
Abstract
Antibodies to blood-stage antigens of Plasmodium falciparum play a pivotal role in human immunity to malaria. During parasite development, multiple proteins are trafficked from the intracellular parasite to the surface of P. falciparum-infected erythrocytes (IEs). However, the relative importance of different proteins as targets of acquired antibodies, and key pathways involved in trafficking major antigens remain to be clearly defined. We quantified antibodies to surface antigens among children, adults, and pregnant women from different malaria-exposed regions. We quantified the importance of antigens as antibody targets using genetically engineered P. falciparum with modified surface antigen expression. Genetic deletion of the trafficking protein skeleton-binding protein-1 (SBP1), which is involved in trafficking the surface antigen PfEMP1, led to a dramatic reduction in antibody recognition of IEs and the ability of human antibodies to promote opsonic phagocytosis of IEs, a key mechanism of parasite clearance. The great majority of antibody epitopes on the IE surface were SBP1-dependent. This was demonstrated using parasite isolates with different genetic or phenotypic backgrounds, and among antibodies from children, adults, and pregnant women in different populations. Comparisons of antibody reactivity to parasite isolates with SBP1 deletion or inhibited PfEMP1 expression suggest that PfEMP1 is the dominant target of acquired human antibodies, and that other P. falciparum IE surface proteins are minor targets. These results establish SBP1 as part of a critical pathway for the trafficking of major surface antigens targeted by human immunity, and have key implications for vaccine development, and quantifying immunity in populations.
Similar content being viewed by others

Introduction
Malaria caused by Plasmodium falciparum remains a major cause of morbidity and mortality worldwide [1]. Clinical symptoms result from the blood-stage replication of the parasite, which multiplies within erythrocytes. After erythrocyte invasion by the merozoite form, the intraerythrocytic parasite undergoes development through ring, trophozoite and schizont stages before rupturing to release merozoites to perpetuate the asexual cycle. P. falciparum-induced modifications of infected erythrocytes (IEs) (reviewed in [2, 3]) include the appearance of knob structures on the IE membrane, created by the deposition of knob-associated histidine-rich protein (KAHRP) molecules on the cytoplasmic face of the erythrocyte membrane. These knob structures enable the adhesion of IEs to vascular receptors and sequestration of IEs to prevent clearance by the spleen. Plasmodium falciparum erythrocyte membrane protein 1 (PfEMP1), which is presented on the IE surface by the knob structures, is a key ligand for IE sequestration in the vasculature [4], and is not found other human malaria species. A central feature in the pathogenesis of P. falciparum malaria is the sequestration of IEs in the vasculature of various organs, which is not prominent in other human malarias such as P. vivax, P. knowlesi, P. ovale, P. malariae.
Plasmodium falciparum IE surface antigens are important targets of acquired antibodies that contribute to immunity and function by opsonizing IEs for phagocytic clearance by monocytes, macrophages and other cells, and by inhibiting vascular adhesion and rosette formation (reviewed in [5]). These antigens are polymorphic, and antibodies that are acquired after exposure to infection show a degree of strain specificity [6, 7]. Antibodies to IE surface antigens have been associated with protection in longitudinal studies (reviewed in [5]) and effective immunity is thought to require the acquisition of a broad repertoire of antibodies to different P. falciparum strains or variants [6, 8]. Efforts to define the nature and importance of IE surface antigens and to quantify which antigens are targeted by naturally acquired immunity have been challenging due to the many IE surface proteins identified, the complexity of multigene families encoding these antigens, and limited tools available. In addition to PfEMP1, other antigens identified on the IE surface include repetitive interspersed family proteins (RIFIN) [9], subtelomeric variable open reading frame proteins (STEVOR) [10–12], surface-associated interspersed gene family proteins (SURFIN) [13, 14] and others. These proteins are encoded by multigene families with variation in expression between isolates, which has presented many challenges to studying them as immune targets. Recent approaches to study immune responses using genetically modified parasites, in a Kenyan coastal population, identified PfEMP1 as a major target of acquired human antibodies [15]. However, the relative importance of PfEMP1 and the significance of other IE surface proteins remain unclear.
The trafficking of antigens to the IE surface is only partly understood, and further detailed knowledge is important for defining and quantifying key targets of protective immunity. Since erythrocytes are devoid of protein trafficking machinery and de novo protein synthesis, P. falciparum has to synthesize and export parasite proteins for trafficking, as well as for parasite development (reviewed in [16]). Several P. falciparum proteins are exported into the host erythrocyte to support the correct trafficking and surface display of proteins in the erythrocyte membrane (reviewed in [2, 5]). The export of PfEMP1, and other IE surface proteins, is highly complex due to their large size, the number of membranes that must be traversed before reaching the IE surface and the involvement of various chaperone proteins (reviewed in [16–18]). Skeleton-binding protein 1 (SBP1) [19] is an important protein of membranous structures in the IE cytosol called Maurer’s clefts (MC) and is required for the loading of PfEMP1 molecules into the MC [20, 21]. Although the role of SBP1 in trafficking PfEMP1 has been established, its role in trafficking other IE surface antigens or membrane-bound proteins is unclear. Orthologues of SBP1 have not yet been reported in other Plasmodium species causing human malaria, but has been identified in the chimpanzee malaria, P. reichenowi, which is a close relative of P. falciparum and also expresses PfEMP1 orthologues [20–22]. This suggests that SBP1 has evolved to export PfEMP1 and possibly other IE surface antigens in P. falciparum, and its relative, P. reichenowi [20, 21]. Transcription of pfsbp1 commences shortly after erythrocyte invasion and persists throughout parasite development [19]. SBP1 is a type 2 integral membrane protein (48 kDa) with the N-terminal domain buried within the MC and the C-terminus exposed within the IE cytoplasm where it interacts with erythrocyte proteins such as LANCL1, spectrin and protein 4.1 [19, 23, 24].
In this study, we aimed to quantify the importance of SBP1 in the trafficking of IE surface antigens that are important targets of protective immunity in P. falciparum and evaluate the importance of PfEMP1 and other IE surface antigens as targets of naturally acquired antibodies, including functional antibodies that mediate the opsonic phagocytosis of IEs, which is an important mechanism for clearance of infection [25, 26]. These aims were achieved by using parasites with a targeted deletion of pfsbp1 or genetically engineered to inhibit var gene and PfEMP1 expression and evaluating antibodies to parental parasites compared to genetically modified parasites. We applied these tools to human studies in Kenya, Malawi and Papua New Guinea (PNG), and included serum samples from children, adults and pregnant women to represent the spectrum of malaria exposure and immunity observed globally.
Results
Generation and characterization of P. falciparum with SBP1 deletion
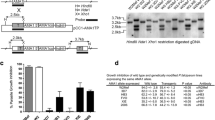
Isolates 3D7 SBP1KO (SBP1 ‘knock-out’) and CS2 SBP1KO have been previously generated and characterized [20, 21]; the transport of PfEMP1 is arrested at MCs or the parasitophorous vacuole, knob formation occurs normally, but IE adhesion by SBP1KO was substantially reduced [20, 21]. In this study, we generated an additional parasite isolate, E8B SBP1KO. E8B parental [27], repeatedly selected for adhesion to immobilized intercellular adhesion molecule-1 (ICAM-1), was transfected with the pHHT-TK-ΔSBP1 plasmid and human-dihydrofolate reductase (DHFR) cassette [20], which encodes resistance to WR99210, and then cycled for integration. Southern blot analyses of the pfsbp1 locus in E8B showed the predicted band sizes of the parental and disrupted loci containing the WR99210 resistance gene hDHFR inserted by double crossover recombination (Fig. 1a). Western blot analyses of IE membrane extracts from pigmented trophozoites confirmed the loss of SBP1 in E8B SBP1KO compared to E8B parental which had a band at approximately 50 kDa (Fig. 1b). Northern blot analysis showed the active transcription of var genes in E8B SBP1KO that were of similar size to E8B parental (Fig. 1c), which suggests that there has not been a switch in var gene expression, consistent with previous findings with SBP1KO parasites [20]. Pigmented trophozoites of E8B SBP1KO had markedly reduced adhesion, with almost no binding to immobilized ICAM-1 (Fig. 1d) and CD36 (Fig. 1e), consistent with the loss of PfEMP1 surface expression. It was shown in previous studies, with repeated failure to select SBP1KO parasites for cytoadhesion, that this phenotype is not due to var gene switching [20].

Phenotypic analyses of pfsbp1 knock-out (SBP1KO) parasites. a Southern blot analysis of the pfsbp1 locus in E8B parental and E8B SBP1KO parasites. Genomic DNA was digested with EcoRI/Bgl II and probed with the 5′ or 3′ targeting sequence. Predicted sizes for the plasmid were 6.9 kb (for both 5′ and 3′ probes) and 4.6 kb for E8B parental (for both 5′ and 3′ probes); predicted sizes for E8B SBP1KO were 2.7 kb (for 5′ probe) and 1.1 kb (for 3′ probe). b Western blot analyses of membrane extracts from mature trophozoite IEs probed with α-SBP1 antibodies. Antibodies detected SBP1 at approximately 50 kDa (arrow) in E8B parental, E8Bvpkd (PfEMP1 knock-down) and E8B-ICAM1 (parental line selected on ICAM1), which was absent in E8B SBP1KO. The band at 100 kDa present in all lanes corresponds to antibody cross reactivity. c Northern blot analysis of var gene transcription by hybridization with a specific var exon 2 sequence. The position of molecular weight standards (kb) is indicated on the left and the arrow represents var transcripts. In both E8B parental and E8B SBP1KO parasites, similar var transcripts were detected. RNA was extracted from highly synchronous ring stage IEs at approximately 10 h post-invasion. Adhesion of IEs to immobilized ICAM-1 (d) and CD36 (e) was substantially reduced in E8B SBP1KO compared to E8B parental. Values are expressed as a percentage of parental parasites binding to ICAM-1 or CD36. Assays were performed twice independently and bars represent median and interquartile ranges of samples tested in triplicate
To further characterize the different SBP1KO parasites, it was important to investigate whether the deletion of pfsbp1 had affected the trafficking and expression of other IE membrane proteins. 3D7 SBP1KO and CS2 SBP1KO were previously reported to retain the typical expression of knobs similar to parental parasites, suggesting that KAHRP expression was unaffected [20, 21]. Here, scanning electron microscopy of pigmented trophozoite-IEs confirmed the expression of knobs by E8B SBP1KO similar to those of E8B parental (Fig. 2a). Additionally, RIFIN and STEVOR proteins localized to the IE membrane remain expressed by both E8B SBP1KO (Fig. 2) and 3D7 SBP1KO (Supplemental Fig S1), similar to their respective parental parasites. Immunofluorescence microscopy with anti-RIF40 antibodies labeled the IE membrane of E8B (Fig. 2b) and 3D7 mature trophozoites (Supplemental Fig S1A). Anti-STEVOR10 antibodies also labeled the IE membrane of mature trophozoites from E8B parental and SBP1KO (Fig. 2c) and 3D7 parental and SBP1KO isolates (Supplemental Fig S1B) by immunofluorescence microscopy. There was no labeling of uninfected erythrocytes by anti-RIF40 or anti-STEVOR10 antibodies and cells were fixed prior to antibody incubations. Furthermore, anti-RIF29 and anti-STEVOR3 antibodies labeled proteins of expected molecular size (~30–40 kDa) in Western blots of IE membrane extracts from E8B (Fig. 2d) and 3D7 mature trophozoites (Supplemental Fig S1C). Immunofluorescence microscopy also confirmed that another IE membrane protein, PfEMP3, remained expressed by E8B SBP1KO (Fig. 2e) and 3D7 SBP1KO (Supplemental Fig S1D), similar to parental parasites. We obtained further evidence that RIFINs remain exposed on the IE surface of the SBP1KO parasites. We examined degradation of RIFINs following treatment of intact IEs with high concentrations of trypsin (1 mg/ml for 30 min), which has been reported to cleave RIFINs [28–30]. Western blotting of IE extracts from pigmented trophozoites with anti-RIF40.2 antibodies demonstrated that trypsin treatment of intact 3D7 parental and 3D7 SBP1KO IEs led to a reduction in the labeling of RIFINs, consistent with the cleavage of RIFINs on the IE surface by trypsin (Fig. 2f). Similarly, trypsin treatment of intact E8B parental and E8B SBP1KO led to a reduction of RIFINs labeled on western blots, suggesting cleavage of surface-exposed RIFIN proteins (Fig. 2f). These findings suggest that the trafficking and expression of several IE membrane proteins other than PfEMP1 occurred normally in the SBP1KO parasites.

Ultrastructural features of the IE membrane and expression of IE membrane proteins by E8B parasites. a Scanning electron microscopy confirmed the expression of surface knob protrusions (arrows) with IEs from E8B SBP1KO similar to that of E8B parental. Representative images are shown and scale bar represents 2 μm in all images. Labeling of RIFIN (b) and STEVOR (c) proteins expressed by pigmented trophozoite-IEs were visualized by immunofluorescence microscopy using specific α-RIF40 and α-STEVOR10 antibodies (green). Despite the lack of PfEMP1 expression, RIFIN and STEVOR were detected in E8B SBP1KO (lower panel), similar to E8B parental (upper panel). The pattern of staining was consistent with the reported labeling of RIFIN and STEVOR in the IE membrane. d Western blot analyses of pigmented trophozoite-IEs membrane extracts probed with α-RIF29 (left) and α-STEVOR3 (right) antibodies, which detected RIFIN and STEVOR in both E8B parental and E8B SBP1KO parasites. Equal loading of parasite material was confirmed with α-HSP70 antibodies (bottom panel). e Pigmented trophozoite-IEs of E8B parental and E8B SBP1KO were probed with α-PfEMP3 antibodies (first column) as a positive control and α-AMA1 antibodies (second column) as a negative control (antibody staining in green, DAPI staining in blue). As expected, the pattern of staining was consistent with the labeling of PfEMP3 in the IE membrane and there was no apparent labeling by anti-AMA1. In all immunofluorescence assays, cells were fixed with a mixture of acetone (90 %) and methanol (10 %) and DAPI was used to stain nuclear DNA (blue). All images were taken at equal exposure for both parasite lines and representative images are shown. f Western blot analyses of protein extracts from purified pigmented trophozoite-IEs (parental and SBP1KO) that were treated with trypsin (+) or control (−). Blots were probed with α-RIF40.2, α-aldolase and α-SBP1 for 3D7 (left) and E8B (right) parasites. There was a reduction in the labeling of RIFINs after trypsin treatment of live IEs. Equal loading of parasite material was confirmed with α-aldolase. α-SBP1 confirmed the absence of SBP1 protein in the SBP1KO parasites and showed that the erythrocyte plasma membrane remained intact during trypsin treatment
Antibodies to the IE surface predominantly target antigens dependent on SBP1 for trafficking
To understand the significance of SBP1 in the trafficking of IE surface antigens that are targeted by naturally acquired antibodies, samples from malaria-exposed individuals were measured by an established flow cytometry-based assay [15] against parental and SBP1KO parasites. The levels of acquired antibodies targeting native antigens expressed on the IE surface were quantified.
Papua New Guinea (PNG)
We tested a selection of serum samples (n = 87) from malaria-exposed adults (n = 46) and children (n = 41) residing in Madang, PNG for antibodies to the surface of erythrocytes infected with 3D7 parental and 3D7 SBP1KO parasites. The overall IgG binding to 3D7 SBP1KO was substantially reduced compared to 3D7 parental (Fig. 3a; p < 0.0001). Sera from most individuals showed a marked reduction in IgG binding to 3D7 SBP1KO compared to 3D7 parental (Fig. 3b). Of those samples that were considered positive for IgG binding (defined as antibody levels greater than mean + 3SD of negative controls) to 3D7 parental (81/87; 93.1 %), there was an 85.6 % reduction in median IgG binding to 3D7 SBP1KO (p = 0.01), suggesting that most antibodies are directed at SBP1-dependent antigens expressed on the IE surface (reflected by the level of IgG binding to 3D7 parental minus 3D7 SBP1KO). To further understand the role of SBP1 in the transport of IE surface antigens, we used the genetically different parasite isolate, E8B. A selection of plasma samples (n = 91) from the same PNG cohort of adults (n = 50) and children (n = 41) were tested for antibodies to the surface of erythrocytes infected with E8B parental and E8B SBP1KO parasites. The overall IgG binding to E8B SBP1KO was greatly reduced compared to E8B parental (Fig. 3c; p < 0.0001). Most plasma samples showed a substantial decrease in IgG binding to E8B SBP1KO (Fig. 3d). Of those samples that were considered positive for IgG binding to E8B parental (72/91; 79.1 %), there was a 61.3 % reduction in median antibody reactivity to E8B SBP1KO (p = 0.001), further indicating that the majority of the antibody response in this cohort was directed towards IE surface antigens that are dependent on SBP1 for trafficking.

Antibodies among children and adults from PNG to surface antigens expressed by P. falciparum-IEs. IgG binding to the surface of erythrocytes infected with 3D7 SBP1KO (a), E8B SBP1KO (c) and was markedly reduced compared to 3D7 parental and E8B parental parasites, respectively. Assays were performed twice independently; bars represent median and interquartile ranges of samples tested in duplicate (n = 87 for 3D7, n = 91 for E8B); p value was calculated using a paired Wilcoxon signed rank test; IgG binding levels are expressed as geometric mean fluorescence intensity (MFI) for all graphs. A representative selection of serum samples from PNG tested for antibodies to 3D7 SBP1KO (b) and E8B SBP1KO (d). Samples were from malaria-exposed children and adults (P3-P25) (b and d) and non-exposed Melbourne residents (Control). IgG binding to 3D7 SBP1KO and E8B SBP1KO was substantially reduced in most individuals, with adults having a higher proportion of antibody response. There was minimal background reactivity observed among sera from Melbourne residents. Assays were performed twice independently; bars represent mean and range of samples tested in duplicate
Antibody levels to 3D7 parental in adults and children from PNG were largely similar (Supplemental Fig S2A; p = 0.15), suggesting that antibodies to IE surface antigens of 3D7 were acquired at a relatively young age in this population. IgG binding to 3D7 SBP1KO was markedly reduced compared to 3D7 parental for both adults (p < 0.0001) and children (p < 0.0001) (Supplemental Fig S2B), indicating that antibodies targeting SBP1-dependent antigens on the IE surface are highly prevalent in both groups. Adults had markedly higher IgG levels than children to E8B parasites (Supplemental Fig S2C; p = 0.0004). Similar to 3D7, IgG binding to E8B SBP1KO was substantially reduced compared to E8B parental for both adults (p < 0.0001) and children (p < 0.0001) (Supplemental Fig S2D). These results further indicate that naturally acquired human antibodies to the IE surface are primarily directed towards SBP1-dependent antigens, among children and adults.
Kenya
To expand these findings in a geographically different population, serum samples (n = 26) from a selection of adults (n = 5) and children (n = 21) residing in Chonyi, Kenya were tested for antibodies to the surface of erythrocytes infected with 3D7 parental and 3D7 SBP1KO parasites. The overall IgG binding to 3D7 SBP1KO was substantially reduced compared to 3D7 parental (Fig. 4a; p < 0.0001). Most individuals showed a marked reduction in IgG binding to 3D7 SBP1KO (Fig. 4b). All 26 samples were classified as positive for IgG binding to 3D7 parental and there was a 90 % reduction in median antibody reactivity to 3D7 SBP1KO (p = 0.0002). Serum samples (n = 15) from adults residing in Kilifi, Kenya were also tested for antibodies to the surface of erythrocytes infected with E8B parental and E8B SBP1KO. The overall IgG reactivity to E8B SBP1KO was markedly reduced compared to E8B parental (Fig. 4c) (p = 0.01). Sera from most individuals showed a substantial reduction in IgG binding to E8B SBP1KO (Fig. 4d). All 15 samples were classified as positive for IgG binding to E8B parental and there was a 81.9 % reduction in median antibody reactivity to E8B SBP1KO (p = 0.15), suggesting that SBP1 is crucial for the trafficking of major antibody targets.

Antibodies among Kenyan individuals to surface antigens expressed by P. falciparum-IEs. IgG binding to the surface of erythrocytes infected with 3D7 SBP1KO (a) and E8B SBP1KO (c) was markedly reduced compared to 3D7 parental and E8B parental. Assays were performed twice independently; bars represent median and interquartile ranges of samples tested in duplicate (n = 26 for 3D7, n = 15 for E8B); p value was calculated using a paired Wilcoxon signed rank test; IgG binding levels are expressed as geometric mean fluorescence intensity (MFI) for all graphs. A representative selection of serum samples tested for antibodies to 3D7 SBP1KO (b) and E8B SBP1KO (d). Samples were from malaria-exposed individuals residing in Kenya (C3–C25; K2–K11) and non-exposed Melbourne residents (control). IgG binding to 3D7 SBP1KO and E8B SBP1KO was substantially reduced in most individuals. There was minimal background reactivity observed among sera from Melbourne residents. Assays were performed twice independently; bars represent mean and range of samples tested in duplicate
Antibodies to pregnancy-associated surface antigens predominantly target SBP1-dependent antigens
Pregnant women are an additional group at high risk of malaria and are infected with specific P. falciparum variants that evade existing immunity and sequester in the placenta [31, 32]. Serum samples (n = 94) from a cohort of pregnant women residing in PNG were tested for antibodies to the IE surface of the placental-binding isolate CS2 parental and CS2 SBP1KO. This isolate expresses VAR2CSA, a PfEMP1 variant that mediates adhesion to CSA in the placenta [33]. The overall IgG binding to CS2 SBP1KO was substantially reduced (90 %) compared to CS2 parental (Fig. 5a; p < 0.0001). Most individuals showed a marked reduction in IgG binding to CS2 SBP1KO (Fig. 5b). When classified according to gravidity, the majority of samples tested from primigravid (PG; n = 33) and multigravid (MG; n = 30) women showed a marked decrease in antibody response to CS2 SBP1KO compared to CS2 parental (Fig. 5c); 19/33 samples (57.6 %) from PG women were positive for IgG binding to CS2 parental and 12/33 samples (36.4 %) were positive for IgG binding to CS2 SBP1KO (p = 0.09); 23/30 samples (76.7 %) and 15/30 samples (50 %) from MG women were positive for IgG binding to CS2 parental and CS2 SBP1KO, respectively (p = 0.05). Of those samples that were considered positive for IgG binding to CS2 parental, IgG binding to CS2 SBP1KO was reduced by 83.1 % in PG women and 91.2 % in MG women. As expected, the level of antibodies was substantially higher in MG women than PG women (Fig. 5c), consistent with previous reports [31, 34]. The level and prevalence of antibodies to CS2 parasites were substantially lower in men compared to pregnant women (Fig. 5c); 9/31 samples (29 %) from men were classified as positive for IgG binding to CS2 parental whereas none of the samples were classified as positive for IgG binding to CS2 SBP1KO (p = 0.001).

Antibodies among pregnant women and men from PNG and Malawi to surface antigens expressed by P. falciparum-IEs. a Among PNG donors, IgG binding to the surface of erythrocytes infected with CS2 SBP1KO parasites was markedly reduced compared to CS2 parental. Assays were performed twice independently; bars represent median and interquartile ranges of samples tested in duplicate (n = 94); p value was calculated using a paired Wilcoxon signed rank test; IgG binding levels are expressed as geometric mean fluorescence intensity (MFI) for all graphs. b A representative selection of samples from PNG pregnant women tested for antibodies to CS2 parental and CS2 SBP1KO. IgG binding to CS2 SBP1KO was substantially reduced in most individuals. There was minimal background reactivity observed among sera from non-exposed Melbourne residents (Control). Assays were performed twice independently; p value was calculated using a paired Wilcoxon signed rank test; bars represent mean and range of samples tested in duplicate. c. Serum samples from pregnant women in PNG tested for antibodies to CS2 parental and CS2 SBP1KO parasites are grouped by gravidity. Samples were from malaria-exposed primigravid (PG; n = 33) and multigravid (MG; n = 30) women and men (n = 31) in PNG. IgG binding to CS2 SBP1KO parasites was substantially reduced compared to CS2 parental in PG women, MG women and men. There was minimal background reactivity observed among sera from non-exposed Melbourne residents (Control). Assays were performed twice independently; p value was calculated using a paired Wilcoxon signed rank test; bars represent median and interquartile ranges of samples tested in duplicate. d Rabbit antibodies (R1–R2) raised against the DBL3 domain of VAR2CSA were tested for antibody recognition to CS2 parental and CS2 SBP1KO. All samples showed a marked reduction in IgG binding to CS2 SBP1KO. PI represents pre-immune serum from these rabbits. Assays were performed twice independently; bars represent mean and range of samples tested in duplicate. e Among pregnant women from Malawi, IgG binding to the surface of erythrocytes infected with CS2 SBP1KO parasites was markedly reduced compared to CS2 parental. Assay was performed once; bars represent median and interquartile ranges of samples tested in duplicate (n = 81); p value was calculated using a paired Wilcoxon signed rank test; IgG binding levels are expressed as geometric mean fluorescence intensity (MFI) for all graphs. f A representative selection of samples tested for antibodies to CS2 parental and CS2 SBP1KO. Samples were from malaria-exposed pregnant women from Malawi (ML1–ML8) and non-exposed Melbourne residents (Control). IgG binding to CS2 SBP1KO was substantially reduced in most individuals. There was minimal background reactivity observed among sera from non-exposed Melbourne residents (Control). Assay was performed once; p value was calculated using a paired Wilcoxon signed rank test; bars represent mean and range of samples tested in duplicate
We confirmed these findings with serum samples from a cohort of pregnant women (n = 71) and men (n = 10) residing in Malawi. Again, overall IgG binding to CS2 SBP1KO was substantially reduced (76.7 %) compared to CS2 parental (Fig. 5e; p < 0.0001). Most individuals showed a marked reduction in IgG binding to CS2 SBP1KO (Fig. 5f). When classified according to gravidity, the majority of samples tested from primigravid (PG; n = 35) and multigravid (MG; n = 36) women showed a marked decrease in antibody response to CS2 SBP1KO compared to CS2 parental (Supplemental Fig S3A); 22/35 samples (62.9 %) from PG women were positive to CS2 parental and 20/35 samples (57.1 %) were positive to CS2 SBP1KO (p = 0.73); 22/36 samples (61.1 %) from MG women were positive for IgG to CS2 parental while 6/36 samples (16.7 %) were positive for CS2 SBP1KO (p = 0.001). The overall IgG binding to CS2 SBP1KO in samples from men residing in the same region was markedly reduced compared to CS2 parental (Supplemental Fig S3A; p = 0.002). The level and prevalence of antibodies to CS2 parasites were substantially lower in men compared to pregnant women (Supplemental Fig S3A).
To confirm the loss of VAR2CSA-PfEMP1 expression in CS2 SBP1KO, we tested rabbit antibodies raised against the DBL3 domain of VAR2CSA expressed by CS2 [35] for recognition of the IE surface of CS2 parental and CS2 SBP1KO parasites. There was markedly lower IgG binding to CS2 SBP1KO (Fig. 5d), suggesting the absence of PfEMP1 on the IE surface of these parasites and supporting the significance of SBP1 in the trafficking of PfEMP1.
Antibodies that mediate opsonic phagocytosis of IEs predominantly target SBP1-dependent antigens on the IE surface
To examine the importance of SBP1 in the trafficking of IE surface antigens targeted by functional antibodies, serum samples from Kenyan adults were tested for their ability to opsonize pigmented trophozoite-IEs for phagocytic clearance by undifferentiated Thp-1 monocytes, an established model of opsonic phagocytosis [15, 36, 37]. IEs were incubated with serum samples for opsonisation by antibodies before co-incubation with THP-1 monocytes was performed to test for phagocytosis. The ability of serum antibodies (n = 25) to promote opsonic phagocytosis activity was substantially reduced (78.4 %) in 3D7 SBP1KO compared to 3D7 parental (Fig. 6a; p < 0.0001). We also tested serum IgG purified from these Kenyan adults (n = 9) for their opsonic phagocytosis activity with E8B parental and E8B SBP1KO. The level of opsonic phagocytosis was markedly reduced by 77.1 % in E8B SBP1KO compared to E8B parental (Fig. 6c; p = 0.0001). All samples showed a reduction in opsonic phagocytosis activity to 3D7 SBP1KO (Fig. 6b) and E8B SBP1KO (Fig. 6d) compared to their respective parental parasites. Furthermore, we tested a selection of plasma samples (n = 28) from adults and children residing in Madang, PNG for their opsonic phagocytosis activity. Similarly, the ability of serum antibodies to opsonize IEs for phagocytosis was substantially reduced (60.8 %) in E8B SBP1KO compared to E8B parental (Fig. 6e; p < 0.0001) and all samples showed a reduction in opsonic phagocytosis activity to E8B SBP1KO (Fig. 6f) compared to E8B parental. Together, our results suggest that antibodies target SBP1-dependent antigens on the IE surface function to opsonise IEs for phagocytosis.

Opsonic phagocytosis of IEs by undifferentiated Thp-1 monocytes. Opsonic phagocytosis activity of serum antibodies from Kenya and PNG was markedly reduced with 3D7 SBP1KO (a) and E8B SBP1KO (c, e) compared to 3D7 parental and E8B parental. Assays were performed twice independently; bars represent median and interquartile ranges [for Kenyan samples, n = 25 for 3D7 (a), n = 9 for E8B (b); for PNG samples, n = 25 for E8B (c)]; p value was calculated using a paired Wilcoxon signed rank test. For all graphs, the level of opsonic phagocytosis is expressed as a percentage of positive control (rabbit antibody raised against human erythrocytes). A representative selection of serum samples from Kenya tested for opsonic phagocytosis activity to 3D7 SBP1KO (b) and E8B SBP1KO (d) and from PNG tested for opsonic phagocytosis activity to E8B SBP1KO (f). Assays were performed twice independently for 3D7 and once for E8B; bars represent mean and range with samples tested in duplicate
We tested serum samples from pregnant women (n = 36) residing in Malawi for their opsonic phagocytosis activity to CS2 parental and CS2 SBP1KO parasites. The ability of serum antibodies to promote opsonic phagocytosis activity was substantially reduced (76.4 %) in CS2 SBP1KO compared to CS2 parental (Fig. 7a; p < 0.0001). All samples showed a reduction in opsonic phagocytosis activity to CS2 SBP1KO (Fig. 7b) compared to their respective parental parasites, suggesting the functional importance of antibodies to SBP1-dependent antigens.

Opsonic phagocytosis of IEs by undifferentiated Thp-1 monocytes using sera from pregnant women in Malawi. Opsonic phagocytosis activity was markedly reduced in CS2 SBP1KO (a) compared to CS2 parental. Assays were performed twice independently; bars represent median and interquartile ranges (n = 36); p value was calculated using a paired Wilcoxon signed rank test. For all graphs, the level of opsonic phagocytosis is expressed as a percentage of positive control (rabbit antibody raised against human erythrocytes). A representative selection of serum samples tested for opsonic phagocytosis activity to CS2 SBP1KO (b). Samples were from malaria-exposed pregnant women residing in Malawi (ML9–19) and non-exposed Melbourne residents (Control). Assays were performed twice independently; bars represent mean and range with samples tested in duplicate
PfEMP1 is the major target of antibodies to the IE surface
To investigate the identity of SBP1-dependent antigens recognized by human antibodies we used P. falciparum that was genetically engineered to suppress PfEMP1 expression (var promoter knock-down, vpkd). These parasites, which have been previously described and characterized in detail, have inhibited PfEMP1 surface expression through the suppression of endogenous var genes [15, 38]. Antibody levels to the surface of pigmented trophozoite-IEs were compared between parental, vpkd and SBP1KO parasites. In a selection of PNG individuals (n = 12), the overall IgG reactivity to E8Bvpkd and E8B SBP1KO was substantially lower than parental E8B. Reactivity to E8B SBP1KO was lower again than to E8Bvpkd (Fig. 8a; p = 0.008). Compared to E8B parental, IgG binding was reduced by 54.5 % to E8Bvpkd and by 76.5 % to E8B SBP1KO. The majority of the samples showed a marked reduction in IgG binding to E8B SBP1KO compared to E8Bvpkd and E8B parental (Fig. 8b). Similar results were seen using adult sera from Kilifi, Kenya (n = 7). (Figure 8c, d; p = 0.02). Compared to E8B parental, IgG binding was reduced by 79.3 % to E8Bvpkd and by 91.6 % to E8B SBP1KO. Similar results were also seen using 3D7 parental, vpkd and SBP1KO parasites (Supplemental Fig S4). For PNG serum (n = 81) IgG binding was reduced by 79.5 % to 3D7vpkd and by 89 % to 3D7 SBP1KO compared to parental 3D7 (Supplemental Fig S4A, S4B). For samples from Kenya, IgG binding was reduced by 76.7 % to 3D7vpkd and by 90 % to 3D7 SBP1KO compared to 3D7 parental (Supplemental Fig S4C, S4D). This suggests that most of the SBP1-dependent antibody targets are PfEMP1.

PfEMP1 is the major target of human antibodies. The overall IgG binding to E8B SBP1KO parasites was markedly reduced compared to E8B parental parasites and further reduced compared to E8Bvpkd, among samples from PNG (a) and Kenya (c). Assays were performed twice independently; bars represent median and interquartile ranges (n = 12 for PNG, n = 7 for Kenya); p value was calculated using a paired Wilcoxon signed rank test; IgG binding levels are expressed as geometric mean fluorescence intensity (MFI) for all graphs. A representative selection of samples tested in parallel for antibodies to E8B parental, E8Bvpkd and E8B SBP1KO. Samples were from malaria-exposed PNG adults (b; P7–P31), Kenyan adults (d; K3–K18) and non-exposed Melbourne residents (Control). There was minimal background reactivity observed among sera from Melbourne residents. Assays were performed twice independently; bars represent mean and range of samples tested in duplicate. e IgG binding to the IE surface of 3D7 parental and 3D7vpkd was trypsin sensitive compared to 3D7 SBP1KO. IEs were incubated with trypsin at 5 or 10 ug/ml, or without trypsin, and tested for IgG reactivity among samples form Kenyan adults. IgG binding levels are expressed as a percentage of their respective 3D7 parental reactivity without trypsin; bars represent mean and SEM of samples tested in duplicate (n = 5) after subtraction with IgG levels from negative Melbourne controls
To understand the nature of residual antibody reactivity to vpkd IEs, compared to SBP1KO IEs, we evaluated the effect of trypsin treatment of IEs on antibody binding (Fig. 8e). The majority of IgG binding to 3D7 parental surface antigens was highly trypsin sensitive, suggesting that antibodies are targeting PfEMP1 on the IE surface, since PfEMP1 is known to be trypsin sensitive [4]. The residual IgG binding observed with 3D7vpkd parasites was also trypsin sensitive, consistent with PfEMP1 being the target of these antibodies. Treatment of 3D7vpkd with trypsin reduced antibody reactivity to levels that were comparable to 3D7 SBP1KO, suggesting that antibody reactivity to 3D7vpkd may represent residual PfEMP1 expression.
Discussion
Our findings indicate that, remarkably, a single parasite protein, SBP1, is required for the transport of IE surface antigens that are the major targets of human antibodies against P. falciparum infections. SBP1 is clearly a crucial component of the trafficking pathway for surface antigens and may represent an important difference between P. falciparum and other human malaria species. Comparing the level of IgG binding to the IE surface between parental and SBP1KO allowed the quantification of antibodies targeting SBP1-dependent antigens and defined a key property of major surface antigens and epitopes. The overall IgG binding to SBP1KO IEs was dramatically reduced compared to parental parasites, indicating that the great majority of the naturally acquired antibodies to the IE surface targeted SBP1-dependent antigens. The decrease in antibody reactivity to SBP1KO parasites was observed with serum antibodies from children and adults from PNG and Kenya, as well as serum antibodies from pregnant women residing in PNG and Malawi. Furthermore, the level of functional antibodies measured through opsonic phagocytosis activity was greatly reduced in the SBP1KO parasites. Comparisons between antibody reactivity to SBP1KO and PfEMP1 knock-down (vpkd) parasites supported the conclusion that PfEMP1 is the dominant target of human antibodies.
Plasmodium falciparum malaria affects children, adults and pregnant women, and has a high burden of disease in many parts of Africa, Asia, South America and the Pacific. Plasmodium falciparum exposure and the development of naturally acquired immunity can vary considerably between risk groups and geographical regions. Therefore, to ensure generalizability of findings we included samples from all three risk groups (children, adults, pregnant women) that represented different populations from Africa (Kenya and Malawi) and other regions (PNG). Additionally, we used three different parasite isolates, with different genetic or phenotypic backgrounds. Prior studies have shown that parasite isolates such as 3D7 and IT4 lines (E8B) are commonly recognized by serum from malaria-exposed individuals across different geographic regions [27, 39, 40]. Strikingly, the same pattern of substantially reduced antibody binding to SBP1KO parasites was observed in all risk groups and populations with each of our three P. falciparum isolates. These repeated observations provide strong evidence for SBP1-dependent IE surface antigens being predominant antibody targets with minimal targeting of non-SBP1-dependent antigens by human immune responses.
The functional mechanisms of antibodies to IE surface antigens that protect against malaria are not fully understood. Antibodies are believed to contribute to protection against symptomatic disease by opsonising IEs for clearance by monocytes or macrophages [25, 26, 41]. Undifferentiated THP-1 monocytes were used in this study because they specifically measure Fc-mediated opsonic phagocytosis and have very little non-opsonic phagocytosis [36]. Antibody-mediated opsonic phagocytosis was greatly reduced in 3D7 SBP1KO, E8B SBP1KO and CS2 SBP1KO IEs compared to their respective parental parasites, providing key evidence that antibodies to SBP1-dependent epitopes on the IE surface function in the opsonic phagocytosis of IEs. It was interesting that some measurable opsonic phagocytosis activity was still detected in the SBP1KO parasites. Although it was low, it may still contribute to immunity and could potentially be accounted for by antibodies to other IE surface antigens such as RIFIN and STEVOR, which are still expressed by the SBP1KO parasites. An additional important mechanism that is thought to contribute to immunity is antibody inhibition of IE adhesion or rosetting (reviewed in [5]). For example, antibodies from pregnant women sera inhibited the binding of IEs to the placental receptor CSA [34, 42], which have been associated with reduced malaria in pregnancy and placental infection [42, 43]. Furthermore, antibodies from children with mild malaria disrupted rosette formation whereas those with severe malaria did not, suggesting that acquired antibodies mediate protection through inhibition of rosette formation [44, 45]. In our study, and previous studies, we show that disruption of SBP1 leads to loss of cytoadhesion ability and antibody recognition, suggesting that antibodies that inhibit adhesion are targeting SBP1-dependent antigens, particularly PfEMP1. IE surface antigens also have other important immune interactions, such as cellular immunity. Prior studies have reported that PfEMP1 may impact on responses by a range of cell types, including dendritic cells, T cells, and innate immune cells [46–50]. Future studies using SBP1KO parasites to better understand these responses may be valuable for understanding the role of PfEMP1 and other surface antigens in cell-mediated immunity.
Antibodies directed to the IE surface of P. falciparum isolates that are prevalent in pregnancy malaria also represent important components of protective immunity [42]. These antibodies are thought to confer protection by inhibiting parasite sequestration in the placenta [31, 42, 51] and promoting the opsonic phagocytosis of IEs [52–54]. The PfEMP1 variant, VAR2CSA, is an important antibody target and vaccine candidate but other antigens have also been proposed [55]. Reflecting previous exposure to CSA-binding isolates that primarily expresses var2csa [56], multigravid (MG) women generally have a higher prevalence of antibodies to placental isolates compared to primigravid (PG) women or men [31, 34, 57] and the findings in this study are consistent with that. IgG binding to CS2 SBP1KO in both PG and MG women was markedly reduced compared to CS2 parental, demonstrating that surface antigens dependent on SBP1 are major targets of acquired antibodies, a substantial proportion of which is presumably directed at VAR2CSA. In our study, rabbit antibodies raised against VAR2CSA were used to confirm the loss of PfEMP1 surface expression in the CS2 SBP1KO parasites.
The importance of PfEMP1 as a target was demonstrated by comparing the levels of IgG binding between parental, vpkd and SBP1KO parasites. The majority of the loss of reactivity of antibodies to SBP1KO parasites could be explained by a loss of PfEMP1, supporting the conclusion that PfEMP1 is the major target of human antibodies to the IE surface. Interestingly, there was a small further reduction in antibody reactivity to SBP1KO compared to vpkd parasites. This was observed in E8B and 3D7 parasites, and similarly for both PNG and Kenyan populations. The residual antibody binding to vpkd compared to SBP1KO might reflect residual PfEMP1 expression in the vpkd parasites, whereas SBP1KO may have inhibited PfEMP1 surface display more efficiently. Reactivity to 3D7vpkd was largely trypsin sensitive, similar to the sensitivity pattern of 3D7 parental parasites, suggesting that the reactivity may be explained by residual PfEMP1 expression on vpkd parasites. PfEMP1 has been described as being highly sensitive to trypsin treatment [4] compared to other antigens such as RIFIN, that have been reported to be resistant to cleavage by low trypsin concentrations [28, 29].
The low antibody reactivity to SBP1KO parasites suggests that antibodies that may target other antigens that are trafficked to the IE surface independent of SBP1 expression are not a prominent feature of the naturally acquired immune response. Possible antigens include RIFIN, STEVOR or SURFIN families which have been previously identified on the IE surface [12, 13, 28–30, 58, 59]. We found that that the expression of other surface proteins, such as RIFIN and STEVOR, were similar in the parental and SBP1KO parasites, suggesting that these surface antigens are not dependent on SBP1. We also provided evidence that RIFINs were exposed on the IE surface, by showing that trypsin treatment of intact IEs led to degradation of RIFINs. Furthermore, exported proteins required for knob formation were still expressed by the SBP1KO parasites. There is a possibility that the expression levels of RIFINs and STEVORs may have changed in the SBP1KO parasites compared to parental and this may contribute to reduced antibody recognition. One previous study [58] reported that a lower proportion of IEs stained positive with STEVOR antibodies (raised against a STEVOR peptide) for 3 lab isolates compared to 4 recently adapted isolates. Currently we lack the tools to quantify the expression of these other antigens on the IE surface of parental and SBP1KO parasites, but there did not appear to be a major difference in expression between parental and KO parasites using western blots and immunofluorescence microscopy, although these methods are not highly quantitative. Rapidly generating SBP1KO clones with clinical isolates is currently not possible. It is unlikely that our findings of reduced antibody recognition of SBP1KO IEs can be explained by the switching of var genes and PfEMP1 variants expressed between parental and SBP1KO parasites as Northern blots showed bands of similar sizes in both parental and SBP1KO parasites, and the disruption of surface-exposed PfEMP1 has been confirmed in three independent parasite isolates. Furthermore, the immune serum used in our assays is not variant-specific and should detect a wide repertoire of PfEMP1 variants, should they be expressed by the SBP1KO parasites. In addition, previous studies have been unable to select the SBP1KO parasites for adhesion to immobilized receptors, thus suggesting that the parasites have not switched to another PfEMP1 variant [20, 21].
These findings have important translational implications for understanding and measuring immunity, and developing interventions against malaria. A strong knowledge of the targets of mechanisms of immunity is crucial for the development of effective vaccines, and our findings further highlight the significance of PfEMP1 as the major target of acquired human antibodies. While PfEMP1 is a known target of immunity, it is important to quantify its role relative to other antigens in order to prioritize antigens for vaccine development, and develop tools to quantify and monitor human immunity, which are currently lacking. There is significant interest in PfEMP1-based vaccines [60–62] and vaccines based on the VAR2CSA-PfEMP1 are progressing to clinical trials. We also demonstrate the value of SBP1KO parasites as a tool to measure and evaluate antibodies to surface antigens, particularly PfEMP1, in populations and potentially in vaccine trials. Unlike other tools, such as isolates with suppressed PfEMP1 expression [15, 38], pfsbp1 gene-KO parasites are phenotypically stable in culture and can be readily applied in different assays and laboratories. Approaches to developing highly efficacious second generation vaccines include the potential use of attenuated whole parasites, including attenuated IEs [63, 64]. Therefore, it is important to define key virulence features and antibody targets. Moreover, our studies highlight a crucially important evolutionary difference between P. falciparum and P. vivax, which has major implications for understanding immunity and disease pathogenesis and the development of future vaccine and therapeutic strategies against these pathogens.
In conclusion, our study has provided major new evidence that the key targets of acquired human antibodies are highly dependent on SBP1 for trafficking to the IE surface. Moreover, finding that PfEMP1 is by far the major target of human antibodies to the IE surface supports efforts to develop vaccines based on this antigen. We also showed that antibodies to SBP1-dependent antigens, predominantly PfEMP1, play a key role in a mechanism of parasite clearance by opsonizing IEs for phagocytosis. These findings highlight an important role for SBP1 in the trafficking of IE surface antigens and define a key feature in the biology and virulence properties that may be different to other human malarias. These findings have major implications for malaria vaccine development, and for quantifying immunity in populations.
Methods
Study population and ethics statement
Serum samples (n = 100) were collected from a community-based cross-sectional study in the Madang Province, PNG [65]. This population experiences year-round malaria transmission with some seasonal variation [66]. Half of these samples were from children with a median age of 7 years (interquartile range, 6–9 years), while the other half were from adults with a median age of 28 years (interquartile range 24–35). For 9 subjects, there was insufficient serum available for inclusion in this study; therefore, a total of 91 subjects were included. Serum samples were also collected from a cross-sectional study of pregnant women in their second trimester (n = 63) who visited the antenatal clinic for routine care, and from men (n = 31), at the Yagaum Health Centre and Modilon Hospital in Madang Province, PNG as previously described [67, 68]. Of these, 33 samples from PGs and 30 samples from MG women were available for testing. In Malawi, samples were collected from pregnant women (n = 85) as described [69]. In Kenya, serum samples (n = 25) were also collected from anonymous adults at the Hospital Blood Donor Service in Kilifi, [15, 70] and from individuals participating in a longitudinal cohort study (n = 296) conducted in Chonyi, Kilifi district as described elsewhere [15, 70, 71]. Ethics approval was obtained from the Kenya Medical Research Institute Ethics Committee, the Medical Research Advisory Council PNG, the College of Medicine Research Ethics Committee in Malawi, the Walter and Eliza Hall Institute Human Research and Ethics Committee and the Alfred Hospital Human Research and Ethics Committee. Written informed consent was obtained from all study participants or their parents or legal guardians.
Plasmodium falciparum culture and isolates
Plasmodium falciparum isolates were maintained in continuous culture and synchronized as previously described [15]. Isolate E8B SBP1KO was generated by transfecting E8B parental [27], derived from a clone of IT4, with the pHHT-TK-ΔSBP1 plasmid and culturing in the presence of WR99210 (2.5 nM), using established methods [72]. Isolates 3D7 SBP1KO and CS2 SBP1KO were generated as previously described [20, 21]. Isolates 3D7vpkd and E8Bvpkd were generated as previously described [15, 38]. IEs were typically used in assays at the pigmented trophozoite developmental stage when they express the surface proteins PfEMP1, RIFIN, STEVOR, SURFIN (except ring stage parasites were used for transcript analysis by northern blotting).
Measuring antibodies to the IE surface by flow cytometry
Measuring IgG binding to the IE surface of pigmented trophozoites was performed with an established flow cytometry-based assay as previously described [15]. IgG binding levels for each sample was expressed as the geometric mean fluorescence intensity (MFI; arbitrary units) for IEs, after subtracting the MFI of uninfected erythrocytes. Samples were tested at a 1/10 dilution and were designated antibody positive if the MFI was greater than the mean plus 3 standard deviation of reactivity seen with sera from non-malaria exposed Australian residents. Cryopreserved pigmented trophozoite IEs used in antibody assays have been previously validated (Beeson et al. unpublished) [15, 73]. Cryopreservation of IEs was performed as described [15]. IEs were incubated with various concentrations of trypsin (0–10 μg/mL; TPCK-treated, Worthington Biomedical) diluted in PBS for 15 min at 37 °C, as previously described [15].
Western blots
Western blots of membrane proteins from pigmented trophozoite-IEs were performed using Triton-X 100-insoluble, SDS-soluble protein extracts of pigmented trophozoite-IEs as previously described [15]. Nitrocellulose membranes (Invitrogen) were probed with affinity-purified rat antibodies against recombinant RIF29 (1/2000) [74, 75], rat antibodies against RIF40.2 (1/2000) [30], affinity-purified mouse antibodies against recombinant STEVOR3 protein (1/2000) [76], rabbit anti-SBP1 (1/1000) [20], rabbit anti-HSP70 antibodies (1/500; provided by Paul Gilson) and rabbit anti-aldolase (1/5000; provided by Jake Baum). Magnet-purified IEs were incubated with trypsin at 1 mg/mL (TPCK-treated, Sigma–Aldrich) or PBS as a negative control for 30 min at 37 °C, and the reaction stopped with trypsin inhibitor (Sigma–Aldrich) before the cells were extracted with SDS loading buffer, as previously described [30].
Northern blots
RNA from highly synchronous ring-staged parasite cultures at 10 h post-invasion was extracted using Trizol (Invitrogen) as previously described [15]. Northern blots were hybridized with a conserved var exon 2 sequence (PlasmoDB accession number PFIT_1219000) and washed at low stringency with 2X SSC and 0.1 % SDS at 55 °C [77].
Plasmodium falciparum cytoadherence assays
Testing for adhesion to immobilized receptors was performed using gelatin-enriched pigmented trophozoite-IEs at 15–20 % parasitemia as previously described [15]. The numbers of bound cells were recorded as IEs per square millimeter and PBS was used as a negative control to record non-specific binding. Adhesion to each receptor was tested in triplicate at these concentrations: ICAM-1 10 μg/mL (Bender MedSystems) and CD36 20 μg/mL (rhCD36/Fc chimera, R&D systems).
Immunofluorescence microscopy
Immunofluorescence microscopy of pigmented trophozoite-IEs were performed as previously described [15]. Slides were incubated with primary antibodies such as anti-ARIF40 (1/50; accession number AF483820) [75], anti-STEVOR10 (1/200) [76], anti-SBP1 (1/500) [20], anti-PfEMP3 (1/500; provided by Melanie Rug) and anti-AMA1 (apical membrane antigen 1; 1/500) [78], followed by the corresponding Alexa Fluor 488-conjugated IgG (1/500, Invitrogen). Images were obtained using a Plan-Apochromat (100 ×/1.40) oil immersion phase-contrast lens (Carl Zeiss) on an AxioVert 200 M microscope (Carl Zeiss) equipped with an AxioCam Mrm camera (Carl Zeiss).
Opsonic phagocytosis assays
Measuring the level of opsonic phagocytosis activity using undifferentiated Thp-1 monocytes was conducted as previously described [15, 36]. The proportion of Thp-1 positive cells with phagocytosed pigmented trophozoite-IEs was counted by flow cytometry (FACSCalibur, BD Biosciences). The level of phagocytosis for each sample was expressed relative to the positive control (Abcam; rabbit antibody raised against human erythrocytes). Negative control samples from non-malaria exposed Australian residents were included in all assays. Purification of IgG was performed using Melon Gel IgG purification kit (Thermo Scientific) according to the manufacturer’s instructions and dialysed into PBS prior to performing antibody assays [79].
Scanning electron microscopy
Square glass coverslips (22 mm) were prepared by smearing a 0.1 % solution of polyethyleneimine (PEI) and dried. Pigmented trophozoite-IE samples were incubated on PEI-coated glass coverslips for 30 min and then immersed in 2 % glutaraldehyde in PBS for 1 h. Coverslips were rinsed thrice in PBS for 10 min before being dehydrated in increasing concentrations of ethanol consisting of 10, 20, 40, 60, 80, and 100 % ethanol in water for 10 min each. The coverslips were then dried in a Balzers CPD 030 Critical Point Dryer (Balzers Pfeiffer, Balzers, Liechtenstein) and mounted onto 25 mm aluminum stubs with double-sided carbon tabs and then gold-coated in a Dynavac “Xenosput” magnetron sputter coater. Cells were imaged with the Philips XL30 field emission scanning electron microscope (Philips, Eindhoven, Netherlands) at a voltage of 2 kV.
Statistical analyses
Non-parametric analytical methods were used to evaluate assay results from cohort studies, as the data obtained were not normally distributed. Differences in adhesion levels between parental and genetically modified parasites were assessed using an unpaired Mann–Whitney test. Differences in antibody levels between parental and genetically modified parasites were assessed using a paired Wilcoxon signed rank test or Kruskal–Wallis test (>2 groups). Statistical analyses were performed using Prism version 5 (GraphPad Software Inc) or Stata 12.0 (StataCorp).
References
World Health Organization (2014) Report 2014. 1–142
Maier AG, Cooke BM, Cowman AF, Tilley L (2009) Malaria parasite proteins that remodel the host erythrocyte. Nat Rev Micro 7:341–354. doi:10.1038/nrmicro2110
Garcia CRS, de Azevedo MF, Wunderlich G et al (2008) Plasmodium in the postgenomic era: new insights into the molecular cell biology of malaria parasites. Int Rev Cell Mol Biol 266:85–156. doi:10.1016/S1937-6448(07)66003-1
Leech JH, Barnwell JW, Miller LH, Howard RJ (1984) Identification of a strain-specific malarial antigen exposed on the surface of Plasmodium falciparum-infected erythrocytes. J Exp Med 159:1567–1575
Chan J-A, Fowkes FJI, Beeson JG (2014) Surface antigens of Plasmodium falciparum-infected erythrocytes as immune targets and malaria vaccine candidates. Cell Mol Life Sci. doi:10.1007/s00018-014-1614-3
Marsh K, Howard RJ (1986) Antigens induced on erythrocytes by P. falciparum: expression of diverse and conserved determinants. Science 231:150–153
Reeder JC, Rogerson SJ, al-Yaman F et al (1994) Diversity of agglutinating phenotype, cytoadherence, and rosette-forming characteristics of Plasmodium falciparum isolates from Papua New Guinean children. Am J Trop Med Hyg 51:45–55
Bull PC, Lowe BS, Kortok M, Marsh K (1999) Antibody recognition of Plasmodium falciparum erythrocyte surface antigens in Kenya: evidence for rare and prevalent variants. Infect Immun 67:733–739
Khattab A, Klinkert M-Q (2006) Maurer’s clefts-restricted localization, orientation and export of a Plasmodium falciparum RIFIN. Traffic 7:1654–1665. doi:10.1111/j.1600-0854.2006.00494.x
Kaviratne M, Khan SM, Jarra W, Preiser PR (2002) Small variant STEVOR antigen is uniquely located within Maurer’s clefts in Plasmodium falciparum-infected red blood cells. Eukaryot Cell 1:926–935
Niang M, Bei AK, Madnani KG et al (2014) STEVOR Is a Plasmodium falciparum erythrocyte binding protein that mediates merozoite invasion and rosetting. Cell Host Microbe 16:81–93. doi:10.1016/j.chom.2014.06.004
Niang M, Yam XY, Preiser PR (2009) The Plasmodium falciparum STEVOR multigene family mediates antigenic variation of the infected erythrocyte. PLoS Pathogol 5:e1000307
Winter G, Kawai S, Haeggström M et al (2005) SURFIN is a polymorphic antigen expressed on Plasmodium falciparum merozoites and infected erythrocytes. J Exp Med 201:1853–1863. doi:10.1084/jem.20041392
Mphande FA, Ribacke U, Kaneko O et al (2008) SURFIN4.1, a schizont-merozoite associated protein in the SURFIN family of Plasmodium falciparum. Malar J 7:116. doi:10.1186/1475-2875-7-116
Chan J-A, Howell KB, Reiling L et al (2012) Targets of antibodies against Plasmodium falciparum-infected erythrocytes in malaria immunity. J Clin Invest. doi:10.1172/JCI62182
Elsworth B, Crabb BS, Gilson PR (2014) Protein export in malaria parasites: an update. Cell Microbiol 16:355–363. doi:10.1111/cmi.12261
Boddey JA, Cowman AF (2013) Plasmodium nesting: remaking the erythrocyte from the inside out. Annu Rev Microbiol 67:243–269. doi:10.1146/annurev-micro-092412-155730
Deponte M, Hoppe HC, Lee MCS et al (2012) Wherever I may roam: protein and membrane trafficking in P. falciparum-infected red blood cells. Mol Biochem Parasitol 186:95–116. doi:10.1016/j.molbiopara.2012.09.007
Blisnick T, Morales Betoulle ME, Barale JC et al (2000) Pfsbp1, a Maurer’s cleft Plasmodium falciparum protein, is associated with the erythrocyte skeleton. Mol Biochem Parasitol 111:107–121
Maier AG, Rug M, O’Neill MT et al (2007) Skeleton-binding protein 1 functions at the parasitophorous vacuole membrane to traffic PfEMP1 to the Plasmodium falciparum-infected erythrocyte surface. Blood 109:1289–1297. doi:10.1182/blood-2006-08-043364
Cooke BM, Buckingham DW, Glenister FK et al (2006) A Maurer’s cleft-associated protein is essential for expression of the major malaria virulence antigen on the surface of infected red blood cells. J Cell Biol 172:899–908. doi:10.1083/jcb.200509122
Otto TD, Rayner JC, Böhme U et al (2014) Genome sequencing of chimpanzee malaria parasites reveals possible pathways of adaptation to human hosts. Nat Commun 5:4754. doi:10.1038/ncomms5754
Blisnick T, Vincensini L, Barale JC et al (2005) LANCL1, an erythrocyte protein recruited to the Maurer’s clefts during Plasmodium falciparum development. Mol Biochem Parasitol 141:39–47. doi:10.1016/j.molbiopara.2005.01.013
Kats LM, Proellocks NI, Buckingham DW et al (2015) Interactions between Plasmodium falciparum skeleton-binding protein 1 and the membrane skeleton of malaria-infected red blood cells. Biochim Biophys Acta. doi:10.1016/j.bbamem.2015.03.038
Celada A, Cruchaud A, Perrin LH (1982) Opsonic activity of human immune serum on in vitro phagocytosis of Plasmodium falciparum infected red blood cells by monocytes. Clin Exp Immunol 47:635–644
Bouharoun-Tayoun H, Attanath P, Sabchareon A et al (1990) Antibodies that protect humans against Plasmodium falciparum blood stages do not on their own inhibit parasite growth and invasion in vitro, but act in cooperation with monocytes. J Exp Med 172:1633–1641
Beeson JG, Brown GV (2004) Plasmodium falciparum-infected erythrocytes demonstrate dual specificity for adhesion to hyaluronic acid and chondroitin sulfate A and have distinct adhesive properties. J Infect Dis 189:169–179. doi:10.1086/380975
Fernandez V, Hommel M, Chen Q et al (1999) Small, clonally variant antigens expressed on the surface of the Plasmodium falciparum-infected erythrocyte are encoded by the rif gene family and are the target of human immune responses. J Exp Med 190:1393–1404
Kyes SA, Rowe JA, Kriek N, Newbold CI (1999) Rifins: a second family of clonally variant proteins expressed on the surface of red cells infected with Plasmodium falciparum. Proc Natl Acad Sci USA 96:9333–9338
Bachmann A, Scholz JAM, Janßen M et al (2015) A comparative study of the localization and membrane topology of members of the RIFIN, STEVOR and PfMC-2TM protein families in Plasmodium falciparum-infected erythrocytes. Malar J 14:274. doi:10.1186/s12936-015-0784-2
Beeson JG, Brown GV, Molyneux ME et al (1999) Plasmodium falciparumIsolates from infected pregnant women and children are associated with distinct adhesive and antigenic properties. J Infect Dis 180:464–472. doi:10.1086/314899
Fried M, Duffy PE (1996) Adherence of Plasmodium falciparum to chondroitin sulfate A in the human placenta. Science 272:1502–1504
Salanti A, Dahlbäck M, Turner L et al (2004) Evidence for the involvement of VAR2CSA in pregnancy-associated malaria. J Exp Med 200:1197–1203. doi:10.1084/jem.20041579
Ricke CH, Staalsoe T, Koram K et al (2000) Plasma antibodies from malaria-exposed pregnant women recognize variant surface antigens on Plasmodium falciparum-infected erythrocytes in a parity-dependent manner and block parasite adhesion to chondroitin sulfate A. J Immunol 165:3309–3316
Avril M, Kulasekara BR, Gose SO et al (2008) Evidence for globally shared, cross-reacting polymorphic epitopes in the pregnancy-associated malaria vaccine candidate VAR2CSA. Infect Immun 76:1791–1800. doi:10.1128/IAI.01470-07
Ataíde R, Hasang W, Wilson DW et al (2010) Using an improved phagocytosis assay to evaluate the effect of HIV on specific antibodies to pregnancy-associated malaria. PLoS One 5:e10807. doi:10.1371/journal.pone.0010807
Feng G, Aitken E, Yosaatmadja F et al (2009) Antibodies to variant surface antigens of Plasmodium falciparum-infected erythrocytes are associated with protection from treatment failure and the development of anemia in pregnancy. J Infect Dis 200:299–306. doi:10.1086/599841
Voss TS, Healer J, Marty AJ et al (2006) A var gene promoter controls allelic exclusion of virulence genes in Plasmodium falciparum malaria. Nature 439:1004–1008. doi:10.1038/nature04407
Aguiar J, Albrecht G, Cegielski P (1992) Agglutination of Plasmodium falciparum-infected erythrocytes from east and west African isolates by human sera from distant geographic regions. Am J Trop Med Hyg 47:621–632
Mackintosh CL, Mwangi T, Kinyanjui SM et al (2008) Failure to respond to the surface of Plasmodium falciparum infected erythrocytes predicts susceptibility to clinical malaria amongst African children. Int J Parasitol 38:1445–1454. doi:10.1016/j.ijpara.2008.03.009
Perraut R, Mercereau-Puijalon O, Mattei D et al (1995) Induction of opsonizing antibod$$$ies after injection of recombinant Plasmodium falciparum vaccine candidate antigens in preimmune Saimiri sciureus monkeys. Infect Immun 63:554–562
Fried M, Nosten F, Brockman A et al (1998) Maternal antibodies block malaria. Nature 395:851–852. doi:10.1038/27570
Ndam NT, Denoeud-Ndam L, Doritchamou J et al (2015) Protective Antibodies against Placental Malaria and Poor Outcomes during Pregnancy, Benin. Emerging Infect Dis 21:813–823. doi:10.3201/eid2105.141626
Carlson J, Helmby H, Hill AV et al (1990) Human cerebral malaria: association with erythrocyte rosetting and lack of anti-rosetting antibodies. Lancet 336:1457–1460
Treutiger CJ, Hedlund I, Helmby H et al (1992) Rosette formation in Plasmodium falciparum isolates and anti-rosette activity of sera from Gambians with cerebral or uncomplicated malaria. Am J Trop Med Hyg 46:503–510
Elliott SR, Payne PD, Duffy MF et al (2007) Antibody recognition of heterologous variant surface antigens after a single Plasmodium falciparum infection in previously naive adults. Am J Trop Med Hyg 76:860–864
D’Ombrain MC, Voss TS, Maier AG et al (2007) Plasmodium falciparum erythrocyte membrane protein-1 specifically suppresses early production of host interferon-gamma. Cell Host Microbe 2:130–138. doi:10.1016/j.chom.2007.06.012
Allsopp CEM, Sanni LA, Reubsaet L et al (2002) CD4 T cell responses to a variant antigen of the malaria parasite Plasmodium falciparum, erythrocyte membrane protein-1, in individuals living in malaria-endemic areas. J Infect Dis 185:812–819. doi:10.1086/339521
Urban BC, Willcox N, Roberts DJ (2001) A role for CD36 in the regulation of dendritic cell function. Proc Natl Acad Sci USA 98:8750–8755. doi:10.1073/pnas.151028698
Simone O, Bejarano MT, Pierce SK et al (2011) TLRs innate immunereceptors and Plasmodium falciparum erythrocyte membrane protein 1 (PfEMP1) CIDR1α-driven human polyclonal B-cell activation. Acta Trop 119:144–150. doi:10.1016/j.actatropica.2011.05.005
Staalsoe T, Shulman CE, Bulmer JN et al (2004) Variant surface antigen-specific IgG and protection against clinical consequences of pregnancy-associated Plasmodium falciparum malaria. Lancet 363:283–289. doi:10.1016/S0140-6736(03)15386-X
Keen J, Serghides L, Ayi K et al (2007) HIV impairs opsonic phagocytic clearance of pregnancy-associated malaria parasites. Plos Med 4:e181. doi:10.1371/journal.pmed.0040181
Tebo AE, Kremsner PG, Luty AJF (2002) Fcgamma receptor-mediated phagocytosis of Plasmodium falciparum-infected erythrocytes in vitro. Clin Exp Immunol 130:300–306
Turrini F, Ginsburg H, Bussolino F et al (1992) Phagocytosis of Plasmodium falciparum-infected human red blood cells by human monocytes: involvement of immune and nonimmune determinants and dependence on parasite developmental stage. Blood 80:801–808
Francis SE, Malkov VA, Oleinikov AV et al (2007) Six genes are preferentially transcribed by the circulating and sequestered forms of Plasmodium falciparum parasites that infect pregnant women. Infect Immun 75:4838–4850. doi:10.1128/IAI.00635-07
Salanti A, Staalsoe T, Lavstsen T et al (2003) Selective upregulation of a single distinctly structured var gene in chondroitin sulphate A-adhering Plasmodium falciparum involved in pregnancy-associated malaria. Mol Microbiol 49:179–191. doi:10.1046/j.1365-2958.2003.03570.x
Staalsoe T, Megnekou R, Fievét N et al (2001) Acquisition and decay of antibodies to pregnancy-associated variant antigens on the surface of Plasmodium falciparum-infected erythrocytes that protect against placental parasitemia. J Infect Dis 184:618–626. doi:10.1086/322809
Blythe JE, Yan Yam X, Kuss C et al (2008) Plasmodium falciparum STEVOR proteins are highly expressed in patient isolates and located in the surface membranes of infected red blood cells and the apical tips of merozoites. Infect Immun 76:3329–3336. doi:10.1128/IAI.01460-07
Goel S, Palmkvist M, Moll K et al (2015) RIFINs are adhesins implicated in severe Plasmodium falciparum malaria. Nat Med 21:314–317. doi:10.1038/nm.3812
Beeson JG, Chan J-A, Fowkes FJ (2013) PfEMP1 as a target of human immunity and a vaccine candidate against malaria. Expert Rev Vaccines 12:105–108. doi:10.1586/erv.12.144
Ghumra A, Semblat J-P, Ataíde R et al (2012) Induction of strain-transcending antibodies against Group A PfEMP1 surface antigens from virulent malaria parasites. PLoS Pathog 8:e1002665. doi:10.1371/journal.ppat.1002665
Hviid L (2010) The role of Plasmodium falciparum variant surface antigens in protective immunity and vaccine development. Hum Vaccin 6:84–89
Good MF, Reiman JM, Rodriguez IB, Ito K (2013) Cross-species malaria immunity induced by chemically attenuated parasites. The Journal of clinical …. doi: 10.1172/JCI66634
Tanner M, Greenwood B, Whitty CJM et al (2015) Malaria eradication and elimination: views on how to translate a vision into reality. BMC Med 13:167. doi:10.1186/s12916-015-0384-6
Terheggen U, Drew DR, Hodder AN et al (2014) Limited antigenic diversity of Plasmodium falciparum apical membrane antigen 1 supports the development of effective multi-allele vaccines. BMC Med 12:183. doi:10.1186/s12916-014-0183-5
Cattani JA, Tulloch JL, Vrbova H et al (1986) The epidemiology of malaria in a population surrounding Madang, Papua New Guinea. Am J Trop Med Hyg 35:3–15
Beeson JG, Ndungu F, Persson KEM et al (2007) Antibodies among men and children to placental-binding Plasmodium falciparum-infected erythrocytes that express var2csa. Am J Trop Med Hyg 77:22–28
Hommel M, Elliott SR et al (2010) Evaluation of the Antigenic Diversity of Placenta-Binding Plasmodium falciparum Variants and the Antibody Repertoire among Pregnant Women. Infect Immun 78:1963–1978. doi:10.1128/IAI.01365-09
Beeson JG, Mann EJ, Elliott SR et al (2004) Antibodies to variant surface antigens of Plasmodium falciparum–infected erythrocytes and adhesion inhibitory antibodies are associated with placental malaria and have overlapping and distinct targets. J Infect Dis 189:540–551
Mwangi TW, Ross A, Snow RW, Marsh K (2005) Case definitions of clinical malaria under different transmission conditions in Kilifi District, Kenya. J Infect Dis 191:1932–1939. doi:10.1086/430006
Polley SD, Mwangi T, Kocken CHM et al (2004) Human antibodies to recombinant protein constructs of Plasmodium falciparum Apical Membrane Antigen 1 (AMA1) and their associations with protection from malaria. Vaccine 23:718–728. doi:10.1016/j.vaccine.2004.05.031
Maier AG, Rug M, O’Neill MT et al (2008) Exported proteins required for virulence and rigidity of Plasmodium falciparum-infected human erythrocytes. Cell 134:48–61. doi:10.1016/j.cell.2008.04.051
Kinyanjui SM, Howard T, Williams TN et al (2004) The use of cryopreserved mature trophozoites in assessing antibody recognition of variant surface antigens of Plasmodium falciparum-infected erythrocytes. J Immunol Methods 288:9–18. doi:10.1016/j.jim.2004.01.022
Petter M, Haeggström M, Khattab A et al (2007) Variant proteins of the Plasmodium falciparum RIFIN family show distinct subcellular localization and developmental expression patterns. Mol Biochem Parasitol 156:51–61. doi:10.1016/j.molbiopara.2007.07.011
Abdel-Latif MS, Khattab A et al (2002) Recognition of variant Rifin antigens by human antibodies induced during natural Plasmodium falciparum infections. Infect Immun 70:7013–7021
Schreiber N, Khattab A, Petter M et al (2008) Expression of Plasmodium falciparum 3D7 STEVOR proteins for evaluation of antibody responses following malaria infections in naive infants. Parasitology 135:155
Duffy MF, Byrne TJ, Elliott SR et al (2005) Broad analysis reveals a consistent pattern of var gene transcription in Plasmodium falciparum repeatedly selected for a defined adhesion phenotype. Mol Microbiol 56:774–788. doi:10.1111/j.1365-2958.2005.04577.x
Leykauf K, Treeck M, Gilson PR et al (2010) Protein kinase a dependent phosphorylation of apical membrane antigen 1 plays an important role in erythrocyte invasion by the malaria parasite. PLoS Pathogol 6:e1000941. doi:10.1371/journal.ppat.1000941
Boyle MJ, Reiling L, Feng G et al (2015) Human antibodies fix complement to inhibit Plasmodium falciparum invasion of erythrocytes and are associated with protection against malaria. Immunity 42:580–590. doi:10.1016/j.immuni.2015.02.012
Acknowledgments
Thanks to all participants in the study. We would like to acknowledge Brian Cooke for provision of the 3D7 SBP1KO isolate, Melanie Rug and Alan Cowman for provision of anti-PfEMP3 antibodies and CS2 SBP1KO parasites, Paul Gilson for provision of anti-HSP70 antibodies, Joe Smith for provision of the rabbit anti-DBL3 antibodies, Gaoqian Feng for assistance with opsonic phagocytosis assays and Simon Crawford for assistance with electron microscopy. Funding was provided by the National Health and Medical Research Council of Australia (Program grant and Senior research fellowship to JG Beeson, project grant to JG Beeson and SJ Rogerson, Infrastructure for Research Institutes Support Scheme Grant), Australian Postgraduate Award to J Chan, and the Australian Research Council (Future fellowships to JG Beeson and FJI Fowkes, Australian Research fellowship to AG Maier). The authors gratefully acknowledge support for the Burnet Institute from the Victorian Operational Infrastructure Support Program.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
18_2016_2267_MOESM1_ESM.pdf
Supplemental Figure S1: Expression of IE membrane proteins by 3D7 parasites. RIFIN (A) and STEVOR (B) proteins expressed by pigmented trophozoite-IEs of 3D7 parental and SBP1KO were visualized by immunofluorescence microscopy using specific α-RIF40 and α-STEVOR10 antibodies (green). RIFIN and STEVOR were detected in 3D7 SBP1KO (middle panel), similar to E8B parental (upper panel). The pattern of staining was consistent with the reported labeling of RIFIN and STEVOR in the IE membrane. The antibodies did not show any labeling of uninfected erythrocytes (uRBC) present in the culture (lower panel).C. Western blot analyses of pigmented trophozoite-IEs membrane extracts probed with α-RIF29 (left) and α-STEVOR3 (right) antibodies, which detected RIFIN at 40 kDa and STEVOR at 30 kDa in both 3D7 parental and 3D7 SBP1KO. Equal loading of parasite material was confirmed with anti-HSP70 antibodies (bottom panel). D. Pigmented trophozoite-IEs of 3D7 parental and 3D7 SBP1KO were probed with α-PfEMP3 antibodies (first column) as a positive control and α-AMA1 antibodies (second column) as a negative control (antibody staining in green, DAPI staining in blue). As expected, the pattern of staining was consistent with the labeling of PfEMP3 in the IE membrane and there was no apparent labeling of AMA1. In all immunofluorescence assays, cells were fixed with a mixture of acetone (90 %) and methanol (10 %) and DAPI was used to stain nuclear DNA (blue). All images were taken at equal exposure for both parasite lines and representative images are shown. (PDF 426 kb)
18_2016_2267_MOESM2_ESM.pdf
Supplemental Figure S2: Comparing the antibody response between adults and children from PNG. A. The level of IgG reactivity to the IE surface of 3D7 parental and 3D7 SBP1KO parasites was similar in both adults (n = 46) and children (n = 41). B. In the same group of adults (n = 46) and children (n = 41) as presented in (A), IgG binding to 3D7 SBP1KO was markedly reduced compared to 3D7 parental for both groups. C. The level of IgG reactivity to the IE surface of E8B parental and E8B SBP1KO parasites was significantly higher in adults (n = 50) compared to children (n = 41). D. In the same group of adults (n = 50) and children (n = 41) as presented in (C), IgG binding to E8B SBP1KO was substantially reduced compared to E8B parental for both groups. In all graphs, bars represent median and interquartile ranges of samples tested in duplicate; IgG binding levels are expressed as geometric mean fluorescence intensity (MFI) and control refers to serum samples from non-exposed Melbourne residents; assays were performed twice independently. The p values in (A) and (C) were calculated using the Mann–Whitney test and the p values in (B) and (D) were calculated using a paired Wilcoxon signed rank test. (PDF 89 kb)
18_2016_2267_MOESM3_ESM.pdf
Supplemental Figure S3: Antibody levels measured as IgG binding by flow cytometry in pregnant women samples. A. Serum samples from pregnant women in Malawi tested for antibodies to CS2 parental and CS2 SBP1KO parasites are grouped by gravidity. Samples were from malaria-exposed primigravid (PG; n = 35) and multigravid (MG; n = 36) women and men (n = 10). IgG binding to CS2 SBP1KO parasites was substantially reduced compared to CS2 parental in PG women, MG women and men. There was minimal background reactivity observed among sera from non-exposed Melbourne residents (Control). Assay was performed once; p value was calculated using a paired Wilcoxon signed rank test; bars represent median and interquartile ranges of samples tested in duplicate. (PDF 55 kb)
18_2016_2267_MOESM4_ESM.pdf
Supplemental Figure S4: PfEMP1 is the major target of human antibodies. The overall IgG binding 3D7 SBP1KO was reduced compared to 3D7vpkd and 3D7 parental in PNG (A) and Kenya (C). Assays were performed twice independently; bars represent median and interquartile ranges (n = 81 for PNG, n = 26 for Kenya); p value was calculated using a paired Wilcoxon signed rank test; IgG binding levels are expressed as geometric mean fluorescence intensity (MFI) for all graphs. A representative selection of samples tested in parallel for antibodies to 3D7 parental, 3D7vpkd and 3D7 SBP1KO. Samples were from malaria-exposed PNG adults (B; P1-P31), Kenyan adults (D; C12-C25) and non-exposed Melbourne residents (Control). IgG binding to 3D7 SBP1KO was substantially reduced in most individuals, compared to 3D7vpkd and 3D7 parental. There was minimal background reactivity observed among sera from Melbourne residents. Assays were performed twice independently; bars represent mean and range of samples tested in duplicate. (PDF 101 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Chan, JA., Howell, K.B., Langer, C. et al. A single point in protein trafficking by Plasmodium falciparum determines the expression of major antigens on the surface of infected erythrocytes targeted by human antibodies. Cell. Mol. Life Sci. 73, 4141–4158 (2016). https://doi.org/10.1007/s00018-016-2267-1
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00018-016-2267-1