
Abstract
The use of subfossil cladocerans is rare for understanding environmental, cultural and archaeological developments of lakes in Europe. In 2007, we collected a 12-m long sediment core from Lake Schalkenmehrener Maar (SMM), Germany for the analyses of subfossil cladocerans. Dating of core was based on tephrochronology, radiocarbon and pollen stratigraphy. Pollen-based chronostratigraphy indicated a decline of species richness and abundance of chydorids during the Lateglacial (ca.14500 cal yr bp) with dominant cold preferring taxa Acroperus harpae Baird and Alona affinis Leydig. During the early Bölling, the abundance of cladocerans increased commensurate with growth of birch (Betula L.) and pine (Pinus L.) trees. Except a spike of Bosmina coregoni Baird during the Younger Dryas, cladoceran assemblages remained stable from the Bölling to the mid-Atlantic period. During the Neolithic (ca. 4300 bc), the abundance of B. coregoni increased sharply with reciprocal decrease in Daphnia. However, as soon as Daphnia was dominant (ca. 4250 bc), a reciprocal decline in abundance of B. coregoni occurred. The mid-Holocene change in cladoceran abundance coincided with the use of hardwood forest. This situation ended at ca. 4000 bc and remained unchanged throughout the Neolithic and Bronze Age (ca. 3000–1200 bc). Low Daphnia abundance indicated reduced water quality in the Hunsrück-Eifel culture (ca. 800 bc). A spike of B. coregoni at ca. ad 150 indicates construction of the Roman Villa Rustica and extensive farming. However, reoccurrence of Daphnia at ca. ad 470 indicates the retreat of the Romans from the Eifel region. From the early Frankish rule (ca. ad 500) to the Medieval period (ca. ad 1500), species richness reduced but abundance of B. coregoni increased indicating a switch in lake ecosystem. The loss of species richness and the lack of precise evidence of the human activity in the region in the past have impeded the restoration of the ecosystem of the Lake SMM.
Similar content being viewed by others

Introduction
The maar crater lakes of the Eifel region of West Germany are highly sensitive to environmental change. Most of Eifel maar crater lakes are deep and have closed basins with anoxic bottom waters creating suitable conditions for the formation of high-resolution laminated sediment records (Brauer et al., 1999a; Schaber & Sirocko, 2005). Abiotic and biotic assemblages preserved in varved lake sediment are excellent indicators of annual and subannual variation in climates (Litt et al., 2001a, b), catchment erosion (Zolitschka & Negendank, 1999; Zolitschka et al., 2003), water chemistry (Schettler & Romer, 1998; Lücke et al., 2003) and palaeoenvironmental changes such as variability in windstorms (Brüchmann & Negendank, 2004; Pfahl et al., 2009). Preservation of some significant anthropogenic signatures that are crucial to understand human civilisation in Central Europe has also been reported in sediment of Eifel maar crater lakes from the beginning of the Neolithic (Gronenborn & Sirocko, 2009; Sirocko, 2009).
Environmental and archaeological investigations of the pre-European culture have demonstrated that land use, waste disposal and pollution were intensified across the Eifel mountain region soon after the Neolithic (Zolitschka et al., 2003). Domestication of animals, invention of agricultural tools and cultivation of cereals were the characteristic features of the cultural and archaeological developments following the Neolithic (Cunliffe, 1997; Bonsall et al., 2002; Bradshaw et al., 2005; Rasmussen, 2005; Rasmussen & Bradshaw, 2005). Meantime, unique cultural values of the individual society also began to develop. Choice of food and clothing and their adaptation became a part of the cultural life in Europe and elsewhere which would reflect on how natural resources being utilised by humans locally over the period (Kuijper & Turner, 1992; Cunliffe, 1997; Erlandson, 2001).
Evidence of leaf foddering and agriculture has been documented in the Eifel maar lakes sediment as far back as 4600 bc (Gronenborn & Sirocko, 2009; Sirocko, 2009). Farming activity across the Eifel region appears to have altered the physical and hydrological properties of majority of the maar lakes. Catchment erosion, nutrient enrichment and runoff may have led to algal blooms and reduced dissolved oxygen levels subsequently altering the trophic status of the lake. Evidence of these changes caused by rapid climate change and anthropogenic disturbances have been archived in a fine temporal resolution of sediment of majority of maar lakes in the region (Creer & Thouveny, 1996). Following the Roman invasion of the Eifel region, a long-term ecological upheavals were experienced by these maar lakes as a result of over-harvesting of natural resources including the cultivation, cattle raising, fishing and deforestation across the region (e.g. Zolitschka et al., 2003).
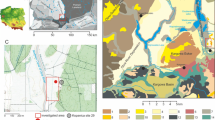
Cladocerans play an intermediary role in lake ecosystems and food web structure and dynamics, being significant grazers of the lower trophic levels and also an important prey items for secondary consumers such as planktivorous fish (Jeppesen et al., 2000). Direct human intervention in lake catchments such as rapid deforestation, cultivation, sedimentation and fishing can have significant effects on cladoceran assemblages via alteration of lake productivity and fish populations (Johansson et al., 2005). Cladocerans have been shown to respond to major environmental changes and its implications in food web structure and dynamics in European lakes (Wagner & Bendorf, 2007). Subfossil assemblages of cladocerans have been used extensively to reconstruct past environmental changes including climate change and eutrophication of many European lakes (e.g. Lotter et al., 1997; Korhola, 1999; Johansson et al., 2005; Kattel et al., 2008). However, a comprehensive study of cladoceran-inferred environmental, cultural and archaeological developments in the pre-historic Europe is rare. Pre-historic Germany, for example, experienced a range of social and political turmoils (Moses, 1976). Several cultures were thrived and vanished in West Germany (Moses, 1976). The period of the Romans, in particular had considerable implications in natural resources including the use of maar lakes in the Eifel region (Zolitschka et al., 2003). In this study, we aim to investigate the role of palaeocladocerans and subfossil ephippia for inferring environmental, cultural and archaeological developments around the Eifel maar lakes region (West Germany) based on a sediment core derived from the Lake Schalkenmehrener Maar (SMM), initially formed by a phreatomagmatic explosion during the Lateglacial period (Fig. 1).

Map of the study site, Lake SMM in Westeifel, Germany
Study site
Lake SMM (50°10′N, 6°50′60″E) is one of 68 maar crater lakes in the Quaternary Westeifel Volcanic Field (Fig. 1) formed ca. 20,000 years ago (Büchel, 1993). The volcanic ejecta of Lake SMM are alkalibasaltsic and were mainly deposited locally to create the tuft ring. The modern maar lake (Fig. 2) is connected to a neighbouring dry maar structure, which has been filled with sediment extending from the Lateglacial into Medieval times (Straka, 1975). The basin of the northern maar lake acted as a sediment trap, thus hindering the discharge of allochthonous clastic particles into the southern maar lake (Hofmann, 1993; Schettler et al., 1999). The catchment of the maar today is 1,299 km2, with a maximum water depth of 21 m, a total volume of 3 Mm3, and maximum water residence time of 9 years (Scharf & Björk, 1992).

Digital elevation model of the modern maar lakes in Westeifel Germany
Physico-chemical variables such as temperature, maximum depth and dissolved oxygen profiles suggest that Lake SMM is a dimictic lake (Schettler et al., 1999). The current metalimnetic dissolved oxygen measurement at thermocline depth is higher (>15 ppm) than the epilimnion (~10 ppm); with values below 1 ppm characterise the hypolimnion. The summer temperature of the lake water reaches to 22°C in the epilimnion and the thermocline usually lies at between 10 and 6°C in the hypolimnion (Seelos & Sirocko, 2009). Although various measures have been taken to restore the Eifel maar lake ecosystems since 1900s, tendency of lake eutrophication is common in many of these lakes including the Lake SMM (Möller & Scharf, 1986; Scharf & Oehms, 1992).
Materials and methods
Coring
Four sediment cores were retrieved from the Lake SMM between 2005 and 2007 for the project, Eifel Laminated Sediment Archives (ELSA). One core was taken using a rope operated freeze corer and the other three were taken with a Niederreiter piston corer. We used the Core SM2 to develop a pollen-based stratigraphy (Fig. 3) and cross-correlated it with Core SM3, which was used for subfossil cladoceran analysis. Other two remaining cores were used for geochemical analyses.

Palynostratigraphy (%) for Core SM2 of Lake SMM
Pollen analysis
Pollen preparation followed the techniques of Berglund & Ralska-Jasiewiczowa (1986) and Faegri & Iversen (1989), modified by Litt (2004) and Kühl & Litt (2007). The samples were sieved through a 200 μm sieve and later filtrated through a 10 μm filter. Lycopodium-spore tablets were added for calibration of the absolute pollen concentration per cm3. Pollen counting was done under a maximum of six hundred fold magnification. The percentage of pollen spectra were calculated using the sum of all pollen taxa displayed in Fig. 3.
Cladoceran analysis
A moderate resolution analysis of the palaeocladoceran records was carried out for the period covering 15000–500 cal yr bp involving 42 samples from the Core SM3. The resolution for the upper 20 samples between 1 and 3 m was at ca. 10 cm intervals, while the lower 22 samples between 3 and 9 m had ca. 25 cm sampling intervals. Differences in sampling resolution after 3 m are due to lack of sufficient sediment sample available for studying at the required depth points. From 6.6 to 5.5 m depth, cladoceran remains were poorly preserved consequently needed a large amount of sediment to analyse. Cladoceran preparation followed the method of Kattel et al. (2008). All slides were examined under a microscope at 100× and 400× magnifications. All cladoceran remains (carapaces, head-shields, post-abdomen, ephippia, and post-abdominal claws) were tabulated separately, but only the most frequently encountered body parts for each taxon were used for estimating taxon abundance. If the two halves of the bilateral carapace came apart, one half was counted as half of an individual. Badly fragmented remains were counted if they contained a clear diagnostic feature as proposed by Goulden (1969). The dry weight percentage of each sediment sample was used to calculate the remains present per gram of dry sediment. At least 200 cladoceran individuals were counted from each sample. Identification was carried out at species level using the method by Frey (1958, 1959, 1986), Whiteside et al. (1978), and Korhola & Rautio (2001). The identification was based on Szeroczyńska & Sarmaja-Korjonen (2007).
Core chronology
A chronology for the changing subfossil assemblages of cladocerans is provided through a core cross-correlation and 14C dated palynological markers evident in the adjacent core (Core SM2). Core cross-correlation was carried out using a set of distinct lithological changes evident in both records (Table 1). Palynological marker layers are shown to have demonstrated an identical pattern of changes to those observed in the varve chronology from Lake Meerfelder Maar (Brauer et al., 1999b; Kubitz, 2000). We used the first and subsequent appearance of hazel (Corylus L.) (ca.ca. 14000 bp and 8850 bc), first appearances of ash (Fraxinus L.) and lime (Tilia L.) (ca. 6600 bc) and the spread of alder (Alnus Mill.) (ca. 4250 bc), beech (Fagus L.) (ca. 1850 bc), hornbeam (Carpinus L.) (ca. ad 450) and grasses such as from the family Poaceae (ca. ad 900) as time markers for comparison with the palynostratigraphy of Core SM2 (Fig. 3) using distinct marker layers (Table 1).
Numerical analysis
Initially, a detrended correspondence analysis (DCA) was carried out for cladoceran subfossil data in CANOCO 4.5 (ter Braak & Smilauer, 2002). Squared-root transformation was performed for the percentage data before analysis. A principal components analysis (PCA) was an appropriate technique to understand the palaeocladoceran data in Core SM3 samples since the gradient length of the first axis of DCA was less than 3. PCA of the cladoceran assemblages were derived from only those cladoceran taxa comprising a minimum of >1% in at least two samples were used during PCA analysis (e.g. Birks, 1995). Because of their poor preservation, eight samples from below 7 m depth were excluded from the PCA analysis. Following the analysis of the remaining 34 samples, the result shows that more than 75% of variability in the cladoceran species data has been explained by the PCA axis 1 followed by the PCA axis 2, which accounted for only 10% of the total variation (Table 2). Summary of the PCA sample scores has been used in cladoceran stratigraphy diagram and will be discussed in result and discussion sections. A dendrogram type cluster analysis followed by Bray-Curtis similarity test for 34 sediment samples was carried out in PRMIER (Clarke & Warwick, 2001; Clarke & Gorley, 2006). Cluster analysis in PRIMER defines groups of cases (in this case depth samples) based on the similarity of multiple variables measured for each case (Clarke & Gorley, 2006). The similarity measure is an appropriate coefficient for exploring biological community similarities (Clarke & Warwick, 2001). The major changes in the cladoceran assemblage following the cluster analysis are presented as dendrogram in Appendix 1—Electronic Supplementary Material.
Results
Lithostratigraphy
Core SM3 (9–6.5 m) is characteristic of the Lateglacial silty gyttja. The dark colour above 6.9 m represents the transition into the Bölling (Fig. 4). At 6.4 m clay dominates gyttja and changes to brownish colour during the Alleröd (Fig. 4). The Laacher See Tephra (LST) represents from 6.3 to 6.2 m (ca. 13200–12800 years bp). Above 6.2 m depth, a lighter dark gyttja with low organic contents dominate. The carbonate content appears to be high from 5.2 to 5 m depth when 8.2 ka Event occurred (Schmidt & Gruhle, 2003). At 4.3 m depth, this become brownish while above 2.8 m this gradually turn darkish brown colour (Fig. 4).

Lithology of the sediment Core SM3 of Lake SMM
Cladoceran stratigraphy
A total of 40 cladoceran taxa were identified in 34 samples. The 26 most abundant species, defined as species occurring more than 1% in at least in two samples are shown in Fig. 5. Unusually the high PCA axis 1 sample scores (75%) indicated higher influence of Daphnia than Bosmina populations in the lake, while axis 2 (10%) reflected the higher influence of littoral to planktonic (L:P) ratios (Fig. 5). The cluster analysis of the 34 samples shows that at least 3 to 4 samples indicated the lowest similarity levels of Lake SMM ecosystems over the past (Appendix 1—Electronic Supplementary Material). Some major changes in Lake SMM occurred during the Lateglacial-Bölling, Stone Age–Bronze Age and Iron Age–Roman period transitions, where the samples during the Stone Age–Bronze Age transition showed ca. 50% similarity levels (Fig. 5; Appendix 1—Electronic Supplementary Material).

Palaeocladoceranstratigraphy for Core SM3 of Lake SMM. Zonation is based on cluster analysis in PRIMER. At least three major zones and several other smaller zones were formed. The major zones are separated by lines at ca. 14000 bp, 4200 bc and 50 bc, respectively. See a dendrogram presented in Appendix 1—Electronic Supplementary Material for further details on level of similarity amongst depth samples. The list of cladoceran species which are not presented in the diagram but were recorded during this study are: Alonopsis elongata Sars, Disparalona rostrata Koch, Alonella exigua Lilljeborg, Alonella excisa Fischer, Pseudochydorus globosus Baird, Pleuroxus uncinatus Baird, Pleuroxus laevis Sars, Peracantha truncata Baird, Anchistropus emarginatus Sars, Leydigia acanthocercoids Fischer, Kurzia latissima Kurz, Sida crystallina O.F. Muller, Bythotrephes longimanus Leydig, Polyphemus pediculus Linne, Holopedium gibberum Zadach
At the bottom section of the stratigraphy covering 8.88–6.8 m (Lateglacial-Bölling transition—ca. 14.5–13.2 kyr bp), most cladoceran species occurring in littoral substrata of the lake were recorded. During the Lateglacial, prior to ca. 14500 cal yr bp, there was a poor preservation of the remains of chydorid cladocerans consisting mainly of three littoral plant-mud-dwelling taxa such as Acroperus harpae Baird, Alona affinis Leydig and Chydorus sphaericus O. F. Müller, respectively (Fig. 5). Pollen-inferred stratigraphy shows the occurrence of Lateglacial vegetation across the vicinity of the Lake SMM with dominant grass species during this period (Fig. 3).
Following 14500 cal yr bp (7.85–7.05 m), during the Meiendorf period, species richness and abundance of chydorid cladocerans gradually began to increase. Littoral chydorid species present during the Lateglacial period continued to dominate the lake (Fig. 5). The records of cladocerans during the Meiendorf were accompanied by shrubs and dwarfs type vegetation at ca. 14500 bp (Fig. 3). However, in the Bölling period from ca. 13300–13200 cal yr bp (7.05–6.80 m), the abundance of the limnetic taxon, Daphnia sp. was high. Meantime, the species richness of littoral cladocerans also continued to increase, where C. sphaericus was the dominant taxon (Fig. 5). Onset of Daphnia in Lake SMM was commensurate with the development of the open forest woodlands such as dominant birch (Betula L.) populations in the lake catchments during the early part of the Bölling (e.g. Litt et al., 2001a, b).
No cladoceran remains were preserved in lake sediment during the Alleröd (ca. 13200–12800 cal yr bp), when the LST (Fig. 4) were predominantly deposited between 6.8 and 6.28 m depths (Fig. 5). However, in the Younger Dryas (6.28–6 m/ca. 12800–11600 cal yr bp), the composition and richness of both limnetic and littoral cladocerans increased. Daphnia was the only key limnetic cladoceran recorded in the maar sediment dominated the total cladoceran populations, while littoral chydorids such as A. harpae, A. affinis and C. sphaericus dominated the littoral cladoceran assemblage during the Younger Dryas (Fig. 5). Subarctic tundra vegetation (Artemesia L., Helianthemum Miller) dominated the catchments of Lake SMM during the Younger Dryas (Fig. 3).
From the Preboreal to until the late Holocene, the species richness and composition of both limnetic and littoral cladocerans increased although there were switches in abundances between limnetic Daphnia and Bosmina as well as total limnetic and littoral cladocerans over different time scales. During the preboreal and boreal periods between 6–5.5 m covering ca. 11600–8550 cal yr bp, the pelagic cladocerans were dominated by Daphnia, and a few remains of Bosmina were also recorded (Fig. 5). The species richness of littoral cladocerans increased, where Alona guttata was the dominant taxon in at least one of the three samples being collected during this interval (Fig. 5). Pollen stratigraphy during this period indicates a gradual increase in warm preferring vegetation such as hazel (Corylus L., Fig. 3). This time was accompanied by the onset of A. guttata (Fig. 5), a mild climate preferring taxon across the temperate Europe (Hofmann, 1996).
During the Atlantic period at 5.5–4.4 m (ca. 6600–4250 bc), concentrations of cladoceran ephippia in sediment increased (Fig. 5). However, the assemblage of Daphnia gradually declined with reciprocal increase in the assemblage of Bosmina coregoni Baird and Bosmina longirostris O. F. Müller (Fig. 5). This period is broadly regarded as Stone Age when the Lake SMM witnessed the 8.2 ka Event. Significant numbers of resting eggs were also produced by cladocerans during this period (Fig. 5). Cluster analysis shows that a major transition in cladoceran assemblages occurred between Stone Age and Bronze Age (Fig. 5). This period, a part of the Subboreal time, extended between ca. 4.4 and 3.4 m (ca. 4250–1850 bc), when the Daphnia populations increased significantly and the abundance of B. coregoni started to decline (Fig. 5). Abundance of a chydorid taxon, C. sphaericus, increased till 3.5 m then gradually began to decline (Fig. 5).
The cluster analysis also shows that a major shift in cladoceran assemblage occurred in transition between the end of the Iron Age and the early Roman period (Fig. 5). The Subatlantic period in West Germany covered mostly the Roman, Migration and Medieval period altogether. In the early Subatlantic period, at ca. 3 m (50 bc), Daphnia populations were recorded low while the abundance of C. sphaericus and A. guttata was relatively high resulting in high L:P taxon ratios (Fig. 5). The response of high cladoceran L:P taxon ratios was also associated with high PCA sample scores in axis 2 (Fig. 5). Meantime, dominant mixed oak-alder (Quercus L.–Alnus Mill.) hardwood forests were recorded in the lake catchment (Fig. 3).
Pollen stratigraphy shows that from the early Roman period, the landscape of the Eifel maar lakes region has changed rapidly with sparse tree populations but dominant cereals and grass densities (Fig. 3). These changes in landscape have been indicated by a number of shifts in the abundance of cladoceran species (e.g. Bosmina vs. Daphnia) or groups (planktonic vs. littoral). There is an overlap between the Roman and the Migration periods. In West Germany, while Romans were retreating, migration began (Schreg & Sirocko, 2009; Sirocko, 2009). During the Roman period at 3–2.6 m (ca. ad 50–450), there was a significant increase in the abundance of limnetic cladocerans mainly Bosmina. A marked abundance of Bosmina coregoni Baird was observed above 2.8 m depth (Fig. 5). While in Migration period from 2.85 to 2.6 m (ca. ad 300–500), Daphnia populations increased briefly but significantly. However, at the end of the Roman rule at ca. ad 471 (2.5 m), Daphnia assemblage showed a reciprocal response to the assemblage of B. coregoni although it was only in one sample (Fig. 5). Only a small proportion of the chydorid cladocerans, such as A. guttata, A. harpae, Camptocercus rectirostris Schödler, Alonella nana Baird and C. sphaericus were recorded during the Migration period (Fig. 5). The density of resting eggs of both limnetic and littoral cladocerans increased significantly during the Migration period (Fig. 5). The abundance of B. coregoni was relatively high and remained at constant level during the Early Medieval period from 2.4 to 2 m (ca. ad 500–900). B. coregoni, along with B. longispina continued to dominate the cladoceran assemblage (Fig. 5) from about 2.4 to 1 m depth corresponding to ca. ad 500–present time (Fig. 5). The abundance of B. coregoni was high throughout the Early Medieval period. The species richness of littoral chydorids increased although their contribution to the total cladoceran assemblage was minimal (Fig. 5). Littoral chydorids such as Graptoleberis testudinaria Fischer, A. affinis, Eurycercus lamellatus O. F. Müller, C. rectirostris Schödler, A. harpae and Pleuroxus trigonellus O. F. Müller increasingly emerged during the Medieval period (Fig. 5). An increased number of ephippia was recorded from, ca. 2.2 and 1.9 m corresponding to ca. ad 680 and 880, respectively (Fig. 5). During the High Medieval period from 2 to 1 m depth (ca. ad 900–1500), the species richness of cladocerans declined slightly (Fig. 5). Only one taxon, B. coregoni made a significant (up to 80%) contribution to the total cladoceran assemblages of the lake (Fig. 5).
Discussion
Lithostratigraphy
Lighter colour gyttja of the Core SM3 just above the 6.2 m depth indicates low organic contents characteristic of oligotrophic sediment deposition. However, high carbonates in sediment from 5.2 to 5 m depth may have been caused by diatom-rich organic contents. The 8.2 ka Event appears to have influenced the sediment colour (Schmidt & Gruhle, 2003). Brownish colour sediment at 4.3 m depth indicates the expansion of alder (Alnus Mill.) trees (Fig. 3). Gyttja with rich organic carbon between 4 and 3 m depths indicate shallow swampy margins possibly as a result of increased runoffs, deforestation and catchment burning across the northern maar lakes of the Eifel region. The light colour (Munsell Soil Colour Chart: 5/1, http://www.color-chart.org/munsell-color-chart.php) gyttja with higher proportion of coarser grained clastic sediment above 4 m depth was possibly a result of extended land use activity. Frequent flash floods during the late Bronze Age, and anthropogenic activity by the Hunsrück-Eifel culture (ca. 700–300 bc) during the Iron Age increased sedimentation rates. The sediment colour during the Roman period became brownish (Munsell Soil Colour Chart: 7/2, http://www.color-chart.org/munsell-color-chart.php) due to a frequent wet and mild climates (Schmidt & Gruhle, 2003).
During the early period of the first millennium (ad 150), a widespread cultivation of cereals particularly in the northern areas of the maar occurred. Today this area is relatively a dry. However, during flood events, a local creek transports runoffs into the lake (Fig. 2). The cultivation of the former swamp must have been occurred at ca. ad 150, when a Roman house “Villa Rustica” was also founded (Schreg & Sirocko, 2009; Sirocko, 2009; Fig. 1). Following the retreat of the Romans at ca. ad 460, an increase in birch (Betula L.) pollen occurred (Fig. 3) indicating the regional reforestation (Schreg & Sirocko, 2009; Sirocko, 2009). A frequent darkish-brown flash flood layers indicate rapid Migration period over a few centuries (beginning at 2.6 m), when beech (Fagus L.) and hornbeam (Carpinus L.) trees expanded across the region. However, the decrease in pollen between 2.5 and 2 m, the beginning of the High Medieval period (ca. ad 900), indicates rapid forest clearances (Fig. 4). Abundant macroscopic plant remains indicates dark flash flood layers that may have occurred in ad 1342 summer (Sirocko & Herbig, unpublished data). In the Medieval period, deforestation was widespread followed by increased arable farming across the central and north-western Europe (Rasmussen, 2005). Varying catchment disturbances between the Roman and the Medieval periods by humans resulted in differences in formation of minerogenic materials (Rasmussen & Bradshaw, 2005).
Environmental, cultural and archaeological significance of palaeocladocerans in Lake SMM
Trophic dynamics in lacustrine system is strongly linked to internal and external factors including climate change and human disturbances. Occurrence of these changes is a part of geological development reflecting environmental, cultural and archaeological changes of the region. Cladocerans archives have potential to respond historical development of lakes (Frey, 1986). When productivity is intensified, species preferring to nutrient-rich water bodies dominate the lake (Crisman & Whitehead, 1978). Fluctuations in occurrence of cladoceran and ephippial assemblages are also closely related to the changes in physico-chemistry, depth and surface water temperature (Lotter et al., 1997).
Human-induced cultural significance of planktonic cladocerans such as B. coregoni, B. longispina and B. longirostris have been reported in Europe (Polcyn, 1996; Szeroczyńska, 1998; Johansson et al., 2005). B. coregoni prefers relatively moderate level of nutrients (mesotrophic condition) compared to B. longispina, which prefers clearer, oligotrophic condition (Hofmann, 1996). B. coregoni tends to respond to both gradual human disturbances and climate-induced changes to lake water quality (Kattel et al., 2008). However, B. longirostris prefers hypertrophic lakes (Polcyn, 1996; Szeroczyńska, 1998). Increased abundance of B. longirostris in Lake SMM during the Neolithic (Fig. 5) indicates a degraded lake condition due to land use change across the region and widespread eutrophication (Hofmann, 1993; Kappes et al., 2000).
Records of some littoral chydorid cladocerans (A. harpae, A. affinis and C. sphaericus) in Lake SMM indicate cold climatic condition in the past. A. harpae is a common cladoceran during the Lateglacial time in Europe (Whiteside, 1970; Hofmann, 1987; Korhola, 1999). A. affinis is also a cold tolerant species usually colonise clear-water vegetated lake beds in the subarctic region (e.g. Harmsworth, 1968; Korhola, 1999). C. sphaericus is distributed to a wide range of habitats in both hemispheres (Whiteside et al., 1978; Korhola, 1999; Kattel & Augustinus, 2010). Indirect effects of cold climates can reduce nutrient levels (e.g. Kattel et al., 2008) when available benthic habitats (e.g. moss or algae layers) support cold climate adapting chydorids. No reconstruction of cladoceran inferred pH is available for Lake SMM, however, some species of littoral chydorids have shown a strong response to varying levels of lake water pH in Europe (Nilssen & Sandøy, 1990). Use of a range of variables including the temperature can help developing a strong inference model for Cladocera-based past environmental and archaeological changes.
Cladoceran eggs infer ecological stress and lacustrine integrity (Szeroczyńska, 1998; Sarmaja-Korjonen, 2004). Cladocerans undergo gamogenesis, a process when female cladocerans change their reproductive state producing haploid resting eggs (Frey, 1982; Sarmaja-Korjonen, 2003). Waste disposal, catchment disturbances and poor water quality are reflected by production of a large number of cladoceran resting eggs (Sarmaja-Korjonen, 2003, 2004). Increased resting eggs in Lake SMM following the widespread human disturbances including deforestation, grazing, cultivation and bank erosion during the Holocene indicate poor ecological integrity (Fig. 5).
Remains of Daphnia and their ephippia indicate past food web structure and dynamics. The relationship between planktivorous fish density (capture per unit effort—CPUE n ) and Daphnia body weight and egg size (dorsal length) is significant in European lakes (Jeppesen et al., 2002). Hypertrophic lakes have high mean Daphnia body weight while low CPUE n is in less disturbed sites (Jeppesen et al., 2002). Although no measurement was taken for Daphnia body size and their ephippia during the course of this study, changes in Daphnia abundance in Lake SMM during the Holocene reflects the dynamic trophic state (e.g. Johansson et al., 2005).
No documents are available for cladoceran-inferred reconstruction of archaeological artifacts and dietary condition of the pre-European people residing across the Lake SMM. However, evidence of archaeological records such as artifacts of nets, fishing gears and small tidal weirs provides the nature of prehistoric human adaptation (Erlandson, 2001). Cladocerans remains infer water level fluctuations and sedimentation of the impounded rivers. Cladoceran remains may provide the evidence of the type of construction materials being used (e.g. woods vs. non-woods) for dam, weir and bridge by prehistoric communities for fishing, milling, irrigating and transporting goods and materials (Mikulski, 1978; Polcyn, 1996). The use of woody materials, for example, indicates increased deforestation. Dams constructed from these materials can have poor cladoceran preservation due to bank erosion and non-stagnant water. However, the use of permanent materials such as stone may increase preservation of cladocerans and an existence of top-down control in ecosystem.
Palaeocladoceran-inferred environmental, cultural and archaeological developments in the Eifel maar lakes region
The qualitative (palaeostratigraphy), quantitative (PCA) and cluster analyses together show the increasing significance of palaeocladocerans and their ephippia for environmental, cultural and archaeological reconstructions in the Eifel maar lakes region, West Germany. The sample scores revealed by PCA axis 1 and 2 are indicative of the value of cladocerans in Lake SMM (Table 2). Planktonic cladocerans particularly, the Daphnia populations are important for shaping the food web structure and dynamics of the lake (axis 1), while L:P ratios is associated with changes in lake levels (axis 2). The high PCA axis 1 scores are unusual, possibly the result of a high species turnover rates of Daphnia during the early part of the Holocene when human disturbances in Lake SMM were relatively less than the latter part of the Holocene. Several investigations in sedimentary analyses of biological and chemical compositions (Möller & Scharf, 1986; Hofmann, 1990, 1993) and pollen analyses (Litt et al., 2001a, b) in the past reveal that the Lateglacial vegetation landscape in the vicinity of Lake SMM was predominantly covered by grasses, Pinus L. and Betula L. trees (Fig. 3). Macroscopic organic remains reveal that growth of mosses and oospores from Characeae was rapid during the Lateglacial (Sirocko & Herbig, unpublished data). The preservation of cladoceran remains was relatively poor during the Lateglacial time. Only a few cold tolerant littoral chydorid taxa such as A. harpae, A. affinis and C. sphaericus represented this period (Harmsworth, 1968; Kattel et al., 2008). However, a decrease in grass pollen concentration from the mid-to-late part of the Meiendorf (ca. 14000 cal yr bp) indicates gradual onset of warming (Litt et al., 2001a). From mid-to-late part of the Meiendorf, pollen spectra indicated a gradual increase in abundance of juniper (Juniperus L.), pine (Pinus L.) and birch (Betula L.) forests (Fig. 3). The Meiendorf landscape appears to have been dominated by open-steppe vegetation with a few trees in wind-shielded river valleys of south-exposed hill slopes. This result is also corroborated by cladoceran remains recorded in Core SM3. Species richness of cladoceran remains increased with limnetic cladocerans such as Daphnia, and the littoral cladocerans such as A. harpae, A. affinis, C. sphaericus and P. trigonellus (Fig. 5). Presence of Daphnia and the increased species richness of littoral chydorids suggests that the condition of Lake SMM during the Meiendorf may have been oligotrophic. However, the occurrence of a few cladoceran species such as A. guttata which prefers relatively mild temperate climates in modern time also indicates the onset of gradual warming of the lake from mid-to-late part of the Meiendorf period (e.g. Hofmann, 1993). Primary production in Eifel maar lakes including the Lake SMM is also reported to have increased during the early part of the postglacial period (Möller & Scharf, 1986).
Pollen spectra of both dwarf and large trees such as birch (Betula L.) increased during the early part of the Bölling (Fig. 3). Litt et al. (2001a, b) reported that the development of open birch (Betula L.) woodlands across the Eifel region during the Bölling period with an increase in non-arboreal pollens. Cladoceran remains were well preserved in this time with dominant abundance of planktonic Daphnia (Fig. 5). Species richness of littoral cladocerans also increased with C. sphaericus dominating the assemblage. Although the result is derived from the only one sample, the occurrence of A. guttata during the Bölling period indicates relatively a mild climatic condition in the region particularly around the Lake SMM (e.g. Harmsworth, 1968). LST is an important time marker of the Eifel maar lakes (Hajdas et al., 1995). For example, LST in Lake Hozmaar (a nearby lake of Lake SMM) has been recorded as 12900 cal yr bp (Zolitschka et al., 2000). Preservation of cladoceran remains was very poor because of tephra deposition in sediment during the Alleröd. The tephra layer in Lake SMM sediment was as high as 10 cm (Zolitschka et al., 2000). Evidence on the effects of geochemistry on diagenesis of cladoceran remains is not available for Lake SMM, however, a high sedimentation rate as well as the volcanically associated geo-chemistry of lake sediment together may have reduced the preservation of the cladoceran remains during the Alleröd.
The Younger Dryas in the Eifel maar lakes region was cold, and was dominated by sub-arctic steppe tundra with heliophytes (Artemisia L., Helianthemum Miller), shrubs including Juniper L., dwarf and sporadic tree birch (Litt et al., 2001a, b). Despite the cold climates during the Younger Dryas, abundance and species richness of cladocerans in Lake SMM was increasing (Fig. 5). The increased abundance of Daphnia in Lake SMM during the later part of the Younger Dryas indicates increasing lake water clarity (Fig. 5). Daphnia remains were also recorded from the crater lake sediment distributed across the continental Gutaiulai mountain region (Romania) during the Younger Dryas, together with cold and dry adapting plants such as pine (Pinus L.), birch (Betula L.) and alder (Alnus Mill.), respectively (Feurdean & Bennike, 2004). The occurrence of cold tolerant littoral chydorid cladocerans such as A. harpae, A. affinis and C. sphaericus in Lake SMM suggests a cold climatic condition across the Eifel maar lakes region during the Younger Dryas.
The transition from the Younger Dryas to the Boreal period was revealed by a significant increase in warm-climate preferring plants such as hazel (Corylus L.) (Feurdean & Bennike, 2004; Fig. 3). During the Preboreal time, the cladoceran assemblage was dominated by Daphnia with only a small proportion of Bosmina (Fig. 5). The species richness of warm tolerant littoral cladocerans increased where A. guttata became abundant in at least one of the three samples being collected during the Preboreal and Boreal periods. Diatom-inferred eutrophication was reported in the nearby Lake Holzmaar during the Boreal period (Brüchmann & Negendank, 2004), however, this result is not coeval with the changes in Lake SMM as Lake SMM was oligotrophic during the Preboreal and Boreal periods (Zolitschka et al., 2000). Sustained Daphnia populations in Lake SMM during the Preboreal and Boreal periods were possibly the result of high primary production supported by continuous groundwater discharge across the catchment. Palaeoproductivity of Lake SMM with respect to diatomaceous matter was controlled by a groundwater discharge from the lake’s catchment at ca. 10200 bp (Schettler et al., 1999).
Lake SMM experienced a short-lived cold stress in the 8.2 ka Event during the early Atlantic period. Based on the microfacies and geochemical analyses of seasonally laminated varved sediment from the Lake Holzmaar in Eifel, the onset and withdrawal of the 8.2 ka summer cooling occurred within a year, and that summer rains were reduced or absent (Prasad et al., 2009). The onset of cooler summers preceded the onset of winter dryness by ca. 28 years (Prasad et al., 2009). Although there was a small sample size, a spike in cladoceran ephippia during this time indicates changes in the lake environment (Fig. 5). In short summers, cladoceran adopt a reproductive strategy for producing ephippia, usually in July that continues through ice-free seasons (Frey, 1982).
The primary production also decreased significantly following the Atlantic period (Möller & Scharf, 1986). Reduced thermocline depth and deep water carbonate precipitation could have caused the reduction of primary production and cladoceran abundance. Carbonate precipitation during the 8.2 ka Event has been recorded in all Eifel maar lakes and appears to be directly related to cold temperature and better air circulation followed by reduced CO2 concentration in the water column (Sirocko, 2009). However, ambiguity exists in defining the nature of the 8.2 ka Event. Firstly, the longer summer cooling or shorter winter cooling or dryness needs to be established (Prasad et al., 2009). Changes in solar variability, North Atlantic Thermohaline circulation (THC), and the North Atlantic Oscillation (NAO) together could have made implications for the 8.2 ka Event as the NAO cycles can be more frequent during the drier periods possibly as a result of changes in the regional temperatures (Prasad et al., 2009).
The mid-Holocene optimum (ca. 6200 bc) in central Europe including the Eifel maar lakes region was indicated by elevated snowlines in the Alps (Nicolussi & Patzelt, 2006) and increased distribution of mixed oak (Quercus L.) hardwood forests with abundant thermophilous taxa (Kalis et al., 2003; Litt et al., 2009). During this period, the assemblage of Daphnia in Lake SMM declined with reciprocal increase in assemblage of B. coregoni and B. longirostris. B. coregoni and B. longirostris are indicators of shifting mesotrophic to eutrophic conditions. The gradual eutrophication occurred when the Neolithic settlers of the Linienbandkeramik started agriculture in southern Germany although there is no archaeological evidence for the settlement at Linienbandkeramik (5500–5100 bc) (Gronenborn & Sirocko, 2009; Sirocko, 2009). The changes in cladoceran assemblage at this time also indicate a gradual eutrophication of Lake SMM. This might have been caused either by well-developed thermocline during warm summer seasons or by increased total organic carbon production from the climate-induced leaf litter decomposition (e.g. Fuhrmann et al., 2004).
The Subboreal period in central Europe was humid with relatively cool climates (Kalis et al., 2003). A pronounced increase in abundance of alder (Alnus Mill.) and gradual decline in elm (Ulmus L.) has been reported during the Subboreal period in many parts of Europe (e.g. Edwards & McDonald, 1991; Parker et al., 2002; Kalis et al., 2003; Rasmussen, 2005; Sirocko, 2009). Abundance of elm (Ulmus L.) was relatively low when the alder (Alnus Mill.) abundance was increasing in Lake SMM (Fig. 3). Daphnia populations increased in the Subboreal period parallel with an increase in the abundance of C. sphaericus (Fig. 5). Based on pollen records, Gronenborn & Sirocko (2009) documented the use of foliage for cattle by the early Neolithic settlers in the nearby Ulmener Maar during the Subboreal period (ca. 4300 bc). However, the decline of elm (Ulmus L.) populations may not have been associated with cattle feeding during this time as the reciprocal response in abundance of alder and elm in Europe is still arguable. A decline in elm populations has been reported as a result of outbreak of the disease during this time (e.g. Parker et al., 2002). Following the mid-Holocene period, a gradual development of the Michelsberger culture is reported to have flourished around Urmitz, the Rhine valley (Gronenborn & Sirocko, 2009; Sirocko, 2009). Increased abundance of C. sphaericus and Daphnia during this period suggests that the development of the Michelsberger culture was not yet disturbing the water quality of Lake SMM. Urmitz is located as far as 50 km away from the Lake SMM and the influence of foliage harvests on raising cattle should be minimal (Gronenborn & Sirocko, 2009; Sirocko, 2009). An increased abundance of Daphnia in Lake SMM indicates a clear water condition. In the case of C. sphaericus, this taxon is reported to have colonised both littoral and open water habitats and responded well to a wide range of environmental gradients including both clean and polluted water bodies (Frey, 1980; Nilssen & Sandøy, 1990; Vijverberg & Boersma, 1997; Korhola, 1999; Kattel & Augustinus, 2010). The second pronounced marker in the lake sediment during the Subboreal period is the increase in beech (Fagus L.) forest after 1850 bc (3.8 m) (Fig. 3). The early Bronze Age farmers may have practiced slash and burn farming which favored the spread of beech (Fagus L.) forests across the central Europe between 3000 and 2000 bc (Gronenborn & Sirocko, 2009; Sirocko, 2009). Pollen and plant macrofossil records of the Dallund Sø, Denmark indicate extensive forest clearance in late Bronze Age (ca. 500 bc) followed by livestock grazing and meadow cultivation (Rasmussen, 2005). Cladoceran assemblages showed a clear response to changes in trophic states of Lake SMM during the Bronze Age (Fig. 5). A decline in Daphnia abundance during the late Bronze Age appears to be the result of reduced water quality caused by forest burning and increased organic rich carbon in the water column (Fig. 5).
The beginning of the Subatlantic period was cold and wet (Van Geel & Bergland, 2000). Increased river inflows to Eifel maar lakes were common during the Subatlantic period (Gronenborn & Sirocko, 2009; Sirocko, 2009). In order to cope with changing climatic conditions, farmers gradually modified farming techniques with inventions of tools including iron ploughs in ca. 800 bc (Gronenborn & Sirocko, 2009; Sirocko, 2009). In the southern and western Germany this period is widely regarded as the Hallstatt Iron Age culture (Gronenborn & Sirocko, 2009; Sirocko, 2009). The Hallstatt culture around the Eifel maar lakes region during ca. 800 bc was called the Hunsrück-Eifel culture (Gronenborn & Sirocko, 2009; Sirocko, 2009). During the Subatlantic period, the Daphnia populations declined sharply, whereas the abundance of C. sphaericus was relatively high. C. sphaericus shows a preference for a wide range of nutrient concentrations (Nilssen & Sandøy, 1990; Vijverberg & Boersma, 1997; Korhola, 1999). Relative abundance of small Alona (A. guttata) was also high during the Subatlantic period. Increased species diversity of small-sized pelagic cladocerans has been reported in Dallund Sø, Denmark between ca. 4830 and 750 bc (Johansson et al., 2005). L:P taxon ratios gradually increased during the early part of the Subatlantic period indicating shallow littoral margins with reduced lake level (Fig. 5). The people of the Hunsrück-Eifel culture may have utilized Lake SMM for various purposes including cultivation and water mills (Gronenborn & Sirocko, 2009; Sirocko, 2009). Occasional flash floods may also have increased sedimentation rates and altered lake depth in the Subatlantic period (Gronenborn & Sirocko, 2009; Sirocko, 2009). Hofmann (1990, 1993) reported that small A. guttata responded very well to widespread human-induced eutrophication across the Eifel maar lakes region. Changes in lake levels alter physical structures of sub-merged macrophytes in shallow water bodies and dynamics of littoral cladoceran assemblages (Whiteside et al., 1978). A relatively high abundance of chydorids such as A. harpae, A. guttata, C. sphaericus and Leydigia leydigi Leydig in the littoral margins of the Lake SMM during the Subatlantic period further strengthens our assumption of a reduced lake level during this time. As previously discussed, the people of Hunsrück-Eifel culture during this time may have used locally available natural resources for domestic consumption (Gronenborn & Sirocko, 2009; Sirocko, 2009). A brief appearance of the sand-preferring taxon, Monospilus dispar Sars at ca. 50 bc in Lake SMM suggests an increasing rate of deforestation followed by catchment erosion and sedimentation. The late Holocene sediment of the Lake SMM was dominated by lithic matter such as Turbidites and dust layers (Heinz et al., 1993) which may also have caused to extend the littoral layers of the lake. The high L:P taxon ratios accompanied by high sample scores of the PCA axis 2 of cladocerans during the early Iron Age are most likely to be the indication of the changes in catchment hydrology due to increased deforestation and sedimentation across the Eifel maar lakes region (Rein & Negendank, 1993; Rein et al., 2007). However, in the late Iron Age, differences in farming practice may have changed the patterns of cladoceran assemblages as indicated by increased sample scores of the PCA axis 2 (Fig. 5). Around this time, an increased abundance of macrophyte-associated chydorid cladoceran and algal taxa, which are indicative of lower lake levels have also been reported in Dallund Sø, Denmark (Bradshaw et al., 2005; Johansson et al., 2005). Hunsrück-Eifel culture (ca. 300 bc), and the Celtic community during this time left their fields with low level of manuring and longer fallow period followed by a more intensive farming (mostly rye, Secale cereal L.) with shorter fallow periods and a higher manuring intensity (Spek et al., 2003; Gronenborn & Sirocko, 2009; Sirocko, 2009).
Following the Roman’s victory against the Celtic people in ca. 58–52 bc, the nature of land use practice changed around the Lake SMM and also the activity of land use change expanded rapidly across the region (Van Haaren, 1988; Rasmussen, 2005). The Romans built roads and cultivated a variety of cereals by clearing off the dense forests as well as founded the “Villa Rustica” in the northwest area of the Lake SMM in ca. ad 150 (Gronenborn & Sirocko, 2009; Sirocko, 2009). The first cereal pollen in Lake SMM occured in ca. ad 150 (Fig. 3). Changes in palaeocladoceran assemblage with marked increase in abundance of B. coregoni (Fig. 5) indicates the alteration of trophic conditions as a result of aquaculture around the maar swamp and farming activity such as grazing cattle and horse. Although, the fish as a staple diet of the Romans is debatable (Kuijper & Turner, 1992), the loss of Daphnia populations and gradual mesotrophication of Lake SMM as indicated by increased abundance of B. coregoni during this time is expected as a result of a widespread cultural activity in the lake. In shallow Danish lakes, a shift in fish stock, for example, percid (mesotrophic) to cyprinid (eutrophic) has caused to switch macrophyte density and cladoceran assemblages followed by a lake water quality from clear to turbid state (e.g. Johansson et al., 2005). Changes in sediment colour and texture also occurred with dominant clastic soil inferring flash flood layers deposited during the late Roman and the beginning of the Migration period (ca. ad 260) (Heinz et al., 1993). For the first time in Roman period, the local German tribes invaded the Romano-Celtic populations in Eifel areas resulting in significant social and political conflicts (Gronenborn & Sirocko, 2009; Sirocko, 2009) implicating the disturbances further in the use of natural resources.
The widespread forest clearances occurred during the Migration period in the Eifel maar lakes region (Zolitschka et al., 2003). At least three pronounced flash flood layers occurred during this period. The River Rhine also experienced a prolonged freezing (Sirocko, 2009). The abrupt climate change and widespread occurrence of human migration during this period may have considerable implications for water quality and ecology of the Lake SMM. Meantime, local German tribes who defeated the Romans were able to settle further south to the Mediterranean. As a result of increased conflict between Germans and Romans, the exploitation of natural resources across the region was further intensified (Schreg & Sirocko, 2009; Sirocko, 2009). Even though the cladoceran remains were derived from only one sample, at around the end of the Roman Empire (ca. ad 471), Daphnia and B. coregoni populations showed a reciprocal response. We argue that the cladoceran response during this time was associated with increased social and cultural changes in the Eifel maar lakes region (also see Johansson et al., 2005). Cladoceran records derived from additional samples can help comprehensive understanding in the effects of conflicting resource use on reciprocal switching of Daphnids and Bosminids in Lake SMM during the Migration period. Although the abundance of chydorid cladocerans declined during the Migration period, an increased density of resting eggs of both limnetic and littoral cladocerans indicates the stress on lake ecosystems (Sarmaja-Korjonen, 2004). Cold climates accompanied by land use activity by local tribes following the demolition of the Roman Villa Rustica and predation pressure by fish could have caused the ecological stress of the Lake SMM during the mid Migration period (e.g. Johansson et al., 2005). Apparently, the area around the Lake SMM may have been abandoned for agriculture during the late Migration and the lake was restored naturally as a clear water state as shown by increased remains of large sized cladocerans such as Daphnia accompanied by increased PCA axis 1 sample scores (Fig. 5). However, increased sample size will be useful to support the assertion of the increasing link between water clarity and ecological changes of the lake during this period.
In the early Medieval period, B. coregoni and B. longispina dominated the cladoceran community. This trend continued till ca. ad 500, when the Frankish people ruled the Eifel region. Pollen records in the sixth century ad suggest that the vegetation was dominated by beech (Fagus L.) and hornbeam (Carpinus L.) trees. Cereal pollens, however, were observed almost continuously during the entire early Medieval period around the Eifel region (Schreg & Sirocko, 2009; Sirocko, 2009) and also further north in the Danish lakes region, Dallund Sø (Bradshaw et al., 2005; Rasmussen, 2005). During the early Medieval time, concentration of grass pollens also increased (Fig. 3) indicating the significance of sheep farming and economic prosperity of the region. Evidence of increased abundance of limnetic cladocerans in Lake SMM suggests that the lake could have been less productive, possibly because the early Medieval period was still less favourable to cold preferring cladoceran taxa. A minimum solar strength was experienced in central Germany during the early Medieval period (Schreg & Sirocko, 2009; Sirocko, 2009). However, the Medieval Europe (ca. ad 500–1500) was regarded as warm with dry summers and mild winters due to a prevailing anticyclonic circulation and persistent westerly air flow (Bradley et al., 2003). The Medieval period witnessed occasionally the extreme weather patterns such as phases with very cold winters and high storm intensities (Glaser, 2008). Anthropogenic stressors were also critical during this time with cumulative impacts on biodiversity and ecosystem of lakes (e.g. Folke et al., 2004; Carpenter & Brock, 2006). Distinguishing natural climate change from anthropogenic effects during the Medieval period is difficult due to their overlapping nature of influence on lake ecosystems. Several factors can bias whilst distinguishing these effects. The taphonomic and edaphic effects obscure representation of cladocerans in lake sediments and minimize accuracy of palaeolimnological reconstructions (e.g. Kattel et al., 2007). Thin section analysis of sedimentary minerals, diatoms, cladocerans and varve counts of Eifel maar lakes have been able to overcome these problems and provided an increasingly accurate evidence of the effects of anthropogenic and climatic change (e.g. windstorms) on lake ecosystems separately (Rein & Negendank, 1993; Rein et al., 2007; Pfahl et al., 2009). The cladoceran records in the Medieval sediment showed relatively poor species richness with only one taxon, B. coregoni dominated the lake (Fig. 5). Increased B. coregoni abundance in Lake SMM during the high Medieval period indicates a gradual change of lake from oligotrophication to mesotrophication due to warm climate and farming activity across the maar catchment (Schreg & Sirocko, 2009; Sirocko, 2009). Increased abundance of small sized Bosmina indicates fishing activity in lakes. Sustained removal of large size fish can lead to greater density of planktivorous fish thereby allowing the smaller cladoceran (e.g. Bosmina) community to flourish (e.g. Leucke et al., 1990). The loss of Daphnia populations in Lake SMM following the Medieval period is unknown although the combination of human impacts on lake catchments and rapidly changing climates, for example, approaching of the Little Ice Age (ad 1500–1700) are anticipated. After the ad 1000, a reciprocal response of Daphnia and B. coregoni assemblages have also indicated major cascading effects on food web and the water quality of the Dallund Sø, Denmark (Johansson et al., 2005). A widespread anthropogenically induced soil erosion and lithic maximum in sediment have also influenced the Lake SMM catchment during the Little Ice Age (Möller & Scharf, 1986; Heinz et al., 1993; Zolitschka, 2002).
Conclusions
Palaeocladoceran records archived in Lake SMM sediment reveal that cladoceran zooplankton have been increasingly sensitive organism to rapid environmental, cultural and archaeological changes across the Eifel maar lakes region (West Germany). Lateglacial periods represented by Bölling, Alleröd, Younger Dryas, Preboreal and Boreal, were inferred by clear water-preferring cladocerans and natural vegetation development across the lake catchments. The lake was oligotrophic in nature during the preboreal and boreal periods with dominant hazel (Corylus L.) forests and grasslands in the catchment. However, the lake went onto eutrophication during the early Atlantic period. Following the 8.2 ka Event, the mixed oak–alder (Quercus L.–Alnus Mill.) forests dominated the lake catchment providing a nutrient rich leaf-litter detritus suitable for littoral cladocerans. An increased abundance of B. longirostris and reduced elm (Ulmus L.) forests at ca. 4300 bc reflects a sign of the first human impacts on lake ecosystems as a result of forest clearance and use of foliage for livestock production. Lake ecosystems returned to an oligotrophic condition during the Subboreal period commensurate with increased abundance of Daphnia. However, the Iron Age–Hunsrück culture across the Eifel maar lakes region during the Subatlantic period led to a mesotrophic condition followed by a reduction of Daphnia populations in the lake. The lake experienced shallow littoral habitats during the early Subatlantic period as indicated by increased L:P taxon ratios. Fishing and waste disposal activities increased in the lake following the foundation of the Roman Villa Rustica in the northern lake catchment in ca. ad 150. The common flash flood layers from this time onward also increased the frequency of catchment soil erosion leading to mesotrophication. However, the lake may have returned to an oligotrophic condition but short-lived during the early Migration period (ca. ad 460) when the Roman Villa Rustica was destroyed and the people abandoned the area. Over the Frankish rule during the fifth century ad, the populations of B. coregoni increased and a reciprocal decrease in Daphnia occurred. Changes in cladoceran population structure and dynamics of the maar from ca. ad 600 through to the Medieval period and to present time appears to be the result of an indirect effect of recent anthropogenic warming and increased land use activity in the lake catchment. Loss of cladoceran species richness from this time has become a major setback for restoring the water quality and healthy ecosystems of the lake. Lack of precise archaeological evidence of human activity around the Lake SMM has further impeded the investigation of the biological values of the Eifel maar lakes to infer environmental, cultural and archaeological developments in the region.
References
Berglund, B. E. & M. Ralska-jasiewiczowa, 1986. Pollen analysis and pollen diagrams. In Berglund, B. E. (ed.), Handbook of Holocene Palaeoecology and Palaeohydrology. Wiley, Chichester: 455–484.
Birks, H. J. B., 1995. Quantitative paleoenvironmental reconstructions. In Maddy, D. & J. S. Brew (eds), Statistical Modelling of Quaternary Science Data. Cambridge University Press, Cambridge: 254 pp.
Bonsall, C., M. G. Macklin, D. E. Anderson & R. W. Payton, 2002. Climate Change and the Adoption of Agriculture in North-West Europe. European Journal of Archaeology 5: 9–23.
Bradley, R. S., M. K. Hughes & H. F. Diaz, 2003. Climate in Medieval time. Science 302: 404–405.
Bradshaw, E. G., P. Rasmussen, H. Nielsen & N. J. Anderson, 2005. Mid-to-late-Holocene land-use change and lake development at Dallund Sø, Denmark: trends in lake primary production as reflected by algal and macrophyte remains. The Holocene 15: 1130–1142.
Brauer, A., C. Endres, C. Gunter, T. Litt, M. Stebich & J. F. W. Negendank, 1999a. High resolution sediment and vegetation responses to Younger Dryas climate change in varved lake sediments from Meerfelder Maar, Germany. Quaternary Science Reviews 18: 321–329.
Brauer, A., C. Endres & J. F. W. Negendank, 1999b. Lateglacial calendar year chronology based on annually laminated sediments from Lake Meerfelder maar, Germany. Quaternary International 61: 17–25.
Brüchmann, C. & J. F. W. Negendank, 2004. Indication of climatically induced natural eutrophication during the early Holocene period, based on annually laminated sediment from Lake Holzmaar, Germany. Quaternary International 123–125: 117–134.
Büchel, G., 1993. Maars of the Westeifel (Germany). In Nagendank, J. F. W. & B. Zolitschka (eds), Paleolimnology of European Maar Lakes. Lecture Notes in Earth Sciences, Vol. 49. Springer, Berlin: 1–14.
Carpenter, S. R. & W. A. Brock, 2006. Rising variance: a leading indicator of ecological transition. Ecology Letters 9: 311–318.
Clarke, K. R. & R. N. Gorley, 2006. PRIMER v6: User Manual/Tutorial. PRIMER-E, Plymouth, UK: 192 pp.
Clarke, K. R. & R. M. Warwick, 2001. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation. PRIMER-E, Plymouth, UK: 38 pp.
Creer, K. M. & N. Thouveny, 1996. The EUROMAARS project: a multidisciplinary approach directed to the reconstruction of palaeoenvironmental, palaeoclimatic and palaeogeomagnetic changes recorded by sediments deposited in maar lakes located in western Europe. Quaternary Science Reviews 15: 99–245.
Crisman, T. L. & D. R. Whitehead, 1978. Paleolimnological studies on small New England (USA) ponds. Part II. Cladocera community responses to trophic oscillations. Polish Archives of Hydrobiology 25: 75–86.
Cunliffe, B., 1997. The Ancient Celts. Oxford University Press, Oxford.
Edwards, K. J. & G. M. McDonald, 1991. Holocene palynology II. Human influence and vegetation change. Progress in Physical Geography 15: 364–391.
Erlandson, J. M., 2001. The archaeology of aquatic adaptations: paradigms for a new millennium. Journal of Archaeological Research 9: 287–350.
Faegri, K. & J. Iversen, 1989. Textbook of Pollen Analysis. John Wiley & Sons, New York.
Feurdean, A. & O. Bennike, 2004. Late Quaternary palaeoecological and palaeoclimatological reconstruction in the Gutaiului Mountains, northwest Romania. Journal of Quaternary Science 19: 809–827.
Folke, C., S. Carpenter, B. Walker, M. Scheffer, T. Elmqvist, L. Gunderson & C. S. Holling, 2004. Regime shifts, resilience and biodiversity in ecosystem management. Annual Review of Ecology, Evolution and Systematics 35: 557–581.
Frey, D. G., 1958. The late-glacial cladoceran fauna of a small lake? Archiv für Hydrobiologie 54: 209–275.
Frey, D. G., 1959. The taxonomic and phylogenetic significance of the headpores of the Chydoridae (Cladocera). Internationale Revue der Gesamten Hydrobiologie 44: 27–50.
Frey, D. G., 1980. On the plurality of Chydorus sphaericus (O. F. Müller) (Cladocera, Chydoridae), and designation of neotype from Sjaelsø, Denmark. Hydrobiologia 69: 83–123.
Frey, D. G., 1982. Contrasting strategies of gamogenesis in northern and southern populations of Cladocera. Ecology 63: 223–241.
Frey, D. G., 1986. Cladocera analysis. In Berglund, B. E. (ed.), Handbook of Holocene Palaeoecology and Palaeohydrology. John Wiley & Sons, Chichester: 667–692.
Fuhrmann, A., T. Fischer, A. Lücke, A. Brauer, B. Zolitschka, B. Horsfield, J. F. W. Negendank, G. H. Schleser & H. Wilkes, 2004. Late Quaternary environmental and climatic changes in central Europe as inferred from the composition of organic matter in annually laminated maar lake sediments. Geochemistry Geophysics Geosystems 5: Q11015.
Glaser, R., 2008. Klimageschichte Mitteleuropas. 1200 Jahre Wetter, Klima, Katastrophen. Mit Prognosen für das 21. Jahrhundert. Wissenschaftliche Buchgesellschaft, Darmstadt.
Goulden, C. E., 1969. Interpretative studies of cladoceran microfossils in lake sediments. Mitteilungen Internationale Vereinigung für Theoretische und Angewandte Limnologie 17: 43–55.
Gronenborn, D. & F. Sirocko, 2009. 4400–3400 BC Viehwirtschaft und Ausbreitung der Michelsberger Kultur. In Sirocko, F. (ed.), Wetter – Klima – Menschheitsentwicklung. Von der Eiszeit bis ins 21. Jahrhundert. Theiss Verlag, Stuttgart: 116–118.
Hajdas, I., B. Zolitschka, S. D. Ivy-Ochs, J. Beer, G. Bonani, S. A. G. Leroy, J. W. Negendank, M. Ramrath & M. Suter, 1995. AMS radiocarbon dating of annually laminated sediments from Lake Holzmaar, Germany. Quaternary Science Reviews 14: 137–143.
Harmsworth, R. V., 1968. The developmental history of Belham Tarn (England) as shown by Animal Microfossils, with special reference to the Cladocera. Ecological Monographs 38: 223–241.
Heinz, T., B. Rein & J. F. W. Negendank, 1993. Sediments and basin analysis of Lake Schalkenmehrener Maar. In Negendank, J. F. W. & B. Zolitschka (eds), Paleolimnology of European Maar Lakes. Lecture Notes in Earth Sciences, Vol. 49. Springer, Berlin: 149–161.
Hofmann, W., 1987. Cladocera in space and time: analysis of lake sediments. Hydrobiologia 145: 315–321.
Hofmann, W., 1990. Weichselian chironomid and cladoceran assemblages from maar lakes. Hydrobiologia 214: 207–211.
Hofmann, W., 1993. Lateglacial/Holocene changes of the climatic and trophic conditions in three Eifel maar lakes as indicated by faunal remains. I. Cladocera. Lecture Notes in Earth Sciences 49: 393–420.
Hofmann, W., 1996. Empirical relationships between cladoceran fauna and trophic state in thirteen northern German lakes: analysis of surficial sediments. Hydrobiologia 318: 195–201.
Jeppesen, E., J. P. Jensen, M. Sondergaard, T. Lauridsen & F. Landkildehus, 2000. Trophic structure, species richness and biodiversity in Danish lakes: changes along a phosphorus gradient. Freshwater Biology 45: 201–218.
Jeppesen, E., J. P. Jensen, S. Amsinck, F. Landkildehus, T. Lauridsen & S. F. Mitchell, 2002. Reconstructing the historical changes in Daphnia mean size and planktivorous fish abundance in lakes from the size of Daphnia ephippia in the sediment. Journal of Paleolimnology 27: 133–143.
Johansson, L. S., S. L. Amsinck, R. Bjerring & E. Jeppesen, 2005. Mid-to-late-Holocene land-use change and lake development at Dallund Sø, Denmark: trophic structure inferred from cladoceran subfossils. The Holocene 15: 1143–1151.
Kalis, A. J., J. Merkt & J. Wunderlich, 2003. Environmental changes during the Holocene Climate Optimum in central Europe – human impact and natural causes. Quaternary Science Reviews 22: 33–79.
Kappes, H., C. Mechenich & U. Sinsch, 2000. Long-term dynamics of Asplanchna priodonta in Lake Windsborn with comments on the diet. Hydrobiologia 432: 91–100.
Kattel, G. R. & P. Augustinus, 2010. Biogeography and taxonomy of New Zealand Chydorid Cladocera (Anomopoda, Chydoridae). Journal of the Royal Society of New Zealand 40: 209–224.
Kattel, G. R., R. W. Battarbee, A. W. Mackay & H. J. B. Birks, 2007. Are Cladoceran fossils in core samples a biased reflection of the Cladoceran communities from which they were derived? Journal of Paleolimnology 38: 157–181.
Kattel, G. R., R. W. Battarbee, A. Mackay & H. J. B. Birks, 2008. Recent ecological change in remote mountain loch: an evaluation of cladocera-based temperature transfer function. Palaeogeography, Palaeoecology, Palaeooceanography 259: 51–76.
Korhola, A., 1999. Distribution patterns of Cladocera in subarctic Fennoscandian lakes and their potential in environmental reconstruction. Ecography 22: 357–373.
Korhola, A. A. & M. Rautio, 2001. Cladocera and other branchiopod crustaceans. In Smol, J. P., H. J. B. Birks & W. M. Last (eds), Tracking Environmental Change Using Lake Sediments, Volume 4: Zoological Indicators. Kluwer Academic Publishers, Dordrecht, The Netherlands: 240 pp.
Kubitz, B., 2000. Die Holozäne Vegetations- und Sieldlungsgeschichte in der Westeifel am Beispiel eines Hochauflösenden Pollendiagrammes aus dem Meerfelder Maar. Dissertationes Botanicae 339: 106 pp.
Kühl, N. & T. Litt, 2007. Quantitative time-series reconstructions of Holsteinian and Eemian temperatures using botanical data. In Sirocko, F., M. Claussen, M. F. S. Goni & T. Litt (eds), The Climate of Past Interglacials, Developments in Quaternary Science, Vol. 7. Elsevier, Amsterdam: 239–254.
Kuijper, W. J. & H. Turner, 1992. Diet of a Roman centurion at Alphen aan den Rijn, The Netherlands, in the first century AD. Review of Palaeobotany and Palynology 73: 187–204.
Leucke, C., M. J. Vanni, J. J. Magnuson, J. F. Kitchel & T. J. Jacobson, 1990. Seasonal regulation of Daphnia populations by planktivorous fish: implications for the spring clear-water phase. Limnology and Oceanography 35: 1718–1733.
Litt, T., 2004. Eifelmaare als Archive für die Vegetations- und Klimageschichte der letzten 15000 Jahre. Berichte der Reinhold-Tüxen-Gesellschaft 16: 87–95.
Litt, T., A. Brauer, T. Goslar, J. Merkt, K. Bałaga, H. Müller, M. Ralska-Jasiewiczowa, M. Stebich & J. F. W. Negndank, 2001a. Correlation and synchronization of Lateglacial continental sequences in northern central Europe based on annually laminated lacustrine sediments. Quaternary Science Reviews 20: 1233–1249.
Litt, T., A. Brauer, H.-U. Schmincke & B. Kromer, 2001b. Environmental response to climatic and volcanic events in central Europe during the Weichselian Lateglacial. Quaternary Science Reviews 22: 7–32.
Litt, T., C. Schölzel, N. Kuhl & A. Brauer, 2009. Vegetation and climate history in the Westeifel Volcanic Field (Germany) during the past 11000 years based on annually laminated lacustrine maar sediments. Boreas 38: 679–690.
Lotter, A. F., H. J. B. Birks, U. Eicher, W. Hofmann & A. Marchetto, 1997. Modern diatom, Cladocera, chironomid, and chrysophyte cyst assemblages as quantitative indicators for the reconstruction of past environmental conditions in the Alps. I. Climate. Journal of Paleolimnology 18: 395–420.
Lücke, A., G. H. Schlesser, B. Zolitschka & J. F. W. Negendank, 2003. A Lateglacial and Holocene organic carbon isotope record of lacustrine palaeoproductivity and climatic change derived from varved lake sediments of Lake Holzmaar, Germany. Quaternary Science Reviews 22: 569–580.
Mikulski, J. S., 1978. Value of some biological indices in case historic of lakes. Verhandlungen der Internationalen Vereinigung Limnologie 20: 992–996.
Möller, W. A. A. & B. W. Scharf, 1986. The content of chlorophyll in the sediment of the volcanic maar lakes in the Eifel region (Germany) as an indicator for eutrophication. Hydrobiologia 143: 327–329.
Moses, J., 1976. Recent social and political history in West Germany. Australian Journal of Politics and History 22: 316–321.
Nicolussi, K. & G. Patzelt, 2006. Klimawandel und Veränderungen an der alpinen Waldgrenze – aktuelle Entwicklungen im Vergleich zur Nacheiszeit. BFW Praxisinformationen 10.
Nilssen, J. P. & S. Sandøy, 1990. Recent lake acidification and cladoceran dynamics: surface sediment and core analyses from lakes in Norway, Scotland and Sweden. Philosophical Transactions of the Royal Society of London B327: 299–309.
Parker, A. G., A. S. Goudie, D. E. Anderson, M. A. Robinson & C. Bonsall, 2002. A review of the mid-Holocene elm decline in the British Isles. Progress in Physical Geography 26: 1–45.
Pfahl, S., F. Sirocko, K. Seelos, S. Dietrich, A. Walter & H. Wernli, 2009. A new windstorm proxy from lake sediments: a comparison of geological and meteorological data from western Germany for the period 1965–2001. Journal of Geophysical Research 114: D18106.
Polcyn, I., 1996. Application of cladocera analysis in archaeology. Circaea, The Journal of the Association for Environmental Archaeology 11: 41–48.
Prasad, S., A. Witt, U. Kienel, P. Dulski, E. Bauer & G. Yancheva, 2009. The 8.2 ka event: evidence for seasonal differences and the rate of climate change in Western Europe. Global and Planetary Change 67: 218–226.
Rasmussen, P., 2005. Mid-to-late-Holocene land-use change and lake development at Dallund Sø, Denmark: vegetation and land-use history inferred from pollen data. The Holocene 15: 1116–1129.
Rasmussen, P. & E. G. Bradshaw, 2005. Mid-to-late-Holocene land-use change and lake development at Dallund Sø, Denmark: study, aims, natural and cultural setting, chronology and soil erosion history. The Holocene 15: 1105–1115.
Rein, B. & J. F. W. Negendank, 1993. Organic carbon contents of sediments from Lake Schalkenmehrener Maar: a palaeoclimate indicator. In Negendank, J. F. W. & B. Zolitschka (eds), Paleolimnology of European Maar Lakes. Lecture Notes in Earth Sciences, Vol. 49. Springer, Berlin: 326 pp.
Rein, B., K. Jäger, Y. Kocot, K. Grimm & F. Sirocko, 2007. Holocene and Eemian varve types of Eifel Maar lakes sediments. In Sirocko, F., M. Claussen, M. F. S. Goni & T. Litt (eds), The Climates of Past Interglacials, Developments in Quaternary Science, Vol. 7. Elsevier, Oxford: 615 pp.
Sarmaja-Korjonen, K., 2003. Chydorid ephippia as indicators of environmental change – biostratigraphical evidence from two lakes in southern Finland. The Holocene 13: 691–700.
Sarmaja-Korjonen, K., 2004. Chydorid ephippia as indicators of past environmental changes – a new method. Hydrobiologia 526: 129–136.
Schaber, K. & F. Sirocko, 2005. Lithologie und Stratigraphieder spätpleistozänen Trockenmaare der Eifel. Mainzer geowissenschaftliche Mitteilungen 33: 295–340.
Scharf, B. & S. Björk, 1992. Limnology of Eifel maar lakes. Archiv für Hydrobiologie/Ergebnisse der Limnologie 38: 348 pp.
Scharf, B. & M. Oehms, 1992. Physical and chemical characteristics of Eifel maar lakes. Advances in Limnology 38: 63–83.
Schettler, G. & R. Romer, 1998. Anthropogenic influences on Pb/Al and lead isotope signature in annually layered Holocene Maar lake sediments. Applied Geochemistry 13: 787–797.
Schettler, G., B. Rein & J. F. W. Negendank, 1999. Geochemical evidence for Holocene palaeodischarge variations in lacustrine records from the Westeifel Volcanic Field, Germany: Schalkenmehrener Maar and Meerfelder Maar. The Holocene 9: 381–400.
Schmidt, B. & W. Gruhle, 2003. Niederschlagsschwankungen in Westeuropa während der letzten 8000 Jahre. Archäologisches Korrespondenzblatt 33: 281–300.
Schreg, R. & F. Sirocko, 2009. 400–500 AD Völkerwanderung und Umweltkrise – das Ende des römischen Weltreiches. In Sirocko, F. (ed.), Wetter – Klima Menschheitsentwicklung. Von der Eiszeit bis ins 21. Jahrhundert. Theiss Verlag, Stuttgart: 150–153.
Seelos, K. & F. Sirocko, 2009. Limnologie der Maarseen und typische Sedimente. In Sirocko, F. (ed.), Wetter – Klima – Menschheitsentwicklung, Von der Eiszeit bis ins 21, Jahrhundert. Theiss Verlag, Stuttgart.
Sirocko, F. (ed.), 2009. Wetter – Klima – Menschheitsentwicklung. Von der Eiszeit bis ins 21. Jahrhundert. Theiss Verlag, Stuttgart: 208 pp.
Spek, T., W. G. Waateringe, M. Kooistra & L. Bakker, 2003. Formation and land-use history of celtic fields in north-west Europe – an interdisciplinary case study at Zeijen, The Netherlands. European Journal of Archaeology 6: 141–173.
Straka, H., 1975. Die spätquartäre Vegetationsgeschichte der Vulkaneifel. Beitr Landespfl Rheinl-Pfalz: 163 pp.
Szeroczyńska, K., 1998. Palaeolimnological investigations in Poland based on Cladocera. Palaeogeography, Palaeoclimatology, Palaeoecology 140: 335–345.
Szeroczyńska, K. & K. Sarmaja-Korjonen, 2007. Atlas of Subfossil Cladocera from Central and Northern Europe, Friends of the Lower Vistula Society, Świecie, Poland: 84 pp.
ter Braak, C. J. F. & P. Smilauer, 2002. CANOCO Reference Manual and CanoDraw for Window’s User’s Guide: Software for Canonical Community Ordination (Version 4.5). Microcomputer Power, Ithaca, NY, USA: 500 pp.
Van Geel, B. & B. E. Bergland, 2000. A causal link between the climatic deterioration around 850 cal BC and a subsequent rise in human population density in NW-Europe? Terra Nostra 5: 126–130.
Van Haaren, C., 1988. Eifelmaare, Landschaftsokologisch-historische Betrachtung und Naturschutz. Planung, Pollichia 16: 518 pp.
Vijverberg, J. & M. Boersma, 1997. Long-term dynamics of small-bodied and large-bodied cladocerans during the eutrophication of a shallow reservoir, with special attention for Chydorus sphaericus. Hydrobiologia 360: 233–242.
Wagner, A. & J. Bendorf, 2007. Climate-driven warming during spring destabilises a Daphnia population: a mechanistic foodweb approach. Oecologia 151: 351–364.
Whiteside, M. C., 1970. Danish Chydorid Cladocera: modern ecology and core studies. Ecology 40: 79–118.
Whiteside, M. C., J. B. Williams & C. P. White, 1978. Seasonal abundance and pattern of chydorid, Cladocera in mud and vegetative habitats. Ecology 59: 1177–1188.
Zolitschka, B., 2002. Late Quaternary sediment yield variations – natural vs human forcing. Zeitschrift fur Geomorphologie N.F., Suppl.-Bd. 128: 1–15.
Zolitschka, B. & J. F. W. Negendank, 1999. High-resolution from European Lakes. Quaternary Science Reviews 18: 885–888.
Zolitschka, B., A. Brauer, J. F. W. Negendank, H. Stockhausen & A. Lang, 2000. Annually dated late Weichselian continental paleoclimate record from the Eifel, Germany. Geology 28: 783–786.
Zolitschka, B., K.-E. Behre & J. Schneider, 2003. Human and climatic impact on the environment as derived from colluvial, fluvial and lacustrine archives – examples from the Bronze Age to the Migration period, Germany. Quaternary Science Reviews 22: 81–100.
Acknowledgements
This work was funded by a grant from the Research Center “Earth System Science” at the University of Mainz, Germany where GRK was a Visiting Scholar. We would like to thank the members of the ELSA team of the Geoscience Department of the University of Mainz, who have assisted for both field and laboratory work of this study. Particularly, we are very grateful to Saskia Rudert and Petra Sigl of the ELSA team for their untiring assistance for drawing figures. We are also thankful to four anonymous referees for their constructive comments plus some very useful remarks by guest editor of this special issue, Hilde Eggermont, all this have improved the quality of this article significantly.
Author information
Authors and Affiliations
Corresponding author
Additional information
Guest editors: H. Eggermont & K. Martens / Cladocera as indicators of environmental change
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Kattel, G., Sirocko, F. Palaeocladocerans as indicators of environmental, cultural and archaeological developments in Eifel maar lakes region (West Germany) during the Lateglacial and Holocene periods. Hydrobiologia 676, 203–221 (2011). https://doi.org/10.1007/s10750-011-0872-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-011-0872-4