
Abstract
Key message
The AtOR gene enhances carotenoid levels in corn by promoting the formation of plastoglobuli when the carotenoid pool is limited, but has no further effect when carotenoids are already abundant.
Abstract
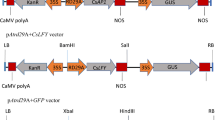
The cauliflower orange (or) gene mutation influences carotenoid accumulation in plants by promoting the transition of proplastids into chromoplasts, thus creating intracellular storage compartments that act as metabolic sink. We overexpressed the Arabidopsis OR gene under the control of the endosperm-specific wheat LMW glutenin promoter in a white corn variety that normally accumulates only trace amounts of carotenoids. The total endosperm carotenoid content in the best-performing AtOR transgenic corn line was 32-fold higher than wild-type controls (~25 µg/g DW at 30 days after pollination) but the principal carotenoids remained the same, suggesting that AtOR increases the abundance of existing carotenoids without changing the metabolic composition. We analyzed the expression of endogenous genes representing the carotenoid biosynthesis and MEP pathways, as well as the plastid fusion/translocation factor required for chromoplast formation, but only the DXS1 gene was upregulated in the transgenic corn plants. The line expressing AtOR at the highest level was crossed with four transgenic corn lines expressing different carotenogenic genes and accumulating different carotenoids. The introgression of AtOR increased the carotenoid content of the hybrids when there was a limited carotenoid pool in the parental line, but had no effect when carotenoids were already abundant in the parent. The AtOR gene therefore appears to enhance carotenoid levels by promoting the formation of carotenoid-sequestering plastoglobuli when the carotenoid pool is limited, but has no further effect when carotenoids are already abundant because high levels of carotenoids can induce the formation of carotenoid-sequestering plastoglobuli even in the absence of AtOR.





Similar content being viewed by others

References
Bai C, Rivera SM, Medina V, Alves R, Vilaprinyo E, Sorribas A et al (2014) An in vitro system for the rapid functional characterization of genes involved in carotenoid biosynthesis and accumulation. Plant J 77:464–475
Bai C, Capell T, Berman J, Medina V, Sandmann G, Christou P et al (2016) Bottlenecks in carotenoid biosynthesis and accumulation in rice endosperm are influenced by the precursor-product balance. Plant Biotechnol J 14:195–205
Berman J, Zorrilla-Lopez U, Farré G, Zhu C, Sandmann G, Twyman RM, Capell T, Christou P (2015) Nutritionally important carotenoids as consumer products. Phytochem Rev 14:727–743
Breitenbach J, Bai C, Rivera SM, Canela R, Capell T, Christou P, Zhu C, Sandmann G (2014) A novel carotenoid, 4-keto-α-carotene, as an unexpected by-product during genetic engineering of carotenogenesis in rice callus. Phytochemistry 98:85–91
Campbell R, Morris WL, Mortimer CL, Misawa N, Ducreux, LJM, Morris JA et al (2015) Optimising ketocarotenoid production in potato tubers: effect of genetic background, transgene combinations and environment. Plant Sci 234:27–37
Cao H, Zhang J, Xu J, Ye J, Yun Z, Xu Q et al (2012) Comprehending crystalline β-carotene accumulation by comparing engineered cell models and the natural carotenoid-rich system of citrus. J Exp Bot 63:4403–4417
Chayut N, Yuan H, Ohali S, Meir A, Sa’ar U, Tzuri G, Zheng Y, Mazourek M Gepstein S, Zhou X, Portnoy V, Lewinsohn E, Schaffer AA, Katzir N, Fei Z, Welsch R, Li L, Burger J, Tadmor Y (2017) Distinct mechanisms of the ORANGE protein in controlling carotenoid flux. Plant Physiol 173:376–389
Crisp P, Walkey DGA, Bellman E, Roberts E (1975) A mutation affecting curd colour in cauliflower (Brassica oleracea L var. Botrytis DC). Euphytica 24:173–176
Enfissi, EMA, Fraser PD, Lois LM, Boronat A, Schuch W, Bramley PM (2005) Metabolic engineering of the mevalonate and non-mevalonate isopentenyl diphosphate-forming pathways for the production of health-promoting isoprenoids in tomato. Plant Biotechnol J 3:17–27
Farré G, Perez-Fons L, Decourcelle M, Breitenbach J, Hem S, Zhu C, Capell T, Christou P, Fraser PD, Sandmann G (2016) Metabolic engineering of astaxanthin biosynthesis in maize endosperm and characterization of a prototype high oil hybrid. Transgenic Res 25:477–489
Fraser PD, Enfissi, EMA, Halket JM, Truesdale MR, Yu D, Gerrish C et al (2007) Manipulation of phytoene levels in tomato fruit: effects on isoprenoids, plastids, and intermediary metabolism. Plant Cell 19:3194–3211
Goo YM, Han EH, Jeong JC, Kwak SS, Yu J, Kim YH et al (2015) Overexpression of the sweet potato IbOr gene results in the increased accumulation of carotenoid and confers tolerance to environmental stresses in transgenic potato. C R Biol 338:12–20
Hugueney P, Bouvier F, Badillo A, D’Harlingue A, Kuntz M, Camara B (1995) Identification of a plastid protein involved in vesicle fusion and/or membrane protein translocation. Proc Natl Acad Sci USA 92:5630–5634
Jayaraj J, Devlin R, Punja Z (2008) Metabolic engineering of novel ketocarotenoid production in carrot plants. Transgenic Res 17:489–501
Kim SH, Ahn YO, Ahn MJ, Jeong JC, Lee HS, Kwak SS (2013) Cloning and characterization of an Orange gene that increases carotenoid accumulation and salt stress tolerance in transgenic sweetpotato cultures. Plant Physiol Biochem 70:445–554
Li L, Paolillo DJ, Parthasarathy MV, Dimuzio EM, Garvin DF, Plant US et al (2001) A novel gene mutation that confers abnormal patterns of β-carotene accumulation in cauliflower (Brassica oleracea var. botrytis). Plant J 26:59–67
Li L, Shan L, Cosman KM, Earle ED, Garvin DF, O’Neill J (2006) β-Carotene accumulation induced by the cauliflower Or gene is not due to an increased capacity of biosynthesis. Phytochemistry 67:1177–1184
Li L, Yang Y, Xu Q, Owsiany K, Welsch R, Chitchumroonchokchai C et al (2012) The or gene enhances carotenoid accumulation and stability during post-harvest storage of potato tubers. Mol Plant 5:339–352.
Lopez AB, Van Eck J, Conlin BJ, Paolillo DJ, O’Neill, JLL (2008) Effect of the cauliflower Or transgene on carotenoid accumulation and chromoplast formation in transgenic potato tubers. J Exp Bot 59:213–223
Lu S, Van Eck J, Zhou X, Lopez AB, O’Halloran DM, Cosman KM et al (2006) The cauliflower Or gene encodes a DnaJ cysteine-rich domain-containing protein that mediates high levels of beta-carotene accumulation. Plant Cell 18:3594–3605
Maass D, Arango J, Wüst F, Beyer P, Welsch R (2009) Carotenoid crystal formation in Arabidopsis and carrot roots caused by increased phytoene synthase protein levels. PLoS ONE 4:e6373
Morris WL, Ducreux, LJM, Hedden P, Millam S, Taylor MA (2006) Overexpression of a bacterial 1–deoxy-D–xylulose 5–phosphate synthase gene in potato tubers perturbs the isoprenoid metabolic network: implications for the control of the tuber life cycle. J Exp Bot 57:3007–3018
Naqvi S, Zhu C, Farre´ G et al (2011) Synergistic metabolism in hybrid corn indicates bottlenecks in the carotenoid path- way and leads to the accumulation of extraordinary levels of the nutritionally important carotenoid zeaxanthin. Plant Biotechnol J 9:384–393
Nogueira M, Mora L, En, EMA, Bramley PM, Fraser PD (2013) Subchromoplast sequestration of carotenoids affects regulatory mechanisms in tomato lines expressing different carotenoid gene combinations. Plant Cell 25:1–20
Paolillo DJ, Garvin DF, Parthasarathy MV (2004) The chromoplasts of Or mutants of cauliflower (Brassica oleracea L var. botrytis). Protoplasma 224:245–253
Park, S, Kim HS, Jung Y, Kim SH, Ji CY, Wang Z, Jeong JC, Lee HS, Lee SY, Kwak SS (2016) Orange protein has a role in phytoene synthase stabilization in sweetpotato. Sci Rep 16:33563. doi:10.1038/srep33563
Rivera S, Vilaró F, Canela R (2011) Determination of carotenoids by liquid chromatography/mass spectrometry: effect of several dopants. Anal Bioanal Chem 400:1339–1346
Rivera SM, Vilaró F, Zhu C, Bai C, Farré G, Christou P, Canela-Garayoa R (2013) Fast quantitative method for the analysis of carotenoids in transgenic maize. J Agric Food Chem 61:5279–5285
Shewmaker CK, Sheehy JA, Daley M, Colburn S, Ke DY (1999) Seed-specific overexpression of phytoene synthase: Increase in carotenoids and other metabolic effects. Plant J 20:401–412
Wurtzel ET (2004) Chapter five genomics, genetics, and biochemistry of maize carotenoid biosynthesis. Recent Adv Phytochem 38:85–110
Yu D, Lydiate DJ, Young LW et al (2008) Enhancing the carot enoid content of Brassica napus seeds by downregulating lycopene epsilon cyclase. Transgenic Res 17:573–585
Yuan H, Owsiany K, Sheeja TE, Zhou X, Rodriguez C, Li Y et al (2015) A Single amino acid substitution in an ORANGE protein promotes carotenoid overaccumulation in arabidopsis. Plant Physiol 169:421–431
Zhou X, Welsch R, Yang Y, Álvarez D, Riediger M, Yuan H et al (2015) Arabidopsis OR proteins are the major posttranscriptional regulators of phytoene synthase in controlling carotenoid biosynthesis. Proc Natl Acad Sci USA 112:3558–3563
Zhu C, Naqvi S, Breitenbach J, Sandmann G, Christou P, Capell T (2008) Combinatorial genetic transformation generates a library of metabolic phenotypes for the carotenoid pathway in maize. Proc Natl Acad Sci USA 105:18232–18237
Zhu C, Bai C, Sanahuja G et al (2010) The regulation of carotenoid pigmentation in flowers. Arch Biochem Biophys 504:132–141
Acknowledgements
Research at the Universitat de Lleida is supported by MINECO, Spain (BIO2014-54426-P; BIO2014-54441-P), by the Catalan Government (2014 SGR 1296 Agricultural Biotechnology Research Group), and by European Union Framework 7, European Research Council IDEAS Advanced Grant BIOFORCE and POC Grant (to PC). G. Farré is supported by a J de la C fellowship.
Author’s contribution
CZ, GS, TC, and PC designed research; JB, UZ, and GF performed research, analyzed data, and wrote the article; VM did TEM and analyzed these data.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Communicated by Salim Al-Babili.
Electronic supplementary material
Below is the link to the electronic supplementary material.
299_2017_2126_MOESM1_ESM.docx
Supplementary material 1 Supplementary Figure 1—RNA blot analysis of AtOR mRNA in wild type (M37W) and two different transgenic lines (OR1 and OR2) transformed with the AtOR gene driven by the wheat low-molecular-weight glutelin promoter. Each lane was loaded with 25 μg total RNA isolated from endosperm tissue Ribosomal RNA stained with ethidium bromide is shown as a loading control. Supplementary Figure 2—Transcript accumulation normalized against ACTIN mRNA in wild-type and transgenic lines presented as means of three technical replicates. Standard error bars were not included due to the use of technical replicates rather than biological replicates (the aim was to confirm transgene expression rather than to compare different transcript profiles). Supplementary Figure 3—Schematic representation of the transgenes expressed in our transgenic plants lines and hybrids. Supplementary Table 1—Carotenoid content and composition in wild-type M37W, transgenic lines OR2, CARO1, CARO2, KETO1 and KETO2, and hybrids ORxCARO1, ORxCARO2, ORxKETO1 and ORxKETO2 (T1 plants) at 30 (*) and 60 DAP (**) (µg/g DW±SE) (n = 3–5 seeds). Abbreviations: Phyt, phytoene; Lyco, lycopene; βcryp, β-cryptoxanthin; βcaro, β-carotene; Lut, lutein; Zea, zeaxanthin; Anthe, antheraxanthin; Viola, violaxanthin; CAROT, carotenoids. Supplementary Table 2—Ketocarotenoid content and composition of wild-type M37W, transgenic lines OR2, CARO1, CARO2, KETO1 and KETO2, and hybrids ORxCARO1, ORxCARO2, ORxKETO1 and ORxKETO2 T1 at 30 (*) and 60 DAP (**) (µg/g DW±SE) (n = 3–5 seeds). Abbreviations: Asta, astaxanthin; Cantha, canthaxanthin; Adonir, adonirubin; Adonix, adonixanthin; 3OHechi, 3-OH-echinenone; KETO, ketocarotenoids. Supplementary Table 3—Oligonucleotide sequences of corn ACTIN, endogenous carotenogenic genes and transgenes for real-time quantitative RT-PCR analysis. (DOCX 114 KB)
Rights and permissions
About this article

Cite this article
Berman, J., Zorrilla-López, U., Medina, V. et al. The Arabidopsis ORANGE (AtOR) gene promotes carotenoid accumulation in transgenic corn hybrids derived from parental lines with limited carotenoid pools. Plant Cell Rep 36, 933–945 (2017). https://doi.org/10.1007/s00299-017-2126-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00299-017-2126-z