in certain regions of the north of Algeria.
Remarkably, low losses occurred despite high prevalence of parasitic mites and pathogens in the colonies at the beginning of the year. V. destructor infestation rates of adults were similar in all regions and subspecies, and therefore do not explain the differences observed in morbidities. Brood infestation rates were not significantly different between the subspecies located in the South. The duration of the brood stops due to winter thus cannot explain the difference in infestation rate compared to A. m. intermissa in the North. Although a significant higher brood infestation rate in A. m. intermissa in the North could explain higher losses in this population, the range of infestation in comparison to the other population was similar and the effect on colony health expected to be weak. The high number of mites recorded both in the brood and on adult workers, in a majority of colonies during spring, was unexpected. Based on the damage thresholds identified in Europe and North America, most of these colonies should have collapsed during the previous winter (Genersch et al. 2010; Guzman-Novoa et al. 2010). The longer brood interruption in the South could limit the reproduction of V. destructor in the colonies (Moretto et al. 1991; Medina-Flores et al. 2014). In line with this hypothesis, mite treatments are less frequent in the South (one, rarely two treatments per year versus two to three in the North), which could result from a lower parasitic pressure at certain critical time of the year, a trend not visible based on a unique mite sampling as performed in our study. The monitoring of V. destructor population dynamics in these honeybee populations could help understand the relationship between the occurrence of the parasite and virus virulence and their effect on honeybee health.
Despite high number of mites recorded in the colonies in spring, the pattern of virus expected from surveys performed in Europe and North America (Tentcheva et al. 2004; vanEngelsdorp et al. 2009; Francis et al. 2013) was not observed. For instance, the high number of CBPV and BQCV positive colonies reported here (90–100 % and 72–92 % of colonies infected, respectively) was higher than expected, since these viruses are not usually associated with V. destructor. CBPV and BQCV occurrence has so far not been associated with colony losses. It is thus unlikely that these viruses generate mortalities in Algeria. The absence of clinical signs reported in other African honey bee subspecies with high BQCV prevalence (Kajobe et al. 2010; Strauss et al. 2013; Muli et al. 2014) supports this idea.
In contrast, the DWV-VDV-1 as well as the ABPV-KBV-IAPV viral complexes is commonly found associated with V. destructor infestations and colony mortalities in Europe. DWV was detected in a large proportion of the colonies, but was not as ubiquitous as expected (de Miranda and Genersch 2010). It showed a lower, but not significantly different, prevalence in A. m. intermissa compared to A. m. sahariensis colonies (61 and 75 %, respectively), in spite of higher mortality records in northern Algeria. This virus is thus not likely to be responsible for the colony mortality observed in north Algeria. Although the prevalence values measured overlap with those recorded in Europe (e.g. Nguyen et al. 2010; Nielsen et al. 2008; Tentcheva et al. 2004), or in Africanized bee populations (Antùnez et al. 2006), it is worth mentioning that most surveys performed in other African honey bee populations have not, or only occasionally, detected this virus (Kajobe et al. 2010; Strauss et al. 2013; Muli et al. 2014; Amakpe et al. 2015). The absence of detection of VDV-1, a virus closely related to DWV (De Miranda and Genersch 2010), in both honey bee subspecies tested here also contrasts with other surveys performed in Europe or the Middle East, where this virus is frequently detected (De Miranda and Genersch 2010; Chejanovsky 2010). IAPV prevalence was also atypical since it was higher in Algeria compared to European as well as other African and Africanized honey bees (e.g. Ai et al. 2012; Blanchard et al. 2008; Cox-Foster et al. 2007; Strauss et al. 2013; Muli et al. 2014; Amakpe et al. 2015). This virus was more prevalent in A. m. sahariensis than in A. m. intermissa (39 and 83 %, respectively). This difference is inverse compared to the mortality trend observed between the subspecies. As a result, the implication of this virus in the higher mortalities observed in A. m. intermissa in north Algeria is unlikely. The marginally significant difference in prevalence of IAPV recorded between Algerian subspecies could be due to different intrinsic resistance levels, but requires further investigations. A discrepancy between honey bee subspecies was not observed for the closely related ABPV. Although ABPV was associated with colony death in Europe (Genersch and Aubert 2010), in this case also, the prevalence and mortality patterns observed do not suggest a negative impact of this virus on Algerian colony survival. In contrast to the relatively high prevalence observed in this country, ABPV was absent or rare in surveys performed in Uganda, South Africa, Kenya and Benin (Kajobe et al. 2010; Strauss et al. 2013; Muli et al. 2014; Amakpe et al. 2015). As in other African or Africanized honey bee populations (Freiberg et al. 2012; Strauss et al. 2013; Muli et al. 2014), SBV was rarely detected in our samples, contrasting with studies in other parts of the world (Tentcheva et al. 2004; Ai et al. 2012; Chen et al. 2006; Antùnez et al. 2006; Reynaldi et al. 2010).
We found multiple virus infections in the colonies screened. Multiple infections are noted in other reports with values reaching 92 % of the colonies screened (Tentcheva et al. 2004; Antùnez et al. 2006; Forgach et al. 2008; Ai et al. 2012; Chen et al. 2004; Ryba et al. 2012). Their effect on honey bee health is yet to be determined. Overall, the high virus prevalence found in the honey bee populations screened in Algeria overlaps with the range associated with colony losses, more detailed surveys in the field are necessary to determine the sanitary status of both subspecies, and a better understanding of virus resistance in these subspecies is required.
The high prevalence of certain viruses in Algerian colonies, together with the high V. destructor infestation levels recorded in spring, was a reason to expect that mites act as vectors of these infections, as was demonstrated in many studies (Dainat et al. 2012a; vanEngelsdorp et al. 2009). However, few mites were found positive for viruses in this survey. With the exception of CBPV and BQCV, which were rarely detected in mite samples elsewhere (Celle et al. 2008; Locke et al. 2014), we expected a high frequency of DWV and IAPV-ABPV in Algerian mite samples. No mites were found positive for IAPV or SBV, while DWV and ABPV were only detected in mites of 10 and 16 % of the A. m. intermissa colonies, respectively. Only ABPV was found in mites of a third of the A. m. sahariensis colonies sampled. Concerning IAPV, our results are in line with those obtained from South Africa, where this virus was only occasionally detected in V. destructor (Strauss et al. 2013). Compared to the situation which prevails in Europe, the pattern found in Algeria supports the idea that viruses such as DWV still display a low virulence in this country, and therefore do not have a strong impact on colony health, especially during winter time. However, more detailed surveys covering several seasons are needed to support this hypothesis. In addition, the occurrence of different evolutionary scenarios in the mite-viruses interaction in Algeria or Africa could affect their prevalence and virulence (Martin et al. 2012; Ryabov et al. 2014) and explain the atypical patterns observed on this continent. Such interactions deserve further investigation in mite-tolerant honey bee populations and might help us understand the evolution of this host-parasite relationship in time and ultimately help design methods to mitigate its negative impact on susceptible populations (Dietemann et al. 2012).
In most of the world, Nosema spp. has been found at high prevalence at colony and apiary level reaching 95–100 % (e.g. Higes et al. 2010; Hong et al. 2011; Nabian et al. 2011). N. ceranae was found at higher prevalence than N. apis in the majority of samples (e.g. Strauss et al. 2013; Gisder et al. 2010; Higes et al. 2010; Nabian et al. 2011). Our survey of microsporidia infections in Algeria confirmed pure infections of N. ceranae in A. m. intermissa (Higes et al. 2009a) and showed for the first time its occurrence in A. m. sahariensis. In both subspecies, average prevalence at colony level was high with values of 100 and 90 %, respectively. No infection by N. apis could be detected. Our data thus correspond to the global trend of N. ceranae dominance and high prevalence with apparently low effects on colony health (Fries 2010). Since no historical data is available, we are not able to confirm or infirm the current hypothesis of a replacement of N. apis by N. ceranae (e.g. Paxton et al. 2007; Higes et al. 2013; Martín-Hernández et al. 2012). Given the total absence of N. apis in Algeria, it might have been completely replaced by N. ceranae. The resulting dominance of N. ceranae could reflect its better adaptation to warm climates (Forsgren and Fries 2010; Martín-Hernández et al. 2012). N. apis distribution is, based on cases for which species was identified, restricted to subequatorial Africa (Mumoki et al. 2014). Under the assumption that the endemic species in Africa is N. apis, this fact suggests that N. ceranae has not reached countries below the equator yet. Alternatively, N. ceranae could be endemic to Northern Africa, and N. apis might never have been present in this region. A more precise picture might emerge after surveying more African countries.
5 Conclusion
We confirmed the presence of common honey bee virus infections in A. m. intermissa and A. m. sahariensis populations, as well as the wide distribution of the gut parasite N. ceranae in Algerian apiaries. Despite an atypical distribution of some of these infections compared to other parts of the world, our results showed few differences in prevalence of viruses and microsporidia or in infestation rates with V. destructor between these two subspecies of honey bees. The impact of these pathogens on colony health in Algeria remains unclear, but appears low overall. Our data suggest that the difference in mortalities observed in the two subspecies could be due to local parameters, such as those related to climate. A more comprehensive knowledge on the virulence and impact of honey bee pathogens is necessary to understand the factors affecting honey bee health and be able to improve beekeeping management to mitigate colony losses.
References
Adjlane, N., Doumandji, S.E., Haddad, N. (2012a) Situation de l'apiculture en Algérie: facteurs menaçant la survie des colonies d'abeilles locales Apis mellifera intermissa. Cah. Agric. 21, 235–241. doi:10.1684/agr.2012.0566
Adjlane, N., Doumandji, S., Haddad, N. (2012b) La prévalence de la nosémose dans les colonies d’abeilles Apis mellifera intermissa dans la région médioseptrionale de l’Algérie. Leb. Sci. J 13, 65–73
Adjlane, N., Kechih, S., Doumandji, S., Haddad, N. (2012c) Survey of American foulbrood in Apis mellifera intermissa colonies in mid-northern region of Algeria. U. Arı Drg. 12, 98–105
Ai, H., Yan, X., Han, R. (2012) Occurrence and prevalence of seven bee viruses in Apis mellifera and Apis cerana apiaries in China. J. Invertebr. Pathol. 109, 160–164
Aizen, M.A., Harder, L.D. (2009) The global stock of domesticated honey bees is growing slower than agricultural demand for pollination. Curr. Biol. 19, 1–4. doi:10.1016/j.cub.2009.03.071
Amakpe, F., De Smet, L., Brunain, M., Ravoet, J., Jacobs, F.J., Reybroeck, W., Sinsin, B., de Graaf, D.C. (2015) Discovery of Lake Sinai virus and an unusual strain of acute bee paralysis virus in West African apiaries. Apidologie . doi:10.1007/s13592-015-0372-z
Antùnez, K., D’Alessandro, B., Corbella, E., Ramallo, G., Zunino, P. (2006) Honeybee viruses in Uruguay. J. Invertebr. Pathol. 91, 67–70
Berthoud, H., Imdorf, A., Haueter, M., Radloff, S., Neumann, P. (2010) Virus infections and winter losses of honey bee colonies (Apis mellifera). J. Apic. Res. 49, 60–65
Blanchard, P., Schurr, F., Celle, O., Cougoule, N., Drajnudel, P., Thiery, R., Faucon, J.P., Ribière, M. (2008) First detection of Israeli acute paralysis virus (IAPV) in France, a dicistrovirus affecting honeybees (Apis mellifera). J. Invertebr. Pathol. 99, 348–350
Breeze, T.D., Vaissière, B.E., Bommarco, R., Petanidou, T., Seraphides, N., et al. (2014) Agricultural policies exacerbate honeybee pollination service supply–demand mismatches across Europe. PLoS ONE 9(1), e82996. doi:10.1371/journal.pone.0082996
Celle, O., Blanchard, P., Schurr, F., Olivier, V., Cougoule, N., Faucon, J.P., Ribière, M. (2008) Detection of Chronic bee paralysis virus (CBPV) genome and RNA replication in various hosts: possible ways of spread. Virus Res. 133, 280–284
Chejanovsky, N. (2010) Paralytic viruses of the honey bee. U. Bee J. 10, 126–134
Chen, Y.P., Siede, R. (2007) Honey bee viruses. Adv. Virus Res. 70, 33–80
Chen, Y.P., Zhao, Y., Hammond, J., Hsu, H., Evans, J., Feldlaufer, M. (2004) Multiple virus infections in the honey bee and genome divergence of honey bee viruses. J. Invertebr. Pathol. 87, 84–93
Chen, Y.P., Pettis, J.S., Collins, A., Feldlaufer, M.F. (2006) Prevalence and transmission of honeybee viruses. Appl. Environ. Microbiol. 72, 606–611
Cornman, R.S., Tarpy, D.R., Chen, Y., Jeffreys, L., Lopez, D., Pettis, J.S., vanEngelsdorp, D., Evans, J.D. (2012) Pathogen webs in collapsing honey bee colonies. PLoS ONE 7, e43562. doi:10.1371/journal.pone.0043562
Cox-Foster, D.L., Conlan, S., Holmes, E.C., Palacios, G., Evans, J.D., et al. (2007) A metagenomic survey of microbes in honey bee colony collapse disorder. Science 318, 283–287
Dainat, B., Evans, J.D., Chen, Y.P., Neumann, P. (2011) Sampling and RNA quality for diagnosis of honey bee viruses using quantitative PCR. J. Virol. Methods 174, 150–152
Dainat, B., Evans, J.D., Chen, Y.P., Gauthier, L., Neumann, P. (2012a) Predictive markers of honey bee colony collapse. PloS One 7, e32151. doi:10.1371/journal.pone.0032151
Dainat, B., Evans, J.D., Chen, Y.P., Gauthier, L., Neumann, P. (2012b) Dead or alive: deformed wing virus and Varroa destructor reduce the life span of winter honeybees. Appl. Environ. Microbiol. 78, 981–987. doi:10.1128/AEM.06537-11
de Miranda, J.R., Genersch, E. (2010) Deformed wing virus. J. Invertebr. Pathol. 103, 48–61
Delaplane, K.S., Mayer, D.F. (2000) Crop pollination by bees. CABI Publishing, New York
Dietemann, V., Pirk, C.W.W., Crewe, R.M. (2009) Is there a need for conservation of honeybees in Africa? Apidologie 40, 285–295
Dietemann, V., Pflugfelder, J., Anderson, D., Charrière, J.-D., Chejanovsky, N., et al. (2012) Varroa destructor: research avenues towards sustainable control. J. Apic. Res. 51, 125–132. doi:10.3896/IBRA.1.51.1.15
Dietemann, V., Nazzi, F., Martin, S.J., Anderson, D., Locke, B. et al. (2013) Standard methods for Varroa research. In V Dietemann; J D Ellis; P Neusmann (Eds), The COLOSS BEEBOOK, Volume II: standard methods for Apis mellifera pest and pathogen research. J. Apic. Res. 52(4). doi: 10.3896/IBRA.1.52.1.09
Forgach, P., Bakonyi, T., Tapaszti, Z., Nowotny, N., Rusvai, M. (2008) Prevalence of pathogenic bee viruses in Hungarian apiaries: situation before joining the European Union. J. Invertebr. Pathol. 98, 235–238
Forsgren, E., Fries, I. (2010) Comparative virulence of Nosema ceranae and Nosema apis in individual European honey bees. Vet. Parasitol. 170, 212–217
Francis, R.M., Nielsen, S.L., Kryger, P. (2013) Varroa-Virus Interaction in collapsing honey bee colonies. PLoS ONE 8, e57540. doi:10.1371/journal.pone.0057540
Freiberg, M., De Jong, D., Message, D., Cox-Foster, D. (2012) First report of sacbrood virus in honey bee (Apis mellifera) colonies in Brazil. Genet. Mol. Res. 11, 3310–3314
Fries, I. (2010) Nosema ceranae in European honey bees (Apis mellifera). J. Invertebr. Pathol. 103, 73–79
Gallai, N., Salles, J.M., Settele, J., Vaissière, B.E. (2008) Economic valuation of the vulnerability of world agriculture confronted with pollinator decline. Ecol. Econ. 68, 810–821
Genersch, E., Aubert, M. (2010) Emerging and re-emerging viruses of the honey bee (Apis mellifera L.). Vet. Res. 41, 121–127
Genersch, E., von der Ohe, W., Kaatz, H., Schroeder, A., Otten, C., et al. (2010) The German bee monitoring project: a long term study to understand periodically high winter losses of honey bee colonies. Apidologie 41, 332–352
Gisder, S., Mockel, N., Linde, A., Genersch, E. (2010) A cell culture model for Nosema ceranae and Nosema apis allows new insights into the life cycle of these important honey bee-pathogenic microsporidia. Environ. Microbiol. 46, 1–10
Guzman-Novoa, E., Eccles, L., Calvete, Y., McGowen, J., Kelly, P.G., Corra-Benitez, A. (2010) Varroa destructor is the main culprit for the death and reduced populations of overwintered honey bee (Apis mellifera) colonies in Ontario, Canada. Apidologie 41, 443–450
Higes, M., Martin-Hernandez, R., Botias, C., Bailon, E.G., Gonzalez-Porto, A.V. (2008) How natural infection by Nosema ceranae causes honeybee colony collapse. Environ. Microbiol. 10, 2659–2669
Higes, M., Martín-Hernández, R., Garrido-Bailón, E., Botías, C., Meana, A. (2009a) The presence of Nosema ceranae (Microsporidia) in North African honey bees (Apis mellifera intermissa). J. Apic. Res. 48, 217–219
Higes, M., Martín-Hernandez, R., Meana, A. (2010) Nosema ceranae in Europe: an emergent type C nosemosis. Apidologie 41, 375–392
Higes, M., Meana, A., Bartolomé, C., Botías, C., Martín-Hernández, R. (2013) Nosema ceranae (Microsporidia), a controversial 21st century honeybee pathogen. Environ. Microbiol. 5, 17–29. doi:10.1111/1758-2229.12024
Hong, I., Woo, S., Choi, Y., Han, S., Kim, N., Kim, H., Han, S., Lee, M., Byeon, K. (2011) Prevalence of Nosema and virus in honey bee (Apis mellifera L.) colonies on flowering period of Acacia in Korea. Mycobiology 39, 317–320
Kajobe, R., Marris, G., Budge, G., Laurenson, L., Cordoni, G., Jones, B., Wilkinis, S., Cuthbertson, A.G., Brown, M.A. (2010) First molecular detection of a viral pathogen in Ugandan honey bees. J. Invertebr. Pathol. 104, 153–156
Li, J., Qin, H., Wu, J., Sadd, B.M., Wang, X., et al. (2012) The prevalence of parasites and pathogens in Asian honey bee Apis cerana in China. PloS One 7(1), 0047955. doi:10.1371/journal.pone.0047955
Locke, B., Forsgren, E., de Miranda, J.R. (2014) Increased tolerance and resistance to virus infections: A possible factor in the survival of Varroa destructor-resistant honey bees (Apis mellifera). PLoS ONE 9(6), e99998. doi:10.1371/journal.pone.0099998
Loucif-Ayad, W., Chefrour, A., Algharibeh, M., Haddad, N. (2013) First detection of deformed wing virus of honeybees in Algeria. Phytoparasitica 41, 445–447
Martin, S.J., Highfield, A.C., Brettell, L., Villalobos, E.M., Budge, G.E., Powell, M., Nikaido, S., Schroeder, D.C. (2012) Global honey bee viral landscape altered by a parasitic mite. Science 336, 1304–1306
Martín-Hernández, R., Botías, C., Bailón, E.G., Martínez-Salvador, A., Prieto, L., Meana, A., Higes, M. (2012) Microsporidia infecting Apis mellifera: coexistence or competition. Is Nosema ceranae replacing Nosema apis? Environ. Microbiol. 14, 2117–2138
Matheson, A. (1993) World bee health report. Bee World 74, 176–212
Medina-Flores, C.A., Guzman-Novoa, E., Hamiduzzaman, M.M., Arechiga-Flores, C.F., Lopez-Carlos, M.A. (2014) Africanized honey bees (Apis mellifera) have low infestation levels of the mite Varroa destructor in different ecological regions in Mexico. Genet. Mol. Res. 13, 7282–7293
Moretto, G., Gonçalves, L.S., De Jong, D., Bichuette, M.Z. (1991) The effects of climate and bee race on Varroa jacobsoni Oud. infestations in Brazil. Apidologie 22, 197–203
Muli, E., Patch, H., Frazier, M., Frazier, J., Torto, B., et al. (2014) Evaluation of the distribution and impacts of parasites, pathogens, and pesticides on honey bee (Apis mellifera) populations in East Africa. PLoS ONE 9(4), e94459. doi:10.1371/journal.pone.0094459
Mumoki, F.N., Fombong, A., Muli, E., Muigai, A.W.T., Masiga, D. (2014) An inventory of documented diseases of African honeybees. Afr. Entomol. 22(3), 473–487. doi:10.4001/003.022.0313
Nabian, S., Ahmadi, K., NazemShirazi, M.H., GeramiSadeghian, A. (2011) First detection of Nosema ceranae, a microsporidian protozoa of European honeybees (Apis mellifera) in Iran. Iran. J. Parasitol. 6, 89–95
Nazzi, F., Brown, S.P., Annoscia, D., Del Piccolo, F., Di Prisco, G., Varricchio, P., Della Vedova, G., Cattonaro, F., Caprio, E., Pennacchio, F. (2012) Synergistic parasite-pathogen interactions mediated by host immunity can drive the collapse of honeybee colonies. PLoS Pathog. 8(6), e1002735. doi:10.1371/journal.ppat.1002735
Nguyen, B.K., Mignon, J., Laget, D., De Graaf, D.C., Jacobs, F.J., Vanengelsdorp, D., Brostaux, Y., Saegerman, C., Haubruge, E. (2010) Honey bee colony losses in Belgium during the 2008–2009 winter. J. Apic. Res. 49, 337–339
Nielsen, S.L., Nicolaisen, M., Kryger, P. (2008) Incidence of acute bee paralysis virus, black queen cell virus, chronic bee paralysis virus, deformed wing virus, Kashmir bee virus and sacbrood virus in honeybees (Apis mellifera) in Denmark. Apidologie 39, 310–314
Paxton, R.J., Klee, J., Korpela, S., Fries, I. (2007) Nosema ceranae has infected Apis mellifera in Europe since at least 1998 and may be more virulent than Nosema apis. Apidologie 38, 558–565
Reynaldi, F.J., Sguazza, G.H., Pecoraro, M.R., Tizzano, M.A., Galosi, C.M. (2010) First report of viral infections that affect Argentine honeybees. Environ. Microbiol. Rep. 2, 123–131
Rosenkranz, P., Aumeier, P., Ziegelmann, B. (2010) Biology and control of Varroa destructor. J. Invertebr. Pathol. 103, S96–S119
Ryabov, E.V., Wood, G.R., Fannon, J.M., Moore, J.D., Bull, J.C., Chandler, D., et al. (2014) A virulent strain of deformed wing virus (DWV) of honeybees (Apis mellifera) prevails after Varroa destructor-mediated, or In Vitro, transmission. PLoS Pathog. 10(6), e1004230. doi:10.1371/journal.ppat.1004230
Ryba, S., Titera, D., Schodelbauerova-Traxmandlova, I., Kindlmann, P. (2012) Prevalence of honeybee viruses in the Czech Republic and coinfections with other honeybee disease. Biologia 67, 590–595
Shimanuki, H., Knox, D.A. (2000) Diagnosis of honey bee diseases. Agriculture Handbook No. AH690. US Department of Agriculture, Beltsville. 53 pp
Spleen, A.M., Lengerich, E.J., Rennich, K., Caron, D., Rose, R., et al. (2013) A national survey of managed honey bee 2011–12 winter colony losses in the United States: results from the Bee Informed Partnership. J. Apic. Res. 52(2), 44–53
Steinhauer, N.A., Rennich, K., Wilson, M.E., Caron, D.M., Lengerich, E.J., Pettis, J.S., Rose, R., Skinner, J.A., Tarpy, D.R., Wilkes, J.T., vanEngelsdorp, D. (2014) A national survey of managed honey bee 2012–2013 annual colony losses in the USA: results from the Bee Informed Partnership. J. Apic. Res. 53, 1–18
Strauss, U., Human, H., Gauthier, L., Crewe, R.M., Dietemann, V., Pirk, C.W.W. (2013) Seasonal prevalence of pathogens and parasites in the savannah honeybee (Apis mellifera scutellata). J. Invertebr. Pathol. 114, 45–52. doi:10.1016/j.jip.2013.05.003
Tentcheva, D., Gauthier, L., Zappulla, N., Dainat, B., Cousserans, F., Colin, M.E., Bergoin, M. (2004) Prevalence and seasonal variations of six bee viruses in Apis mellifera L. and Varroa destructor mite populations in France. Appl. Environ. Microbiol. 70, 7185–7191
Van der Zee, R., Pisa, L., Andonov, S., Brodschneider, R., Charrière, J.-D., et al. (2012) Managed honey bee colony losses in Canada, China, Europe, Israel and Turkey, for the winters of 2008–9 and 2009–10. J. Apic. Res. 51(1), 100–114
Van Der Zee, R., Brodschneider, R., Brusbardis, V., Charrière, J.D., Chlebo, R., Coffey, M.F., et al. (2014) Results of international standardised beekeeper surveys of colony losses for winter 2012–2013: analysis of winter loss rates and mixed effects modelling of risk factors for winter loss. J. Apic. Res. 53(1), 19–34. doi:10.3896/IBRA.1.53.1.02
vanEngelsdorp, D., Evans, J.D., Saegerman, C., Mullin, C., Haubruge, E., et al. (2009) Colony Collapse Disorder: A descriptive study. PLoS ONE 4(8), e6481. doi:10.1371/journal.pone.0006481
vanEngelsdorp, D., Hayes Jr., J., Underwood, R.M., Pettis, J.S. (2010) A survey of honey bee colony losses in the United States, fall 2008 to spring. J. Apic. Res. 49, 7–14
vanEngelsdorp, D., Caron, D., Hayes, J., Underwood, R., Henson, M., et al. (2012) A national survey of man-aged honey bee 2010–11 winter colony losses in the USA: results from the Bee Informed Partnership. J. Apic. Res. 51(1), 115–124
Winston, M.L. (1987) The biology of honey bee. Harvard University Press, Cambridge
Yang, B., Peng, G., Li, T., Kadowaki, T. (2013) Molecular and phylogenetic characterization of honey bee viruses, Nosema microsporidia, protozoan parasites, and parasitic mites in China. Ecol. Evol. 3(2), 298–311
Acknowledgments
N. A. was funded by a Short Term Scientific Mission from the Prevention of Honeybee Colony Losses (COLOSS) network (COST Action FA 0803). We thank two anonymous reviewers for their constructive comments on an earlier draft of the manuscript. We would like to acknowledge beekeepers, beekeeping associations and l'association du développement de l'apiculture dans la Mitidja (ADAM - Blida) for participating to this study.
Author information
Authors and Affiliations
Corresponding author
Additional information
Manuscript editor: David Tarpy
Profils viraux et parasitaires des sous-espèces algériennes d’abeilles, Apis mellifera intermissa et A . m . sahariensis
Apis mellifera intermissa / Apis mellifera sahariensis / virus / Nosema / Varroa destructor / perte de colonies / Algérie
Atypische Viren- und Parasitenmuster bei Unterarten der Honigbiene, Apis mellifera intermissa und A . m . sahariensis , in Algerien
Apis mellifera intermissa / Apis mellifera sahariensis / virus / Nosema / Varroa destructor / Völkerverluste / Algerien
Electronic supplementary material
Below is the link to the electronic supplementary material.
ESM 1
(PDF 135 kb)
Rights and permissions
About this article

Cite this article
Adjlane, N., Dainat, B., Gauthier, L. et al. Atypical viral and parasitic pattern in Algerian honey bee subspecies Apis mellifera intermissa and A. m. sahariensis . Apidologie 47, 631–641 (2016). https://doi.org/10.1007/s13592-015-0410-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13592-015-0410-x