
Abstract
Increasing temperature and consequent changes in climate adversely affect plant growth and development, resulting in catastrophic loss of wheat productivity. For each degree rise in temperature, wheat production is estimated to reduce by 6%. A detailed overview of morpho-physiological responses of wheat to heat stress may help formulating appropriate strategies for heat-stressed wheat yield improvement. Additionally, searching for possible management strategies may increase productivity and sustainability of growing wheat. The major findings from this review are as follows: (1) heat stress significantly reduces seed germination and seedling growth, cell turgidity, and plant water-use efficiency; (2) at a cellular level, heat stress disturbs cellular functions through generating excessive reactive oxygen species, leading to oxidative stress; (3) the major responses of wheat to heat stress include the enhancement of leaf senescence, reduction of photosynthesis, deactivation of photosynthetic enzymes, and generation of oxidative damages to the chloroplasts; (4) heat stress also reduces grain number and size by affecting grain setting, assimilate translocation and duration and growth rate of grains; (5) effective approaches for managing heat stress in wheat include screening available germplasm under field trials and/or employing marker-assisted selection, application of exogenous protectants to seeds or plants, mapping quantitative trait locus conferring heat resistance and breeding; (6) a well-integrated genetic and agronomic management option may enhance wheat tolerance to heat. However, the success of applying various techniques of heat stress management requires greater understanding of heat tolerance features, molecular cloning, and characterization of genes. The overall success of the complex plant heat stress management depends on the concerted efforts of crop modelers, molecular biologists, and plant physiologists.
Similar content being viewed by others

Contents
-
1. Introduction
-
2. Plant responses to heat stress
-
2.2.1 Water relations
-
2.2.3 Oxidative damage
-
2.2.4 Respiration
-
2.3.2 Starch synthesis
-
1 Introduction
Various environmental stresses affecting plant growth and development have attained a serious concern in the context of possible climate change. Contemporary agriculture faces a tremendous environmental pressure across the globe. Foley et al. (2011) suggested several management options including conservation tillage, adopting yield gap strategies, increasing cropping efficiencies that could be greatly effective to minimize environmental impacts and for sustainable crop production. However, the most remarkable environmental concern in agriculture is the increase of global temperature. With regard to global climate models, the mean ambient temperature is predicted to increase by 1–6°C by the end of twenty-first century (De Costa 2011). Such increase of global temperature may have a significant influence on agricultural productivity in accordance with the severity of the high temperature, drought, salinity, waterlogging, and mineral toxicity stresses. High temperature-induced heat stress is expressed as the rise in air temperature beyond a threshold level for a period sufficient to cause injury or irremediable damage of crop plants in general (Teixeira et al. 2013). The heat stress situation is aggravated when soil temperature increases as a result of increase in air temperature associated with decline in soil moisture. Thus, heat stress has appeared as a great menace to successful crop production in the world (Kumar et al. 2012; Lobell and Gourdji 2012; Gourdji et al. 2013).
Wheat (Triticum aestivum L.), the most widely cultivated cereal crop belonging to Poaceae family, is the largest contributor with nearly 30% of the world grain production and 50% of the world grain trade. FAO estimated that the world would require additional 198 million tonnes of wheat by 2050 to accomplish the future demands, for which wheat production need to be increased by 77% in the developing countries (Sharma et al. 2015). However, the temperature anomaly distribution is changing toward higher temperatures and the anomalies are increased (Hansen et al. 2012). Such a situation over the crop growing season has already been reported to reduce wheat productivity in the many regions of the world (Fontana et al. 2015; Mueller et al. 2015). Some indicators of heat stress effects in wheat are illustrated in Fig. 1.

Indicators of heat stress are stay green characteristics, rapid ground coverage and early heading and phenology (photo from a USDA research project)
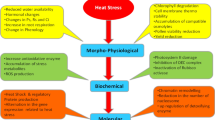
Wheat is very receptive to heat stress (Gupta et al. 2013a). Low latitude zones, where around 100 million hectares of wheat are cultivated, are predominantly heat prone areas worldwide (Braun et al. 2010). Asseng et al. (2014) tested 30 wheat crop models where mean temperatures in the growing season ranged from 15 to 32°C with artificial heating. The results obtained indicate that warming already decreased grain yield at a majority of the wheat-growing locations. The simulated median temperature impact on declining wheat yield varied widely, and the average yields for the periods between 1981 and 2010 decreased; ranging between 1 and 28% across 30 sites of the world; for an increase in temperature of 2°C; and this value rose to between 6 and 55% for a temperature of 4°C. Also they estimated that global wheat production falls by 6% for each 1°C of further temperature increase. The low latitudes showed a marked increase in simulated yield variability with higher temperature than that observed at high latitudes. This greater relative yield decline was due to the higher reference temperature (Challinor et al. 2014). Mondal et al. (2013) stated that the effects of heat stress on plants are very complex resulting in alteration of growth and development, changes in physiological functions, and reduced grain formation and yield (Fig. 2). Heat stress causes alteration of plant water relations (Hasanuzzaman et al. 2012, 2013), reduction of photosynthetic capacity (Almeselmani et al. 2012; Ashraf and Harris 2013), decreases of metabolic activities (Farooq et al. 2011) and changes of hormones (Krasensky and Jonak 2012), production of oxidative reactive species (Wang et al. 2011), promotion of ethylene production (Hays et al. 2007), reduction of pollen tube development, and increases of pollen mortality (Oshino et al. 2011) in wheat. During the period from 1880 to 2012, the Earth’s system warmed by 0.85°C (IPCC 2014). This warming period will continue and is predicted to rise between the range of 1.5–4.0°C in the future (Wheeler and Von Braun 2013). The climatic factors like changes in temperature, precipitation, CO2, weather variability, and soil moisture deficit would have positive or negative effects on crop production (Joshi and Kar 2009). The deleterious impacts of climate change on crop production are challenging the food security of the world, and it is predicted that sustaining wheat production will be impacted more by increasing temperature (Tripathi et al. 2016). Climate change could strongly affect wheat production, accounting for 21% of food and 200 million hectares of farmland worldwide (Ortiz et al. 2008).

Major effects of heat stress on plants growth and development. Pn, Rs, and Ci indicate photosynthesis, stomatal conductance, and intercellular CO2 concentration, respectively
Climate change impacts on crop production are highly diverse. Deryng et al. (2014) contributed greatly to current understanding of climate change impacts on crops under heat stress and elevated CO2 environment. The heat stress occurs usually for rising of canopy temperature that depends on air and soil temperature, soil and canopy properties, and loss of soil moisture (Fig. 3). High temperature affects crops in different ways including poor germination and plant establishment, reduced photosynthesis, leaf senescence, decreased pollen viability, and consequently production of less grains with smaller grain size (Ugarte et al. 2007; Asseng et al. 2011). Such effect varies depending on the crops, cultivars, and phenological stages. Due to global warming and changes in the climate pattern, it is imperative to determine the effects of heat stress and possible ways of improving heat tolerance for the success of wheat production under heat stress environment. Reidsma et al. (2010) approached various models of adaptation measures based on climate impact assessment. Deryng et al. (2014) considered choice of cultivars and changing sowing dates as adaptive measures under extreme heat stress conditions. Some other adaptation measures are surface cooling by irrigation (Lobell et al. 2008), antioxidants defense (Suzuki et al. 2011; Caverzan et al. 2016), and osmoprotectants (Farooq et al. 2011; Kaushal et al. 2016).

Schematic illustration of linking between climate-induced heat stress occurrence, plant responses to heat stress, and adaptation measures in the farmers’ field

Schematic diagram showing the exploration of wheat genetic resources and identifying wheat genotypes tolerant to heat stress, breeding strategies followed for genetic enhancement of diverse wheat genetic resource and adaptation strategies needed for managing genotypes considering phenology under heat stress environment for high yield potentials
However, development of heat-tolerant wheat varieties and generation of improved pre-breeding materials for any breeding program in future is crucial in meeting the food security (Ortiz et al. 2008). Proteomic and transcriptomic data are important to identifying genes and proteins that respond to environment, and affects yield and quality of wheat. However, more information regarding this is required to develop wheat variety that can adapt to climate change-induced high temperature (Altenbach 2012). In this context, this review covers an overview of the current work reported on heat-induced adverse effects and various crucial management strategies to address the heat stress situation in wheat.
2 Plant responses to heat stress
Heat stress affects various plant processes leading to morpho-physiological alterations in wheat plants, hindering the development processes and eventually resulting into great yield loss (McClung and Davis 2010; Grant et al. 2011). Plant responses to heat stress differ significantly with the extent and duration of temperature, and the growth stages encountering the stress (Ruelland and Zachowski 2010). Some common effects of heat stress on growth and productivity, grain development, and yield of wheat are presented in Table 1.
2.1 Morphological and growth responses
The primary effect of heat stress is the impediment of seed germination and poor stand establishment in many crops including wheat (Johkan et al. 2011; Hossain et al. 2013). Ambient temperature around 45°C severely affects embryonic cell in wheat which reduces crop stands through impairing seed germination and emergence (Essemine et al. 2010). Heat stress mostly affects the plant meristems and reduces plant growth by promoting leaf senescence and abscission, and by reducing photosynthesis (Kosova et al. 2011). Heat stress ranging from 28 to 30°C may alter the plant growth duration by reducing seed germination and maturity periods (Yamamoto et al. 2008). Warm environment produces lower biomass compared to plants grown under optimum or low temperature. Day and night temperature around 30 and 25°C, respectively, may have severe effects on leaf development and productive tiller formation in wheat (Rahman et al. 2009). However, the prevalence of reproductive stage heat stress has been found to be more detrimental in wheat production (Nawaz et al. 2013). One degree rise in average temperature during reproductive phase can cause severe yield loss in wheat (Bennett et al. 2012; Yu et al. 2014). High temperature stress degenerates mitochondria, changes the protein expression profiles, reduces ATP accumulation, and oxygen uptake in imbibing wheat embryos, resulting in increased occurrence of loss of seed quality relating to seed mass, vigor, and germination (Balla et al. 2012; Hampton et al. 2013). Increase in temperature of 1–2°C reduces seed mass by accelerating seed growth rate and by shortening the grain-filling periods in wheat (Nahar et al. 2010).
2.2 Physiological responses
2.2.1 Water relations
Plant water status is generally found to be most erratic under changing ambient temperature. High temperature seems to cause dehydration in plant tissue and subsequently restricts growth and development of plants. During flowering, a temperature of 31°C is generally considered as an upper limit of maintaining water status of a crop (Atkinson and Urwin, 2012). With a concomitant increase in leaf temperature, wheat plants exposed to heat stress substantially decrease the water potential and the relative water content in leaves, and eventually reduce photosynthetic productivity (Farooq et al. 2009). Simultaneously, the rate of transpiration and plant growth are severely affected. Almeselmani et al. (2009) observed that high temperature (35/25°C) imposed after tillering showed a significantly reduction of water potential in wheat, and the reduction was higher in genotypes susceptible to heat stress. In general, different antioxidants are associated with dehydration tolerance and are stimulated under heat stress. This is because of increased transpiration in stressed leaf and dropping of osmotic potential (Ahmad et al. 2010). Heat stress also increases hydraulic conductivity of cell membrane as well as plant tissues primarily for increased aquaporin activity (Martinez-Ballesta et al. 2009) and to a greater extent for reduced water viscosity (Cochard et al. 2007).
2.2.2 Photosynthesis, photosystems, and leaf senescence
Photosynthesis is the most sensitive physiological event leading to poor growth performance in wheat (Feng et al. 2014). A major effect of heat stress is the reduction in photosynthesis resulting from decreased leaf area expansion, impaired photosynthetic machinery, premature leaf senescence, and associated reduction in wheat production (Ashraf and Harris 2013; Mathur et al. 2014). The reaction sites of heat-induced injury are stroma and thylakoid lamellae of chloroplast where carbon metabolism and photochemical reactions occur, respectively. In wheat, heat stress causes disruption of thylakoid membranes, thereby inhibiting the activities of membrane-associated electron carriers and enzymes, which ultimately results in a reduced rate of photosynthesis (Ristic et al. 2008). The inactivation of chloroplast enzymes, mainly induced by oxidative stress, may also reduce the rate of leaf photosynthesis. Reduction of net photosynthetic rate due to heat stress is often attributed to increased non-photorespiratory processes (Ainsworth and Ort 2010). The researchers opined that impediment of photosynthetic activities is the result of reduced soluble protein, Rubisco and Rubisco binding proteins (Parry et al. 2011; Hasanuzzaman et al. 2013). Wheat leaf exposed to a high temperature around 40°C either in dark or light causes a great change in Rubisco and Rubisco activase and such changes are irreversible under dark conditions (Mathur et al. 2011).
In photosynthesizing tissues, photosystem-II is much responsive to heat stress (Marutani et al. 2012) but photosystem-I is relatively stable (Mathur et al. 2014). Heat stress firstly, damages the complex phenomena of photosystem-II and secondly, changes the photosynthetic behavior. The stress causes suppression of carbon assimilation due to inactivation of Rubisco activase in wheat. The reduction of carbon assimilation reduces ROS generation which, in turn, reduces protein synthesis and inhibits repairing of damaged photosystem-II (Murata et al. 2007; Allakhverdiev et al. 2008). Prasad et al. (2008a) also explained the sensitivity of photosystem-II where increasing fluidity of thylakoid membrane and transport of electron to heat stress are commonly observed. It is manifested that temperature >40°C dissociates the light harvesting complex-II Chl a/b-proteins from the photosystem-II (Iwaia et al. 2010). Heat stress damaging and disordering of thylakoid membranes is also responsible for the cessation of photophosphorylation (Dias et al. 2009a). At high temperature, the key regulatory enzyme of Rubisco, i.e., Rubisco activase is reported to be dissociated causing a reduction in the photosynthetic capacity of leaf in wheat (Raines 2011).
Leaf senescence is one of the inimitable symptoms of heat injury characterized by structural changes in the chloroplast followed by a vacuolar collapse, and thereafter a loss of plasma membrane integrity and interference of cellular homeostasis (Khanna-Chopra 2012). Thus, heat stressed wheat plants have been found to be experienced senescence-related metabolic changes (Ciuca and Petcu 2009). Inhibition of chlorophyll biosynthesis under heat stress (>34°C) may hasten leaf senescence in wheat (Asseng et al. 2013). Wheat plant exposed to heat stress during maturity enhanced leaf senescence, accentuated the loss of chloroplast integrity, and accelerated the turn-down of photosystem-II-mediated electron transport (Haque et al. 2014). However, a large diurnal variation in temperature is also responsible for the promotion of flag leaf senescence in wheat (Zhao et al. 2007).
2.2.3 Oxidative damage
Plants exposed to heat stress often leads to the generation of destructive ROS, including singlet oxygen (1O2), superoxide radical (O2−), hydrogen peroxide (H2O2), and hydroxyl radical (OH−) responsible for generating oxidative stress (Marutani et al. 2012; Suzuki et al. 2012). Oxidative stress notably increased membrane peroxidation and decreased membrane thermo-stability in many plants including wheat (Savicka and Skute 2010). Hydroxyl radicals react with almost all constituents of cells. Continual heat stress in plants may cause accumulation of ROS in cell plasma membrane with depolarization of cell membrane, activation of ROS-producing enzyme RBOHD and trigger of programmed cell death (Mittler et al. 2011). Miller et al. (2009) found that heat stress increased O2—production in root by 68% and malondialdehyde (MDA) content in leaf by 27% at the early stages, and 58% at the later stage of seedling development. However, plants have antioxidant mechanisms for escaping the excess ROS. Several studies have shown that antioxidants superoxide dismutase (SOD), ascorbate peroxidase (APX) and catalase (CAT) glutathione reductase (GR), and peroxidase (POX) have ameliorating effects of heat stress in wheat (Suzuki et al. 2011; Caverzan et al. 2016).
2.2.4 Respiration
Heat stress changes mitochondrial activities by affecting respiration. The rate of respiration increases with increasing temperature, but at a certain level of temperature, it diminishes due to damage of respiratory apparatus (Prasad et al. 2008b). The increased rate of respiratory carbon loss due to heat stress in the rhizosphere reduced production of ATP and enhanced the generation of ROS (Huang et al. 2012). This is because heat stress affects the solubility of CO2 and O2, and the kinetics of Rubisco (Cossani and Reynolds 2012). Almeselmani et al. (2012) observed that the rate of respiration in flag leaf of wheat was significantly higher in heat susceptible varieties under heat stress (35/25°C day/night) when compared with that of control (23/18°C day/night).
2.3 Grain growth and development
The optimum temperature for wheat anthesis and grain filling ranges from 12 to 22°C (Shewry 2009). Plants exposed to temperatures above >24°C during reproductive stage significantly reduced grain yield and yield reduction continued with increasing duration of exposure to high temperature (Prasad and Djanaguiraman 2014).
2.3.1 Grain number, grain filling, and grain quality
Heat stress reduces the number of grains leading to lower harvest index in wheat (Lukac et al. 2011). However, the influence of heat stress on both the number and size of grains varies with the growth stages encountering heat stress. For instance, temperatures above 20°C between spike initiation and anthesis speed up the development of the spike but reduce the number of spikelets and grains per spike (Semenov 2009). Heat stress adversely affects pollen cell and microspore resulting into male sterility (Anjum et al. 2008). Even high temperature of above 30°C during floret development may cause complete sterility in wheat depending on genotypes (Kaur and Behl 2010). In wheat, the anther produced under 3 days heat stress during anthesis was found to be structurally abnormal and nonfunctional florets (Hedhly et al. 2009). Day/night high temperature of 31/20°C may also cause shrinking of grains resulting from changing structures of the aleurone layer and cell endosperm (Dias et al. 2008).
Grain-filling stage in wheat is very sensitive to high temperature (Farooq et al. 2011). Heat stress generally accelerates the rate of grain-filling and shortens the grain-filling duration (Dias and Lidon 2009a). However, the grain growth rate and duration decreased in plants having different grain weight stability (Vijayalakshmi et al. 2010). In wheat, grain-filling duration may be decreased by 12 days with the increase of 5°C temperature above 20°C (Yin et al. 2009). The increase in night temperature is more responsive, shortens the grain-filling period, and reduces the grain yield than that of day temperature. Night temperatures of 20 and 23°C reduced the grain-filling period by 3 to 7 days (Prasad et al. 2008a). Recently, Song et al. (2015) observed a significant reduction in the rate of grain filling in wheat cultivars at day/night temperature of 32/22°C when compared with that of 25/15°C.
Heat stress affects grain quality of many cereals and legumes, essentially because of limitation of assimilates and less remobilization of nutrients. Heat stress hardly affects the protein concentration of grain in wheat (Lizana and Calderini 2013), but a strong correlation was observed between leaf nitrogen content and grain protein (Iqbal et al. 2017). However, wheat plants are capable of adopting a heat shock by developing thermo-tolerance for the improvement of the grain quality and yield (Sharma-Natu et al. 2010). Although build-up of protein in wheat grain is not significant under heat stress, the processing quality traits are reported important. Li et al. (2013) found that heat tends to diminish flour quality by reducing gluten strength-related parameters lactic acid retention capacity and mixograph peak time. However, wheat plants experiencing heat stress early in grain filling were found to have high content of grain protein (Castro et al. 2007). Increased grain protein content is associated with sedimentation index and intensity of essential amino acids. With decreased levels of amino acids, heat stress decreases the sedimentation index (Dias et al. 2008).
2.3.2 Starch synthesis
Wheat grain contains 60–75% starch of its total dry weight (Sramkova et al. 2009). Heat stress significantly limits starch biosynthesis in grains of wheat but caused a remarkable increase in total soluble sugar and protein (Sumesh et al. 2008; Asthir and Bhatia 2014). Liu et al. (2011) observed that heat-shock treatment above 30°C resulted in a significant increase of grain starch and limited the dry matter accumulation in grain of wheat. Around 97% of activity was lost due to the decrease in soluble starch synthase at 40°C, resulting in reducing grain growth and starch accumulation in wheat (Chauhan et al. 2011). High temperature stress (35/27°C) imposed at seedling stage significantly reduce soluble sugar accumulation and biomass yield in wheat (Wang et al. 2014).
2.3.3 Translocation of photosynthetic products
Photosynthetic products in the form of sucrose and glutamine are essentially translocated to the reproductive sinks for seed development. Under heat stress conditions, the source and sink limitations may reduce the growth and development of crop plants. Seed-set and -filling can also be restricted by source and/or sink limitations (Lipiec et al. 2013). When photosynthesis is inhibited by heat stress, stem reserves during pre-anthesis period are recognized as source of carbon for supporting grain filling (Mohammadi et al. 2009). In wheat, heat stress reduced N remobilization. The grain filling of wheat is seriously impaired by heat stress due to reduction in current leaf and ear photosynthesis. In case of heat-induced source limitation, plants seek to explore alternative source of assimilates to remobilize into the grains. At this time, stem reserves of water soluble carbohydrate and its greater translocation to reproductive organs are vitally important for supporting grain growth and development (Talukder et al. 2013). However, assimilate translocation occurring through both symplastic and apoplastic pathway is substantially reduced at high temperature. High temperature at the pre-anthesis period increased carbohydrate translocation from stem to grain resulting to less reduction of starch content in grains of wheat at the post-anthesis heat stress (Wang et al. 2012). In future, research directing to assimilate partitioning and phenotypic flexibility is suggested by Iqbal et al. (2017).
3 Managing heat stress
It is evident that heat stress adversely affects the growth and development of wheat plants. Such effects can be managed principally through producing appropriate plant genotypes together with adjustment of relevant agronomic practices (Asseng et al. 2011; Chapman et al. 2012). Various efforts have been made to produce heat-tolerant genotypes using the knowledge gained until now on the responses of wheat plant to heat stress. For sustainable wheat production in heat-stressed areas, the two most imperative strategies can be followed: (a) introduction of genetically modified or transgenic wheat cultivars by selecting molecular and biotechnological means coupled with conventional breeding approaches and (b) inducing several agronomic management strategies so far experiencing heat stress management under field conditions. A schematic diagram showed identification of wheat genotypes tolerant to heat stress, and breeding and adaptation strategies for managing wheat genotypes under heat stress environment (Fig. 4).
3.1 Genetic management
Breeding is an adaptation response of crops under changing environment. Therefore, it requires the evaluation of genetic diversity for adaptation to future climate change conditions, and thereby the selection and induction of stress inducible genes of genetic resources for developing new varieties in the production systems (Chapman et al. 2012). Breeding for heat tolerance is still in the preliminary stage and therefore, much attention is given to the genetic improvement of wheat to heat stress. In recent years, several studies have been done to find out wheat genotypes tolerant to heat stress (Kumar et al. 2010; Sareen et al. 2012; Kumari et al. 2013; Nagar et al. 2015).
3.1.1 Screening and breeding for heat tolerance
In breeding program, various physiological approaches have been found to be effective in Australia and several developing countries. The methodology includes screening genetic resources for identification of genetic bases for heat tolerance in crops. From this, a desired new plant types can be developed following physiological crossing of novel trait combinations, to combat future climate that comprises high temperature events (Reynolds and Langridge 2016). Screening wheat genotypes under natural heat stress condition in various spatial environments is difficult. Therefore, no consistent selection criterion has been established to evaluate diverse genetic materials for tolerance to heat stress. Selection criteria and screening methods for identifying better wheat genetic materials tolerant to heat stress are generally approached based on characteristics associated with higher grain yield under the adverse heat stress situation. In this regards, researchers suggested some indirect selection criteria for developing heat tolerance in wheat (Table 2).
Sharma et al. (2013) found susceptibility index as a consistent parameter while selecting wheat genotypes tolerant to heat stress. Mason et al. (2010) have given emphasis on quantitative trait loci (QTL) mapping of each yield attributes as susceptibility index and its collective contribution to heat tolerance and grain yield stability. Recent data shows that thylakoid membrane stability is highly associated with the heat tolerance capacity of wheat. Mass screening using stay-green character may be done for heat tolerance of wheat genotypes. Kumar et al. (2010) followed this method in evaluating stay-green trait of wheat and found a correlation with terminal heat tolerance in wheat. In general, morphological traits like early ground cover, leaf rolling, biomass, and also several physiological traits, such as leaf chlorophyll content, photosynthetic rate, flag leaf stomatal conductance, membrane thermostability, and stem reserves have been found to be associated with cellular thermotolerance in wheat plants. Recent advances in molecular science contributed greatly to understand the complexity of stress response mechanisms under heat stress conditions. Asthir (2015a, b) emphasized on the knowledge of molecular pathways and protective mechanisms to breed heat stress-tolerant plants. Heat tolerance is obviously a polygenic trait, and the above tools also aid in analyzing the genetic basis of plant thermotolerance. Wang et al. (2016) proposed a useful utilization of some transcription factors to improve multiple stress tolerance of crops. QTL mapping and subsequent marker-assisted selection made it possible to better understanding the heat tolerance in plants (Heffner et al. 2009). Recent studies reveal that several QTLs are available and can be used for developing heat tolerance in wheat. For example, QTLs for heat tolerance has been identified for grain weight and grain-filling duration (Mason et al. 2010; Paliwal et al. 2012), senescence-related traits (Vijayalakshmi et al. 2010), and canopy temperature (Paliwal et al. 2012). Mason et al. (2010) also identified QTLs related to yield and yield attribute traits and suggested that the spike of wheat could be used for locating QTL’s genomic zone for heat tolerance. Besides, others recognized QTLs on chromosomes 2B and 5B and 4A in wheat under heat stress conditions (Pinto et al. 2010). The electrolyte leakage is an indication of reduced cell membrane thermo-stability (CMT) and reflects the performance of wheat genotypes subjected to in vitro heat shock. Genotypes generating heat shock proteins (HSPs) can withstand heat stress as they protect proteins from heat-induced damage (Farooq et al. 2011). The genotypic differences in CMT tolerance in wheat at different growth stages were also reported (Kumar et al. 2013b; Asthir et al. 2013). The findings also suggested that the abundance of small heat shock protein and superoxide dismutase during milky-dough stage plays a vital role in the biosynthesis of starch granule, and this will help to develop heat-tolerant wheat cultivars containing high quality grains. For this, a simple, quick, and less costly screening method is required for a large number of germplasm to develop heat-tolerant wheat cultivars. As such, SPAD chlorophyll meter could be used for high throughput screening of wheat germplasm for heat tolerance (Ristic et al. 2007).
3.1.2 Biotechnological approach for improving heat tolerance
Genetic engineering and transgenic approaches can alleviate the adverse effects of heat stress by improving heat tolerance (Chapman et al. 2012). It involves the incorporation of genes of interest into the desired plants to improve plant tolerance to heat stress (Zheng et al. 2012). However, the complexity of the genomic pattern makes it difficult to research for genetic modification in wheat. Heat stress for a longer period increases protein synthesis elongation factor (EF-Tu) in chloroplast which is associated with heat tolerance in wheat. The constitutive expression of EF-Tu in transgenic wheat protected leaf proteins against thermal aggregation, reduced thylakoid membranes disruption, enhanced photosynthetic capability, and resisted pathogenic microbes infection (Fu et al. 2012). The wheat genotypes accruing more EF-Tu showed better tolerance to heat stress than those exposed to less EF-Tu (Ristic et al. 2008). Recently, many transcription factors (TFs) involved in various abiotic stresses have been found and engineered to improve stress tolerance in crops (Wang et al. 2016). Genome sequences of many plants are recently generated for improvement of stress tolerance. Clavijo et al. (2016) confirmed three known and identified one novel genome rearrangement of wheat. They used relatively inexpensive sequencing technologies and anticipated that researchers will use the approaches illustrated to sequence multiple wheat varieties. This will bring a large scale structural changes that are known to play a major role in the adaptation of the wheat crop to different stressful environments.
3.2 Agronomic management
Wheat can be grown successfully in a warmer environment through manipulating some agronomic management practices (Ortiz et al. 2008). Adoption of various agronomic practices like (i) water conserve techniques (ii) the appropriate amount and methods of fertilization (iii) maintaining proper time and methods of sowing, and (iv) the application of exogenous protectants can effectively alleviate the adverse impact of heat stress in wheat (Singh et al. 2011b).
3.2.1 Conserving soil moisture
A continuous supply of water is necessary for sustaining the grain-filling rate and duration, and grain size in wheat. This could not be possible in rain-fed wheat growing area, but here, mulching can be the best option for maintaining optimum moisture and thermal regimes in the soil system. Straw mulch conserves soil moisture by reducing soil evaporation (Chen et al. 2007). However, mulching is advocated to avoid yield reduction in wheat when reduced tillage is practiced (Glab and Kulig 2008). Increasing the productivity of wheat using mulch under heat stress and water deficit environment has been reported elsewhere (Chakraborty et al. 2008). Application of organic mulches preserves better soil moisture and improves plant growth and development, subsequently increases water and nitrogen use efficiency which may reduce (Singh et al. 2011b). This practice has been found to be very effective in wheat production under adverse heat stress conditions in temperate and tropical regions.
3.2.2 Nutrient management and planting time
Adequate and balanced supply of mineral nutrients is essential in plants exposed to temperature stress (Waraich et al. 2012). Application of nitrogen, phosphorus, and potassium at the post-anthesis period enriches grain proteins when the day and night temperatures remain 24 and 17°C, respectively, but effects are nullified for higher day and night temperature. Foliar spray of nutrients is very effective and can alleviate such adverse effect of heat stress on wheat. Application of potassium orthophosphate (KH2PO4) as a foliar spray after anthesis could be an alternative technique to increase the heat tolerance of wheat. Potassium orthophosphate causes a delay in the heat stress-induced leaf senescence and enhances grain yield (Dias and Lidon 2010). The advantages of NO3 − through delaying abscisic acid synthesis and promoting cytokinin activity, and similarly K+ induced increasing photosynthetic activities and assimilates accumulation are well recognized for increasing grain yield under heat stress environment (Singh et al. 2011a).
The exogenous application of calcium promotes heat tolerance in plants (Waraich et al. 2011). Calcium (Ca) application in the form of CaCl2 increased the malondialdehyde (MDA) content and stimulated the activities of guaiacol peroxidase, SOD, and CAT in wheat, which could be the reasons for the induction of heat tolerance. In bread wheat genotypes, Ca accumulation also seems to be linked with a higher tolerance to heat stress, possibly because this nutrient can shield chlorophylls from photo-destruction and can maintain stomata functioning, thus attenuating the heat stress effects through transpiration (Dias et al. 2009b).
Adequate supply of magnesium (Mg) was identified as an effective nutritional strategy to minimize heat stress-related losses in wheat production. Mengutay et al. (2013) found that wheat plants suffering from Mg deficiency were susceptible to heat stress, and sufficient Mg in the form of MgSO4.7H2O effectively alleviated the detrimental effect of heat stress (light/dark 35/28°C). Zinc (Zn) deficiency and heat stress also affect the wheat productivity by reducing kernel growth and chloroplast function (Peck and McDonald 2010). Heat stress generally increases Zn concentration in grain mostly due to remobilization from the shoot (Dias and Lidon 2009b). Therefore, Zn has also been proven to be effective in improving heat tolerance in wheat.
In general, late sowing wheat varieties faces severe temperature stress, shortens the heading and maturity duration, ultimately affecting final yield and grain quality (Hossain and Teixeira de Silva 2012; Hakim et al. 2012). Therefore, it is recommended for the development of high yielding wheat cultivars adapted to semiarid environments to select the genotypes with early maturity and a relatively long time to heading (Al-Karaki 2012). Hence, the early planting and the genotypes with early maturity and a relatively long time to heading are advocated to evade terminal heat stress and accelerate grain filling (Khichar and Niwas 2007; Al-Karaki 2012). Therefore, the maintaining appropriate planting time is one of the most important agronomic practices for getting optimum plant growth and yield of wheat under heat-stressed environment (Kajla et al. 2015).
Modification in planting method could alleviate the adverse impact of heat stress during the reproduction stage of wheat. Permanent bed planting under zero-tillage with crop residue retention has already been proposed as the possible means for improving heat stress tolerance in wheat plants for Northwest Mexico. Planting of wheat in conventional tillage with straw mulch increased water holding capacity, organic carbon, and total nitrogen in soil and improve tillering capacity resulting to mitigate the high temperature-induced reduction of grain weight at the late grain filling stage (Tang et al. 2013).
3.2.3 Use of exogenous protectants
In recent times, exogenously applied several growth-promoting protectants such as osmoprotectants, phytohormones, signaling molecules and trace elements have resulted in the potential to protect the plants by neutralizing the harmful and adverse effects of heat stress (Sharma et al. 2012; Upreti and Sharma 2016). Exogenously applications of these substances improve thermotolerance in wheat under heat stress by managing the ROS (Farooq et al. 2011) and upregulating the antioxidant capacity (Hemantaranjan et al. 2014). Treating thermo-sensitive wheat plants with several protectants, such as arginine, putrescine (Put), and α-tocopherol (vitamin E) have already established their roles in thermo-tolerance. External application of these molecules have ameliorating effects against oxidative stress through activation of various enzymatic viz. superoxide dismutase, catalase, ascorbate peroxidase, glutathione reductase and non-enzymatic viz. ascorbic acid, tocopherol, and glutathione antioxidants (Balla et al. 2007). The extensively used various plant bio-regulators in horticultural crops can be potentially used in field crops including wheat, and their prospects are now whipped out as in emerging stress alleviating technology in heat stress environment (Ratnakumar et al. 2016). The protective effects of exogenous application of arginine, putrescine, and tocopherol on heat-stressed wheat plants are shown in Table 3. Under abiotic stress condition, naturally occurring several intracellular polyamines (PAs), such as spermine, spermidine, and putrescine, can play vital roles for sustainable crop production. Recently, research directed towards polyamine biosynthesis, catabolism, and its role in abiotic stress tolerance is gaining priority (Gupta et al. 2013b; Rangan et al. 2014).
3.2.4 Bacterial seed treatment
Varietal improvement through the breeding program is time-consuming and costly, and gene transformation technology is not well perceived by many stakeholders. Therefore, using biological control agents like fungi and bacteria are now considered as an alternative method of improving heat tolerance (Raaijmakers et al. 2009). Plant growth-promoting rhizobacteria are found to be compatible and having a beneficial effect on the growth of wheat plants under heat stress (Nain et al. 2010). Seed treatment with rhizobacteria and foliar spray of various organic and inorganic agents enhanced heat tolerance in wheat (Yang et al. 2009). Seed inoculation with rhizobacteria also significantly improved heat tolerance in wheat (Anderson and Habiger 2012). Seed treatment with Bacillus amyloliquefaciens UCMB5113 and Azospirillum brasilense NO40 strains were also found to be effective to increase heat tolerance of wheat seedlings by reducing ROS generation (Abd El-Daim et al. 2014).
4 Conclusions and future perspective
In the recent past, heat stress was found to lead to enormous loss of wheat productivity worldwide. Despite carrying out intensive studies on the deleterious effects of heat stress in wheat, in-depth understanding of the mechanism of heat tolerance remains elusive. So, heat stress tolerance mechanism is vital for developing a notable strategy of wheat management under heat stress and forth seeing climate change settings. To generate heat-tolerant high yielding crops, metabolic and development processes associated with heat stress and energy regulation must be systematically understood. Although a considerable progress has been achieved in understanding the heat stress effects on wheat, yet there is a need for further understanding of the biochemical and molecular basis of heat tolerance for improvement of the crop yield from upcoming warmer environments. Molecular knowledge of response and tolerance mechanisms to harvest sustainable grain yields must be investigated. To recognize this, the functional genomic approach would be supportive in the response of wheat to heat stress.
It is well established that classical and modern molecular genetics tools integrated with the agronomic management practices can overcome the complexity of the heat syndrome. This is why the different biochemical and molecular approaches and agronomic options are required to explore the actual effects of heat stress on final crop yield. Moreover, the exogenous applications of protectants have revealed advantageous effects on heat tolerance improvement in wheat. Applying microorganisms seem to be a useful tool in agriculture to ameliorate the negative effects of heat stress on wheat plants, but further studies are needed to identify and optimize the parameters involved in successful microbial performance.
Thus, conventional breeding and modern biotechnological and molecular tools are an important area for future research. The actual basis of applying these methods is whether the plants contain heat tolerance or not. Wheat genotypes are found to express a substantial level of heat tolerance, although complete tolerance has not been found hitherto. Most alarming is that no selection criteria of heat tolerance have been established. Recently, heat sensitivity indexes for thousand kernel weight and grain filling duration have been developed.
Ultimately, an intimate collaboration and efforts amongst molecular biologists, plant physiologists, and breeders is required. A system-wide phenome to genome analysis is required to make possible an accurate trait mapping, introgression of superior alleles, or cloning of major QTLs for heat tolerance, and such combination will enable us to identify genes involved in heat tolerance and also the relationships between phenotypes and genotypes. For obtaining effective heat tolerance, the transgenic approach must be pooled with marker-assisted breeding programs for heat stress-related genes and QTLs.
In view of foreseen global warming, knowledge relating to molecular basis and mechanism of tolerance is considered to pave the way for engineering plants that can withstand heat stress and give satisfactory yield. Despite the fact that there is a possibility for application of EF-Tu in developing heat tolerant and disease resistant wheat varieties by modulating its expression levels, additional studies are mandatory to explore the mechanism of action of wheat EF-Tu relative to heat tolerance. It is important to note that molecular study affirms increasing economic crop yield, but full potential yield expression requires estimation of yield at crop level. So, crop modeling system studies are vital to improving heat stress tolerance and grain yield in wheat.
References
Abd El-Daim IA, Bejai S, Meijer J (2014) Improved heat stress tolerance of wheat seedlings by bacterial seed treatment. Plant Soil 379:337–350. doi:10.1007/s11104-014-2063-3
Ahmad P, Jaleel CA, Salem MA, Nabi G, Sharma S (2010) Roles of enzymatic and nonenzymatic antioxidants in plants during abiotic stress. Crit Rev Biotechnol 30:161–175. doi:10.3109/07388550903524243
Ainsworth EA, Ort DR (2010) How do we improve crop production in a warming world? Plant Physiol 154:526–530. doi:10.1104/pp.110.161349
Al-Karaki GN (2012) Phenological development-yield relationships in durum wheat cultivars under late-season high-temperature stress in a semiarid environment. International Scholarly Res Network 2012:1–7. doi:10.5402/2012/456856
Allakhverdiev SI, Kreslavski VD, Klimov VV, Los DA, Carpentier R, Mohanty P (2008) Heat stress: an overview of molecular responses in photosynthesis. Photosynth Res 98:541–550. doi:10.1007/s11120-008-9331-0
Almeselmani M, Deshmukh PS, Sairam RK (2009) High temperature stress tolerance in wheat genotypes: role of antioxidant defense enzymes. Acta Agron Hungar 57:1–14
Almeselmani M, Deshmukh PS, Chinnusamy V (2012) Effect of prolonged high temperature stress on respiration, photosynthesis and gene expression in wheat (Triticum aestivum L.) varieties differing in their thermotolerance. Plant Stress 6:25–32
Altenbach SB (2012) New insights into the effects of high temperature, drought and post-anthesis fertilizer on wheat grain development. J Cereal Sci 56:39–50. doi:10.1016/j.jcs.2011.12.012
Anderson M, Habiger J (2012) Characterization and identification of productivity-associated Rhizobacteria in wheat. Appl Environ Microbiol 78:4434–4446. doi:10.1128/AEM.07466-11
Anjum F, Wahid A, Javed F, Arshad M (2008) Influence of foliar applied thiourea on flag leaf gas exchange and yield parameters of bread wheat (Triticum aestivum) cultivars under salinity and heat stresses. Int J Agri Biol 10:619–626
Ashraf M, Harris PJC (2013) Photosynthesis under stressful environments: an overview. Photosynthetica 51:163–190. doi:10.1007/s11099-013-0021-6
Asseng S, Ewert F, Martre P, Rotter RP, Lobell DB, Cammarano D, Kimball BA, Ottman MJ, Wall GW, White JW et al (2014) Rising temperatures reduce global wheat production. Nat Clim Chang 5:143–147. doi:10.1038/nclimate2470
Asseng S, Foster I, Turner NC (2011) The impact of temperature variability on wheat yields. Glob Chang Biol 17:997–1012. doi:10.1111/j.1365-2486.2010.02262.x
Asseng S, Royce R, Cammarano D (2013) Temperature routines in wheat, workshop modeling wheat response to high temperature. Proceedings, Vol. VIII, p. 128. CIMMYT, Mexico, DF (Mexico). Jun 19–21
Asthir B (2015a) Mechanisms of heat tolerance in crop plants. Biologia Plant 59:620–628. doi:10.1007/s10535-015-0539-5
Asthir B (2015b) Protective mechanisms of heat tolerance in crop plants. J Plant Interactions 10:202–210. doi:10.1080/17429145.2015.1067726
Asthir B, Bala S, Bains NS (2013) Metabolic profiling of grain carbon and nitrogen in wheat as influenced by high temperature. Cereal Res Commun 41:230–242. doi:10.1556/CRC.2012.0036
Asthir B, Bhatia S (2014) In vivo studies on artificial induction of thermotolerance to detached panicles of wheat (Triticum aestivum L) cultivars under heat stress. J Food Sci Tech 51:118–123. doi:10.1007/s13197-011-0458-1
Asthir B, Koundal A, Bains NS (2012) Putrescine modulates antioxidant defense response in wheat under high temperature stress. Biol Plant 56:757–761. doi:10.1007/s10535-012-0209-1
Atkinson NJ, Urwin PE (2012) The interaction of plant biotic and abiotic stresses: from genes to the field. J Exp Bot 63:3523–3543. doi:10.1093/jxb/ers100
Bahar B, Yildirim M, Yucel C (2011) Heat and drought resistance criteria in spring bread wheat (Triticum aestivum L.): Morpho-physiological parameters for heat tolerance. Sci Res Essays 6:2212–2220. doi:10.5897/SRE11.418
Balla K, Bedo Z, Veisz O (2007) Heat stress induced changes in the activity of antioxidant enzymes in wheat. Cereal Res Commun 35:197–200
Balla K, Karsai I, Bencze S, Veisz O (2012) Germination ability and seedling vigour in the progeny of heat-stressed wheat plants. J Acta Agron Hung 60:299–308. doi:10.1556/AAgr.60.2012.4.1
Bennani S, Nsarellah N, Birouk A, Ouabbou H, Tadesse W (2016) Effective selection criteria for screening drought tolerant and high yielding bread wheat genotypes. Universal J Agril Res 4:134–142. doi:10.13189/ujar.2016.040404
Bennett D, Izanloo A, Reynolds M, Kuchel H, Langridge P, Schnurbusch T (2012) Genetic dissection of grain yield and physical grain quality in bread wheat (Triticum aestivum L.) under water limited environments. Theor Appl Genet 125:255–271. doi:10.1007/s00122-012-1831-9
Braun HJ, Atlin G, Payne T (2010) Multi-location testing as a tool to identify plant response to global climate change. In: Reynolds MP (ed) Climate change and crop production. CABI, Oxfordshire, pp 115–138
Castro M, Peterson CJ, Rizza MD, Dellavalle PD, V’azquez D, Ibanez V, Ross A (2007) Influence of heat stress on wheat grain characteristics and protein molecular weight distribution. In: Buck HT, Nisi JE, Salomon N (eds) Wheat production in stressed environment. Springer, Dordrecht, pp 365–371
Caverzan A, Casassola A, Brammer SA (2016) Antioxidant responses of wheat plants under stress. Genet Mol Biol 39:1–6. doi:10.1590/1678-4685-GMB-2015-010
Chakraborty D, Nagarajan S, Aggarwal P, Gupta VK, Tomar RK, Garg RN, Sahoo RN, Sarkar A, Chopra UK, Sarma KSS, Kalra N (2008) Effect of mulching on soil and plant water status, and the growth and yield of wheat (Triticum aestivum L.) in a semi-arid environment. Agric Water Manag 95:1323–1334. doi:10.1016/j.agwat.2008.06.001
Challinor AJ, Watson J, Lobell DB, Howden SM, Smith DR, Chhetri N (2014) A meta-analysis of crop yield under climate change and adaptation. Nat Clim Chang 4:287–291. doi:10.1038/nclimate2153
Chapman SC, Chakraborty S, Dreccer MF, Howden SC (2012) Plant adaptation to climate change-opportunities and priorities in breeding. Crop Pasture Sci 63:251–268. doi:10.1071/CP11303
Chauhan H, Khurana N, Tyagi A, Khurana J, Khurana P (2011) Identification and characterization of high temperature stress responsive genes in bread wheat (Triticum aestivum) and their regulation at various stages of development. Plant Mol Biol 75:35–51. doi:10.1007/s11103-010-9702-8
Chen SY, Zhang XY, Pei D, Sun HY, Chen SL (2007) Effects of straw mulching on soil temperature, evaporation and yield of winter wheat: field experiments on the North China plain. Ann Appl Biol 150:261–268. doi:10.1111/j.1744-7348.2007.00144.x
Ciuca M, Petcu E (2009) SSR markers associated with membrane stability in wheat (Triticum aestivum L.) Romanian Agric Res 26:21–24
Clavijo BJ, Venturini L, Schudoma C, Accinelli GG, Kaithakotti G, Wright J, Borrill P, Kettleborough G, Heavens D, Chapman H, Lipscombe J, Barker T, Lu F, McKenzie N, Raats D, Ramirez-Gonzalez RH, Coince A, Peel N, Percival-Alwyn L, Duncan O, Trosch J, Yu G, Bolser DM, Namaati G, Kerhornou A, Spannagl M, Gundlach H, Haberer G, Davey RP, Fosker C, Palma FD, Phillips A, Millar AH, Kersey PJ, Uauy C, Krasileva KV, Swarbreck D, Bevan MW, Clark MD (2016) An improved assembly and annotation of the allohexaploid wheat genome identifies complete families of agronomic genes and provides genomic evidence for chromosomal translocations. Genome Res 27:1–12http://www.genome.org/cgi/doi/10.1101/gr.217117.116
Cochard H, Venisse JS, Barigah TS, Brunel N, Herbette S, Guilliot A, Tyree MT, Sakr S (2007) Putative role of aquaporins in variable hydraulic conductance of leaves in response to light. Plant Physiol 143:122–133. doi:10.1104/pp.106.090092
Cossani CM, Reynolds MP (2012) Physiological traits for improving heat tolerance in wheat. Plant Physiol 160:1710–1718. doi:10.1104/pp.112.207753
De Costa WAJM (2011) A review of the possible impacts of climate change on forests in the humid tropics. J Natl Sci Found Sri 39:281–302. doi:10.4038/jnsfsr.v39i4.3879
Deryng D, Conway D, Ramankutty N, Price J, Warren R (2014) Global crop yield response to extreme heat stress under multiple climate change futures. Environ Res Lett 9:1–13. doi:10.1088/1748-9326/9/3/034011
Dhanda SS, Munjal R (2012) Heat tolerance in relation to acquired thermotolerance for membrane lipids in bread wheat. Field Crops Res 135:30–37. doi:10.1016/j.fcr.2012.06.009
Dias AS, Bagulho AS, Lidon FC (2008) Ultrastructue and biochemical traits of bread and durum wheat grains under heat stress. Brazz J Plant Physiol 20:323–333. doi:10.1590/S1677-04202008000400008
Dias AS, Lidon FC (2009a) Evaluation of grain filling rate and duration in bread and durum wheat under heat stress after anthesis. J Agron Crop Sci 195:137–147. doi:10.1111/j.1439-037X.2008.00347.x
Dias AS, Lidon FC (2009b) Heat stress in Triticum: kinetics of Cu and Zn accumulation. Brazilian J Plant Physiol 21:135–142
Dias AS, Lidon FC (2010) Bread and durum wheat tolerance under heat stress: a synoptical overview. Emir J Food Agric 22:412–436. doi:10.9755/ejfa.v22i6.4660
Dias AS, Lidon FC, Ramalho JC (2009a) IV. Heat stress in Triticum: kinetics of Fe and Mg accumulation. Brazilian J Plant Physiol 21:153–164. doi:10.1590/S1677-04202009000200008
Dias AS, Lidon FC, Ramalho JC (2009b) I. Heat stress in Triticum: kinetics of Ca and Mg accumulation. Brazilian J Plant Physiol 21:123–134. doi:10.1590/S1677-04202009000200005
Essemine J, Ammar S, Bouzid S (2010) Impact of heat stress on germination and growth in higher plants: physiological, biochemical and molecular repercussions and mechanisms of defence. J Biol Sci 10:565–572. doi:10.3923/jbs.2010.565.572
Farooq M, Bramley H, Palta JA, Siddique KHM (2011) Heat stress in wheat during reproductive and grain-filling phases. Crit Rev Plant Sci 30:491–507. doi:10.1080/07352689.2011.615687
Farooq M, Wahid A, Kobayashi N, Fujita D, Basra SMA (2009) Plant drought stress: effects, mechanisms and management. Agron Sustain Dev 29:185–212. doi:10.1051/agro:2008021
Feng B, Liu P, Li G, Dong ST, Wang FH, Kong LA, Zhang JW (2014) Effect of heat stress on the photosynthetic characteristics in flag leaves at the grain-filling stage of different heat-resistant winter wheat varieties. J Agron Crop Sci 200:143–155. doi:10.1111/jac.12045
Foley JA, Ramankutty N, Brauman KA, Cassidy ES, Gerber JS, Johnston M, Mueller ND, O’Connell C, Ray DK, West PC, Balzer C, Bennett EM, Carpenter SR, Hill J, Monfreda C, Polasky S, Rockstrom J, Sheehan J, Siebert S, Tilman D, Zaks DPM (2011) Solutions for a cultivated planet. Nature. doi:10.1038/nature10452
Fontana G, Toreti A, Ceglar A, De Sanctis G (2015) Early heat waves over Italy and their impacts on durum wheat yields. Nat. Hazards Earth Syst. Sci 15:1631–1637. doi:10.5194/nhess-15-1631-2015
Fu J, Momclovic I, Prasad V (2012) Molecular bases and improvement of heat tolerance in crop plants. In: Josipovic S, Ludwig E (eds) Heat stress: causes. Prevention and Treatments. Nova Science, USA, pp 185–214
Glab T, Kulig B (2008) Effect of mulch and tillage system on soil porosity under wheat (Triticum aestivum). Soil Tillage Res 99:169–178
Gourdji S, Mathews KL, Reynolds M, Crossa J, Lobell DB (2013) An assessment of wheat yield sensitivity and breeding gains in hot environments. Proc Biol Sci 280:1471–2954. doi:10.1098/rspb.2012.2190
Grant RF, Kimball BA, Conley MM, White JW, Wall GW, Ottman MJ (2011) Controlled warming effects on wheat growth and yield: field measurements and modeling. Agron J 103(6):1742–1754. doi:10.2134/agronj2011.0158
Gupta NK, Agarwal S, Agarwal VP, Nathawat NS, Gupta S, Singh G (2013a) Effect of short-term heat stress on growth, physiology and antioxidative defence system in wheat seedlings. Acta Physiol Plant 35:1837–1842. doi:10.1007/s11738-013-1221-1
Gupta K, Dey A, Gupta B (2013b) Plant polyamines in abiotic stress responses. Acta Physiol Plant 35:2015–2036. doi:10.1007/s11738-013-1239-4
Hakim MA, Hossain A, Jaime A, da Silva T, Zvolinsky VP, Khan MM (2012) Yield, protein and starch content of twenty wheat (Triticum aestivum L.) genotypes exposed to high temperature under late sowing conditions. J Sci Res 4:477–489. doi:10.3329/jsr.v4i2.8679
Hampton JG, Boelt B, Rolston MP, Chastain TG (2013) Effects of elevated CO2 and temperature on seed quality. J Agric Sci 151:154–162. doi:10.1017/S0021859612000263
Hansen J, Sato M, Ruedy R (2012) Perception of climate change. Proc Natl Acad Sci 109:14726–14727. doi:10.1073/pnas.1205276109
Haque MS, Kjaer KH, Rosenqvist E, Sharma DK, Ottosen CO (2014) Heat stress and recovery of photosystem II efficiency in wheat (Triticum aestivum L.) cultivars acclimated to different growth temperatures. Environ Exp Bot 99:1–8. doi:10.1016/j.envexpbot.2013.10.017
Hasanuzzaman M, Hossain MA, da Silva JAT, Fujita M (2012) Plant responses and tolerance to abiotic oxidative stress: antioxidant defenses is a key factor. In: Bandi V, Shanker AK, Shanker C, Mandapaka M (eds) Crop stress and its management: perspectives and strategies. Springer, Berlin, pp 261–316
Hasanuzzaman M, Nahar K, Alam MM, Roychowdhury R, Fujita M (2013) Physiological, biochemical, and molecular mechanisms of heat stress tolerance in plants. Int J Mol Sci 14:9643–9684. doi:10.3390/ijms14059643
Hays DB, Do JH, Mason RE, Morgan G, Finlayson SA (2007) Heat stress induced ethylene production in developing wheat grains induces kernel abortion and increased maturation in a susceptible cultivar. J Plant Sci 172:1113–1123. doi:10.1016/j.plantsci.2007.03.004
Hedhly A, Hormaza JI, Herrero M (2009) Global warming and sexual plant reproduction. Trends Plant Sci 14:30–36. doi:10.1016/j.tplants.2008.11.001
Heffner EL, Sorrells ME, Jannink JL (2009) Genomic selection for crop improvement. Crop Sci 49:1–12. doi:10.2135/crop-sci2008.08.0512
Hemantaranjan A, Nishant Bhanu A, Singh MN, Yadav DK, Patel PK, Singh R, Katiyar D (2014) Heat stress responses and Thermotolerance. Adv plants Agric Res 1:00012. doi:10.15406/apar.2014.01.00012
Hossain A, Sarker MAZ, Saifuzzaman M, Teixeira da Silva JA, Lozovskaya MV, Akhter MM (2013) Evaluation of growth, yield, relative performance and heat susceptibility of eight wheat (Triticum aestivum L.) genotypes grown under heat stress. Int J Plant Production 7:615–636
Hossain A, Teixeira de Silva JA (2012) Phenology, growth and yield of three wheat (Triticum aestivum L.) varieties as affected by high temperature stress. Not Sci Biol 4:97–109. doi:10.15835/nsb437879
Huang B, Rachmilevitch S, Xu J (2012) Root carbon and protein metabolism associated with heat tolerance. J Exp Bot 63:3455–3465. doi:10.1093/jxb/ers003
Hurkman WJ, Vensel WH, Tanaka CK, Whitehand L, Altenbach SB (2009) Effect of high temperature on albumin and globulin accumulation in the endosperm proteome of the developing wheat grain. J Cereal Sci 49:12–23. doi:10.1016/j.jcs.2008.06.014
Hussain S, Jamil M, Napar AA, Rahman R, Bano A, Afzal F, Kazi AG, Mujeeb-Kazi A (2016) Heat stress in wheat and interdisciplinary approaches for yield maximization. In: Azooz MM, Ahmad P (eds) Plant-environment interaction: responses and approaches to mitigate stress, John Wiley & Sons, pp161–182
IPCC (Intergovernmental Panel on Climate Change) (2014) Summary for policymakers. In: Stocker TF, Qin D, Plattner GK, Tignor M, Allen SK, Boschung J, Nauels A, Xia Y, Bex V, Midgley PM (eds) Climate change 2013: the physical science basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge
Iqbal M, Raja NI, Yasmeen F, Hussain M, Ejaz M, Shah MA (2017) Impacts of heat stress on wheat: a critical review. Adv Crop Sci Tech 5:251–259. doi:10.4172/2329-8863.1000251
Iwaia M, Yokonoa M, Inadab N, Minagawa J (2010) Live-cell imaging of photosystem II antenna dissociation during state transitions. Proc Natl Acad Sci U S A 107:2337–2342. doi:10.1073/pnas.0908808107
Johkan M, Oda M, Maruo T, Shinohara Y (2011) Crop production and global warming. In: Casalegno S (ed) Global warming impacts-case studies on the economy, human health, and on urban and natural environments. Rijeka, Croatia, pp 139–152
Joshi NL, Kar A (2009) Contingency crop planning for dry land areas in relation to climate change. Indian J Agron 54(2):237–243
Kajla M, Yadav VK, Chhokar RS, Sharma RK (2015) Management practices to mitigate the impact of high temperature on wheat. J Wheat Res 7:1–12
Kaur V, Behl RK (2010) Grain yield in wheat as affected by short periods of high temperature, drought and their interaction during pre- and post-anthesis stages. Cereal Res Commun 38:514–520. doi:10.1556/crc.38.2010.4.8
Kaushal N, Bhandari K, Siddique KHM, Nayyar H (2016) Food crops face rising temperatures: An overview of responses, adaptive mechanisms, and approaches to improve heat tolerance. Cogent Food Agric 2:1134380
Khalil SI, El-Bassiouny HMS, Hassanein RA, Mostafa HA, El-Khawas SA, Abd El-Monem AA (2009) Antioxidant defense system in heat shocked wheat plants previously treated with arginine or putrescine. Aust J Basic Appl Sci 3:1517–1526
Khan, AA, Kabir MR (2014) Evaluation of spring wheat genotypes (Triticum aestivum L.) for wheat stress tolerance using different stress tolerance indices Cercetari Agronomice in Moldova, XLVII: 49–63
Khanna-Chopra R (2012) Leaf senescence and abiotic stresses share reactive oxygen species-mediated chloroplast degradation. Protoplasma 249:469–481. doi:10.1007/s00709-011-0308-z
Khichar ML, Niwas R (2007) Thermal effect on growth and yield of wheat under different sowing environments and planting systems. Indian J Agric Res 41:92–96
Kosova K, Vitamvas P, Prasil IT, Renaut J (2011) Plant proteome changes under abiotic stress-contribution of proteomics studies to understanding plant stress response. J Proteome 74:1301–1322. doi:10.1016/j.jprot.2011.02.006
Krasensky J, Jonak C (2012) Drought, salt, and temperature stress-induced metabolic rearrangements and regulatory networks. J Exp Bot 63:1593–1608. doi:10.1093/jxb/err460
Kumar S, Singh R, Nayyar H (2013b) α-tocopherol application modulates the response of wheat (Triticum aestivum l.) seedlings to elevated temperatures by mitigation of stress injury and enhancement of antioxidants. J Plant Growth Regul 32:307–314. doi:10.1007/s00344-012-9299-z
Kumar RR, Sharma SK, Goswami S, Singh GP, Singh R, Singh K, Pathak H, Rai RD (2013a) Characterization of differentially expressed stress-associated proteins in starch granule development under heat stress in wheat (Triticum aestivum L.) Indian J Biochem Biophys 50:126–138
Kumar S, Singh R, Grover M, Singh AK (2012) Terminal heat-an emerging problem for wheat production. Biotechnol Today 2:7–9
Kumar U, Joshi AK, Kumari M, Paliwal R, Kumar S, Roder MS (2010) Identification of QTLs for stay green trait in wheat (Triticum aestivum L.) in the ‘Chirya 3’ x ‘Sonalika’ population. Euphytica 174:437–445. doi:10.1007/s10681-010-0155-6
Kumari M, Pudake RN, Singh VP, Joshi AK (2013) Association of staygreen trait with canopy temperature depression and yield traits under terminal heat stress in wheat (Triticum aestivum L.) Euphytica 190:87–97. doi:10.1007/s10681-012-0780-3
Li YF, Wu Y, Hernandez-Espinosa N, Pena RJ (2013) Heat and drought stress on durum wheat: responses of genotypes, yield, and quality parameters. J Cereal Sci 57:398–404. doi:10.1016/j.jcs.2013.01.005
Lipiec J, Doussan C, Nosalewicz A, Kondracka K (2013) Effect of drought and heat stresses on plant growth and yield: a review. Int Agrophys 27:463–477. doi:10.2478/intag-2013-0017
Liu P, Guo W, Jiang Z, Pu H, Feng C, Zhu X, Peng Y, Kuang A, Little CR (2011) Effects of high temperature after anthesis on starch granules in grains of wheat (Triticum aestivum L.) J Agril Sci 149:159–169. doi:10.1017/S0021859610001024
Lizana XC, Calderini DF (2013) Yield and grain quality of wheat in response to increased temperatures at key periods for grain number and grain weight determination: considerations for the climatic change scenarios of Chile. J Agril Sci 151:209–221. doi:10.1017/S0021859612000639
Lobell DB, Bonfils CJ, Kueppers LM, Snyder MA (2008) Irrigation cooling effect on temperature and heat index extremes. Geophys Res Lett 35:L09705. doi:10.1029/2008GL034145
Lobell DB, Gourdji SM (2012) The influence of climate change on global crop productivity. Plant Physiol 160:1686–1697. doi:10.1104/pp.112.208298
Lopes MS, Reynolds MP (2012) Stay-green in spring wheat can be determined by spectral reflectance measurements (normalized difference vegetation index) independently from phenology. J Exp Bot 63:3789–3798. doi:10.1093/jxb/ers071
Lukac M, Gooding MJ, Griffiths S, Jones HE (2011) Asynchronous flowering and within-plant flowering diversity in wheat and the implications for crop resilience to heat. Ann Bot 109:843–850. doi:10.1093/aob/mcr308
Martinez-Ballesta MC, Lopez-Perez L, Muries B, Munoz-Azcarate O, Carvajal M (2009) Climate change and plant water balance: the role of aquaporins - a review. In: Lichtfouse E (ed) Climate change, intercropping, Pest control and beneficial microorganisms. Springer, Netherlands, pp 71–89. doi:10.1007/978-90-481-2716-05
Marutani Y, Yamauchi YKY, Mizutani M, Sugimoto Y (2012) Damage to photosystem II due to heat stress without light-driven electron flow: involvement of enhanced introduction of reducing power into thylakoid membranes. Planta 236:753–761. doi:10.1007/s00425-012-1647-5
Mason RE, Mondal S, Beecher FW, Pacheco A, Jampala B, Ibrahim AMH, Hays DB (2010) QTL associated with heat susceptibility index in wheat (Triticum aestivum L.) under short-term reproductive stage heat stress. Euphytica 174:23–436. doi:10.1007/s10681-010-0151-x
Mathur S, Agrawal D, Jajoo A (2014) Photosynthesis: response to high temperature stress. J Photochem Photobiol B: Biol 137:116–126
Mathur S, Jajoo A, Mehta P, Bharti S (2011) Analysis of elevated temperature-induced inhibition of photosystem II using chlorophyll a fluorescence induction kinetics in wheat leaves (Triticum aestivum). Plant Biol 13:1–6. doi:10.1111/j.1438-8677.2009.00319.x
McClung CR, Davis SJ (2010) Ambient thermometers in plants: from physiological outputs towards mechanisms of thermal sensing. Curr Biol 20:1086–1092. doi:10.1016/j.cub.2010.10.035
Mengutay M, Ceylan Y, Kutman UB, Cakmak I (2013) Adequate magnesium nutrition mitigates adverse effects of heat stress on maize and wheat. Plant Soil 368:57–72. doi:10.1007/s11104-013-1761-6
Miller G, Schlauch K, Tam R, Cortes D, Torres MA, Shulaev V, Dangl JL, Mittler R (2009) The plant NADPH oxidase RBOHD mediates rapid, systemic signaling in response to diverse stimuli. Sci Signal 2:1–10. doi:10.1126/scisignal.2000448
Mittler R, Vanderauwera S, Suzuki N, Miller G, Tognetti VB, Vandepoele K, Goller M, Shulaev V, Breusegem FV (2011) ROS signaling: the new wave? Trends Plant Sci 16:300–309. doi:10.1016/j.tplants.2011.03.007
Mohammadi M, Karimizadeh RA, Naghavi MR (2009) Selection of bread wheat genotypes against heat and drought tolerance on the base of chlorophyll content and stem reserves. J Agric Soc Sci 5:119–122
Mondal S, Singh RP, Crossa J, Huerta-Espino J, Sharma I, Chatrath R, Singh GP, Sohu VS, Mavi GS, Sukaru VSP, Kalappanavarg IK, Mishra VK, Hussain M, Gautam NR, Uddin J, Barma NCD, Hakim A, Joshi AK (2013) Earliness in wheat: a key to adaptation under terminal and continual high temperature stress in south Asia. Field Crops Res 151:19–26. doi:10.1016/j.fcr.2013.06.015
Mueller B, Hauser M, Iles C, Rimi RH, Zwiers FW, Wan H (2015) Lengthening of the growing season in wheat and maize producing regions. Weather Clim Extrem 9:47–56. doi:10.1016/j.wace.2015.04.001
Murata N, Takahashi S, Nishiyama Y, Allakhverdiev SI (2007) Photoinhibition of photosystem II under environmental stress. Biochim Biophys Acta 1767:414–421. doi:10.1016/j.bbabio.2006.11.019
Nagar S, Singh VP, Arora A, Dhakar R, Ramakrishnan S (2015) Assessment of terminal heat tolerance ability of wheat genotypes based on physiological traits using multivariate analysis. Acta Physiol Plant 37:257. doi:10.1007/s11738-015-2017-2
Nahar K, Ahamed KU, Fujita M (2010) Phenological variation and its relation with yield in several wheat (Triticum aestivum L.) cultivars under normal and late sowing mediated heat stress condition. Not Sci Biol 2:51–56
Nain L, Rana A, Joshi M, Jadhav SD, Kumar D, Shivay YS, Paul S, Prasanna R (2010) Evaluation of synergistic effects of bacterial and cyanobacterial strains as biofertilizers for wheat. Plant Soil 331:217–230. doi:10.1007/s11104-009-0247-z
Nawaz A, Farooq M, Cheema SA, Wahid A (2013) Differential response of wheat cultivars to terminal heat stress. Int J Agric Biol 15:1354–1358
Ortiz R, Sayre KD, Govaerts B, Gupta R, Subbarao GV, Ban T, Hodson D, Dixon JM, Ortiz-Monasterio JI, Reynolds M (2008) Climate change: can wheat beat the heat? Agric Ecosys Environ 126:46–58. doi:10.1016/j.agee.2008.01.019
Oshino T, Miura S, Kikuchi S, Hamada K, Yano K, Watanabe M, Higashitani A (2011) Auxin depletion in barley plants under high-temperature conditions represses DNA proliferation in organelles and nuclei via transcriptional alterations. Plant Cell Environ 34:284–290. doi:10.1111/j.1365-3040.2010.02242.x
Paliwal R, Röder MS, Kumar U, Srivastava JP, Joshi AK (2012) QTL mapping of terminal heat tolerance in hexaploid wheat (T. aestivum L.) Theor Appl Genet 125:561–575. doi:10.1007/s00122-012-1853-3
Parry MAJ, Reynolds M, Salvucci ME, Raines C, Andralojc PJ, Zhu XG, Price GD, Condon AG, Furbank RT (2011) Raising yield potential of wheat. II Increasing photosynthetic capacity and efficiency. J Exp Bot 62:453–467. doi:10.1093/jxb/erq304
Peck AW, McDonald GK (2010) Adequate zinc nutrition alleviates the adverse effects of heat stress in bread wheat. Plant Soil 337:355–374. doi:10.1007/s11104-010-0532-x
Pinto RS, Reynolds MP, Mathews KL, McIntyre CL, Olivares-Villegas J-J, Chapman SC (2010) Heat and drought adaptive QTL in a wheat population designed to minimize confounding agronomic effects. Theor Appl Genet 121:1001–1021. doi:10.1007/s00122-010-1351-4
Pradhan GP, Prasad PVV (2015) Evaluation of wheat chromosome translocation lines for high temperature stress tolerance at grain filling stage. PLoS One 10:e0116620. doi:10.1371/journal.pone.0116620
Prasad PVV, Djanaguiraman M (2014) Response of floret fertility and individual grain weight of wheat to high temperature stress: sensitive stages and thresholds for temperature and duration. Funct Plant Biol 41:1261–1269. doi:10.1071/FP14061
Prasad PVV, Pisipati SR, Ristic Z, Bukovnik U, Fritz AK (2008a) Impact of night time temperature on physiology and growth of spring wheat. Crop Sci 48:2372–2380. doi:10.2135/cropsci2007.12.0717
Prasad PVV, Staggenborg SA, Ristic Z (2008b) Impacts of drought and/or heat stress on physiological, developmental, growth, and yield processes of crop plants. In: Ahuja LH, Saseendran SA (eds) Response of crops to limited water: understanding and modeling water stress effects on plant growth processes. ASA, CSSA, Madison, pp 301–355
Raaijmakers JM, Paulitz TC, Steinberg C, Alabouvette C, Moënne-Loccoz Y (2009) The rhizosphere: a playground and battlefield for soilborne pathogens and beneficial microorganisms. Plant Soil 321:341–361. doi:10.1007/s11104-008-9568-6
Radhika, Thind SK (2014) Photosynthetic functioning as a selection criteria for maximum yield potential in some wheat genotypes under late planting heat stress conditions. Int J Sci Res Manag 2:974–982
Rahman MA, Chikushi J, Yoshida S, Karim AJMS (2009) Growth and yield components of wheat genotypes exposed to high temperature stress under control environment. Bangladesh J Agric Res 34:361–372. doi:10.3329/bjar.v34i3.3961
Raines CA (2011) Increasing photosynthetic carbon assimilation in C3 plants to improve crop yield: current and future strategies. Plant Physiol 155:36–42. doi:10.1104/pp.110.168559
Rangan P, Subramani R, Kumar R, Singh AK, Singh R (2014) Recent advances in polyamine metabolism and abiotic stress tolerance. BioMed Res Int 2014:239621, 9 pages. doi:10.1155/2014/239621
Ratnakumar P, Khan MIR, Minhas PS, Farooq MA, Sultana R, Per TS, Deokate PP, Khan NA, Singh Y, Rane J (2016) Can plant bio-regulators minimize crop productivity losses caused by drought, salinity and heat stress? An integrated review. J Appl Bot Food Qual 89:113–125. doi:10.5073/JABFQ.2016.089.014
Reidsma P, Ewert F, Lansink AO, Leemans R (2010) Adaptation to climate change and climate variability in European agriculture: the importance of farm level responses. Eur J Agron 32:91–102. doi:10.1016/j.eja.2009.06.003
Reynolds M, Langridge P (2016) Physiological breeding. Curr Opin Plant Biol 31:162–171. doi:10.1016/j.pbi.2016.04.005
Ristic Z, Bukovnik U, Momcilovic I, Fu J, Prasad PVV (2008) Heat-induced accumulation of chloroplast protein synthesis elongation factor, EF-Tu, in winter wheat. J Plant Physiol 165:192–202. doi:10.1016/j.jplph.2007.03.003
Ristic Z, Bukovnik U, Prasad PVV (2007) Correlation between heat stability of thylakoid membranes and loss of chlorophyll in winter wheat under heat stress. Crop Sci 47:2067–2073. doi:10.2135/cropsci2006.10.0674
Ruelland E, Zachowski A (2010) How plants sense temperature. Environ Exp Bot 69:225–232. doi:10.1016/j.envexpbot.2010.05.011
Sareen S, Tyagi BS, Tiwari V, Sharma I (2012) Response estimation of wheat synthetic lines to terminal heat stress using stress indices. J Agric Sci 4:97–104. doi:10.5539/jas.v4n10p97
Savicka M, Skute N (2010) Effects of high temperature on malondialdehyde content, superoxide production and growth changes in wheat seedlings (Triticum aestivum L.) Ekologija 56:26–33. doi:10.2478/v10055-010-0004-x
Semenov MA (2009) Impacts of climate change on wheat in England and Wales. J R Soc Interface 6:343–350. doi:10.1098/rsif.2008.0285
Sharma A, Rawat RS, Verma JS, Jaiswal JP (2013) Correlation and heat susceptibility index analysis for terminal heat tolerance in bread wheat. J Central Eur Agri 14:57–66. doi:10.5513/JCEA01/14.2.1233
Sharma I, Tyagi BS, Singh G, Venkatesh K, Gupta OP (2015) Enhancing wheat production- a global perspective. Indian J Agril Sci 85:3–13
Sharma P, Jha AB, Dubey RS, Pessarakli M (2012) Reactive oxygen species, oxidative damage and antioxidative defence mechanisms in plants under stressful conditions. J Bot 2012:1–26. doi:10.1155/2012/217037
Sharma RC, Tiwari AK, Ortiz-Ferrara G (2008) Reduction in kernel weight as a potential indirect selection criterion for wheat grain yield under heat stress. Plant Breed 127:241–248. doi:10.1111/j.1439-0523.2007.01460.x
Sharma-Natu P, Sumesh KV, Ghildiyal MC (2010) Heat shock protein in developing grains in relation to Thermotolerance for grain growth in wheat. J Agron Crop Sci 196:76–80. doi:10.1111/j.1439-037X.2009.00390.x
Shewry PR (2009) Wheat. J Exper Bot 60:1537–1553. doi:10.1093/jxb/erp058
Sikder S, Paul NK (2010) Effects of post-anthesis heat stress on stem reserves mobilization, canopy temperature depression and floret sterility of wheat cultivars. Bangladesh J Bot 39:51–55. doi:10.3329/bjb.v39i1.5526
Singh A, Singh D, Gill BS et al (2011a) Planting time, methods, and practices to reduce the deleterious effects of high temperature on wheat. In: the proceedings of international conference on preparing agriculture for climate change. Punjab Agricultural University, Ludhiana, pp 338–339 February, 6-8
Singh A, Singh D, Kang JS, Aggarwal N (2011b) Management practices to mitigate the impact of high temperature on wheat: a review. IIOABJ 2:11–22
Song WF, Zhao LJ, Zhang XM, Zhang YM, Li JL, Zhang LL, Song QJ, Zhao HB, Zhang YB, Zhang CL, Xin WL, Sun LF, Xiao ZM (2015) Effect of timing of heat stress during grain filling in two wheat varieties under moderate and very high temperature. Indian. J Genet 75:121–124. doi:10.5958/0975–6906.2015.00018.8
Sramkova Z, Gregova E, Sturdik E (2009) Chemical composition and nutritional quality of wheat grain. Acta Chim Slov 2:115–138
Sumesh KV, Sharma-Natu P, Ghildiyal MC (2008) Starch synthase activity and heat shock protein in relation to thermal tolerance of developing wheat grains. Biol Plant 52:749–753. doi:10.1007/s10535-008-0145-x
Suzuki N, Koussevitzky S, Mittler R, Miller G (2012) ROS and redox signalling in the response of plants to abiotic stress. Plant Cell Environ 35:259–270. doi:10.1111/j.1365-3040.2011.02336.x
Suzuki N, Miller G, Morales J, Shulaev V, Torres MA (2011) Respiratory burst oxidases: the engines of ROS signaling. Curr Opin Plant Biol 14:691–699
Talukder ASMHM, McDonald GK, Gill GS (2013) Effect of short-term heat stress prior to flowering and at early grain set on the utilization of water-soluble carbohydrate by wheat genotypes. Field Crop Res 47:1–11
Talukder SK, Babar MA, Vijaylakshmi K, Poland J, Prasad PVV, Bowden R, Fritz A (2014) Mapping QTL for the traits associated with heat tolerance in wheat (Triticum aestivum L.) BMC Genet 15:97. doi:10.1186/s12863-014-0097-4
Tang Y, Xiaoli W, Chaosu L, Chun W, Xiaoling M, Gang H (2013) Long-term effect of year-round tillage patterns on yield and grain quality of wheat. Plant Product Sci 16:365–373
Teixeira EI, Fischer G, Velthuizen HV, Walter C, Ewert F (2013) Global hot-spots of heat stress on agricultural crops due to climate change. Agric For Meteorol 170:206–215. doi:10.1016/j.agrformet.2011.09.002
Trethowan RM, Mujeeb-Kazi A (2008) Novel germplasm resources for improving environmental stress tolerance of hexaploid wheat. Crop Sci 48:1255–1265. doi:10.2135/cropsci2008.02.0082
Tripathi A, Tripathi DK, Chauhan DK, Kumar N, Singh GS (2016) Paradigms of climate change impacts on some major food sources of the world: a review on current knowledge and future prospect. Agric Ecosyst Environ 216:356–373. doi:10.1016/j.agee.2015.09.034
Ugarte C, Calderini DF, Slafer GA (2007) Grain weight and grain number responsiveness to pre-anthesis temperature in wheat, barley and triticale. Field Crops Res 100:240–248. doi:10.1016/j.fcr.2006.07.010
Upreti KK, Sharma M (2016) Role of plant growth regulators in abiotic stress tolerance. In: Rao NKS, Shivashankara KS, Laxman RH (eds.), Abiotic stress physiology of horticultural crops, pp 19-46. doi:10.1007/978-81-322-2725-02
Vijayalakshmi K, Fritz AK, Paulsen GM, Bai G, Pandravada S, Gill BS (2010) Modeling and mapping QTL for senescence-related traits in winter wheat under high temperature. Mol Breed 26:163–175. doi:10.1007/s11032-009-9366-8
Wang H, Wang H, Shao H, Tang X (2016) Recent advances in utilizing transcription factors to improve plant abiotic stress tolerance by transgenic technology. Front Plant Sci 7:67. doi:10.3389/fpls.2016.00067
Wang X, Cai J, Jiang D, Liu F, Dai T, Cao W (2011) Pre-anthesis high-temperature acclimation alleviates damage to the flag leaf caused by post-anthesis heat stress in wheat. J Plant Physiol 168:585–593. doi:10.1016/j.jplph.2010.09.016
Wang X, Cai J, Liu F, Dai T, Cao W, Wollenweber B, Jiang D (2014) Multiple heat priming enhances thermo-tolerance to a later high temperature stress via improving subcellular antioxidant activities in wheat seedlings. Plant Physio Biochem 74:185–192. doi:10.1016/j.plaphy.2013.11.014
Wang X, Cai J, Liu F, Jin M, Yu H, Jiang D, Wollenweber B, Dai T, Cao W (2012) Pre-anthesis high temperature acclimation alleviates the negative effects of post-anthesis heat stress on stem stored carbohydrates remobilization and grain starch accumulation in wheat. J Cereal Sci 55:331–336. doi:10.1016/j.jcs.2012.01.004
Waraich EA, Ahmad R, Ashraf MY, Saifullah AM (2011) Improving agricultural water use efficiency by nutrient management in crop plants. Acta Agric Scand, Section B Plant Soil Sci 61:291–304. doi:10.1080/09064710.2010.491954
Waraich EA, Ahmad R, Halim A, Aziz T (2012) Alleviation of temperature stress by nutrient management in crop plants: a review. J Soil Sci Plant Nutr 12:221–244. doi:10.4067/S0718-95162012000200003
Wheeler T, Von Braun J (2013) Climate change impacts on global food security. Science 341:508–513. doi:10.1126/science.1239402
Yamamoto Y, Aminaka R, Yoshioka M, Khatoon M, Komayama K, Takenaka D, Yamashita A, Nijo N, Inagawa K, Morita N, Sasaki T, Yamamoto Y (2008) Quality control of photosystem II: impact of light and heat stresses. Photosynth Res 98:589–608. doi:10.1007/s11120-008-9372-4
Yang J, Kloepper JW, Ryu CM (2009) Rhizosphere bacteria help plants tolerate abiotic stress. Trends Plant Sci 14:1–4. doi:10.1016/j.tplants.2008.10.004
Yin XY, Guo W, Spiertz JH (2009) A quantitative approach to characterize sink-source relationships during grain filling in contrasting wheat genotypes. Field Crops Res 114:119–126. doi:10.1016/j.fcr.2009.07.013
Yu Q, Li L, Luo Q, Eamus D, Xu S, Chen C, Wang E, Liu J, Nielsen DC (2014) Year patterns of climate impact on wheat yields. Int J Climatol 34:518–528. doi:10.1002/joc.3704
Zhao H, Dai TB, Jing Q, Jiang D, Cao WX (2007) Leaf senescence and grain filling affected by post-anthesis high temperatures in two different wheat cultivars. Plant Growth Regul 51:149–158. doi:10.1007/s10725-006-9157-8
Zheng B, Chenu K, Dreccer MF, Chapman SC (2012) Breeding for the future: what are the potential impacts of future frost and heat events on sowing and flowering time requirements for Australian bread wheat (Triticum aestivium) varieties? Glob Chang Biol 18:2899–2914. doi:10.1111/j.1365-2486.2012.02724.x
Author information
Authors and Affiliations
Corresponding author
About this article

Cite this article
Akter, N., Rafiqul Islam, M. Heat stress effects and management in wheat. A review. Agron. Sustain. Dev. 37, 37 (2017). https://doi.org/10.1007/s13593-017-0443-9
Accepted:
Published:
DOI: https://doi.org/10.1007/s13593-017-0443-9